
Fragmentation of Coastal Grasslands by Plantations and Spontaneous Spread of Invasive Pines in the Southern Pampa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Design and Field Sampling
2.3. Data Analysis
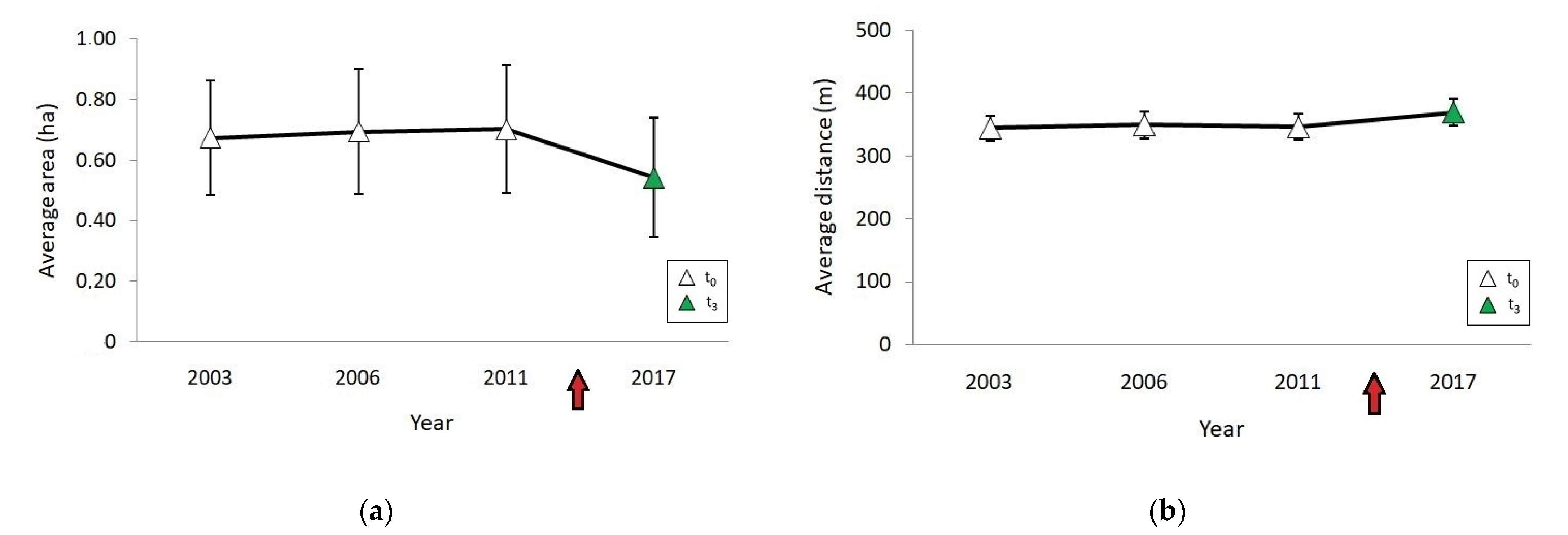
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baldi, G.; Guerschman, J.P.; Paruelo, J.M. Characterizing fragmentation in temperate South America grasslands. Agric. Ecosyst. Environ. 2006, 116, 197–208. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Vasallo, M.; Farley, K.A.; Piñeiro, G.; Garbulsky, M.F.; Nosetto, M.D.; Jackson, R.B.; Paruelo, J.M. Forestación en pastizales: Hacia una visión integral de sus oportunidades y costos ecológicos. Agrociencias 2006, 10, 109–124. [Google Scholar]
- Modernel, P.; Rossing, W.H.A.; Dogliotti, S.; Picasso, V.; Tittonell, P. Land use change and ecosystem service provision in Pampas and Campos grasslands of southern South America. Environ. Res. Lett. 2016, 11, 113002. [Google Scholar] [CrossRef]
- Baeza, S.; Paruelo, J.M. Land use/land cover change (2000–2014) in the Rio de la Plata grasslands: An analysis based on MODIS NDVI time series. Remote Sens. 2020, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Hoekstra, J.M.; Boucher, T.M.; Ricketts, T.H.; Roberts, C. Confronting a biome crisis: Global disparities of habitat loss and protection. Ecol. Lett. 2005, 8, 23–29. [Google Scholar] [CrossRef]
- Suttie, J.M.; Reynolds, S.G.; Batello, C. Grasslands of the World; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2005. [Google Scholar]
- Soriano, A. Rio de la Plata Grasslands. In Natural Grasslands; Coupland, R.T., Ed.; Elsevier: Amsterdam, The Netherlands, 1991; pp. 367–407. [Google Scholar]
- Burkart, R. Las Áreas Protegidas de la Argentina. In La Situación Ambiental Argentina 2005; Brown, A., Martínez Ortíz, U., Acerbiand, M., Corcuera, J., Eds.; Fundación Vida Silvestre: Buenos Aires, Argentina, 2006; pp. 399–403. [Google Scholar]
- Pretelli, M.G.; Isacch, J.P.; Cardoni, D.A. Species-area relationships of specialist versus opportunistic pampas grassland birds depend on the surrounding landscape matrix. Ardeola 2018, 65, 3–23. [Google Scholar] [CrossRef]
- Zalba, S.M.; Villamil, C.B. Woody plant invasion in relictual grasslands. Biol. Invasions 2002, 4, 55–72. [Google Scholar] [CrossRef]
- Kittlein, M.J.; Mora, M.S.; Vassallo, A.I.; de Durana, F.; Ricciardulli, M.G.; Tizón, F.R. Dunas del Sureste Bonaerense. In Identificación de Áreas Valiosas de Pastizal (AVPs) en las Pampas y Campos de Argentina, Uruguay y sur de Brasil; Bilenca, D., Miñarro, F., Eds.; Fundación Vida Silvestre: Buenos Aires, Argentina, 2004; pp. 76–77. [Google Scholar]
- Celsi, C.E.; Mac-Lean, D.; Yezzi, A.; Triches, M. Dunas Costeras de la Pampa Austral. Biodiversidad, Ecología y Conservación Entre el río Quequén Salado y el Balneario Pehuen-Có; Fundación de Historia Natural Félix de Azara: Buenos Aires, Argentina, 2010. [Google Scholar]
