
The Influence of Climate Change on Three Dominant Alpine Species under Different Scenarios on the Qinghai–Tibetan Plateau


Abstract
:1. Introduction
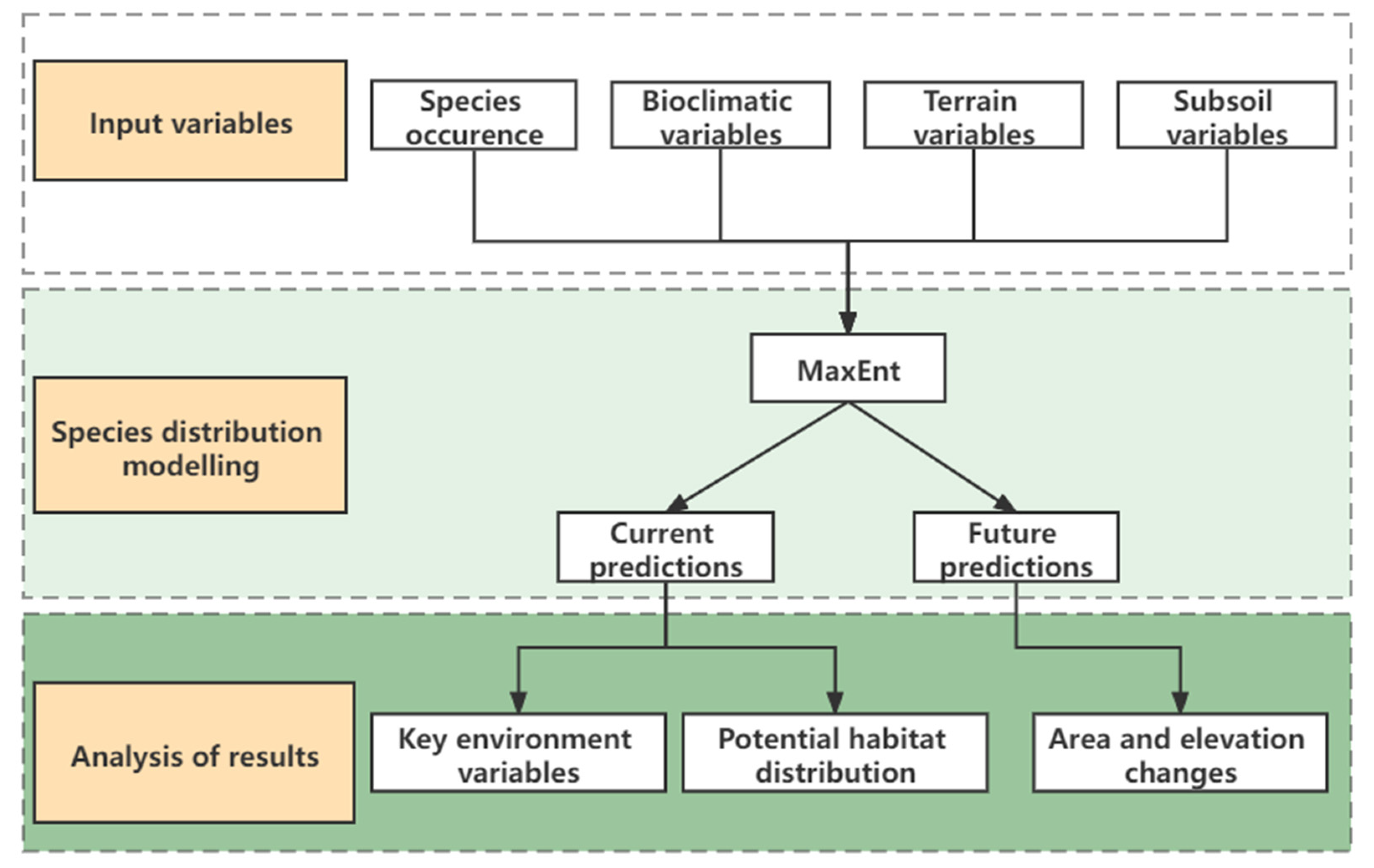
2. Materials and Methods
2.1. Study Area
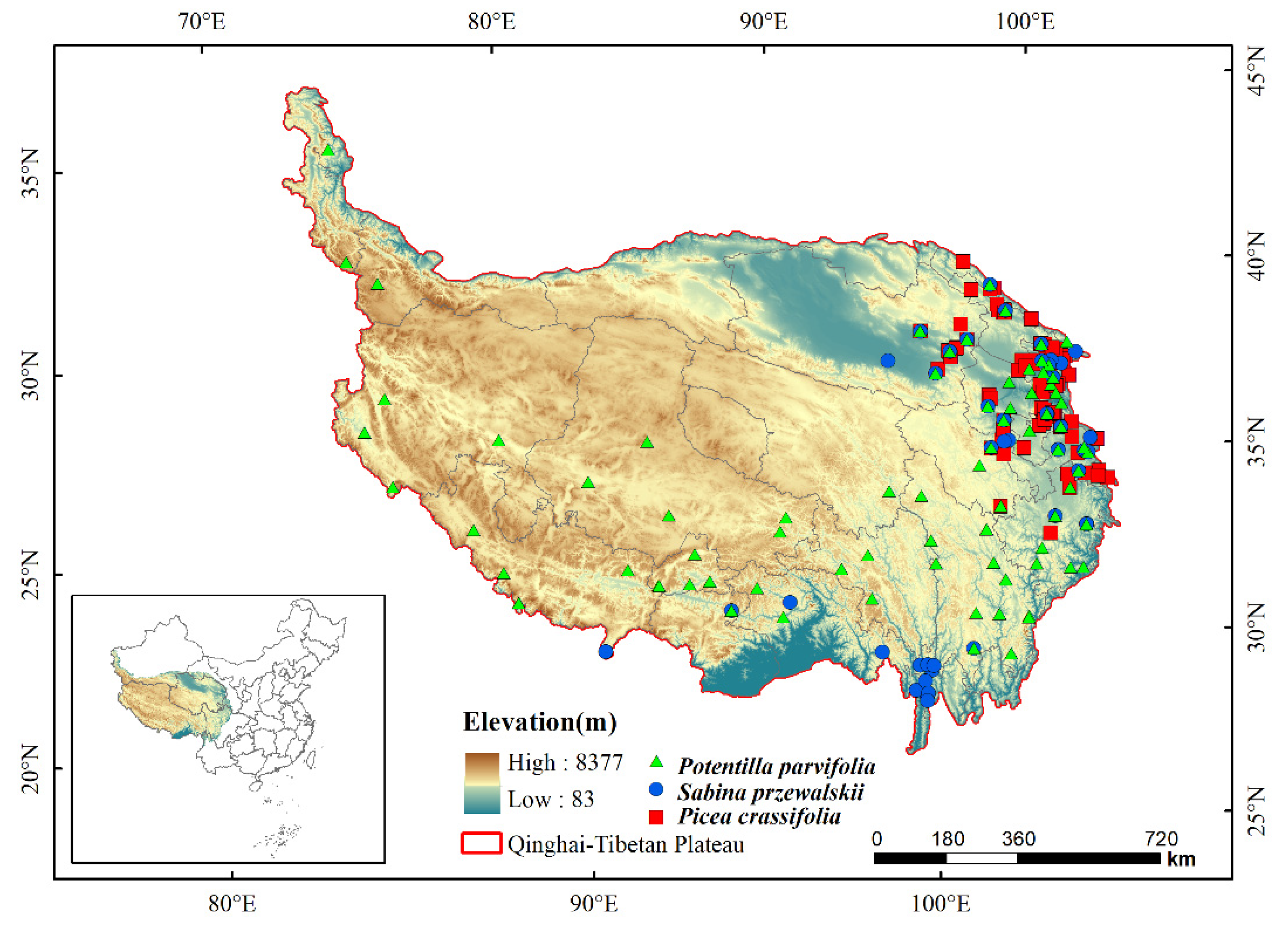
2.2. Occurrence Data
2.3. Environment Variables
2.3.1. Climate Data
2.3.2. Topographic Data
2.3.3. Soil Property Data
2.3.4. Processing and Selection of Environment Variables
2.4. Distribution Modeling
2.4.1. Accuracy Assessment
2.4.2. The Area and Elevation Changes of the Habitat Suitability
3. Results
3.1. Model Assessment and Key Environmental Variables
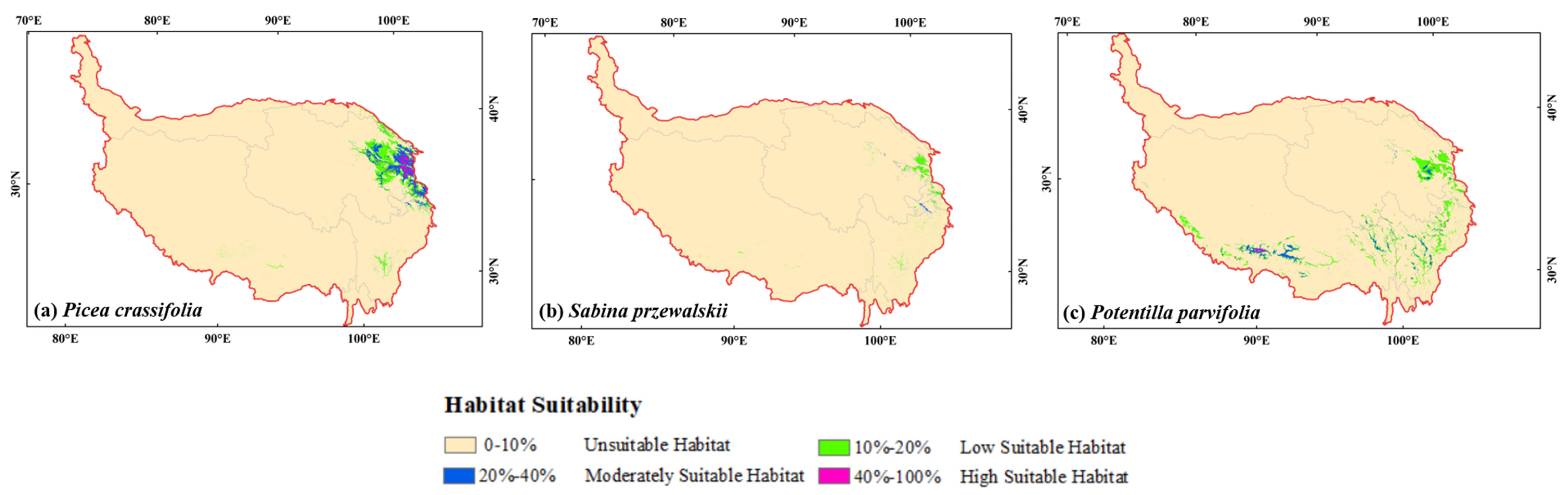
