

Algae and Cyanobacteria Diversity and Bioindication of Long-Term Changes in the Hula Nature Reserve, Israel



Abstract
:1. Introduction
- To compile the list of algae and cyanobacteria species from the past century references and our monitoring data and analyze it by bioindication and statistics;
- To compare the algae and cyanobacteria community of Lake Hula before drainage with it in the Hula Nature Reserve;
- To actualize the bioindication methods for monitoring the ecological situation in the aquatic ecosystem.
2. Materials and Methods
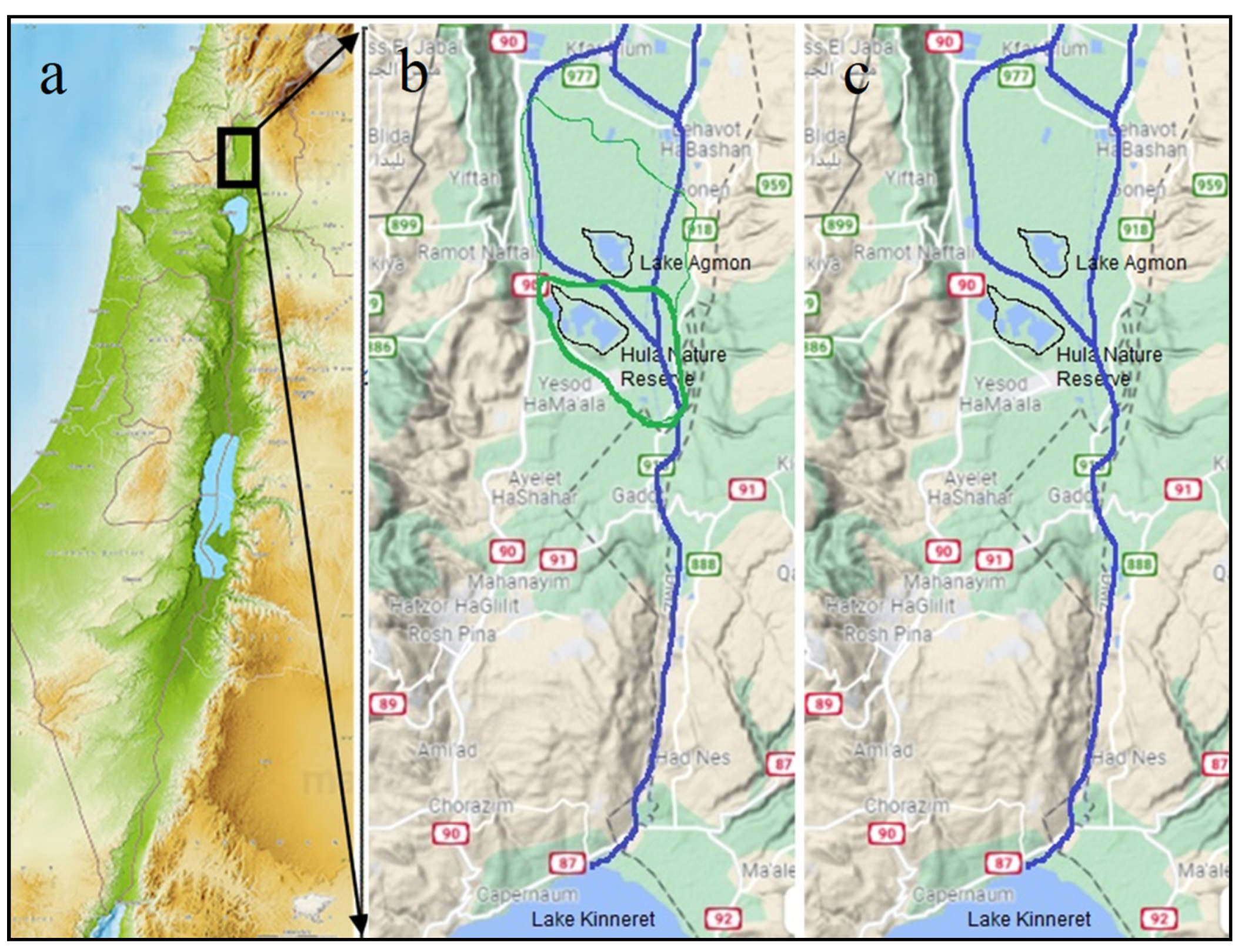
2.1. Description of Study Site
2.2. Sampling and Laboratory Studies
2.3. Determination of Water Physicochemical Properties
2.4. Historical Data Analysis
2.5. Bioindication Analysis
2.6. Statistical Analysis
2.7. Nygaard Species Indices for Trophic State Assessment
- Index 1 (NY-1) = Myxophyceae/Desmidiaceae
- Index 2 (NY-2) = Chlorococcales/Desmidiaceae