- Bilenca, D.; Miñarro, F. Identificación de Áreas Valiosas de Pastizal (AVPs) en las Pampas y Campos de Argentina, Uruguay y sur de Brasil; Fundación Vida Silvestre: Buenos Aires, Argentina, 2004. [Google Scholar]
- Guadagnin, D.L.; Zalba, S.M.; Costa Gorriz, B.; Fonseca, C.R.; Nebbia, A.J.; Cuevas, Y.A.; Emer, C.; Germain, P.; Wendland, E.; Perello, L.F.; et al. Árvores e Arbustos Exóticos Invasores no Bioma Pampa—Questões Ecológicas, Culturais e Socioeconômicas de Umdesafiocrescente. In Campos Sulinos: Conservação e uso Sustentable da Biodiversidade; de Patta Pillar, V., Müller, S.C., de Souza, Z.M., Ávila Jacques, A.V., Eds.; Ministerio do Meio Ambiente: Brasilia, Brasil, 2009; pp. 300–316. [Google Scholar]
- Dadon, J.R. El Impacto del Turismo Sobre los Recursos Naturales Costeros en la Costa Pampeana. In Zona Costera de la Pampa Argentina; Dadon, J.R., Matteucci, S.D., Eds.; Lugar Editorial: Buenos Aires, Argentina, 2002; pp. 101–121. [Google Scholar]
- Cuevas, Y.A.; Zalba, S.M. Control de pinos invasores en el parque provincial Ernesto Tornquist (Buenos Aires): Áreas prioritarias y análisis de costos. BioScriba 2009, 2, 76–89. [Google Scholar]
- Fonseca, C.; Guadagnin, D.L.; Emer, C.; Masciadri, S.; Germain, P.; Zalba, S.M. Invasive alien plants in the Pampas grasslands: A tri-national cooperation challenge. Biol. Invasions 2013, 15, 1751–1763. [Google Scholar] [CrossRef]
- Marchante, H.; Marchante, E.; Freitas, F. Invasion of the Portuguese Dune Ecosystem by the Exotic Species Acacia longifolia (Andrews) Wild: Effects at Community Level. In Plant Invasions: Ecological Threats and Management Solutions; Child, L.E., Brock, J.H., Brundu, G., Prach, K., Pysek, P., Wade, P.M., Williamson, M., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 75–85. [Google Scholar]
- Pauchard, A.; García, A.; Zalba, S.; Sarasola, M.; Zenni, R.; Ziller, S.; Nuñez, M.A. Pine Invasions in South America: Reducing Their Ecological Impacts Through Active Management. In Biological Invasions in Changing Ecosystems: Vectors, Ecological Impacts, Management and Predictions; Canning Clode, J., Ed.; De Gruyter Open Ltd.: Warsaw, Poland, 2016; pp. 318–342. [Google Scholar]
- Alberio, C.; Comparatore, V. Patterns of woody plant invasion in an Argentinean coastal grassland. Acta Oecologica 2014, 54, 65–71. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M.C.; Chen, X.; Corlett, R.T.; Didham, R.K.; Ding, P.; Holt, R.D.; Holyoak, M.; Hu, G.; Hughes, A.C.; Jiang, L.; et al. Habitat fragmentation and biodiversity conservation: Key findings and future challenges. Landsc. Ecol. 2016, 31, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. 2006, 81, 117–142. [Google Scholar] [CrossRef] [PubMed]
- Didham, R.K. Ecological Consequences of Habitat Fragmentation. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological consequences of ecosystem fragmentation: A review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Archer, S.; Boutton, T.W.; Hibbard, K.A. Trees in Grasslands: Biogeochemical Consequences of Woody Plant Expansion. In Global Biogeochemical Cycles and Their Interrelationship with Climate; Schulze, E.A., Harrison, S.P., Heimann, M., Holland, E.A., Lloyd, J., Prentice, I.C., Schimel, D.S., Eds.; Academic Press: London, UK, 2001; pp. 115–130. [Google Scholar]