3.2. Potential Distribution of Three Species at Current Climate Scenarios
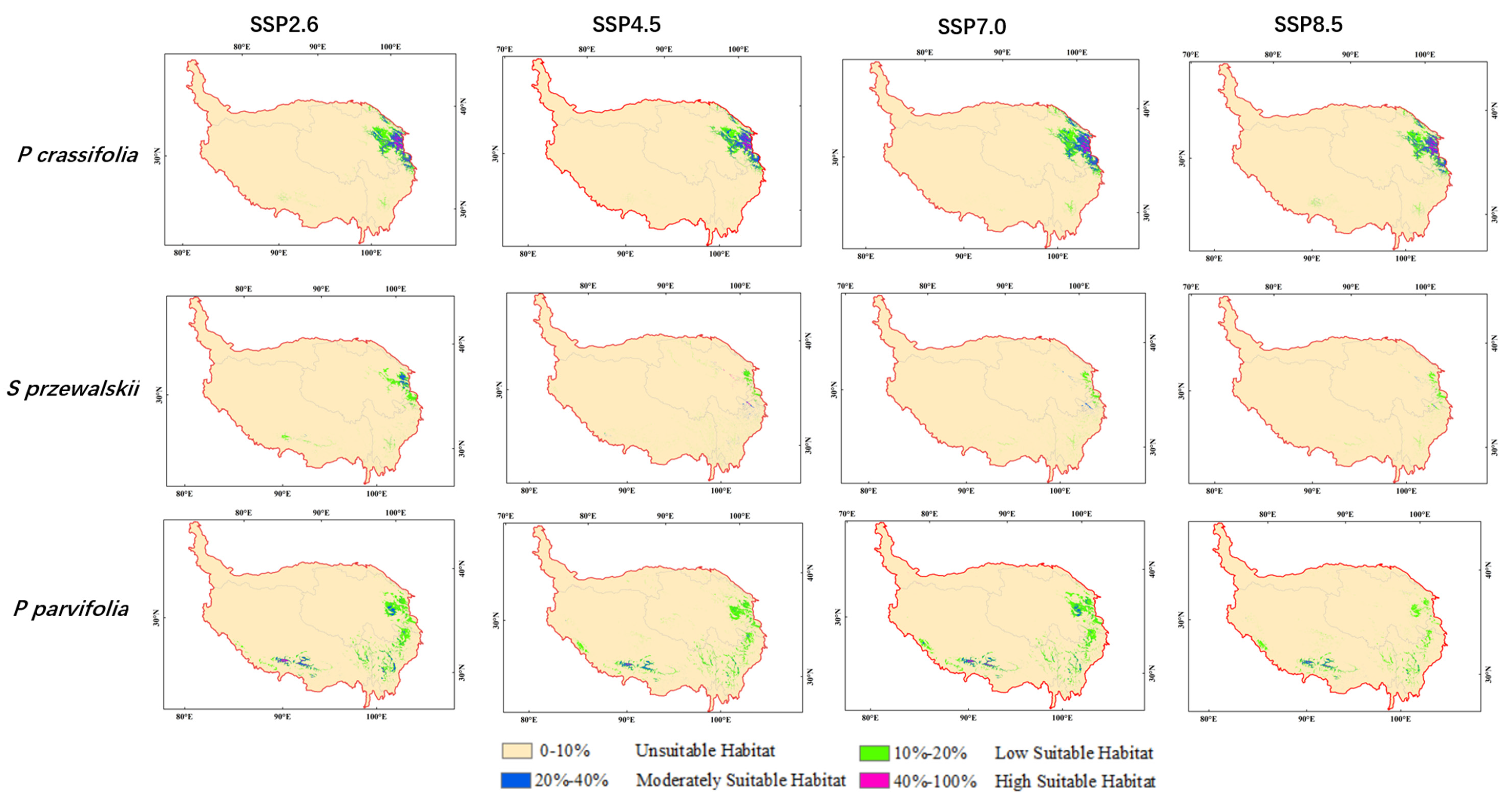
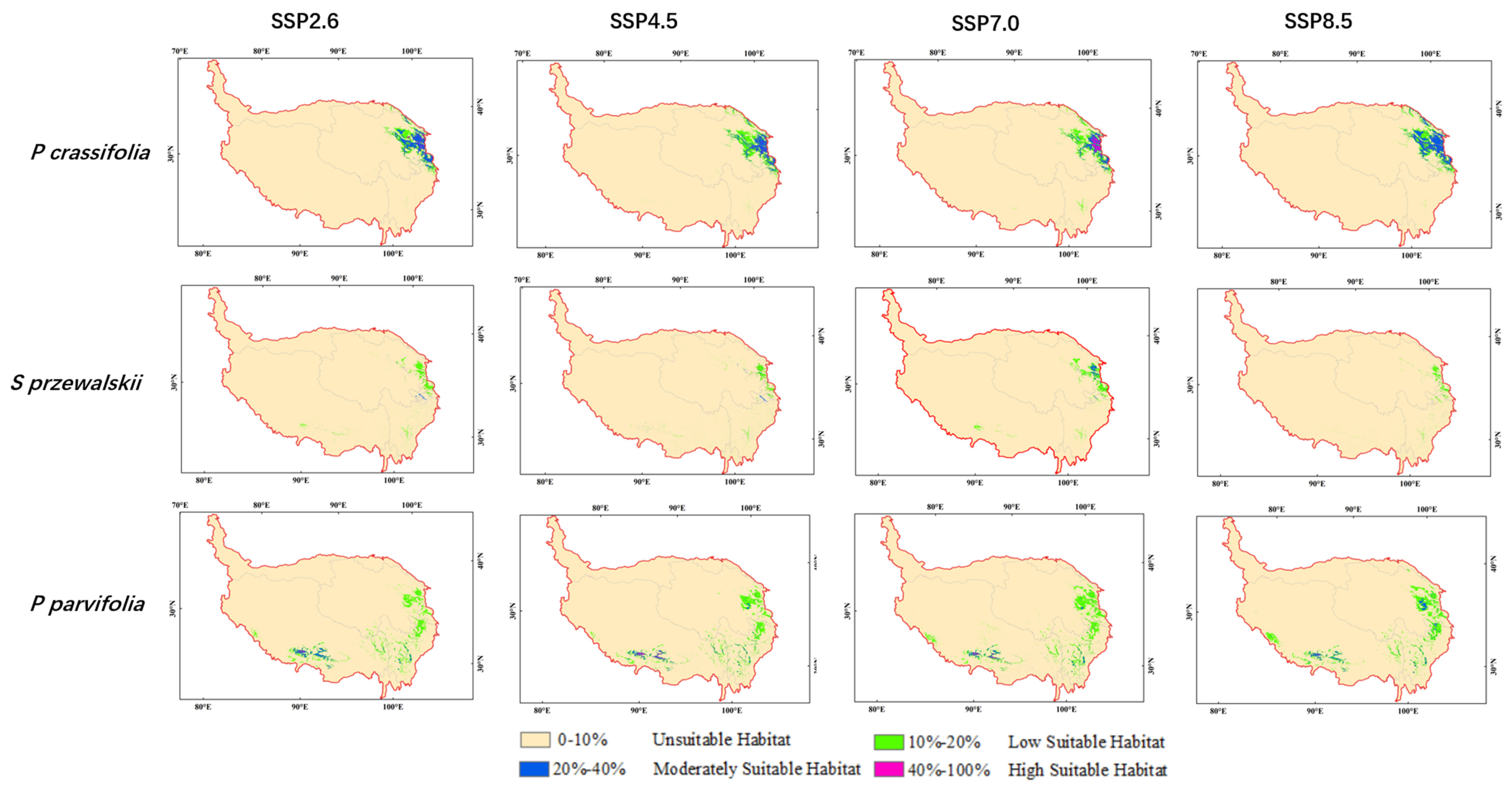
3.3. Potential Distribution of Three Species under Future Climate Scenarios
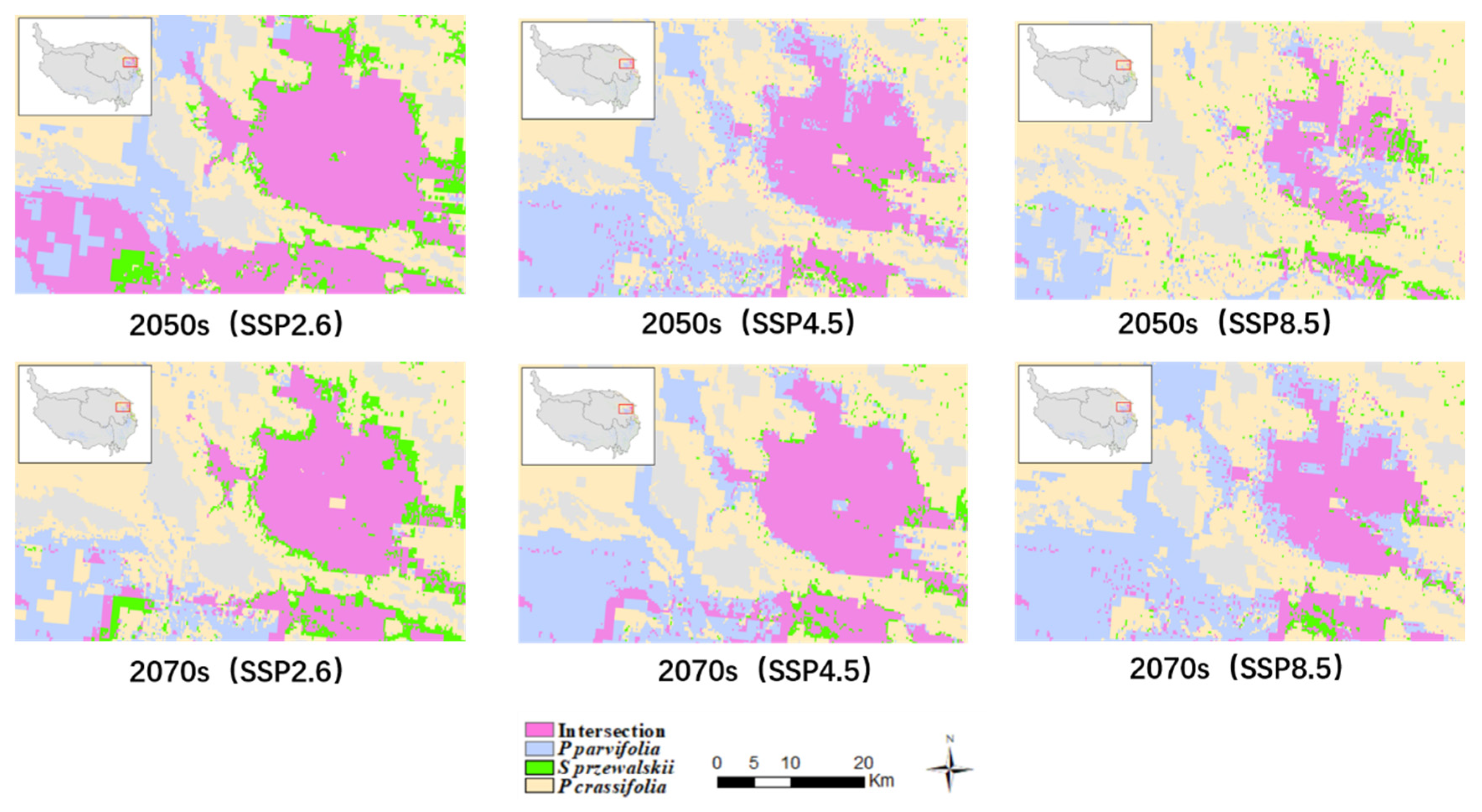
3.4. The Changes of the Intersection Distribution of Three Species
4. Discussion
4.1. Influence of Environmental Variables on the Potential Distribution of Three Species
4.2. Average Elevation Changes of Potential Suitable Habitat for Three Model Species
4.3. Influence of Other Factors on the Potential Distribution of Three Species