- Index 3 (NY-3) = Centrales/Pennales
- Index 4 (NY-4) = Euglenineae/(Myxophyceae + Chlorococcales)
- Index 5 (NY-5) = (Myxophyceae + Chlorococcales + Centrales + Euglenineae)/Desmidiaceae
3. Results
3.1. Water Chemistry
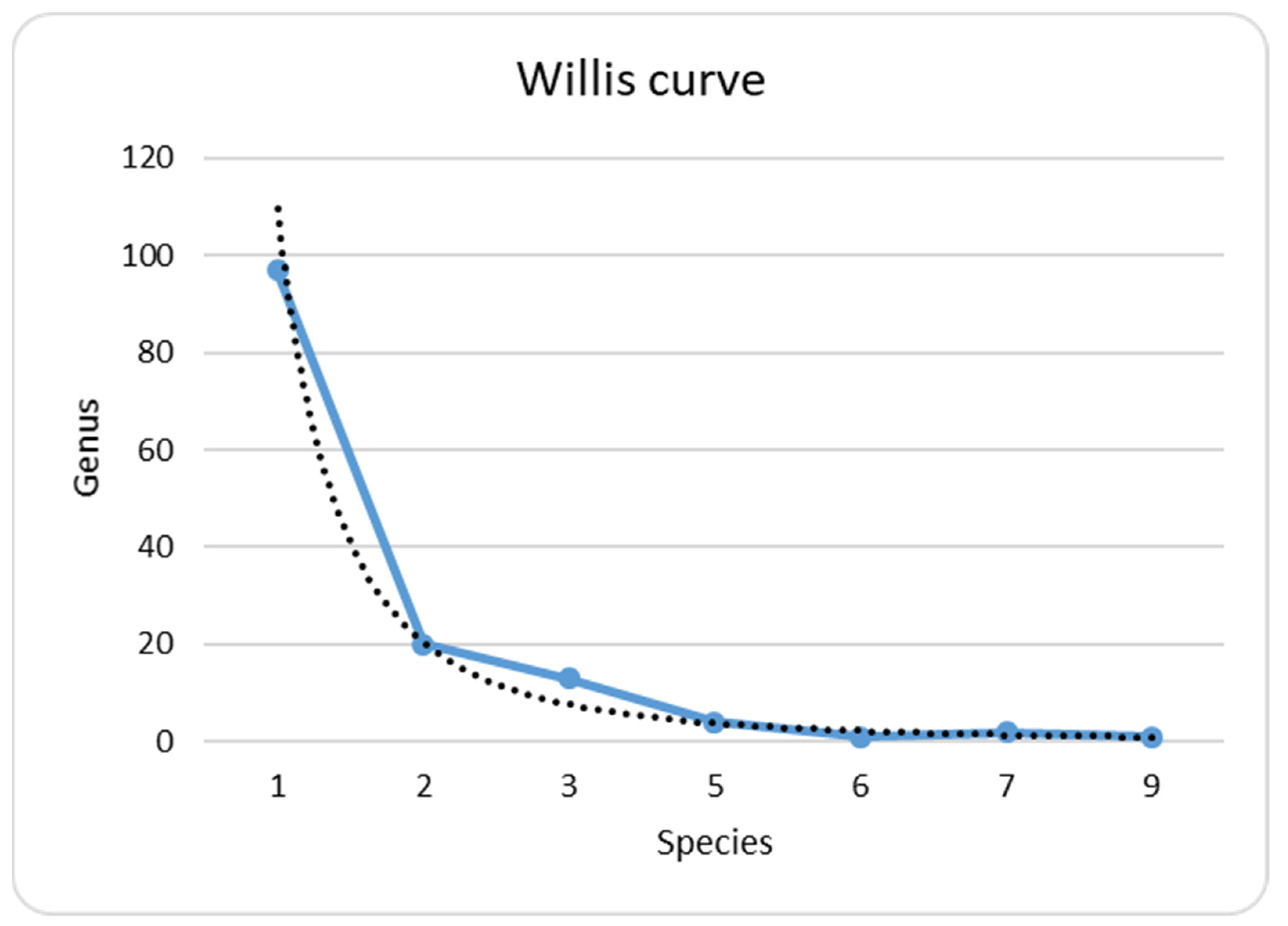
3.2. Species Richness
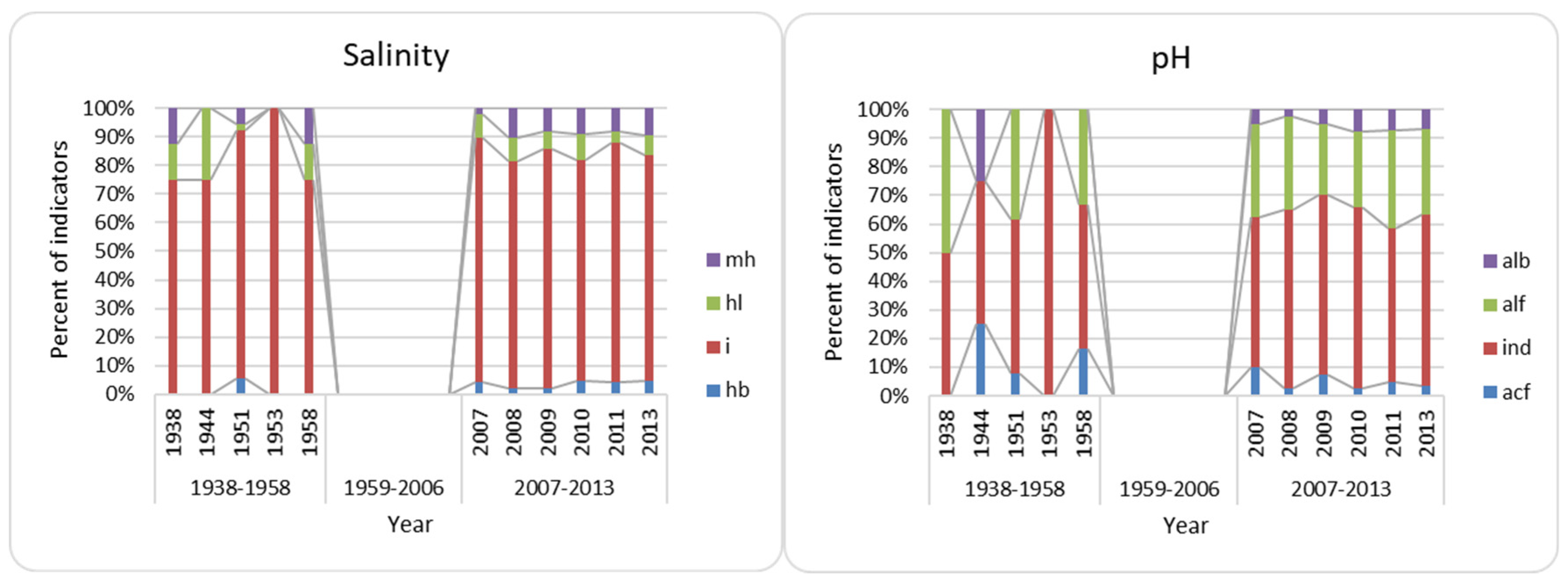
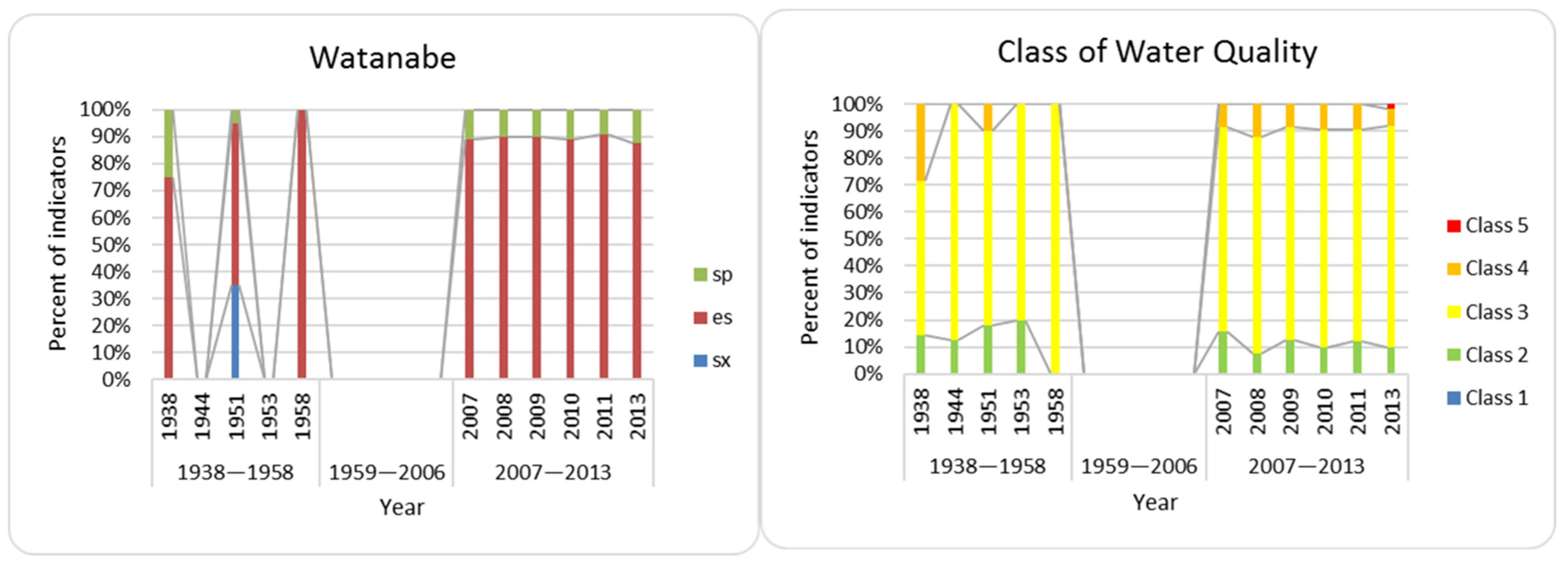
3.3. Bioindication
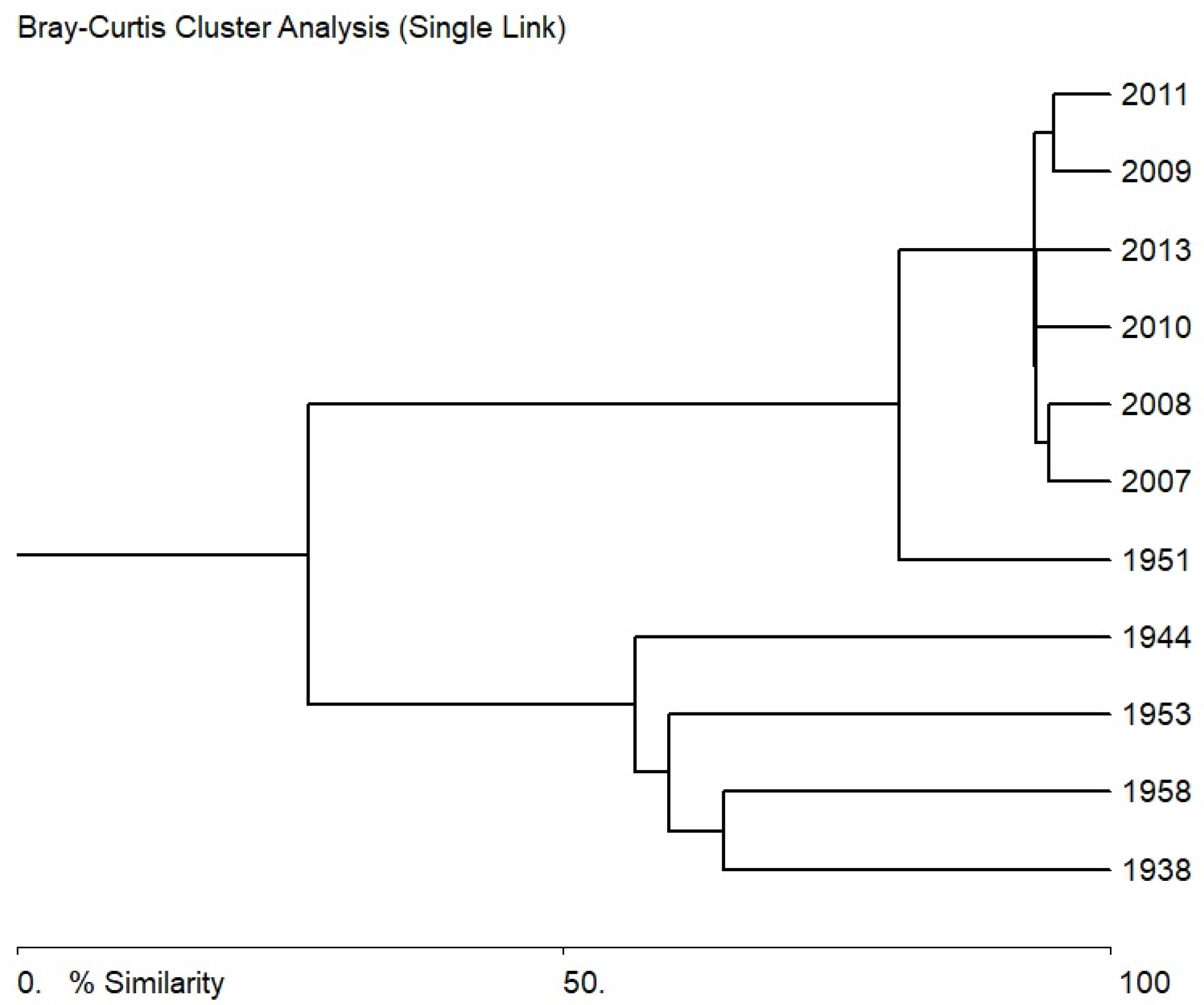
3.4. Statistical Analysis
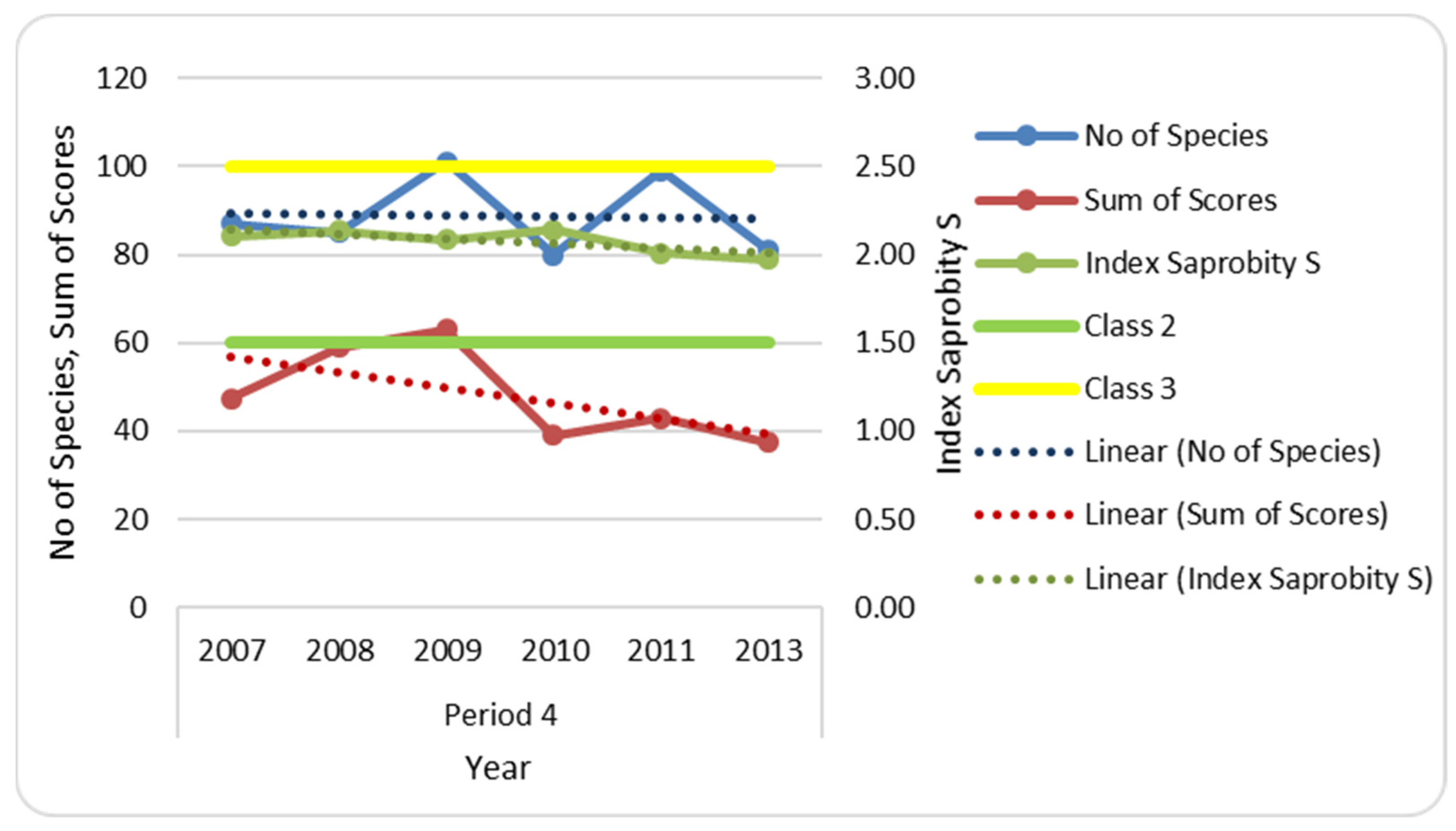
3.5. Index Saprobity S and Organic Pollution
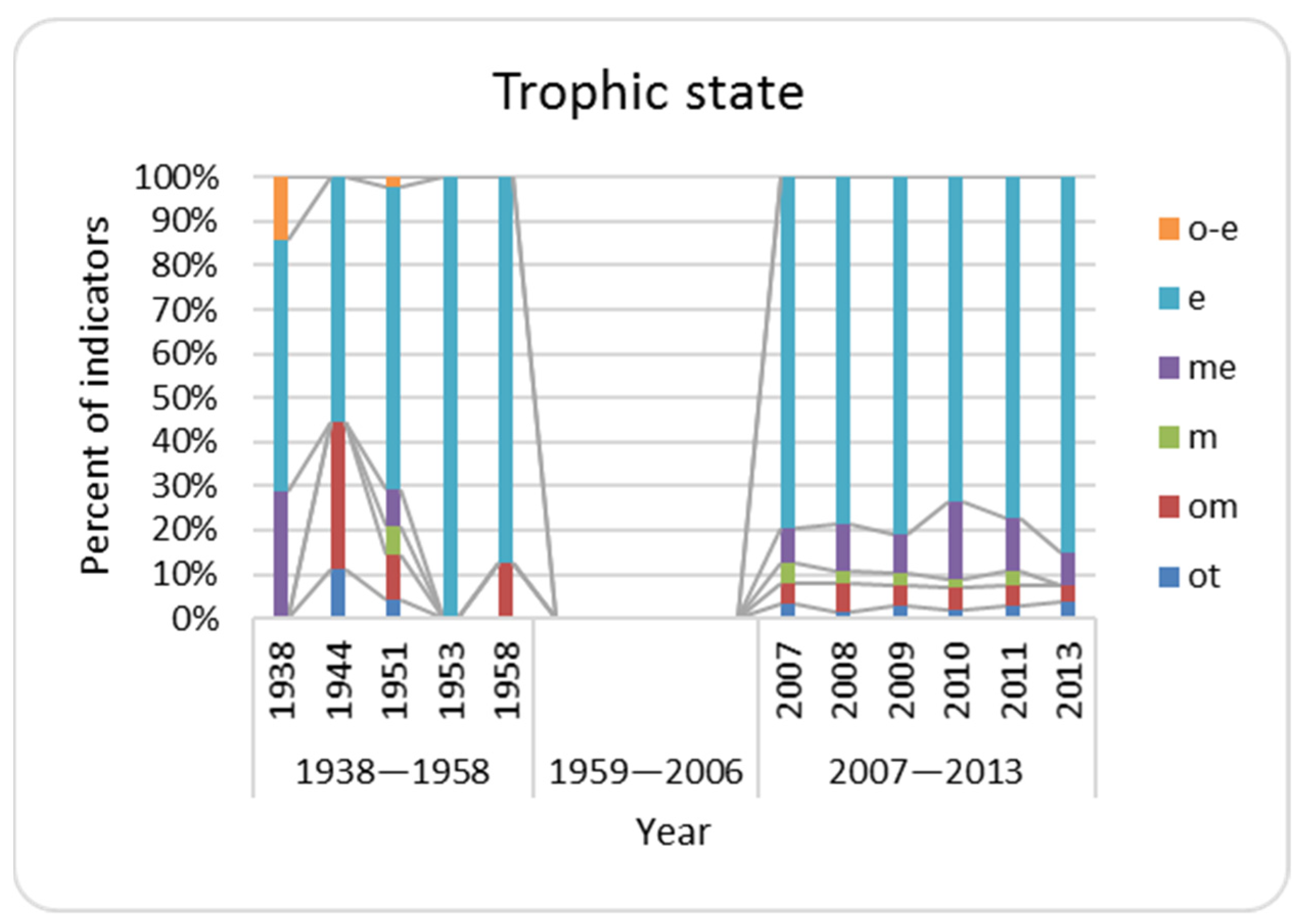
3.6. Trophic State Indices
4. Discussion
5. Conclusions
- For the first time, for long-term bioindication analysis used compiled data from references previous decades published and modern monitoring results;
- Despite the sporadical character of algae and cyanobacteria studies in the Hula Lake/Hula Nature Reserve, 225 species and intraspecific taxa belonging to eight