- Primack, R.; Rozzi, R.; Feinsinger, P.; Dirzo, R.; Massardo, F. Fundamentos de Conservación Biológica: Perspectivas Latinoamericanas; Fondo de Cultura Económico: Mexico City, Mexico, 2001. [Google Scholar]
- Yezzi, A.; Nebbia, A.J.; Zalba, S.M. Fragmentación de pastizales psamófilos por plantaciones de pinos: Efectos sobre la riqueza y la composición vegetal. Ecol. Austral. 2018, 28, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, P.R.; Rodríguez, M.E. Determinantes de la composición florística y efecto de borde en un fragmento de bosque en el Guaviare, Amazonia colombiana. Colomb. For. 2008, 11, 5–18. [Google Scholar] [CrossRef]
- Lord, J.M.; Norton, D.A. Scale and the spatial concept of fragmentation. Conserv. Biol. 1990, 4, 197–202. [Google Scholar] [CrossRef]
- Grace, J.B.; Smith, M.D.; Grace, S.L.; Stohlgren, T.J. Interactions between Fire and Invasive Plants in Temperate Grasslands of North America. In Proceedings of the Invasive Species Workshop: The Role of Fire in the Control and Spread of Invasive Species; Galley, K.E.M., Wilson, T., Eds.; Tall Timbers Research Station: Tallahassee, FL, USA, 2001; pp. 40–65. [Google Scholar]
- Vega, E.; Peters, E. Conceptos Generales Sobre el Disturbio y sus Efectos en los Ecosistemas. In Conservación de Ecosistemas Templados de Montaña en México; Sánchez, O., Vega, E., Peters, E., Monroy-Vilchis, O., Eds.; Secretaría de Medio Ambiente y Recursos Naturales; Instituto Nacional de Ecología; Secretaría de Medio Ambiente; Recursos Naturales y Pesca; Comisión Nacional para el conocimiento y Uso de la Biodiversidad: Mexico City, Mexico, 2003; pp. 137–150. [Google Scholar]
- Wu, G.L.; Zhao, L.P.; Wang, D.; Shi, Z.H. Effects of time-since-fire on vegetation composition and structures in semi-arid perennial grassland on the Loess Plateau, China. Clean Soil Air Water 2014, 42, 98–103. [Google Scholar] [CrossRef]
- Van Langevelde, F.; de Groot, C.; Groen, T.A.; Heitkonig, I.M.A.; Gaigher, I. Effect of patches of woody vegetation on the role of fire in tropical grasslands and savannas. Int. J. Wildland Fire 2014, 23, 410–416. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Végvári, Z.; Hartel, T. Grassland fires in Hungary Experiences of nature conservationists on the effects of fire on biodiversity. Appl. Ecol. Environ. Res. 2014, 12, 267–283. [Google Scholar] [CrossRef]
- Bachinger, L.M.; Brown, L.R.; van Rooyen, M.W. The effects of fire-breaks on plant diversity and species composition in the grasslands of the Loskop Dam Nature Reserve, South Africa. Afr. J. Range Forage Sci. 2016, 33, 21–32. [Google Scholar] [CrossRef]
- Richardson, D.M. Forestry trees as invasive aliens. Conserv. Biol. 1998, 12, 18–26. [Google Scholar] [CrossRef]
- Zalba, S.M.; Cuevas, Y.A.; Boó, R. Aleppo pine invasion after a wildfire in an Argentinean grassland nature reserve. J. Environ. Manag. 2008, 88, 539–546. [Google Scholar] [CrossRef]