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Araújo, M.B.; Rahbek, C. How Does Climate Change Affect Biodiversity? Science 2006, 313, 1396–1397. [Google Scholar] [CrossRef]
- Beaumont, L.J.; Pitm, A.J.; Poulsen, M.; Hughes, L. Where will species go? Incorporating new advances in climate modelling into projections of species distributions. Glob. Chang. Biol. 2007, 13, 1368–1385. [Google Scholar] [CrossRef]
- Lenoir, J.; Gegout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.G.; Wu, T.W.; Zhang, J.; Yao, J.C.; Fang, Y.J. Comparison of CMIP6 and CMIP5 simulations of precipitation in China and the East Asian summer monsoon. Int. J. Climatol. 2020, 40, 6423–6440. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.D.; Chen, B.D. Climatic warming in the Tibetan Plateau during recent decades. Int. J. Climatol. 2000, 20, 1729–1742. [Google Scholar] [CrossRef]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Fox, R.; Thomas, C.D. The distributions of a wide range of taxonomic groups are expanding polewards. Glob. Chang. Biol. 2006, 12, 450–455. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355. [Google Scholar] [CrossRef] [PubMed]
- Naudiyal, N.; Wang, J.; Ning, W.; Gaire, N.P.; Peili, S.; Yanqiang, W.; Jiali, H.; Ning, S. Potential distribution of Abies, Picea, and Juniperus species in the sub-alpine forest of Minjiang headwater region under current and future climate scenarios and its implications on ecosystem services supply. Ecol. Indic. 2021, 121, 107131. [Google Scholar] [CrossRef]