phyla were revealed in 1938–2013. This species list is fairly large for such a small territory but is still far from exhaustion and can be enriched with subsequent works;
- Comparison of Lake Hula and Hula Nature Reserve algae and cyanobacteria flora reveals many similarities; however, the bioindication noticed salinity and organic pollution increase in a modern time. The increase seems to be caused by water flowing into the Hula Nature Reserve water body;
- The use of bioindication methods and our own created database of aquatic flora species ecological preferences made it possible to mark the environmental dynamics of water bodies only based on the species composition of algae and cyanobacteria. Our work shows the special relevance of bioindication for monitoring aquatic ecosystems in situations of impossibility to obtain hydrological data and count the exact species abundance and biomass.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Junk, W.J.; Shuqing, A.; Finlayson, C.M.; Gopal, B.; Kvet, J.; Stephen, A.; Mitchell, S.A.; Mitsch, W.J.; Robarts, R.D. Current state of knowledge regarding the world’s wetlands and their future under global climate change: A synthesis. Aquat. Sci. 2013, 75, 151–167. [Google Scholar] [CrossRef] [Green Version]
- Pollingher, U.; Zohary, T.; Fishbein, T. Algal flora in the Hula valley—Past and present. Isr. J. Plant Sci. 1998, 46, 155–168. [Google Scholar] [CrossRef]
- Payne, R.J. A longer-term perspective on human exploitation and management of peat wetlands: The Hula Valley, Israel. Mires Peat. 2012, 4, 1–9. [Google Scholar]
- Hambright, K.D.; Zohary, T. Lakes Hula and Agmon: Destruction and creation of wetland ecosystems in northern Israel. Wetl. Ecol. Manag. 1998, 6, 83–89. [Google Scholar] [CrossRef]
- Freshwater Ecoregions of the World, (FEOW). Available online: https://www.feow.org/ (accessed on 21 September 2021).
