
Evolution of Beak and Feather Disease Virus across Three Decades of Conservation Intervention for Population Recovery of the Mauritius Parakeet

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mauritius Parakeet Sampling, DNA Extraction, PCR, and Sequencing
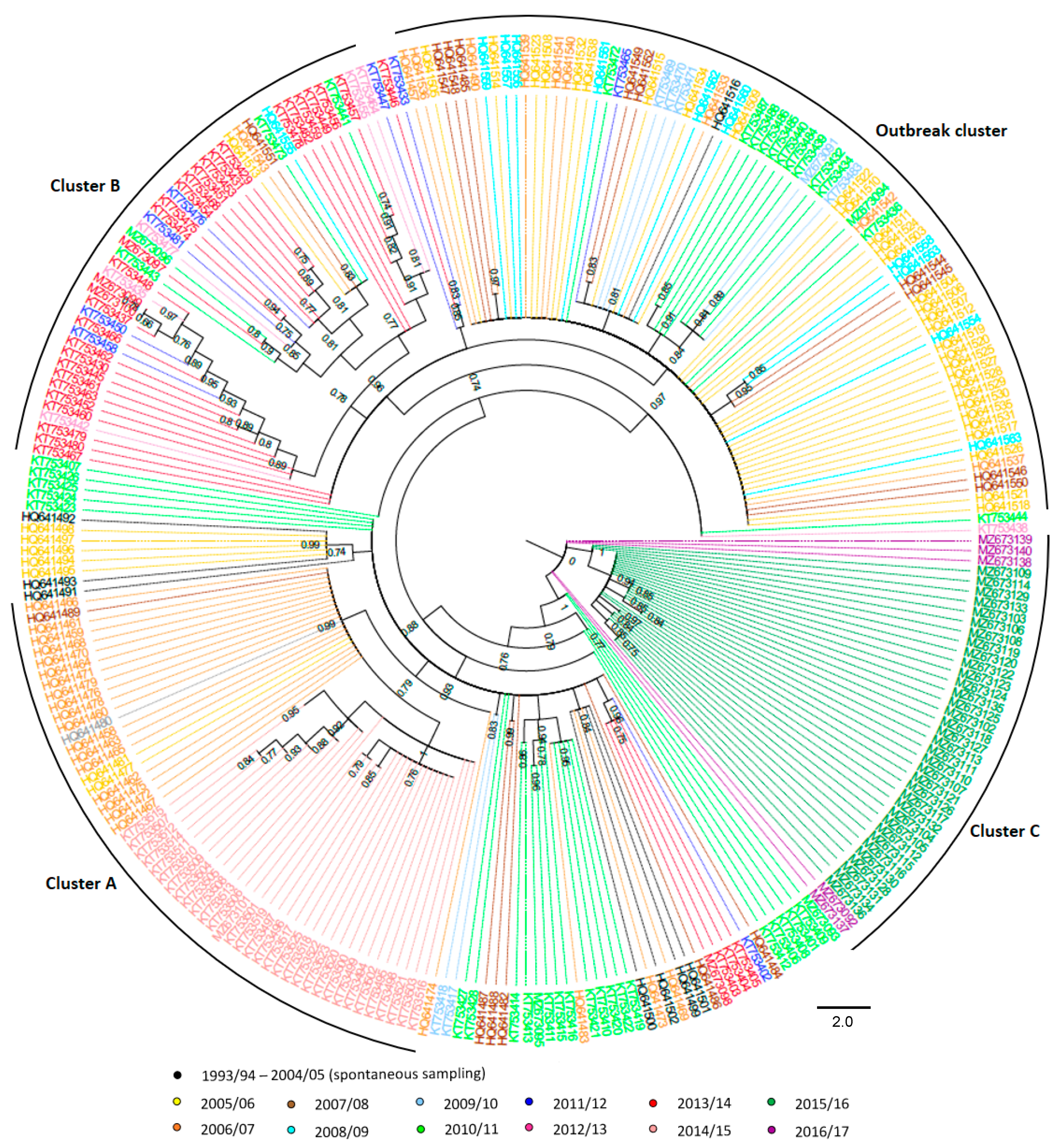
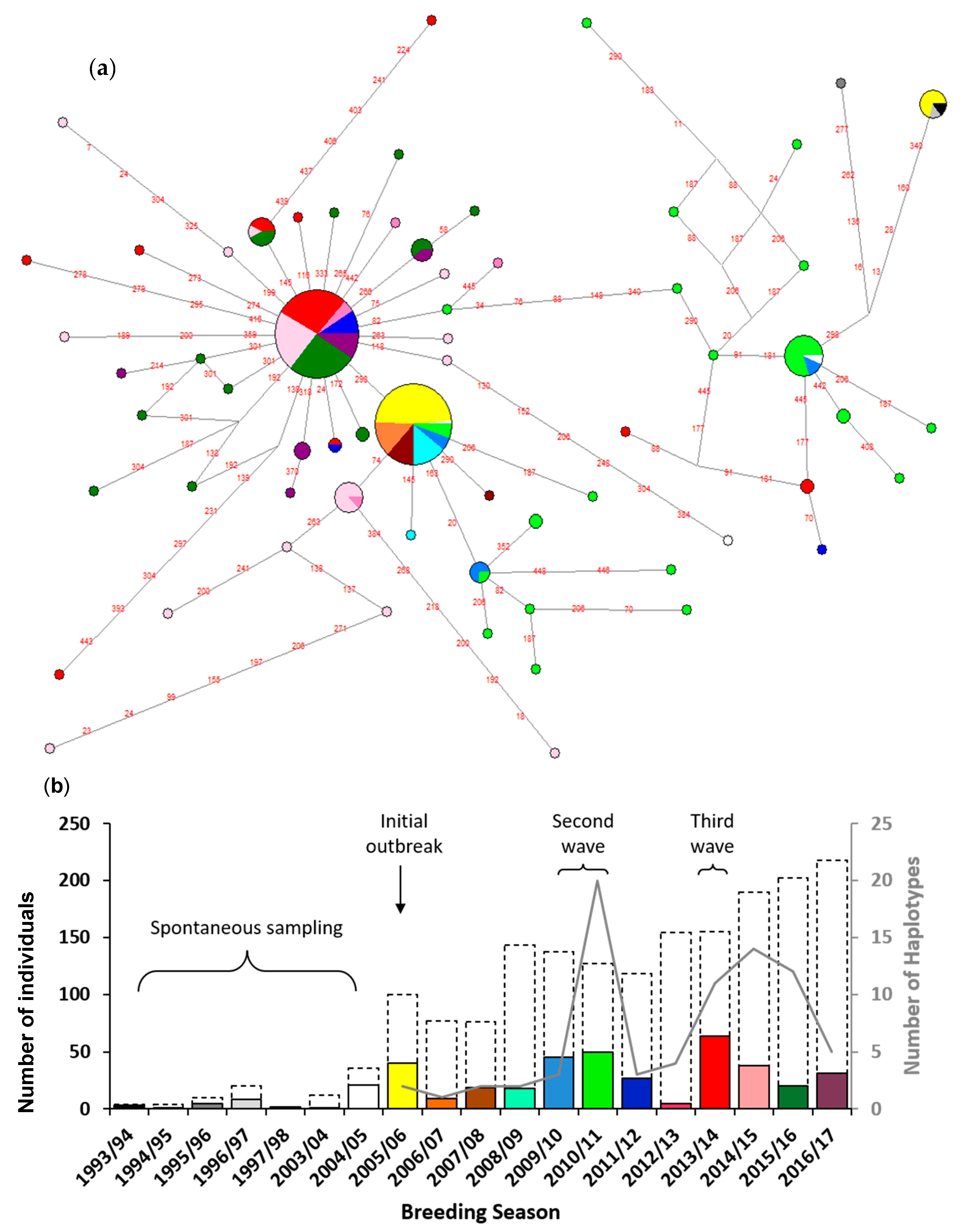
2.2. BFDV Phylogeny and Haplotype Network
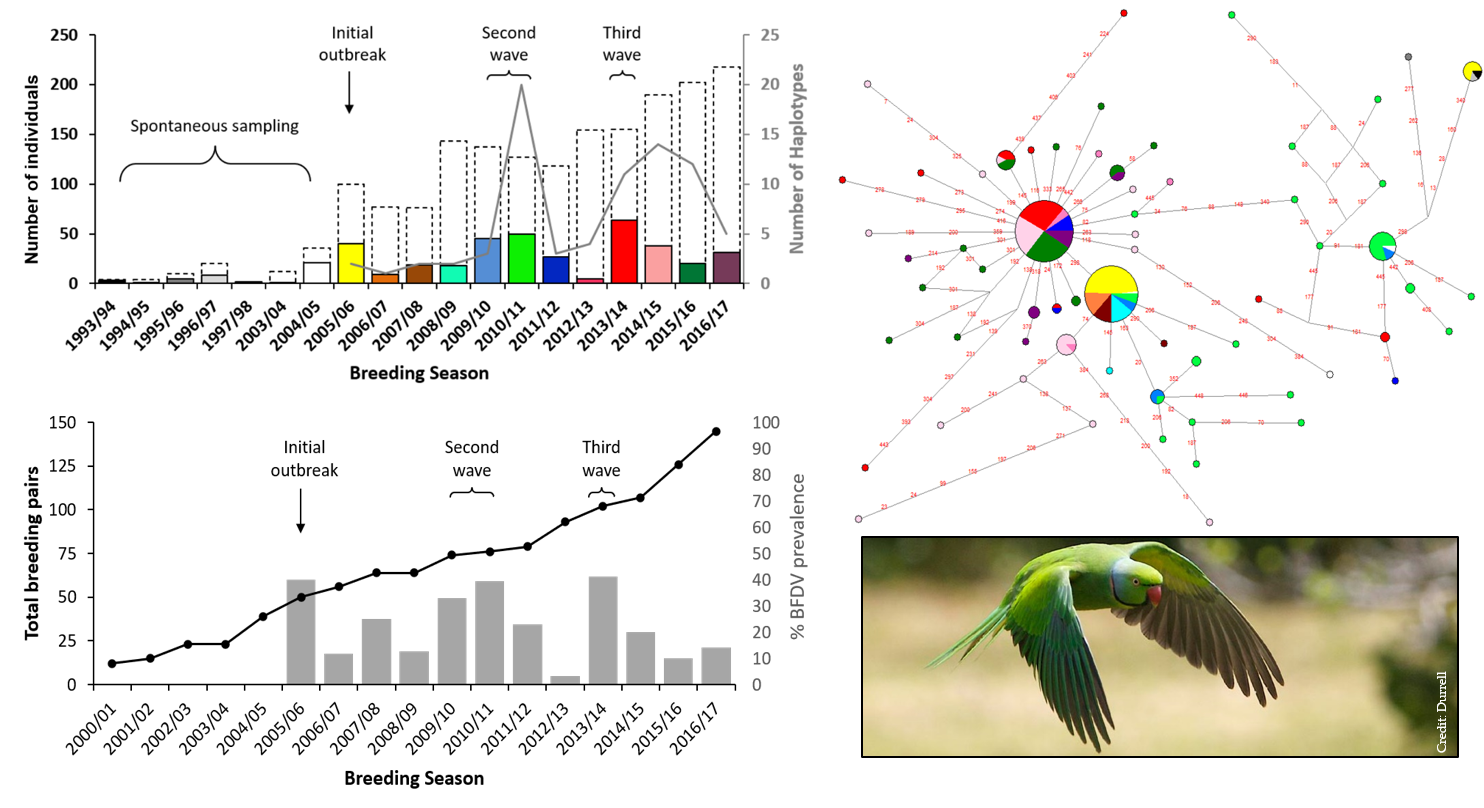
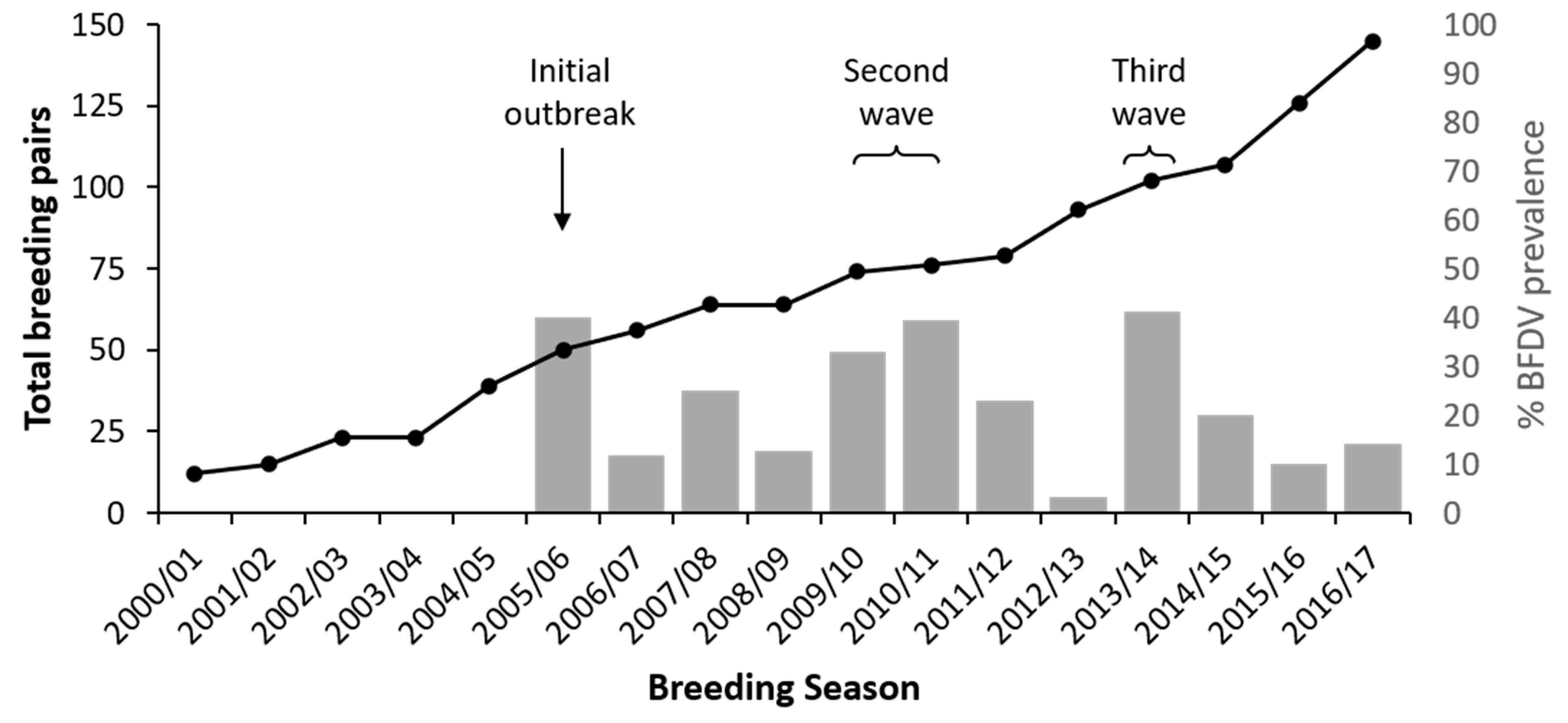
3. Results
4. Discussion
4.1. Multiple Waves of BFDV Infection Following Initial Outbreak