- Cuevas, Y.A.; Zalba, S.M. Recovery of native grasslands after removing invasive pines. Restor. Ecol. 2010, 18, 711–719. [Google Scholar] [CrossRef]
- Didham, R.K.; Tylianakis, J.M.; Gemmell, N.J.; Rand, T.A.; Ewers, R.M. Interactive effects of habitat modification and species invasion on native species decline. Trends Ecol. Evol. 2007, 22, 489–496. [Google Scholar] [CrossRef] [PubMed]
- With, K.A. Assessing the risk of invasive spread in fragmented landscapes. Risk Anal. 2004, 24, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Yates, E.D.; Levia, D.F.; Williams, C.L. Recruitment of three non-native invasive plants into a fragmented forest in southern Illinois. For. Ecol. Manag. 2004, 190, 119–130. [Google Scholar] [CrossRef]
- Yezzi, A.; Nebbia, A.J.; Zalba, S.M. Fragmentation and grassland plants: Individual and transgenerational effects. Plant Ecol. 2020, 12, 1275–1291. [Google Scholar] [CrossRef]
- Yezzi, A.; Nebbia, A.J.; Zalba, S.M. Interaction between fire and fragmentation in the successional stages of coastal dune grasslands of the southern Pampas, Argentina. Sci. Rep. 2019, 9, 15109. [Google Scholar] [CrossRef] [Green Version]
- Yezzi, A.; Nebbia, A.J.; Zalba, S.M. Synergistic Interactions of Grassland Fragmentation, Fire and Invasive Woody Plants in the Argentine Pampas. In Advances in Environmental Research Volume 77; Daniels, J.A., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2020; pp. 43–70. [Google Scholar]
- Cabrera, A.L. Fitogeografía de la República Argentina. Boletín De La Soc. Argent. De Botánica 1971, 14, 1–42. [Google Scholar]
- Campo de Ferreras, A.; Capelli de Steffens, A.; Diez, P. El Clima del Suroeste Bonaerense; Departamento de Geografía y Turismo, Universidad Nacional del Sur, Bahía Blanca: Buenos Aires, Argentina, 2004. [Google Scholar]
- Zalba, S.M.; Nebbia, A.J. Neosparton darwinii (Verbenaceae) a restricted endemic species. Is it also endangered? Biodivers. Conserv. 1999, 8, 1585–1593. [Google Scholar] [CrossRef]
- Pieterse, P.J.; Cairns, A.L. The population dynamics of the weed Acacia longifolia (Fabaceae) in the absence and presence of fire. Siud Afrik. Basboutydskrif 1988, 145, 25–27. [Google Scholar]
- Bowers, M.A.; Dooley, J.L. A controlled, hierarchical study of habitat fragmentation: Responses at the individual, patch, and landscape scale. Landsc. Ecol. 1999, 14, 381–389. [Google Scholar] [CrossRef]
- Mueller-Dombois, D.; Ellenberg, E. Aims and Methods of Vegetation Ecology; John Wiley & Sons: Hoboken, NJ, USA, 1974. [Google Scholar]
- Cabrera, A.L. Flora de la Provincia de Buenos Aires; Colección Científica del INTA: Buenos Aires, Argentina, 1965. [Google Scholar]
- Zuloaga, F.O.; Morrone, O.; Belgrano, M. Catálogo de las Plantas Vasculares del Cono Sur; Monographs in Systematic Botany 107; MO Botanical Garden Press: St. Louis, MI, USA, 2008. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Santos, T.; Tellería, J.L. Pérdida y fragmentación del hábitat: Efecto sobre la conservación de las especies. Ecosistemas 2006, 15, 3–12. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Upper Saddle River, NJ, USA, 1996. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat; Universidad Nacional de Córdoba: Córdoba, Argentina, 2008. [Google Scholar]