- Hambright, K.D.; Zohary, T. The Hula Valley (Northern Israel) Wetlands Rehabilitation Project. In An International Perspective on Wetland Rehabilitation; Streever, W., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999; pp. 73–180. [Google Scholar]
- Cohen-Shacham, E.; Dayan, T.; Feitelson, E.; de Groot, R.S. Ecosystem service trade-offs in wetland management: Drainage and rehabilitation of the Hula, Israel. Hydrol. Sci. 2011, 56, 1582–1601. [Google Scholar] [CrossRef]
- The List of Wetlands of International Importance. Published 6 July 2021. Available online: https://www.ramsar.org/sites/default/files/documents/library/sitelist (accessed on 21 September 2021).
- Rayss, T.; Katchalsky, E. On the plankton in Lake Hula. Hatevah Ve’Haaretz 1938, 5, 483–490. (In Hebrew) [Google Scholar]
- Rayss, T. Materiaux pour Ia flore algologique de Ia Palestine I. Les cyanophycees. Palest. J. Bot. Jerus. Ser. 1944, 3, 94–113. (In French) [Google Scholar]
- Rayss, T. Materiaux pour Ia flore algologique de Ia Palestine II. Les algues des eaux con tinentales. Palest. J. Bot. Jerusalem Ser. 1951, 5, 71–95. (In French) [Google Scholar]
- Komarovsky, B. To the knowledge of the Hula plankton. Alon Hadayagim 1958, 15, 20–28. (In Hebrew) [Google Scholar]
- Ehrlich, A. Quarternary diatoms of the Hula basin (Northern Israel). Geol. Surv. Lsr. Bull. Jerus. Isr. 1973, 58, 1–39. [Google Scholar]
- Sherman, J.W.; Patrick, R. The waters of Merom: A study of Lake Huleh, 7. Diatom stratigraphy of the 54-m core. Arch. Hydrobiol. 1981, 92, 199–221. [Google Scholar]
- Directive 2000/60/EC of the European Parliament and of the Council establishing a framework for Community action in the field of water policy. Off. J. Eur. Communities 2000, 327, 1–72. Available online: http://ec.europa.eu/environment/water/water-framework/index_en.html (accessed on 21 September 2021).
- UNEP/IPCS. Training Module No. 3. Section C, Ecological Risk Assessment, Prepared by The Edinburgh Centre for Toxicology. Available online: https://www.epa.gov/risk/ecological-risk-assessment (accessed on 21 September 2021).
- Carlson, R.E. A trophic state index for lakes. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Bomfim, E.O.; Kraus, C.N.; Bonnet, M.P.; Lobo, M.T.M.P.S.; Nogueira, I.S.; Peres, L.G.M.; Boaventura, G.R.; Laques, A.E.; Garnier, J.; Marques, D.M. Trophic state index validation based on phytoplankton functional groups approach in Amazon floodplains lakes. Inland Waters 2019, 9, 309–319. [Google Scholar] [CrossRef]
- Bere, T. Are diatom-based biotic indices developed in eutrophic, organically enriched waters reliable monitoring metrics in clean waters? Ecol. Indic. 2016, 62, 312–316. [Google Scholar] [CrossRef]
- Gal, G.; Zohary, T. Development and application of a sustainability index for a lake ecosystem. Hydrobiologia 2017, 800, 207–223. [Google Scholar] [CrossRef]
- Nygaard, G. Hydrobiological studies on some Danish ponds and lakes, II: The quotient hypothesis and some little known or new phytoplankton organisms. K. Dan. Vidensk. Selsk. 1949, 7, 1–242. [Google Scholar]
- Zohary, M.; Orshansky, G. The vegetation of the Huleh plain. Palest. J. Bot. Jerus. Ser. 1947, 4, 90–104. [Google Scholar]
- Livneh, M. The Hula drainage project. Ecol. Environ. 1994, 4, 211–219. (In Hebrew) [Google Scholar]
- Salingar, Y. Hula Project Base Map; GIS Division, Keren Kayemet L’Israel: Israel, Jerusalem, 1995. [Google Scholar]
- Zohary, T.; Hambright, K.D. Lake Hula Lake Agmon. The Israel Review of Arts and Letters 1999, 109. Available online: https://www.mfa.gov.il/MFA/MFA-Archive/1999/Pages/Lake%20Hula%20%E2%80%93%20Lake%20Agmon.aspx (accessed on 23 September 2021).
- Avisar, D.; Fox, A.S. Chemical evolution and vegetation response in an altered wetland ecosystem, Hula Valley, Israel (1988–2004). J. Environ. Sci. Health 2012, 47, 1096–1103. [Google Scholar] [CrossRef]
- Huber–Pestalozzi, G. Das Phytoplankton des Süßwassers: Systematik und Biologie, Teil 4., Euglenophycean; E. Schweizerbart’sche Verlagsbuchhandlung (Nägele u. Obermiller): Stuttgart, Germany, 1969; pp. 1–405. [Google Scholar]
- Ettl, H. Xantophyceae. In Süßwasserflora von Mitteleuropa 3 (1); Gustav Fisher Verlag: Stuttgart, Germany, 1978; 530p. [Google Scholar]
- Komárek, J.; Fott, B. Chlorophyceae (Griinalgen), Ordnung Chlorococcales. In Huber-Pestalozzi, G. Das Phytoplankton des Siisswassers 16, Teil 7(1). Die Binnengewasser; Schweizerbartsche Buchhandl: Stuttgart, Germany, 1983; Volume XVI, 1044p. [Google Scholar]
- Cox, E.J. Identification of Freshwater Diatoms from Live Material; Chapman & Hall: London, UK, 1996; 158p. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 1, Teil: Chroococcales. In Süßwasserflora von Mitteleuropa, Bd. 19/1; Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1999; 548p. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2. Teil: Oscillatoriales. In Süßwasserflora von Mitteleuropa, Bd. 19(2); Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Elsevier GmbH: München, Germany, 2005; 759p. [Google Scholar]
- John, D.M.; Whitton, B.A.; Brook, A.J. (Eds.) The Freshwater Algal Flora of the British Isles: An Identification Guide to Freshwater and Terrestrial Alga; Cambridge University Press: Cambridge, UK, 2002; pp. 1–702. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org (accessed on 21 September 2021).
