
Description of Lepiotaceous Fungal Species of the Genera Chlorophyllum, Clarkeinda, Macrolepiota, Pseudolepiota, and Xanthagaricus, from Laos and Thailand




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Morphological Study
2.2. Molecular Study
3. Results
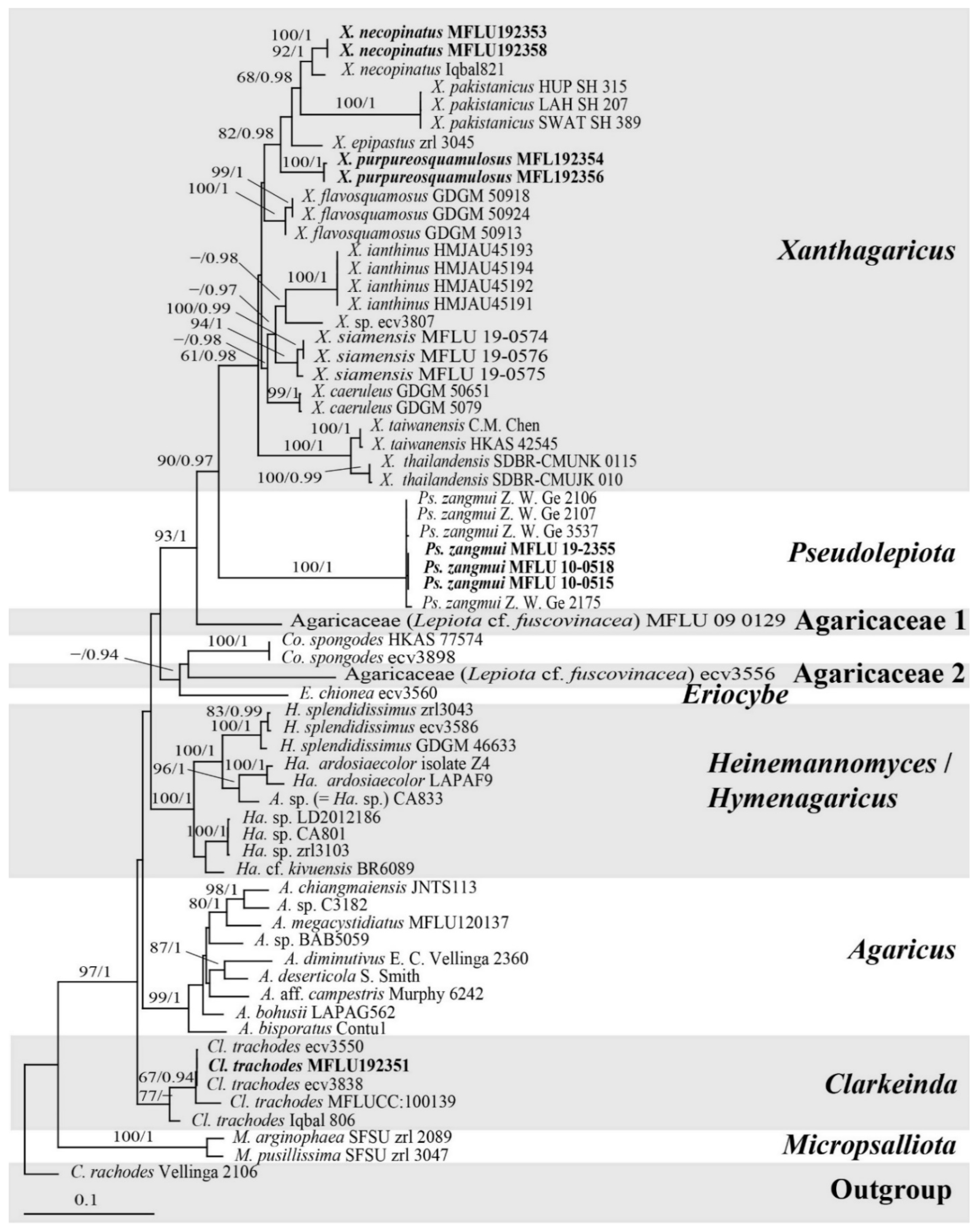
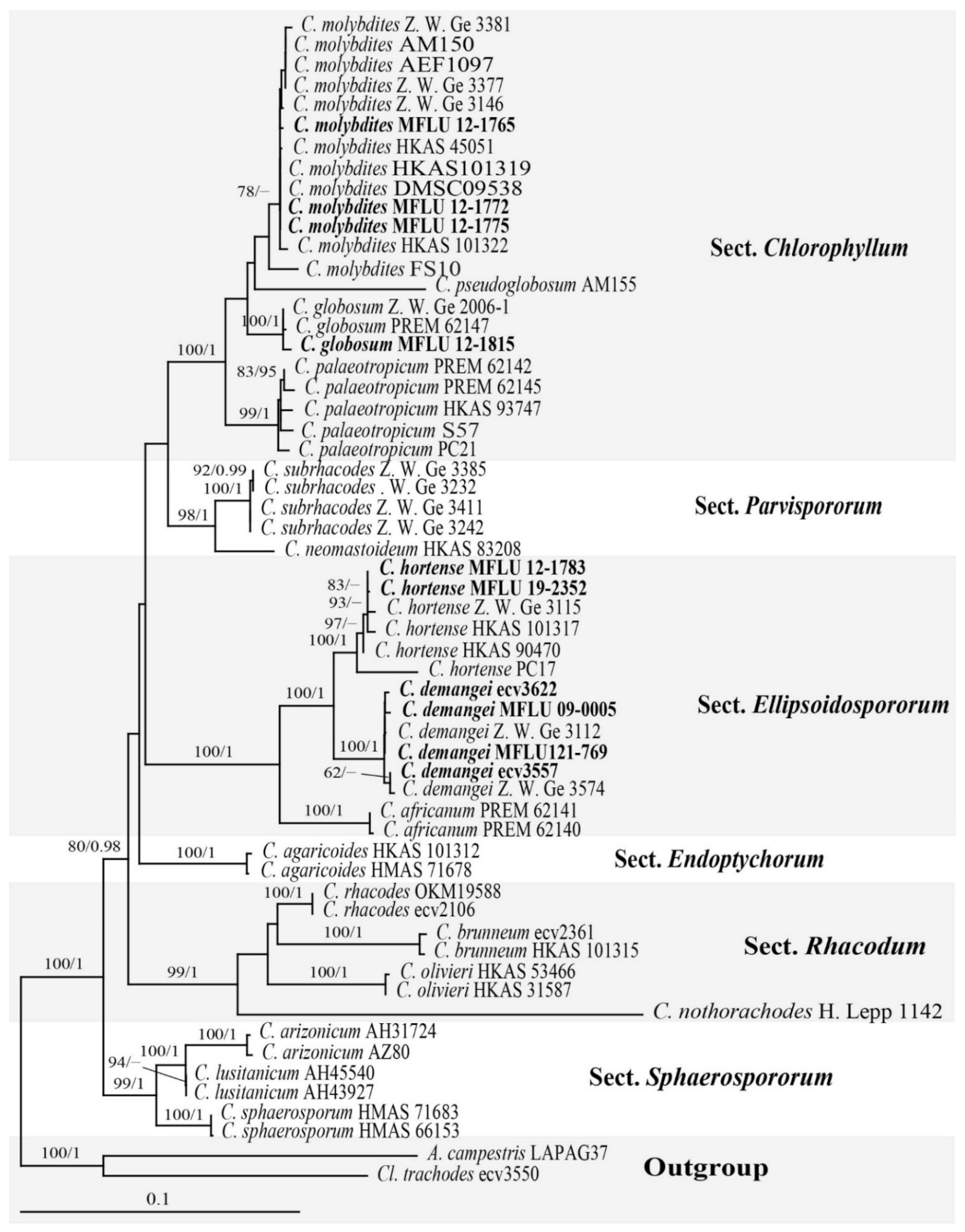
3.1. Result of the Phylogenetic Analyses
3.2. Taxonomy
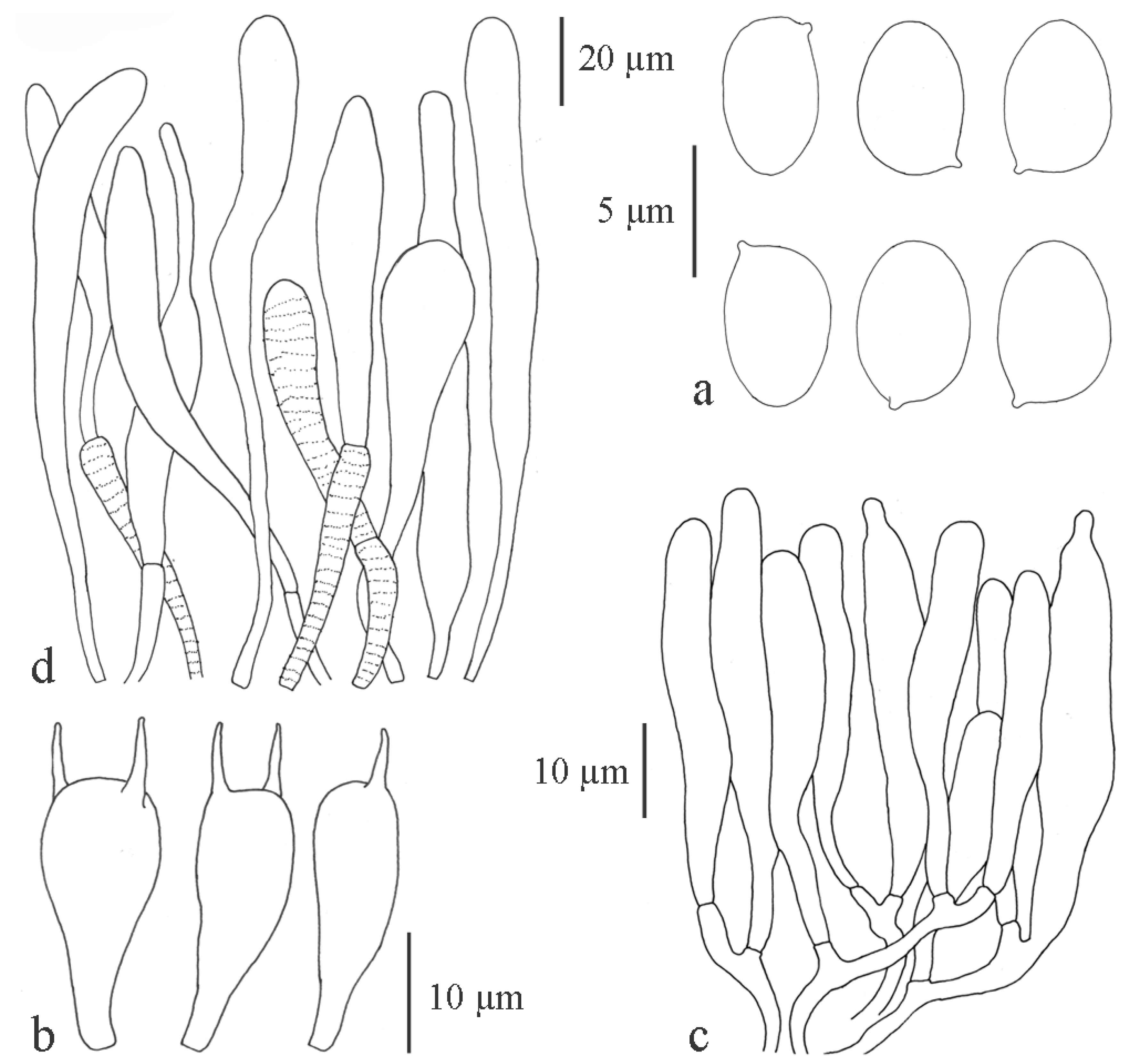
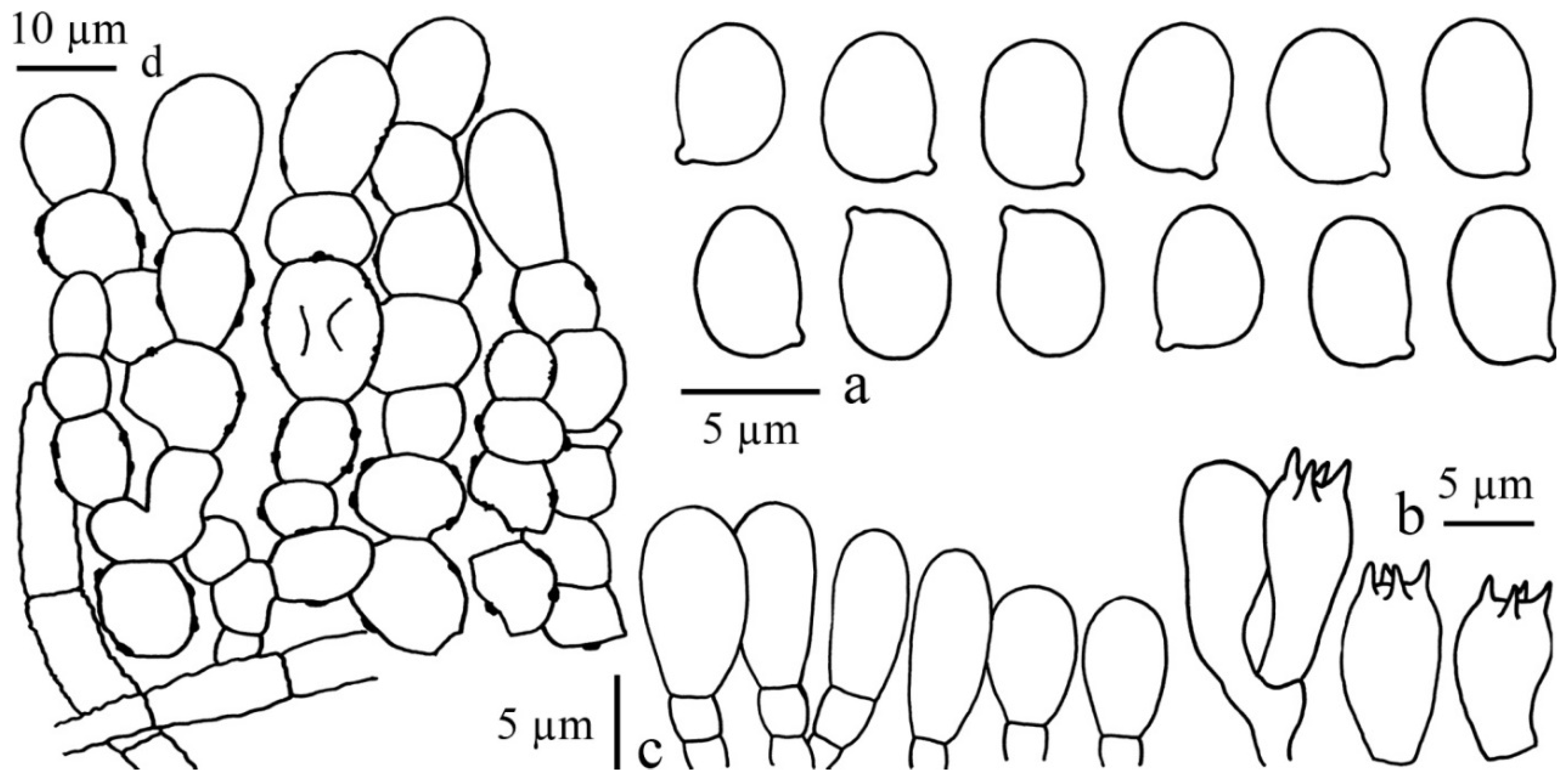
3.2.1. Chlorophyllum Massee
Chlorophyllum demangei (Pat.) Z.W. Ge & Zhu L. Yang, MycoKeys 33: 80 (2018)
Chlorophyllum globosum (Mossebo) Vellinga, Mycotaxon 83: 416 (2002)
Chlorophyllum hortense (Murrill) Vellinga, Mycotaxon 83: 416 (2002)
Chlorophyllum molybdites (G. Mey.) Massee, Kew Bull. 1898: 136 (1898)
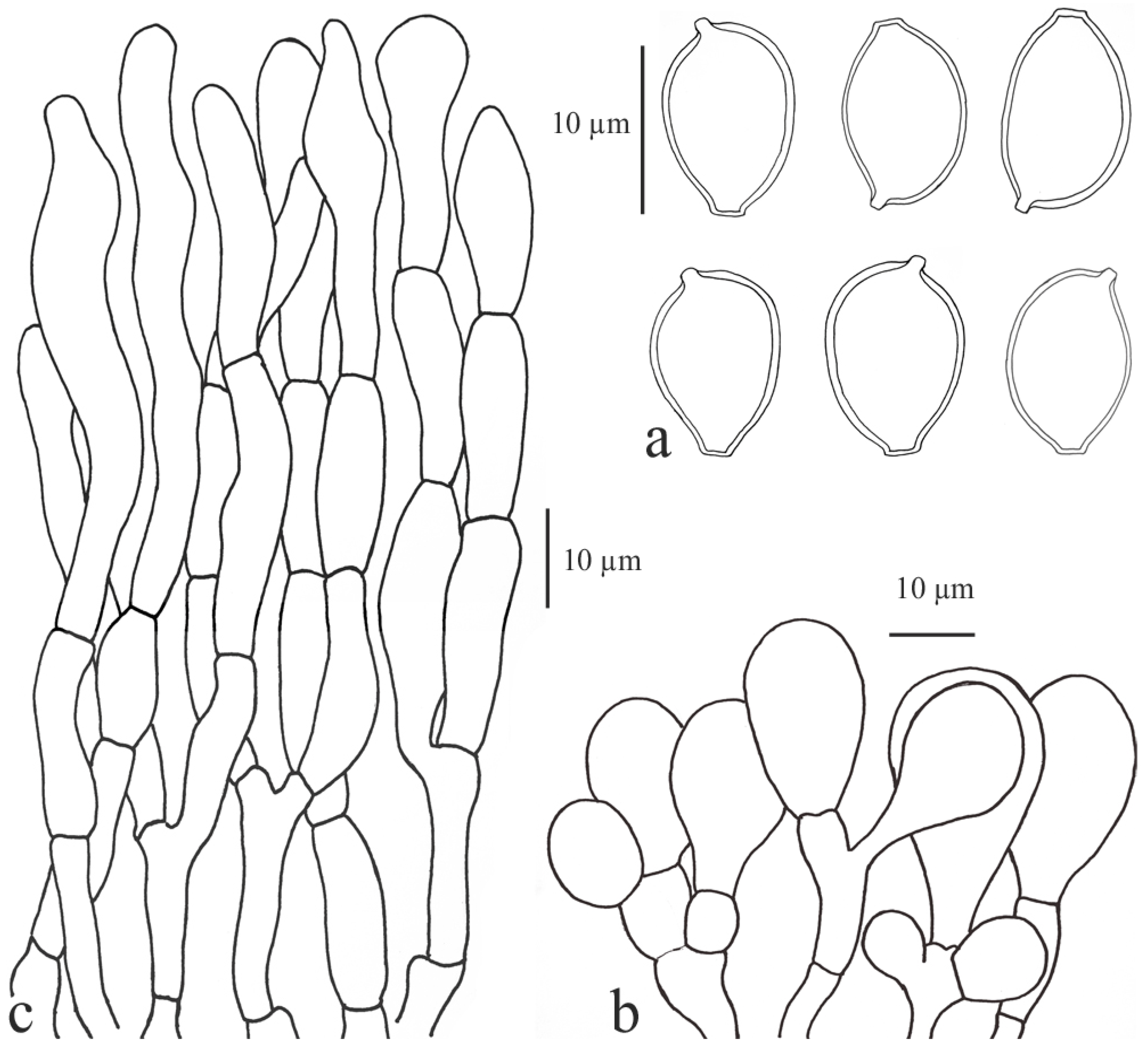
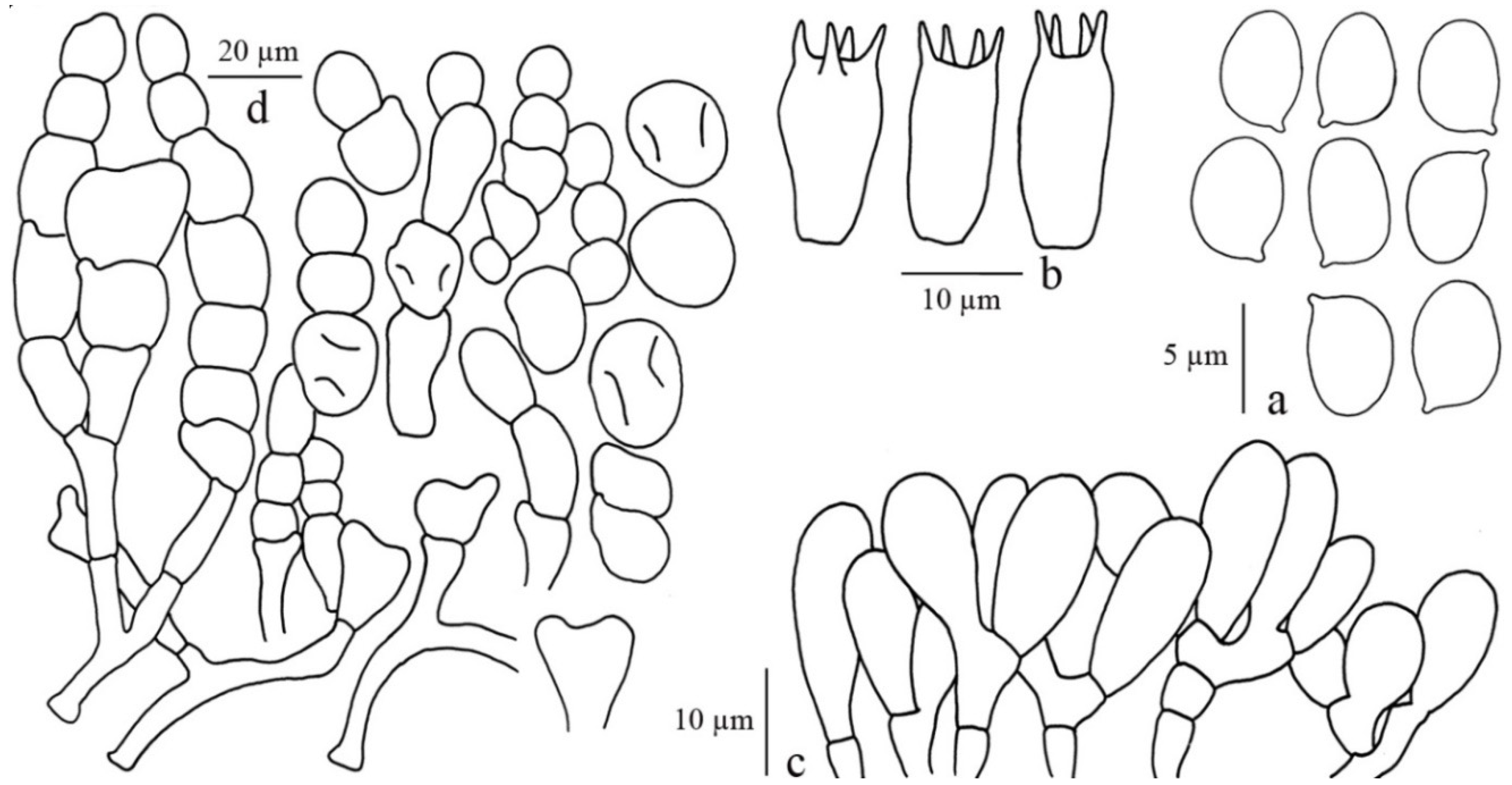
3.2.2. Clarkeinda Kuntze