4.2. BFDV in the Context of Ongoing Conservation Management
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brooks, D.R.; Ferrao, A.L. The historical biogeography of co-evolution: Emerging infectious diseases are evolutionary accidents waiting to happen. J. Biogeogr. 2005, 32, 1291–1299. [Google Scholar] [CrossRef]
- Yap, T.A.; Koo, M.S.; Ambrose, R.F.; Wake, D.B.; Vredenburg, V.T. Averting a North American biodiversity crisis. Science 2015, 349, 481–482. [Google Scholar] [CrossRef] [Green Version]
- Lyles, A.M.; Dobson, A.P. Infectious disease and intensive management: Population dynamics, threatened hosts, and their parasites. J. Zoo Wildl. Med. 1993, 24, 315–326. [Google Scholar]
- Lips, K.R.; Brem, F.; Brenes, R.; Reeve, J.D.; Alford, R.A.; Voyles, J.; Carey, C.; Livo, L.; Pessier, A.P.; Collins, J.P. Emerging infectious disease and the loss of biodiversity in a neotropical amphibian community. Proc. Natl. Acad. Sci. USA 2006, 103, 3165–3170. [Google Scholar] [CrossRef] [Green Version]
- Dobson, A.; Foufopoulos, J. Emerging infectious pathogens of wildlife. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2001, 356, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, D.M.; Carver, S.; Jones, M.E.; Krkošek, M.; Skerratt, L.F. Emerging infectious diseases of wildlife: A critical perspective. Trends Parasitol. 2015, 31, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Altizer, S.; Harvell, D.; Friedle, E. Rapid evolutionary dynamics and disease threats to biodiversity. Trends Ecol. Evol. 2003, 18, 589–596. [Google Scholar] [CrossRef]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2020-3. Available online: http://www.iucnredlist.org/ (accessed on 1 August 2021).