- Korde, N.V. The methods of biological studies for the bottom deposits of lakes (the field methods of biological analysis). In Life in Fresh Waters of USSR; Pavlovsky, E.N., Zhadin, V.I., Eds.; Nauka Press: Moscow, Russia, 1956; pp. 383–413. [Google Scholar]
- Barinova, S. How to Align and Unify the Cell Counting of Organisms for Bioindication. Int. J. Environ. Sci. Nat. Resour. 2017, 2, 555585. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, G.; Chapman, D.; Beim, A.; UNESCO/WHO/UNEP. Chapter 5, The use of biological material. In Water Quality Assessments—A Guide to Use of Biota, Sediments and Water in Environmental Monitoring, 2nd ed.; Chapman, D., Ed.; E & FN Spon: London, UK, 1996; 626p, Available online: https://apps.who.int/iris/handle/10665/41850 (accessed on 21 September 2021).
- Barinova, S. Systemic Criteria for the Analysis of Alpha- and Gamma-Diversity of Freshwater Algae. Int. J. Environ. Sci. Nat. Res. 2017, 4, 555633. [Google Scholar] [CrossRef]
- Barinova, S.S.; Belous, Y.P.; Tsarenko, P.M. Algal Indication of Water Bodies in Ukraine; Haifa University Press: Haifa, Israel; Kiev, Ukraine, 2019; pp. 1–367. (In Russian) [Google Scholar]
- Sládeček, V. System of water quality from the biological point of view. Arch. Für Hydrobiologie. Beih. Ergeb. Der Limnol. 1973, 7, 1–218. [Google Scholar]
- Sládeček, V. Diatoms as indicators of organic pollution. Acta Hydroch. Hydrobiol. 1986, 14, 555–566. [Google Scholar] [CrossRef]
- Barinova, S. On the Classification of Water Quality from an Ecological Point of View. Int. J. Environ. Sci. Nat. Resour. 2017, 2, 1–8. [Google Scholar] [CrossRef]
- Love, J.; Selker, R.; Marsman, M.; Jamil, T.; Dropmann, D.; Verhagen, A.J.; Ly, A.; Gronau, Q.F.; Smira, M.; Epskamp, S.; et al. JASP: Graphical statistical software for common statistical designs. J Stat. Softw. 2019, 88, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J. Biodiversity improves water quality through niche partitioning. Nature 2011, 472, 86–89. [Google Scholar] [CrossRef]
- Watson, S.B.; McCauley, E.; Dawning, J.A. Patterns in phytoplankton taxonomic composition across temperate lakes of different nutrient status. Limnol. Oceanogr. 1997, 42, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Katsiapia, M.; Moustaka-Gouni, M.; Sommer, U. Assessing ecological water quality of freshwaters: PhyCoI—a new phytoplankton community Index. Ecol. Inform. 2016, 31, 22–29. [Google Scholar] [CrossRef]
- Jones, R.F. Report of the Percy Sladen Expedition to Lake Huleh: A Contribution to the Study of the Fresh Waters of Palestine: The Plant Ecology of the District. J. Ecol. 1940, 28, 357–376. [Google Scholar] [CrossRef]
- Nishri, A. Long-Term Impacts of Draining a Watershed Wetland on a Downstream Lake, Lake Kinneret, Israel. Air Soil Water Res. 2011, 4, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Singh, Y.; Singh, G.; Khattar, J.S.; Barinova, S.; Kaur, J.; Kumar, S.; Singh, D.P. Assessment of water quality condition and spatiotemporal patterns in selected wetlands of Punjab, India. Environ. Sci. Pollut. Res. Int. 2021, 9, 1–17. [Google Scholar] [CrossRef]
- Foti, R.; del Jesus, M.; Rinaldo, A.; Rodriguez-Iturbe, I. Signs of critical transition in the Everglades wetlands in response to climate and anthropogenic changes. Proc. Natl. Acad. Sci. USA 2013, 110, 6296–6300. [Google Scholar] [CrossRef] [Green Version]
- Barinova, S.; Nevo, E. Algal diversity of the Akko Park wetlands in the Bahai Gardens (Haifa, Israel). Transylv. Rev. Syst. Ecol. Res. Wetl. Divers. 2012, 14, 55–79. [Google Scholar]
- Klymiuk, V.; Barinova, S.; Fatiukha, A. Algal bio-indication in assessment of hydrological impact on ecosystem in wetlands of “Slavyansky Resort”. Transylv. Rev. Syst. Ecol. Res. Wetl. Divers. 2015, 17, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Barinova, S.; Krupa, E.; Romanova, S. The role of planktonic algae in the ecological assessment of storage-reservoirs of the Ili-Balkhash basin. Transylv. Rev. Syst. Ecol. Res. Wetl. Divers. 2018, 20, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Novoselova, T.; Barinova, S.; Protasov, A. Long-term dynamics of trophic state indicators in phytoplankton of the cooling reservoir of a nuclear power plant. Transylv. Rev. Syst. Ecol. Research. Wetl. Divers. 2021, 23, 1–14. [Google Scholar] [CrossRef]
- Tsarenko, P.M.; Bilous, O.P.; Kryvosheia-Zakharova, O.M.; Lilitska, H.H.; Barinova, S. Diversity of Algae and Cyanobacteria and Bioindication Characteristics of the Alpine Lake Nesamovyte (Eastern Carpathians, Ukraine) from 100 years Ago to the Present. Diversity 2021, 13, 256. [Google Scholar] [CrossRef]
- Sigee, D.C. Freshwater Microbiology: Biodiversity and Dynamic Interactions of Microorganisms in the Aquatic Environment; John Wiley & Sons: Chichester, UK; Hoboken, NJ, USA, 2004; 524p. [Google Scholar] [CrossRef]
- Khalil, S.; Hussain, M.; Ahmed, I.; Shah, M.; Alam, J.; Gul, S.; Islam, M.; Azhar, N.; Bibi, A.; Safeer, S. Conservation and management of lakes and ponds in the perspective of Banjosa Lake Rawalkot, Azad Kashmir, Pakistan. Sylwan 2021, 165, 25–38. [Google Scholar]