Clarkeinda trachodes (Berk.) Singer, Lilloa 22: 413 (1951) [1949]
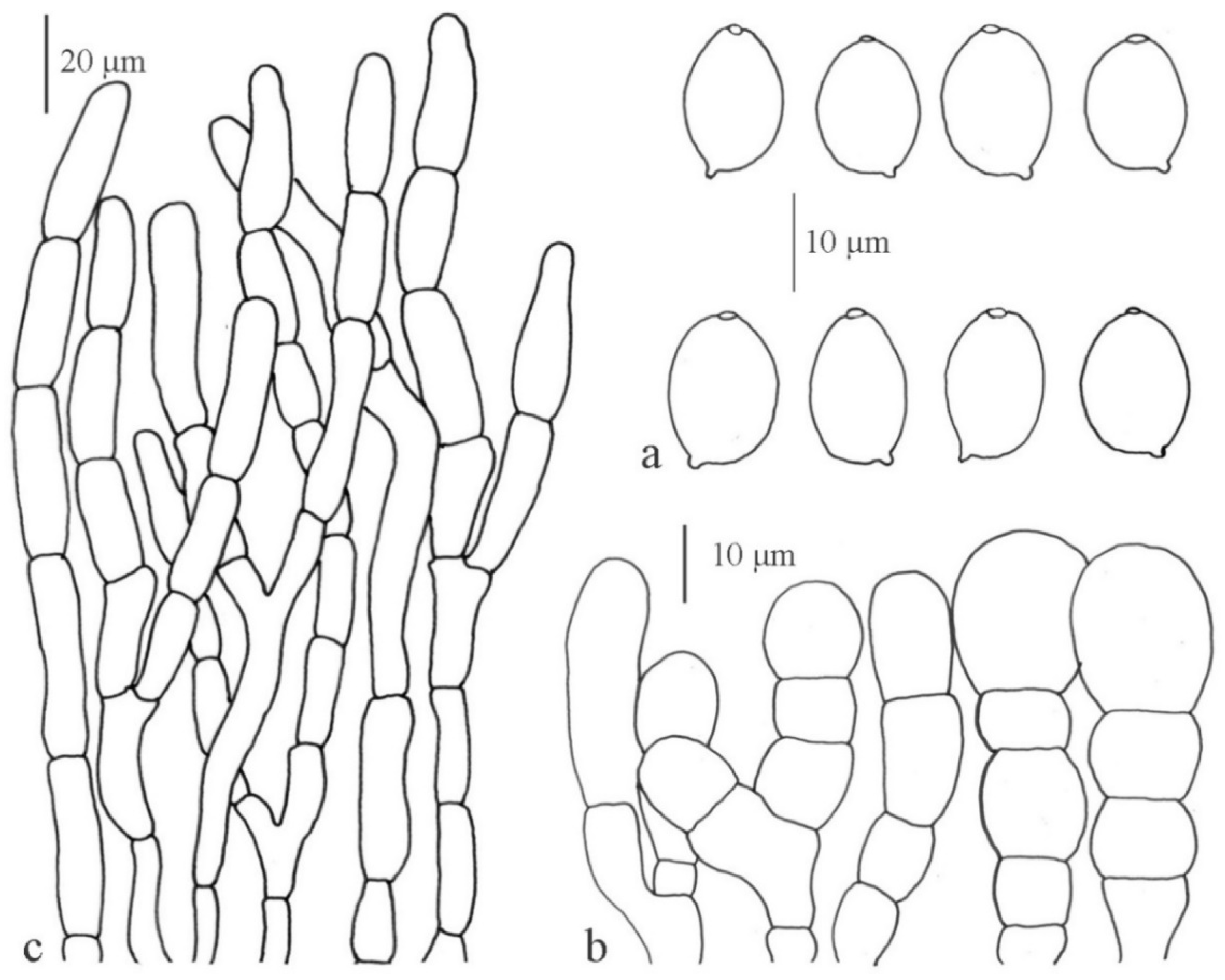
3.2.3. Macrolepiota Singer
Macrolepiota detersa Z. W. Ge, Zhu. L. Yang and Vellinga, Fungal Diversity 45: 83 (2010)
Macrolepiota dolichaula (Berk. and Broome) Pegler and Rayner, Kew Bull. 23: 365 (1969)
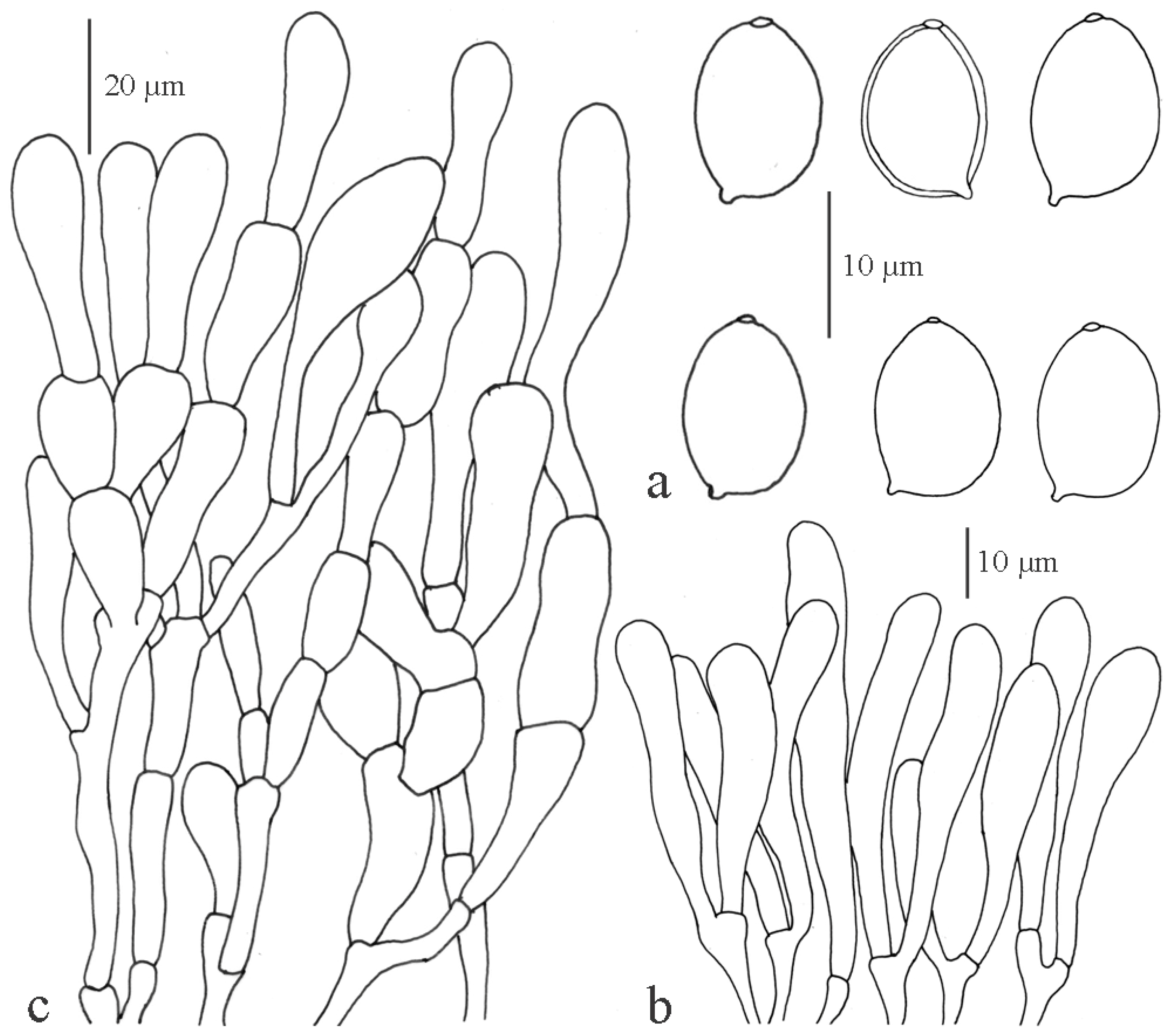
Macrolepiota excelsa Vellinga, Sysouph., Thongkl. and K.D. Hyde sp. nov.
Macrolepiota velosa Vellinga and Zhu L. Yang, Mycotaxon 85: 184 (2003)
3.2.4. Pseudolepiota Z.W. Ge
Pseudolepiota zangmui Z.W. Ge in Ge and Yang, Phytotaxa 312: 252 (2017)
3.2.5. Xanthagaricus (Heinem.) Little Flower, Hosag. and T.K. Abraham
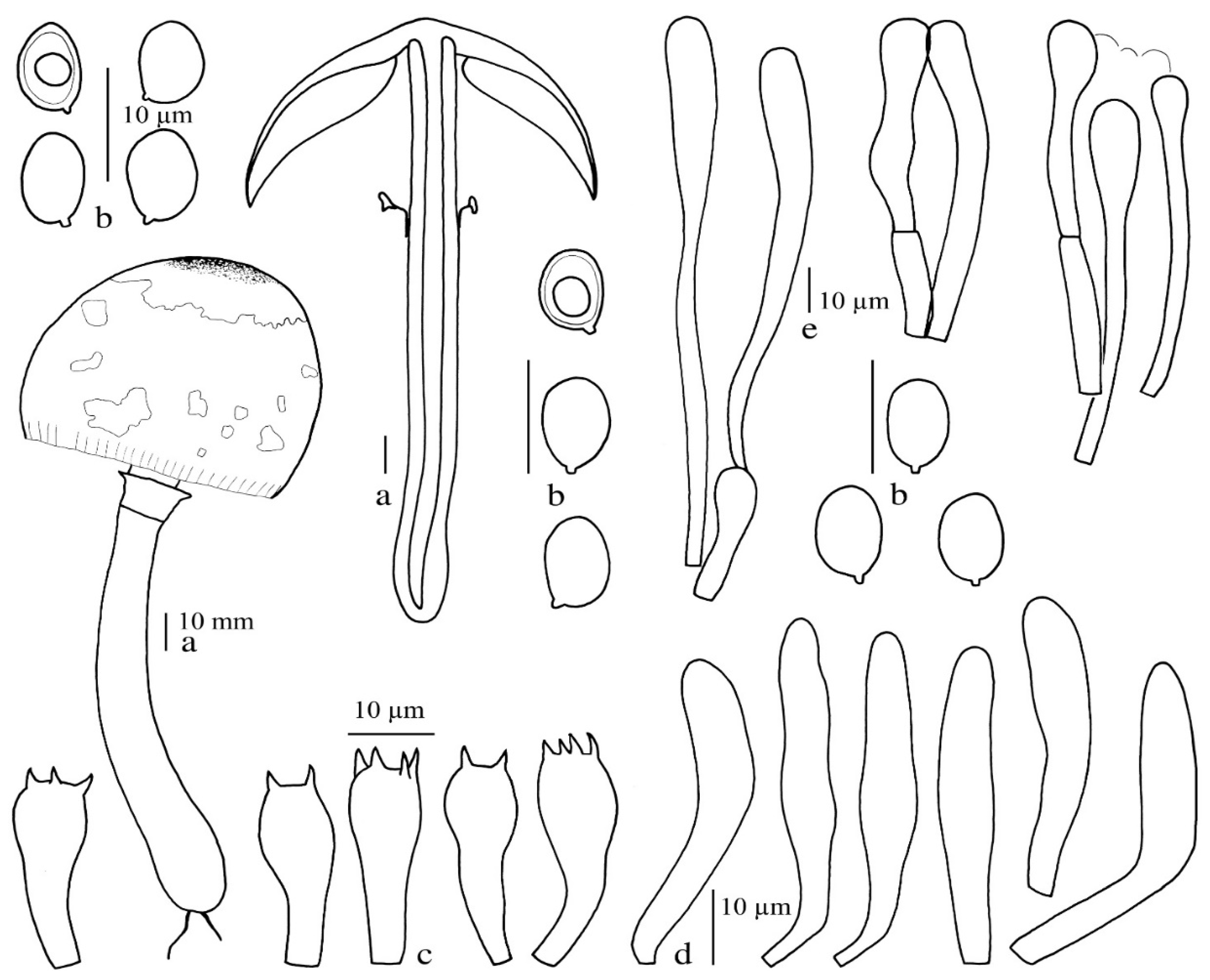
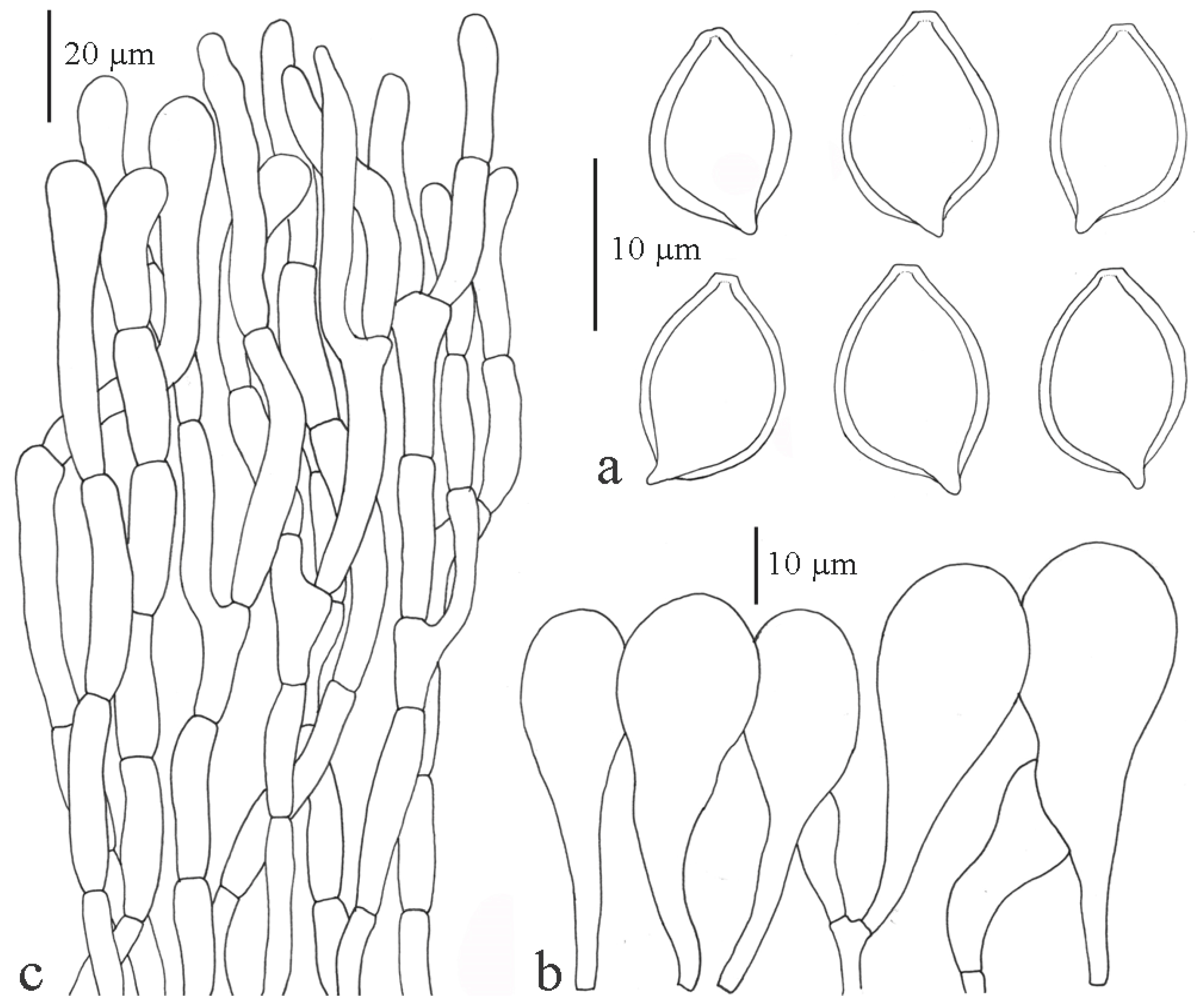
Xanthagaricus purpureosquamulosus Sysouph., Thongkl. and K.D. Hyde sp. nov.