- Khalesi, B.; Bonne, N.; Stewart, M.; Sharp, M.; Raidal, S. A comparison of haemagglutination, haemagglutination inhibition and PCR for the detection of Psittacine beak and feather disease virus infection and a comparison of isolates obtained from loriids. J. Gen. Virol. 2005, 86, 3039–3046. [Google Scholar] [CrossRef] [PubMed]
- Pass, D.A.; Perry, R.A. The pathology of Psittacine beak and feather disease. Aust. Vet. J. 1984, 61, 69–74. [Google Scholar] [CrossRef]
- Raidal, S.R.; Sarker, S.; Peters, A. Review of Psittacine beak and feather disease and its effect on Australian endangered species. Aust. Vet. J. 2015, 93, 466–470. [Google Scholar] [CrossRef]
- Ritchie, B.W.; Niagro, F.D.; Lukert, P.D.; Steffens, W.L., III; Latimer, K.S. Characterization of a new virus from cockatoos with psittacine beak and feather disease. Virology 1989, 171, 83–88. [Google Scholar] [CrossRef]
- Sarker, S.; Ghorashi, S.A.; Forwood, J.K.; Bent, S.J.; Peters, A.; Raidal, S.R. Phylogeny of beak and feather disease virus in cockatoos demonstrates host generalism and multiple-variant infections within Psittaciformes. Virology 2014, 460, 72–82. [Google Scholar] [CrossRef]
- Kondiah, K.; Albertyn, J.; Bragg, R.R. Genetic diversity of the rep gene of Beak and feather disease virus in South Africa. Arch. Virol. 2006, 151, 2539–2545. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.; Faulkes, C.G.; Greenwood, A.G.; Jones, C.G.; Kaiser, P.; Lyne, O.D.; Black, S.A.; Chowrimootoo, A.; Groombridge, J.J. Tracking viral evolution during a disease outbreak: The rapid and complete selective sweep of a Circovirus in the endangered Echo parakeet. J. Virol. 2012, 86, 5221–5229. [Google Scholar] [CrossRef] [Green Version]
- Heath, L.; Martin, D.P.; Warburton, L.; Perrin, M.; Horsfield, W.; Kingsley, C.; Rybicki, E.P.; Williamson, A.-L. Evidence of unique genotypes of Beak and feather disease virus in Southern Africa. J. Virol. 2004, 78, 9277–9284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, P.A.; Anderson, I.L.; Lambert, D.M. Evidence for specificity of Psittacine beak and feather disease viruses among avian hosts. Virology 2003, 306, 109–115. [Google Scholar] [CrossRef]
- Ritchie, B.W.; Niagro, F.D.; Lukert, P.D.; Latimer, K.S.; Steffens III, W.L.; Pritchard, N. A review of Psittacine beak and feather disease: Characteristics of the PBFD virus. J. Assoc. Avian Vet. 1989, 3, 143–150. [Google Scholar] [CrossRef]
- Peters, A.; Patterson, E.I.; Baker, B.G.B.; Holdsworth, M.; Sarker, S.; Ghorashi, S.A.; Raidal, S.R. Evidence of Psittacine beak and feather disease virus spillover into wild critically endangered Orange-bellied parrots (Neophema chrysogaster). J. Wildl. Dis. 2014, 50, 288–296. [Google Scholar] [CrossRef]
- Sarker, S.; Patterson, E.I.; Peters, A.; Baker, G.B.; Forwood, J.K.; Ghorashi, S.A.; Holdsworth, M.; Baker, R.; Murray, N.; Raidal, S.R. Mutability dynamics of an emergent single stranded DNA virus in a naïve host. PLoS ONE 2014, 9, e8537. [Google Scholar] [CrossRef] [Green Version]
- Fogell, D.J.; Martin, R.O.; Groombridge, J.J. Beak and feather disease virus in wild and captive parrots: An analysis of geographic and taxonomic distribution and methodological trends. Arch. Virol. 2016, 161, 2059–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wikelski, M.; Foufopoulos, J.; Vargas, H.; Snell, H. Galápagos Birds and Diseases: Invasive Pathogens as Threats for Island Species. Ecol. Soc. 2004, 9, 5. [Google Scholar] [CrossRef]