- Regan, H.M.; Crookston, J.B.; Swab, R.; Franklin, J.; Lawson, W.M. Habitat fragmentation and altered fire regime create trade-offs for an obligate seeding shrub. Ecology 2010, 91, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, D.A.; Armenteres, D.; Bennett, A.F.; Brotons, L.; Clarke, M.F.; Doherty, T.S.; Haslem, A.; Kelly, L.K.; Sato, C.F.; Sitters, H.; et al. How fire interacts with habitat loss and fragmentation. Biol. Rev. 2021, 96, 976–998. [Google Scholar] [CrossRef]
- García, D. Efectos biológicos de la fragmentación de hábitats: Nuevas aproximaciones para resolver un viejo problema. Ecosistemas 2011, 20, 1–10. [Google Scholar]
- Fox, B.J.; Fox, M.D. The Susceptibility of Natural Communities to Invasion. In Ecology of Biological Invasions: An Australian Perspective; Groves, R.H., Burdon, J.J., Eds.; Australian Academy of Science: Canberra, Australia, 1986; pp. 57–66. [Google Scholar]
- Lozon, J.D.; MacIsaac, H.J. Biological invasions: Are they dependent on disturbance? Environ. Rev. 1997, 5, 131–144. [Google Scholar] [CrossRef]
- Didham, R.K.; Tylianakis, J.M.; Hutchison, M.A.; Ewers, R.M.; Gemmell, N.J. Are invasive species the drivers of ecological change? Trends Ecol. Evol. 2005, 20, 471–474. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire and Invasive Species in Mediterranean-Climate Ecosystems of California. In Proceedings of the Invasive Species Workshop: The Role of Fire in the Control and Spread of Invasive Species; Galley, K.E.M., Wilson, T.P., Eds.; Tall Timbers Research Station: Tallahassee, FL, USA, 2001; pp. 81–94. [Google Scholar]
- Amodeo, M.R.; Vázquez, B.; Zalba, S.M. Generalist dispersers promote germination of an alien fleshy-fruited tree invading natural grasslands. PLoS ONE 2017, 12, e0172423. [Google Scholar] [CrossRef]
- Marozas, V. Effect of the coniferous forest–grassland edge on ground vegetation in the mixed European forest zone, Lithuania. Dendrobiology 2014, 71, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Gross, K.L.; Mittelbach, G.G.; Reynolds, H.L. Grassland invasibility and diversity: Responses to nutrients, seed input, and disturbance. Ecology 2005, 86, 476–486. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Turkington, R. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 2005, 86, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Dechoum, M.; Rejmánek, M.; Castellani, T.T.; Zalba, S.M. Limited seed dispersal may explain differences in forest colonization by the Japanese raisin tree (Hovenia dulcis Thunb.), an invasive alien tree in Southern Brazil. Trop. Conserv. Sci. 2015, 8, 610–622. [Google Scholar] [CrossRef] [Green Version]
- Clements, F.E. Nature and structure of the climax. J. Ecol. 1936, 24, 252–284. [Google Scholar] [CrossRef]