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity—Up to 96% of fungi in northern Thailand are novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Vellinga, E.C.; Sysouphanthong, P.; Hyde, K.D. The family Agaricaceae: Phylogenies and two new white-spored genera. Mycologia 2011, 103, 494–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]
- Kalichman, J.; Kirk, P.M.; Matheny, P.B. A compendium of generic names of agarics and Agaricales. Taxon 2020, 69, 425–447. [Google Scholar] [CrossRef]
- Sysouphanthong, P.; Hyde, K.D.; Chukeatirote, E.; Bahkali, A.H.; Vellinga, E.C. Lepiota (Agaricales) in northern Thailand-1 Lepiota section Stenosporae. Mycotaxon 2011, 117, 53–85. [Google Scholar] [CrossRef]
- Sysouphanthong, P.; Hyde, K.D.; Chukeatirote, E.; Bahkali, A.H.; Vellinga, E.C. Lepiota (Agaricales) in northern Thailand–2 Lepiota section Lepiota. Cryptogam. Mycol. 2012, 33, 25–42. [Google Scholar] [CrossRef]
- Sysouphanthong, P.; Hyde, K.D.; Vellinga, E.C.; Chukeatirote, E. Diversity of Lepiota (Agaricales) in northern Thailand. Mycol. Int. J. Fungal. Biol. 2013, 4, 22–28. [Google Scholar]