- Hambright, K.D.; Bar-Ilan, I.; Eckert, W. General water chemistry and quality in a newly-created subtropical wetland lake. Wetl. Ecol. Manag. 1998, 6, 121–132. [Google Scholar] [CrossRef]
- Hering, D.; Feld, C.K.; Moog, O.; Ofenbock, T. Cook book for the development of a multimeric index for biological condition of aquatic ecosystems: Experiences from the European AQEM and STAR projects and related initiatives. Hydrobiologia 2006, 566, 311–324. [Google Scholar] [CrossRef]
- Laplace-Treyture, C.; Feret, T. Performance of the Phytoplankton Index for Lakes (IPLAC): A multimetric phytoplankton index to assess the ecological status of water bodies in France. Ecol. Indic. 2016, 69, 686–698. [Google Scholar] [CrossRef] [Green Version]
- Marchetto, A.; Bachisio, M.; Padedda, B.M.; Mariani, M.A.; Luglie, A.; Sechi, N. A numerical index for evaluating phytoplankton response to changes in nutrient levels in deep mediterranean reservoirs. J. Limnol. 2009, 68, 106–121. [Google Scholar] [CrossRef] [Green Version]
- Barbiero, R.P. A multi-lake comparison of epilithic diatom communities on natural and artificial substrates. Hydrobiologia 2000, 438, 157–170. [Google Scholar] [CrossRef]
- Serruya, C.; Pollingher, U. Lakes of the Warm Belt; Cambridge University Press: Cambridge, UK; London, UK, 1983; 569p. [Google Scholar]
- Barinova, S.; Tavassi, M.; Glassman, H.; Nevo, E. Algal indication of pollution in the Lower Jordan River, Israel. Appl. Ecol. Environ. Res. 2010, 8, 19–38. [Google Scholar] [CrossRef]
- Barinova, S.S.; Anissimova, O.V.; Nevo, E.; Jarygin, M.M.; Wasser, S.P. Diversity and Ecology of Algae from Nahal Qishon, Northern Israel. Plant Biosyst. 2004, 138, 245–259. [Google Scholar] [CrossRef]
- Barinova, S.S.; Tavassi, M.; Nevo, E. Algal communities of the Hadera River (Israel) under dramatic niche changes. Cent. Eur. J. Biol. 2010, 5, 507–521. [Google Scholar] [CrossRef]
- Barinova, S.S.; Nevo, E. The Upper Jordan River algal communities are evidence of long-term climatic and anthropogenic impacts. J. Water Resour. Prot. 2010, 2, 507–526. [Google Scholar] [CrossRef] [Green Version]
- Barinova, S.; Krassilov, V.A. Algal diversity and bio-indication of water resources in Israel. Int. J. Environ. Resour. 2012, 1, 62–72. [Google Scholar]
- Florescu, M.H.; Maxim, M.M.; Momeu, L.; Cîmpean, M.; Battes, K.P. Wetland algal communities from Balta Mică a Brăilei Nature Park (Romania). Transylv. Rev. Syst. Ecol. Res. Wetl. Divers. 2015, 17, 23–44. [Google Scholar] [CrossRef] [Green Version]
- Barinova, S.; Romanov, R. Charophyte Communities in the Ein Afeq Natural Reserve, Israel. Nat. Resour. Conserv. 2015, 3, 31–44. [Google Scholar] [CrossRef]
- Bănăduc, D.; Curtean-Bănăduc, A.; Cianfaglione, K.; Akeroyd, J.R.; Cioca, L.-I. Proposed Environmental Risk Management Elements in a Carpathian Valley Basin, within the Roşia Montană European Historical Mining Area. Int. J. Environ. Res. Public Health 2021, 18, 4565. [Google Scholar] [CrossRef] [PubMed]
- Török, Z. The Romanian Wetland Inventory Project. Analele Științifice/Sci. Ann. 2003, 9, 161–173. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Observed Cells | Estimate | Score |
|---|---|---|
| 1–5 cells/chamber | Occasional | 1 |
| 10–15 cells/chamber | Rare | 2 |
| 25–30 cells/chamber | Common | 3 |
| 1 cell over a transect | Frequent | 4 |
| Several cells over a transect | Very frequent | 5 |
| A cell in every field of view | Abundant | 6 |
| Trophic State | Index 1 | Index 2 | Index 3 | Index 4 | Index 5 |
|---|---|---|---|---|---|
| Oligotrophic-dystrophic phase | 0 | 0.0–0.3 | 0 | 0 | 0–0.3 |
| Oligotrophic-acidotrophic phase | 0 | 0–0.1 | 0 | 0 | 0–0.1 |
| Oligotrophic | 0–0.4 | 0–0.7 | 0 | 0–0.2 | 0.25–1.0 |
| Mesotrophic | 0.1–0.5 | 0.2–0.6 | 0–0.75 | 0.1–1.0 | 1.1–1.2 |
| Slightly eutrophic | 0.8–1.0 | 0.7–1.0 | 0.2–1.5 | 0–0.2 | 2.0–2.25 |
| Moderately eutrophic | 1.4–2.0 | 1.25–1.4 | 0.6–1.7 | 0 | 3.5–4.4 |
| Eutrophic | 1.2–3.0 | 2.1–3.5 | 1.25–3.0 | 0 | 4.3–8.75 |
| Eutrophic of mixotrophic phase | 0.9–2.7 | 2.2–2.5 | 0.2–0.5 | 0 | 3.3–5.3 |
| NS | QG | Trophic State |
|---|---|---|
| 0–2 | >60 | Ultraoligotrophic |
| 2–4 | 46–60 | Oligotrophic |
| 4–6 | 31–45 | Mesotrophic |
| 6–8 | 16–30 | Eutrophic |
| >8 | <16 | Hypertrophic |
| Parameter | 1938–1958 [2,47,48] | 2011 Our Data | 2011 Average Our Data |
|---|---|---|---|
| Water temperature, °C | 4–36 | 19.9–25.7 | 22.50 |
| TDS, mg L−1 | 224–373 | 255–1675 | 734.75 |
| Sulfate, mg L−1 | ≤373 | nd | nd |
| Nitrate-N, mg L−1 | nd | 1.6–7.7 | 5.67 |
| Ammonia-N, mg L −1 | 0.06 | nd | nd |
| Chloride, mg L−1 | 15–50 | nd | nd |
| pH | 7.2–8.6 | 7.1–7.9 | 7.61 |
| Electrical conductivity, ms cm−1 | nd | 0.36–2.26 | 1.01 |
| Phylum | 1938 | 1944 | 1951 | 1953 | 1958 | 2007 | 2008 | 2009 | 2010 | 2011 | 2013 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Hula before drainage | Hula Nature Reserve monitoring | ||||||||||
| Bacillariophyta | 5 | 0 | 27 | 0 | 2 | 11 | 11 | 12 | 12 | 15 | 10 |
| Charophyta | 2 | 0 | 2 | 0 | 0 | 5 | 6 | 6 | 4 | 5 | 3 |
| Chlorophyta | 0 | 0 | 26 | 5 | 2 | 39 | 33 | 37 | 28 | 38 | 33 |
| Cryptophyta | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 2 | 2 | 2 | 1 |