- Trinkel, M.; Cooper, D.; Packer, C.; Slotow, R. Inbreeding depression increases susceptibility to bovine tuberculosis in lions: An experimental test using an inbred-outbred contrast through translocation. J. Wildl. Dis. 2011, 47, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Spurgin, L.G.; Illera, J.C.; Padilla, D.P.; Richardson, D.S. Biogeographical patterns and co-occurrence of pathogenic infection across island populations of Berthelot’s pipit (Anthus berthelotii). Oecologia 2012, 168, 691–701. [Google Scholar] [CrossRef]
- Russell, J.C.; Kueffer, C. Island Biodiversity in the Anthropocene. Annu. Rev. Environ. Resour. 2019, 44, 31–60. [Google Scholar] [CrossRef]
- Sax, D.F.; Gaines, S.D.; Brown, J.H. Species invasions exceed extinctions on islands worldwide: A comparative study of plants and birds. Am. Nat. 2002, 160, 766–783. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.G. The larger land-birds of Mauritius. In Studies of Mascarene Island Birds; Diamond, A.W., Ed.; Cambridge University Press: Cambridge, UK, 1987; pp. 208–300. [Google Scholar]
- Henshaw, S.; Blackwell, L.; Yeung Shi Chung, C.; Bielsa, M.; Bertille, H. Mauritian Wildlife Foundation: Echo Parakeet (Psittacula eques) Management Report 2017/2018; Mauritian Wildlife Foundation: Vacoas, Mauritius, 2018. [Google Scholar]
- Lovegrove, T.G.; Nieuwland, A.B.; Green, S. Interim Report on the Echo Parakeet Conservation Project; Mauritian Wildlife Foundation: Vacoas, Mauritius, 1995. [Google Scholar]
- Greenwood, A.G. Veterinary support for in situ avian conservation programmes. Bird Conserv. Int. 1996, 6, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Fogell, D.J.; Groombridge, J.J.; Tollington, S.; Canessa, S.; Henshaw, S.; Zuel, N.; Jones, C.G.; Greenwood, A.; Ewen, J.G. Hygiene and biosecurity protocols reduce infection prevalence but do not improve fledging success in an endangered parrot. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tollington, S.; Greenwood, A.; Jones, C.G.; Hoeck, P.; Chowrimootoo, A.; Smith, D.; Richards, H.; Tatayah, V.; Groombridge, J.J. Detailed monitoring of a small but recovering population reveals sublethal effects of disease and unexpected interactions with supplemental feeding. J. Anim. Ecol. 2015, 84, 969–977. [Google Scholar] [CrossRef] [Green Version]
- Hume, J.P. Reappraisal of the Parrots (Aves: Psittacidae) from the Mascarene Islands, with Comments on Their Ecology, Morphology, and Affinities; Magnolia Press: Auckland, New Zealand, 2007; ISBN 1175-5326. [Google Scholar]
- Bruford, M.W.; Hanotte, O.; Brookfield, J.F.Y.; Burke, T. Single-locus and multilocus DNA fingerprinting. In Molecular Genetic Analysis of Populations: A Practical Approach; Hoelzel, A.R., Ed.; IRL Press: Oxford, UK, 1998; pp. 287–336. [Google Scholar]
- Ypelaar, I.; Bassami, M.R.; Wilcox, G.E.; Raidal, S.R. A universal polymerase chain reaction for the detection of Psittacine beak and feather disease virus. Vet. Microbiol. 1999, 68, 141–148. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Stöver, B.C.; Müller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1.4.2. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 17 July 2015).
- Fluxus Technology Ltd Network 10.2.0.0. Available online: http://www.fluxus-engineering.com/netwinfo.htm (accessed on 1 August 2021).
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.; Sánchez-Gracia, A. DNA Sequence Polymorphism v 6.12.03. 2018. Available online: http://www.ub.edu/dnasp/ (accessed on 23 May 2021).