- Sysouphanthong, P.; Guo, J.; Hyde, K.D.; Xu, J.; Mortimer, P.E. Lepiota thailandica (Agaricaceae), a new species from Thailand. Phytotaxa 2016, 245, 262–270. [Google Scholar] [CrossRef]
- Sysouphanthong, P.; Hyde, K.D.; Chukeatirote, E.; Bahkali, A.H.; Vellinga, E.C. Verrucospora vulgaris (Agaricaceae, Agaricales), a rare tropical species and a new record for Thailand. Chiang Mai J. Sci. 2013, 40, 289–293. [Google Scholar]
- Ge, Z.W.; Jacobs, A.; Vellinga, E.C.; Sysouphanthong, P.; van der Walt, R.; Lavorato, C.; An, Y.F.; Yang, Z.L. A multi-gene phylogeny of Chlorophyllum (Agaricaceae, Basidiomycota): New species, new combination and infrageneric classification. MycoKeys 2018, 32, 65–90. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.K.; Bera, S.; Paloi, S.; Rakshit, S.; Tarafder, E.; Sherpa, A.R.; Acharya, K. Lepiotaceous fungi of West Bengal, India: The genus Chlorophyllum. Phytotaxa 2020, 451, 113–131. [Google Scholar] [CrossRef]
- Loizides, M.; Alvarado, P.; Dimou, D.; Zervakis, G.I.; Thines, M.; Telle, S.; Konstantinou, G.; Gube, M. Multiple evolutionary origins of sequestrate species in the genus Chlorophyllum. Mycologia 2020, 112, 400–422. [Google Scholar] [CrossRef] [PubMed]
- Vellinga, E.C.; de Kok, R.P.J.; Bruns, T.D. Phylogeny and taxonomy of Macrolepiota (Agaricaceae). Mycologia 2003, 95, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Vellinga, E.C. Genera in the family Agaricaceae—Evidence from nrITS and nrLSU sequences. Mycol. Res. 2004, 108, 354–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gube, M. Ontogeny and Phylogeny of Gasteroid Members of Agaricaeae (Basidiomycetes). Ph.D. Thesis, Friedrich-Schiller-Universität, Jena, Germany, 2009. Available online: https://www.db-thueringen.de/receive/dbt_mods_00014723 (accessed on 10 December 2021).