| Cyanobacteria | 0 | 9 | 0 | 0 | 4 | 15 | 16 | 18 | 15 | 19 | 14 |
| Euglenozoa | 1 | 0 | 7 | 0 | 0 | 8 | 9 | 10 | 8 | 12 | 11 |
| Miozoa | 1 | 0 | 3 | 0 | 0 | 5 | 3 | 4 | 3 | 3 | 4 |
| Ochrophyta (Chrysophyceae) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Ochrophyta (Eustigmatophyceae) | 0 | 0 | 0 | 0 | 0 | 2 | 3 | 7 | 4 | 4 | 4 |
| Ochrophyta (Xanthophyceae) | 0 | 0 | 3 | 0 | 0 | 1 | 2 | 3 | 4 | 1 | 1 |
| Ochrophyta (Synurophyceae) | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| No of Species | 9 | 9 | 71 | 5 | 8 | 87 | 85 | 101 | 80 | 99 | 81 |
| Average Sum of Scores | 47.35 | 59.01 | 63.01 | 39 | 42.8 | 37.41 | |||||
| Index Saprobity S | 2.10 | 2.13 | 2.09 | 2.14 | 2.01 | 1.97 | |||||
| Class of Water Quality | |||||||||||
| Class 2 | 1 | 1 | 9 | 1 | 0 | 11 | 5 | 10 | 6 | 9 | 6 |
| Class 3 | 4 | 7 | 36 | 4 | 8 | 52 | 52 | 63 | 50 | 58 | 49 |
| Class 4 | 2 | 0 | 5 | 0 | 0 | 6 | 8 | 7 | 6 | 7 | 4 |
| Class 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Watanabe | |||||||||||
| sx | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| es | 3 | 0 | 12 | 0 | 2 | 8 | 9 | 9 | 8 | 10 | 7 |
| sp | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 |
| Oxygen | |||||||||||
| aer | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 1 | 1 | 2 |
| str | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| st-str | 7 | 0 | 48 | 5 | 5 | 45 | 45 | 49 | 41 | 50 | 45 |
| st | 2 | 0 | 19 | 0 | 2 | 10 | 11 | 12 | 9 | 12 | 10 |
| Temperature | |||||||||||
| cool | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| temp | 3 | 0 | 17 | 0 | 1 | 7 | 7 | 7 | 7 | 10 | 6 |
| eterm | 1 | 0 | 3 | 0 | 2 | 5 | 6 | 6 | 4 | 6 | 5 |
| warm | 0 | 1 | 2 | 0 | 0 | 2 | 2 | 2 | 1 | 3 | 2 |
| Habitat | |||||||||||
| B | 2 | 0 | 7 | 0 | 0 | 2 | 3 | 3 | 4 | 5 | 3 |
| P-B | 7 | 9 | 44 | 4 | 6 | 48 | 47 | 53 | 44 | 51 | 44 |
| P | 0 | 0 | 19 | 1 | 4 | 30 | 28 | 33 | 24 | 35 | 27 |
| Salinity | |||||||||||
| hb | 0 | 0 | 3 | 0 | 0 | 2 | 1 | 1 | 2 | 2 | 2 |
| i | 6 | 3 | 45 | 5 | 6 | 41 | 38 | 41 | 34 | 41 | 34 |
| hl | 1 | 1 | 1 | 0 | 1 | 4 | 4 | 3 | 4 | 2 | 3 |
| mh | 1 | 0 | 3 | 0 | 1 | 1 | 5 | 4 | 4 | 4 | 4 |
| pH | |||||||||||
| acf | 0 | 1 | 3 | 0 | 1 | 4 | 1 | 3 | 1 | 2 | 1 |
| ind | 3 | 2 | 21 | 2 | 3 | 21 | 23 | 25 | 24 | 22 | 18 |
| alf | 3 | 0 | 15 | 0 | 2 | 13 | 12 | 10 | 10 | 14 | 9 |
| alb | 0 | 1 | 0 | 0 | 0 | 2 | 1 | 2 | 3 | 3 | 2 |
| Autotropy-Heterotrophy | |||||||||||
| ats | 0 | 0 | 7 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 1 |
| ate | 3 | 0 | 11 | 0 | 2 | 7 | 7 | 6 | 7 | 9 | 5 |
| hne | 1 | 0 | 2 | 0 | 0 | 3 | 4 | 4 | 4 | 3 | 3 |
| hce | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Trophic State | |||||||||||
| ot | 0 | 1 | 2 | 0 | 0 | 2 | 1 | 2 | 1 | 2 | 2 |
| om | 0 | 3 | 5 | 0 | 1 | 3 | 4 | 3 | 3 | 3 | 2 |
| m | 0 | 0 | 3 | 0 | 0 | 3 | 2 | 2 | 1 | 2 | 0 |
| me | 2 | 0 | 4 | 0 | 0 | 5 | 7 | 6 | 10 | 8 | 4 |
| e | 4 | 5 | 33 | 5 | 7 | 50 | 51 | 55 | 42 | 51 | 45 |
| o-e | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Hula before Drainage | Hula Nature Reserve Monitoring | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Index | 1938 | 1944 | 1951 | 1953 | 1958 | 2007 | 2008 | 2009 | 2010 | 2011 | 2013 |
| NS | na | na | 6.6 | na | na | 12.4 | 11.6 | 9.3 | 12.8 | 17.3 | 29.0 |
| Trophic state | na | na | e | na | na | hy | hy | hy | hy | hy | hy |
| QG | 6 | 0 | 48 | 0 | 2 | 27 | 26 | 38 | 25 | 28 | 19 |
| Trophic state | hy | hy | o | hy | hy | e | e | m | e | e | e |
| NY | |||||||||||
| index 1 | na | na | 0 | na | na | 3 | 3.2 | 3.6 | 3.75 | 4.75 | 7 |
| index 2 | na | na | 11 | na | na | 7.6 | 6.4 | 7 | 6.75 | 8.75 | 16 |
| index 3 | 0.67 | na | 0.24 | na | 1 | 0.38 | 0.57 | 0.5 | 0.33 | 0.36 | 0.43 |
| index 4 | 0.5 | 0 | 0.32 | 0 | 0.4 | 0.15 | 0.19 | 0.19 | 0.19 | 0.22 | 0.24 |
| index 5 | na | na | 17 | na | na | 12.8 | 12.2 | 13.4 | 13.25 | 17.5 | 30 |
| Trophic state | |||||||||||
| index 1 | na | na | o | na | na | e | e | na | na | na | na |
| index 2 | na | na | na | o | na | na | na | na | na | na | na |
| index 3 | m | na | m | o | se | m | m | m | m | m | m |
| index 4 | m | o | m | o | m | o | o | o | o | m | m |
| index 5 | na | na | na | na | na | na | na | na | na | na | na |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barinova, S.; Alster, A. Algae and Cyanobacteria Diversity and Bioindication of Long-Term Changes in the Hula Nature Reserve, Israel. Diversity 2021, 13, 583. https://doi.org/10.3390/d13110583
Barinova S, Alster A. Algae and Cyanobacteria Diversity and Bioindication of Long-Term Changes in the Hula Nature Reserve, Israel. Diversity. 2021; 13(11):583. https://doi.org/10.3390/d13110583
Chicago/Turabian StyleBarinova, Sophia, and Alla Alster. 2021. "Algae and Cyanobacteria Diversity and Bioindication of Long-Term Changes in the Hula Nature Reserve, Israel" Diversity 13, no. 11: 583. https://doi.org/10.3390/d13110583
APA StyleBarinova, S., & Alster, A. (2021). Algae and Cyanobacteria Diversity and Bioindication of Long-Term Changes in the Hula Nature Reserve, Israel. Diversity, 13(11), 583. https://doi.org/10.3390/d13110583