- Fu, Y.X.; Li, W.H. Statistical tests of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef] [PubMed]
- Easterday, W.R. The first step in the success or failure of emerging pathogens. Proc. Natl. Acad. Sci. USA 2020, 117, 29271–29273. [Google Scholar] [CrossRef]
- Fogell, D.J.; Martin, R.O.; Bunbury, N.; Lawson, B.; Sells, J.; McKeand, A.M.; Tatayah, V.; Trung, C.T.; Groombridge, J.J. Trade and conservation implications of new Beak and feather disease virus detection in native and introduced parrots. Conserv. Biol. 2018, 32, 1325–1335. [Google Scholar] [CrossRef] [Green Version]
- Todd, D. Circoviruses: Immunosuppressive threats to avian species: A review. Avian Pathol. 2000, 29, 373–394. [Google Scholar] [CrossRef]
- Raisin, C. Conservation Genetics of the Mauritius Parakeet. Ph.D. Thesis, University of Kent, Kent, UK, 2010. [Google Scholar]
- Lacy, R.C.; Ballou, J.D.; Pollak, J.P. PMx: Software package for demographic and genetic analysis and management of pedigreed populations. Methods Ecol. Evol. 2012, 3, 433–437. [Google Scholar] [CrossRef]
- Raisin, C.; Frantz, A.C.; Kundu, S.; Greenwood, A.G.; Jones, C.G.; Zuel, N.; Groombridge, J.J. Genetic consequences of intensive conservation management for the Mauritius parakeet. Conserv. Genet. 2012, 13, 707–715. [Google Scholar] [CrossRef]
- Duxbury, E.M.L.; Day, J.P.; Vespasiani, D.M.; Thüringer, Y.; Tolosana, I.; Smith, S.C.L.; Tagliaferri, L.; Kamacioglu, A.; Lindsley, I.; Love, L.; et al. Host-pathogen coevolution increases genetic variation in susceptibility to infection. Elife 2019, 8, e46440. [Google Scholar] [CrossRef] [PubMed]
- McCallum, H.; Dobson, A. Disease, habitat fragmentation and conservation. Hungarian Q. 2008, 49, 2041–2049. [Google Scholar] [CrossRef] [Green Version]
- Tatayah, R.V.V.; Malham, J.; Haverson, P.; Van de Wetering, J. Design and provision of nest boxes for echo parakeets Psittacula eques in Black River Gorges National Park, Mauritius. Conserv. Evid. 2007, 4, 16–19. [Google Scholar]
- Julian, L.; Piasecki, T.; Chrzastek, K.; Walters, M.; Muhire, B.; Harkins, G.W.; Martin, D.P.; Varsani, A. Extensive recombination detected among Beak and feather disease virus isolates from breeding facilities in Poland. J. Gen. Virol. 2013, 94, 1086–1095. [Google Scholar] [CrossRef] [Green Version]
- Jackson, B.; Varsani, A.; Holyoake, C.; Jakob-Hoff, R.; Robertson, I.; McInnes, K.; Empson, R.; Gray, R.; Nakagawa, K.; Warren, K. Emerging infectious disease or evidence of endemicity? A multi-season study of Beak and feather disease virus in wild Red-crowned parakeets (Cyanoramphus novaezelandiae). Arch. Virol. 2015, 160, 2283–2292. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fogell, D.J.; Tollington, S.; Tatayah, V.; Henshaw, S.; Naujeer, H.; Jones, C.; Raisin, C.; Greenwood, A.; Groombridge, J.J. Evolution of Beak and Feather Disease Virus across Three Decades of Conservation Intervention for Population Recovery of the Mauritius Parakeet. Diversity 2021, 13, 584. https://doi.org/10.3390/d13110584
Fogell DJ, Tollington S, Tatayah V, Henshaw S, Naujeer H, Jones C, Raisin C, Greenwood A, Groombridge JJ. Evolution of Beak and Feather Disease Virus across Three Decades of Conservation Intervention for Population Recovery of the Mauritius Parakeet. Diversity. 2021; 13(11):584. https://doi.org/10.3390/d13110584
Chicago/Turabian StyleFogell, Deborah J., Simon Tollington, Vikash Tatayah, Sion Henshaw, Houshna Naujeer, Carl Jones, Claire Raisin, Andrew Greenwood, and Jim J. Groombridge. 2021. "Evolution of Beak and Feather Disease Virus across Three Decades of Conservation Intervention for Population Recovery of the Mauritius Parakeet" Diversity 13, no. 11: 584. https://doi.org/10.3390/d13110584
APA StyleFogell, D. J., Tollington, S., Tatayah, V., Henshaw, S., Naujeer, H., Jones, C., Raisin, C., Greenwood, A., & Groombridge, J. J. (2021). Evolution of Beak and Feather Disease Virus across Three Decades of Conservation Intervention for Population Recovery of the Mauritius Parakeet. Diversity, 13(11), 584. https://doi.org/10.3390/d13110584