- Gube, M.; Vellinga, E.C.; Alvarado, P.; Polemis, E.; Dimou, D.M.; Zervakis, G.I.; Loizides, M. (2749) Proposal to conserve Chlorophyllum against the additional name Secotium (Agaricaceae). Taxon 2020, 69, 819–820. [Google Scholar] [CrossRef]
- Chandrasrikul, A.; Suwanarit, P.; Sangwanit, U.; Lumyong, S.; Payapanon, A.; Sanoamuang, N.; Pukahuta, C.; Petcharat, V.; Sardsud, U.; Duengkae, K.; et al. Checklist of Mushrooms (Basidiomycetes) in Thailand; Office of Natural Resources and Environmental Policy and Planning: Bangkok, Thailand, 2011; 448p.
- Soytong, K. Mushrooms and Macrofungi in Thailand; Siritham Offset Publishers Ltd.: Ubonratchathani, Thailand, 1994; 151p. (In Thai) [Google Scholar]
- Høiland, K.; Schumacher, T. Agarics, clavarioid and some heterobasidiomycetous fungi from Northern Thailand. Nord. J. Bot. 1982, 2, 265–271. [Google Scholar] [CrossRef]
- Berkeley, M.J. Decades of fungi. Decade XV-XIX. Ceylon fungi. Lond. J. Bot. 1847, 6, 479–514. [Google Scholar]
- Hosen, M.I.; Ge, Z.W. Clarkeinda trachodes (Agaricales, Basidiomycetes), first record from Bangladesh. Mycotaxo 2011, 118, 331–336. [Google Scholar] [CrossRef]
- Leelavathy, K.K.; Zachariah, S.; Sankaran, K.V. Clarkeinda trachodes—An agaric new to India. Mycologia 1981, 73, 204–207. [Google Scholar]
- Verma, R.K.; Tiwari, C.; Parihar, J.; Kumar, S. Diversity of macro-fungi in central India-II: Clarkeinda trachodes. Van Sangyan. 2016, 3, 17–20. [Google Scholar]
- Pegler, D.N. Agaric Flora of Sri Lanka. Kew Bull. Addit. Ser. 1986, 12, 1–519. [Google Scholar]
- Yang, Z.L. Clarkeinda trachodes an agaric new to China. Acta Bot. Yunnanica 1991, 13, 279–282. [Google Scholar]
- Vellinga, E.C.; Noordeloos, M.E. Glossary. In Flora Agaricina Neerlandica 5; Noordeloos, M.E., Kuyper, T.W., Vellinga, E.C., Eds.; A.A. Balkema Publishers: Tokyo, Japan, 2001; pp. 6–11. [Google Scholar]
- Vellinga, E.C. Chlorophyllum and Macrolepiota (Agaricaceae) in Australia. Aust. Syst. Bot. 2003, 16, 361–370. [Google Scholar] [CrossRef]
- Lebel, T.; Syme, A. Sequestrate species of Agaricus and Macrolepiota from Australia: New species and combinations and their position in a calibrated phylogeny. Mycologia 2012, 104, 496–520. [Google Scholar] [CrossRef] [PubMed]
- Junopat, M.; Junopat, W.; Jongjitvimol, T.; Chiengkul, A.; Sanpote, P. Survey of macrofungi in Doi Vieng La Wildlife reserve, Mae Hong Son Province. Naresuan Univ. Sci. J. 2006, 2, 175–181. [Google Scholar]
- Klinhom, U. Checklist of edible mushrooms in Northeast Thailand of survey from 2002–2007 (I). Inf. Mushroom Cultiv. 2008, 13, 8–14. (In Thai) [Google Scholar]
- Sanoamuang, N. Wild Mushrooms of Thailand: Biodiversity and Utilization; Khon Kaen University: Universal Graphic & Trading Ltd.: Bangkok, Thailand, 2010; 424p. (In Thai) [Google Scholar]
- Ge, Z.W.; Yang, Z.L. Pseudolepiota zangmui gen. et sp. nov. (Agaricaceae, Basidiomycota), a new white-spored mushroom from China. Phytotaxa 2017, 312, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Heinemann, P.; Little Flower, S.R. Hymenagaricus (Agaricaceae) de Kerala (Inde) et de Sri Lanka. Bull. Du Jard. Bot. Natl. De Belg. 1984, 54, 151–182. [Google Scholar] [CrossRef]
- Hosen, M.I.; Song, Z.P.; Gates, G.; Karunarathna, S.C.; Chowdhury, M.S.M.; Li, T.H. Two new species of Xanthagaricus and some notes on Heinemannomyces from Asia. MycoKeys 2017, 28, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kumla, J.; Suwannarach, N.; Sringernyuang, K.; Lumyong, S. Xanthagaricus thailandensis sp. nov. (Agaricales, Basidiomycota), from northern Thailand. Phytotaxa 2018, 348, 109–117. [Google Scholar] [CrossRef]
- Liu, S.Y.; Liu, J.K.; Syouphanthong, P.; Hyde, K.D.; Lumyong, S. Xanthagaricus siamensis sp. nov. (Agaricaceae), a new species with dull green lamellae from northern Thailand. Phytotaxa 2020, 437, 14–22. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978; 243p. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML webservers. Syst. Biol. 2008, 75, 758–771. [Google Scholar] [CrossRef]
- Nylander, J.A.A. Modeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, R.D. TreeView: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Li, G.J.; Hyde, K.D.; Zhao, R.L.; Hongsanan, S.; Abdel-Aziz, F.A.; Abdel-Wahab, M.A.; Alvarado, P.; Alves Silva, G.; Ammirati, J.F.; Ariyawansa, H.A.; et al. Fungal diversity notes 253–366: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 78, 1–237. [Google Scholar] [CrossRef]
- Patouillard, N. Champignons nouveaux du Tonkin. Bull. Trimest. De La Soc. Mycol. De Fr. 1907, 23, 69–79. [Google Scholar]
- Mossebo, D.C.; Akoa, A.; Eteme, R.A. Macrolepiota globosa et Nothopanus nsimalenensis, deux nouvelles espèces fongiques du Cameroun. Mycotaxon 2000, 76, 267–278. [Google Scholar]
- Vellinga, E.C. New combinations in Chlorophyllum. Mycotaxon 2002, 83, 415–417. [Google Scholar]
- Akers, B.P.; Sundberg, W.J. Leucoagaricus hortensis: Some synonyms from Florida and taxonomic observations. Mycotaxon 1997, 62, 401–419. [Google Scholar]
- Kumari, B.; Atri, N.S.; Kaur, M. Chlorolepiota indica sp. nov. A new species from India. Mycoscience 2013, 54, 257–259. [Google Scholar] [CrossRef]
- Reid, D.A.; Eicker, A. A comprehensive account of Chlorophyllum molybdites. Bot. Bull. Acad. Sin. 1991, 32, 317–333. [Google Scholar]
- Vellinga, E.C. Een tropisch paddestoelenparadijs. Coolia 1990, 33, 78–79. [Google Scholar]
- Watling, R. A striking addition to the British mycoflora. Mycologist 1991, 5, 23. [Google Scholar] [CrossRef]
- Læssøe, T.; Pedersen, O.S.; Sysouphanthong, P. An Introduction to the Edible, Poisonous and Medicinal Fungi of Northern Laos; Privately Published: Bangkok, Thailand, 2019; 192p. [Google Scholar]
- Bijeesh, C.; Vrinda, K.B.; Pradeep, C.K. Mushroom poisoning by Chlorophyllum molybdites in Kerala. J. Mycopathol. Res. 2017, 54, 477–483. [Google Scholar]
- Wang, N.; Zhao, Z.; Gao, J.; Tian, E.; Yu, W.; Li, H.; Zhang, J.; Xie, R.; Zhao, X.; Chen, A. Rapid and Visual Identification of Chlorophyllum molybdites With Loop-Mediated Isothermal Amplification Method. Front. Microbiol. 2021, 12, 638315. [Google Scholar] [CrossRef]
- Ge, W.; Yang, Z.L.; Vellinga, E.C. The genus Macrolepiota (Agaricaceae, Basidiomycota) in China. Fungal Divers. 2010, 45, 81–98. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.J.; Lee, H.; Park, M.S.; Kim, C.; Wisitrassameewong, K.; Lupala, A.; Park, K.H.; Kim, M.J.; Fong, J.J.; Lim, Y.W. Macrolepiota in Korea: New records and a new species. Mycobiology 2019, 47, 310–368. [Google Scholar] [CrossRef] [Green Version]
- Pegler, D.N. A revision of the genus Lepiota from Ceylon. Kew Bull. 1972, 27, 155–202. [Google Scholar] [CrossRef]
- Pegler, D.N. A preliminary Agaric Flora of East Africa; Kew Bull. Additional Series 6: 1–615; Her Majesty’s Stationery Office: London, UK, 1977.