- Cook, W.M.; Yao, J.; Foster, B.L.; Holt, R.D.; Patrick, L.B. Secondary succession in an experimentally fragmented landscape: Community patterns across space and time. Ecology 2005, 86, 1267–1279. [Google Scholar] [CrossRef] [Green Version]
- De las Heras, J.; Martínez, J.J.; Herranz, J.M. Impacto ecológico de los incendios forestales. Rev. De Estud. 1991, 29, 105–117. [Google Scholar]
- Rejmánek, M.; Richardson, D.M. What attributes make some plant species more invasive? Ecology 1996, 77, 1655–1661. [Google Scholar] [CrossRef]
- Tapias, R.; Gil, L.; Fuentes-Utrilla, P.; Pardos, J.A. Canopy seed banks in Mediterranean pines of south-eastern Spain: A comparison between Pinus halepensis Mill., P. pinaster Ait., P. nigra Arn. and P. pinea L. J. Ecol. 2001, 89, 629–638. [Google Scholar] [CrossRef]
- Mandle, L.; Bufford, J.; Schmidt, I.; Daehler, C. Woody exotic plant invasions and fire: Reciprocal impacts and consequences for native ecosystems. Biol. Invasions 2011, 13, 1815–1827. [Google Scholar] [CrossRef]
- García, R.A.; Franzese, J.; Policelli, N.; Sasal, Y.; Zenni, R.D.; Nuñez, M.A.; Taylor, K.; Pauchard, A. Non-Native Pines are Homogenizing the Ecosystems of South America. In From Biocultural Homogenization to Biocultural Conservation. Ecology and Ethics; Rozzi, R., May, R.H., Chapin, F.S., Massardo, F., Gavin, M.C., Klaver, I.J., Pauchard, A., Nuñez, M.A., Simberloff, D., Eds.; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Chapman and Hall: London, UK, 1958. [Google Scholar]
- Tilman, D. Community invasibility, recruitment limitation and grassland biodiversity. Ecology 1997, 78, 81–92. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | F | p |
|---|---|---|
| Context (C) | 20.28 | <0.0001 ** |
| Area (A) | 2.55 | <0.0001 ** |
| Time (T) | 5.73 | <0.0001 ** |
| C × A | 2.15 | 0.0006 ** |
| C × T | 3.65 | <0.0001 ** |
| A × T | 1.11 | 0.2551 NS |
| C × A × T | 0.79 | 0.9207 NS |
| Fragment Size (ha) | Before Fire (t0) | After Fire (t3) | ||
|---|---|---|---|---|
| No. | Relative Frequency | No. | Relative Frequency | |
| <0.05 | 3 | 0.20 | 3 | 0.23 |
| 0.1 | 3 | 0.20 | 0 | 0.00 |
| 0.5 | 3 | 0.20 | 5 | 0.38 |
| 2 | 3 | 0.20 | 5 | 0.38 |
| >2 | 3 | 0.20 | 0 | 0.00 |
| Total | 15 | 1 | 13 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yezzi, A.; Nebbia, A.; Zalba, S. Fragmentation of Coastal Grasslands by Plantations and Spontaneous Spread of Invasive Pines in the Southern Pampa. Diversity 2021, 13, 637. https://doi.org/10.3390/d13120637
Yezzi A, Nebbia A, Zalba S. Fragmentation of Coastal Grasslands by Plantations and Spontaneous Spread of Invasive Pines in the Southern Pampa. Diversity. 2021; 13(12):637. https://doi.org/10.3390/d13120637
Chicago/Turabian StyleYezzi, Alejandra, Ana Nebbia, and Sergio Zalba. 2021. "Fragmentation of Coastal Grasslands by Plantations and Spontaneous Spread of Invasive Pines in the Southern Pampa" Diversity 13, no. 12: 637. https://doi.org/10.3390/d13120637
APA StyleYezzi, A., Nebbia, A., & Zalba, S. (2021). Fragmentation of Coastal Grasslands by Plantations and Spontaneous Spread of Invasive Pines in the Southern Pampa. Diversity, 13(12), 637. https://doi.org/10.3390/d13120637