- Petch, T. Revision of Ceylon Fungi. Annals royal botanic gardens. Peradeniya 1907, 4, 21–68. [Google Scholar]
- Petch, T.; Bisby, G.R. The fungi of Ceylon. Perad. Manua 1950, 6, 1–111. [Google Scholar]
- Natarajan, K.; Manjula, P. South Indian Agaricales. Indian J. Bot. 1981, 4, 50–59. [Google Scholar]
- Yang, Z.L. Type studies on Agarics described by N. Patouillard (and his co-authors) from Vietnam. Mycotaxon 2000, 75, 431–476. [Google Scholar]
- Chiu, W.F. The Amanitaceae of Yunnan. Sci. Rep. Natl. Tsing Hua Univ. Ser. B Biol. Psychol. Sci. 1948, 3, 165–178. [Google Scholar]
- Sysouphanthong, P.; Bouamanivong, S.; Salichan, T. Some Mushrooms in Houayyang Preserves; Biotechnology and Ecology Institute: Vientiane, Laos, 2018; 87p. (In Lao) [Google Scholar]
- Aberdeen, J.E.C. Notes on Lepiota in the Kew Herbarium. Kew Bull. 1962, 16, 129–137. [Google Scholar] [CrossRef]
- Grgurinovic, C.A. Larger Fungi of South Australia; Botanic Gardens of Adelaide; State Herbarium; Flora of South Australia Handbooks Committee: Adelaide, Australia, 1997.
- Mbaluto, C.; Runo, S.; Wanyoike, W.; Onyango, C.; Kimani, W.; Jagger, H.; Otieno, D.O. Macrolepiota aberdarense, a new edible mushroom from Kenya. Curr. Res. Environ. Appl. Mycol. J. Fungal. Biol. 2018, 8, 247–253. [Google Scholar] [CrossRef]
- Pegler, D.N.; Rayner, R.W. A contribution to the Agaric flora of Kenya. Kew Bull. 1969, 23, 347–412. [Google Scholar] [CrossRef]
- Redhead, S.A. Bully for Coprinus—A story of manure, minutiae, and molecules. McIlvainea 2001, 14, 5–14. [Google Scholar] [CrossRef]
- Vellinga, E.C.; Yang, Z.L. Volvolepiota and Macrolepiota—Macrolepiota velosa, a new species from China. Mycotaxon 2003, 85, 183–186. [Google Scholar]
- Hosen, M.I.; Song, Z.; Gates, G.; Li, T.H. Xanthagaricus caeruleus, a new species with ink-blue lamellae from southeast China. Mycoscience 2017, 59, 190–194. [Google Scholar] [CrossRef]
- Wang, F.; Qi, L.L.; Zhou, X.; Li, Y. A new species and a new record of Xanthagaricus (Agaricaceae, Agaricales) from China. Phytotaxa 2018, 371, 241–250. [Google Scholar] [CrossRef]
- Berkeley, M.J.; Broome, C. The fungi of Ceylon (Hymenomycetes, from Agaricus to Cantharellus). Bot. J. Linn. Soc. 1871, 11, 494–567. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Afshan, N.S.; Ahmad, H.; Sher, H.; Khalid, A.N. Xanthagaricus pakistanicus sp. nov. (Agaricaceae), the first report of the genus from Pakistan. Turk. J. Bot. 2018, 42, 123–133. [Google Scholar] [CrossRef]





























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Voucher Number | Country/Collection Number | GenBAnk Accession Number | |
|---|---|---|---|---|
| ITS | LSU | |||
| Agaricus aff. campestris | Murphy 6242 | USA | HM488744 | - |
| Agaricus bisporatus | Contu1 | - | AF432882 | - |
| Agaricus bohusii | LAPAG562 | - | KM657928 | KR006613 |
| Agaricus deserticola | M. Smith | USA | HM488747 | - |
| Agaricus diminutivus | Vellinga 2360 | USA | AF482831 | AF482877 |
| Agaricus megacystidiatus | MFLU 12-0137 | Thailand | NR_119953 | - |
| Agaricus sp. | NTS113 | Thailand | JF514531 | - |
| Agaricus sp. | C3182 | Togo | KJ540956 | - |
| Agaricus sp. | BAB5059 | India | KR155104 | - |
| Chlorophyllum rhacodes | Vellinga 2106 | Netherlands | AF482849 | - |
| Clarkeinda trachodes | Iqbal 806 | Bangladesh | - | MG462712 |
| Clarkeinda trachodes | ecv3838 | Thailand | HM488750 | HM488771 |
| Clarkeinda trachodes | ecv3550 | Thailand | HM488751 | KY418837 |
| Clarkeinda trachodes | MFLUCC:100139 | Sri Lanka | KU845621 | - |
| Clarkeinda trachodes | MFLU 19-2351 | Thailand | MN099351 | - |
| Coniolepiota spongodes | ecv3898 | Thailand | HM488755 | - |
| Coniolepiota spongodes | HKAS 77574 | Bangladesh | KC625531 | KC625530 |
| Eriocybe chionea | ecv3560 | Thailand | HM488752 | HM488773 |
| Heinemannomyces splendidissimus | ecv3586 | Thailand | HM488760 | HM488769 |
| Heinemannomyces splendidissimus | zrl3043 | Thailand | JF691559 | - |
| Heinemannomyces splendidissimus | GDGM 46633 | China | MF621038 | MF621039 |
| Hymenagaricus ardosiicolor | LAPAF9 | Togo | JF727840 | - |
| Hymenagaricus ardosiicolor | isolateZ4 | Tanzania | KM360160 | - |
| Hymenagaricus cf. kivuensis | BR6089 | Burundi | KM982454 | - |
| Hymenagaricus sp. | CA833 | Thailand | JF727858 | - |
| Hymenagaricus sp. | CA801 | Thailand | JF727859 | |
| Hymenagaricus sp. | zrl3103 | Thailand | KM982450 | KM982452 |
| Hymenagaricus sp. | LD2012186 | Thailand | KM982451 | KM982453 |
| Micropsalliota arginophaea | SFSU zrl 2089 | China | HM436616 | HM436578 |
| Micropsalliota pusillissima | SFSU zrl 3047 | China | HM436645 | HM436594 |
| Pseudolepiota zangmui | Z. W. Ge 3537 | China | KY768925 | - |
| Pseudolepiota zangmui | Z. W. Ge 2106 | China | KY768927 | - |
| Pseudolepiota zangmui | Z. W. Ge 2107 | China | KY768926 | - |
| Pseudolepiota zangmui | Z. W. Ge 2175 | China | KY768928 | MG742049 |
| Pseudolepiota zangmui | MFLU 10-0515 | Thailand | KX904355 | - |
| Pseudolepiota zangmui | MFLU 10-0518 | Thailand | KX904356 | - |
| Pseudolepiota zangmui | MFLU 19-2355 | Thailand | MN099352 | - |
| Xanthagaricus caeruleus | GDGM 50651 | China | MF039088 | MF039086 |
| Xanthagaricus caeruleus | GDGM 50794 | China | MF039089 | MF039087 |
| Xanthagaricus epipastus | zrl 3045 | Thailand | HM436649 | HM436609 |
| Xanthagaricus flavosquamosus | GDGM 50913 | China | MF351627 | - |
| Xanthagaricus flavosquamosus | GDGM 50924 | China | MF351628 | - |
| Xanthagaricus flavosquamosus | GDGM 50918 | China | MF351629 | MF351631 |
| Xanthagaricus ianthinus | HMJAU45193 | China | MH166760 | - |
| Xanthagaricus ianthinus | HMJAU45192 | China | MH166761 | - |
| Xanthagaricus ianthinus | HMJAU45191 | China | MH166762 | - |
| Xanthagaricus ianthinus | HMJAU45194 | China | MH166764 | - |
| Xanthagaricus necopinatus | Iqbal821 | Bangladesh | MF351626 | MF351630 |
| Xanthagaricus necopinatus | MFLU 19-2353 | Thailand | MN480544 | - |
| Xanthagaricus necopinatus | MFLU 19-2358 | Thailand | MN480545 | - |
| Xanthagaricus pakistanicus | LAH SH 207 | Pakistan | KY621555 | KY621554 |
| Xanthagaricus pakistanicus | HUP SH 315 | Pakistan | KY621556 | - |
| Xanthagaricus pakistanicus | SWAT SH 389 | Pakistan | KY621557 | - |
| Xanthagaricus purpureosquamulosus | MFLU 19-2354 | Thailand | MN099353 | MN097917 |
| Xanthagaricus purpureosquamulosus | MFLU 19-2356 | Thailand | MN099354 | MN097918 |
| Xanthagaricus siamensis | MFLU 19-0575 | Thailand | MN176991 | MN176981 |
| Xanthagaricus siamensis | MFLU 19-0574 | Thailand | MN176992 | MN176982 |
| Xanthagaricus siamensis | MFLU 19-0576 | Thailand | MN176993 | MN176983 |
| Xanthagaricus sp. | ecv3807 | Thailand | HM488761 | HM488770 |
| Xanthagaricus taiwanensis | C.M. Chen 3636 | Taiwan | DQ006271 | DQ006270 |
| Xanthagaricus taiwanensis | HKAS 42545 | Taiwan | DQ490633 | - |
| Xanthagaricus thailandensis | CMUJK010 | Thailand | MG256663 | MG256665 |
| Xanthagaricus thailandensis | CMUJK0115 | Thailand | MG256664 | MG256666 |
| Taxon | Voucher/Collection Number | Country | GenBank Accession Number | ||
|---|---|---|---|---|---|
| ITS | LSU | rpb2 | |||
| Agaricus campestris | LAPAG370 | China | KM657927 | KR006607 | KT951556 |
| Chlorophyllum africanum | PREM 62140 | South Africa | MG741961 | MG742041 | MG742070 |
| Chlorophyllum africanum | PREM 62141 | South Africa | MG741963 | MG742042 | MG742071 |
| Chlorophyllum agaricoides | HKAS 101312 | Russia | MG742003 | MG742020 | MG742050 |
| Chlorophyllum agaricoides | HMAS 71678 | China | MG742004 | MG742021 | MG742051 |
| Chlorophyllum arizonicum | AH31724 | Mexico | KR233490 | KR233499 | - |
| Chlorophyllum arizonicum | Trappe 11481 | USA | HQ020416 | HQ020419 | - |
| Chlorophyllum brunneum | HKAS 101315 | Italy | MG742013 | MG742022 | MG742052 |
| Chlorophyllum brunneum | ecv2361 | USA | AY083206 | AF482886 | HM488804 |
| Chlorophyllum demangei | Z. W. Ge 3112 | China | MG741965 | MG742027 | MG742056 |
| Chlorophyllum demangei | Z. W. Ge 3574 | China | MG741964 | MG742025 | MG742055 |
| Chlorophyllum demangei | ecv3622 | Thailand | HM488765 | - | - |
| Chlorophyllum demangei | MFLU 12-1769 | Thailand | KJ524556 | MN097907 | - |
| Chlorophyllum demangei | MFLU 09-0005 | Thailand | KJ524555 | MN097908 | - |
| Chlorophyllum demangei | ecv3557 | Thailand | MN582745 | - | - |
| Chlorophyllum globosum | Z. W. Ge 2006-1 | China | MG741995 | MG742023 | - |
| Chlorophyllum globosum | PREM 62147 | South Africa | MG742002 | MG742024 | MG742053 |
| Chlorophyllum globosum | MFLU 12-1815 | Thailand | KJ524553 | MN097909 | - |
| Chlorophyllum hortense | HKAS 101317 | China | MG741967 | MG742026 | MG742054 |
| Chlorophyllum hortense | Z. W. Ge 3115 | China | MG741968 | MG742028 | MG742057 |
| Chlorophyllum hortense | HKAS 90470 | China | MG741971 | MG742029 | MG742058 |
| Chlorophyllum hortense | MFLU 12-1783 | Thailand | KJ524554 | MN097910 | - |
| Chlorophyllum hortense | MFLU 19-2352 | Thailand | MN099355 | MN097916 | MN816433 |
| Chlorophyllum hortense | PC17 | Philippines | MN099356 | MN097915 | - |
| Chlorophyllum lusitanicum | AH45540 | Spain | KR233482 | KR233491 | - |
| Chlorophyllum lusitanicum | AH43927 | Spain | KR233483 | KR233492 | - |
| Chlorophyllum molybdites | HKAS 45051 | China | MG741985 | MG742030 | MG742059 |
| Chlorophyllum molybdites | Z. W. Ge 3381 | USA | MG741993 | MG742034 | MG742063 |
| Chlorophyllum molybdites | Z. W. Ge 3146 | China | MG741987 | MG742031 | MG742060 |
| Chlorophyllum molybdites | HKAS 101322 | Italy | MG741988 | MG742032 | MG742061 |
| Chlorophyllum molybdites | Z. W. Ge 3377 | USA | MG741992 | MG742033 | MG742062 |
| Chlorophyllum molybdites | MFLU 12-1772 | Thailand | KJ524557 | MN097911 | - |
| Chlorophyllum molybdites | MFLU 12-1765 | Thailand | KJ524559 | MN097912 | - |
| Chlorophyllum molybdites | MFLU 12-1775 | Thailand | KJ524558 | MN097913 | - |
| Chlorophyllum molybdites | S57 | Sudan | MK541941 | - | - |
| Chlorophyllum molybdites | J. States AEF1097 | USA | AF482836 | - | - |
| Chlorophyllum molybdites | DMSC09538 | Thailand | KP229775 | - | - |
| Chlorophyllum molybdites | AM150 (CUH) | India | KM190077 | - | - |
| Chlorophyllum molybdites | HKAS 101319 | Thailand | MG741994 | - | - |
| Chlorophyllum molybdites | FS10 | India | MK855510 | - | - |
| Chlorophyllum neomastoideum | HKAS 83208 | China | MG741976 | MG742035 | MG742064 |
| Chlorophyllum nothorachodes | H. Lepp 1142 | Australia | AF482855 | - | - |
| Chlorophyllum olivieri | HKAS 31587 | Germany | MG742016 | MG742036 | MG742065 |
| Chlorophyllum olivieri | HKAS 53466 | Germany | MG742017 | MG742037 | MG742066 |
| Chlorophyllum palaeotropicum | PREM 62142 | South Africa | MG741978 | MG742038 | MG742067 |
| Chlorophyllum palaeotropicum | PREM 62145 | South Africa | MG741982 | MG742039 | MG742068 |
| Chlorophyllum palaeotropicum | HKAS 93747 | Benin | MG741983 | MG742040 | MG742069 |
| Chlorophyllum palaeotropicum | pc21 | Philippines | MN099357 | MN097914 | - |
| Chlorophyllum pseudoglobosum | AM155 | India | KP642506 | KR080484 | - |
| Chlorophyllum rhacodes | ecv2106 | Netherlands | AF482849 | AY176345 | - |
| Chlorophyllum rhacodes | OKM19588 | USA | U85312 | U85277 | HM488803 |
| Chlorophyllum sphaerosporum | HMAS 66153 | China | MG742011 | MG742043 | MG742072 |
| Chlorophyllum sphaerosporum | HMAS 71683 | China | MG742012 | MG742044 | MG742073 |
| Chlorophyllum subrhacodes | Z. W. Ge 3411 | USA | MG741975 | MG742045 | MG742074 |
| Chlorophyllum subrhacodes | Z. W. Ge 3232 | USA | MG741973 | MG742046 | MG742075 |
| Chlorophyllum subrhacodes | Z. W. Ge 3385 | USA | MG741972 | MG742048 | MG742077 |
| Chlorophyllum subrhacodes | Z. W. Ge 3242 | USA | MG741974 | MG742047 | MG742076 |
| Clarkeinda trachodes | ecv3550 | Thailand | HM488751 | KY418837 | HM488802 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sysouphanthong, P.; Thongklang, N.; Liu, J.-K.; Vellinga, E.C. Description of Lepiotaceous Fungal Species of the Genera Chlorophyllum, Clarkeinda, Macrolepiota, Pseudolepiota, and Xanthagaricus, from Laos and Thailand. Diversity 2021, 13, 666. https://doi.org/10.3390/d13120666
Sysouphanthong P, Thongklang N, Liu J-K, Vellinga EC. Description of Lepiotaceous Fungal Species of the Genera Chlorophyllum, Clarkeinda, Macrolepiota, Pseudolepiota, and Xanthagaricus, from Laos and Thailand. Diversity. 2021; 13(12):666. https://doi.org/10.3390/d13120666
Chicago/Turabian StyleSysouphanthong, Phongeun, Naritsada Thongklang, Jian-Kui Liu, and Else C. Vellinga. 2021. "Description of Lepiotaceous Fungal Species of the Genera Chlorophyllum, Clarkeinda, Macrolepiota, Pseudolepiota, and Xanthagaricus, from Laos and Thailand" Diversity 13, no. 12: 666. https://doi.org/10.3390/d13120666
APA StyleSysouphanthong, P., Thongklang, N., Liu, J. -K., & Vellinga, E. C. (2021). Description of Lepiotaceous Fungal Species of the Genera Chlorophyllum, Clarkeinda, Macrolepiota, Pseudolepiota, and Xanthagaricus, from Laos and Thailand. Diversity, 13(12), 666. https://doi.org/10.3390/d13120666