- Xu, Z.; Zhao, C.; Feng, Z.; Peng, H.; Chao, W. The impact of climate change on potential distribution of species in semi-arid region: A case study of Qinghai spruce (Picea crassifolia) in Qilian Mountain, Gansu province, China. In Proceedings of the 2009 IEEE International Geoscience and Remote Sensing Symposium, Cape Town, South Africa, 12–17 July 2009. [Google Scholar]
- Kearney, M.R.; Wintle, B.A.; Porter, W.P. Correlative and mechanistic models of species distribution provide congruent forecasts under climate change. Conserv. Lett. 2010, 3, 203–213. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Bradter, U.; Kunin, W.E.; Altringham, J.D.; Thom, T.J.; Benton, T.G. Identifying appropriate spatial scales of predictors in species distribution models with the random forest algorithm. Methods Ecol. Evol. 2013, 4, 167–174. [Google Scholar] [CrossRef]
- Stockwell, D.; Peters, D. The GARP modelling system: Problems and solutions to automated spatial prediction. Int. J. Geogr. Inf. Sci. 1999, 13, 143–158. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Chen, X.; Xin, X. Short commentary on CMIP6 Scenario Model Intercomparison Project (ScenarioMIP). Progress. Inquisitiones De Mutat. Clim. 2019, 15, 519–525. [Google Scholar] [CrossRef]
- Riahi, K.; van Vuuren, D.P.; Kriegler, E.; Edmonds, J.; O’Neill, B.C.; Fujimori, S.; Bauer, N.; Calvin, K.; Dellink, R.; Fricko, O.; et al. The Shared Socioeconomic Pathways and their energy, land use, and greenhouse gas emissions implications: An overview. Glob. Environ. Chang. Hum. Policy Dimens. 2017, 42, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Milne, R.I.; Cadotte, M.W.; Wu, Z.-Y.; Provan, J.; Zhu, G.-F.; Gao, L.-M.; Li, D.-Z. Protect Third Pole’s fragile ecosystem. Science 2018, 362, 1368. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Jia, K.; Zhao, W.W.; Liu, S.L.; Wei, X.Q.; Wang, B. Spatio-temporal changes of ecological vulnerability across the Qinghai-Tibetan Plateau. Ecol. Indic. 2021, 123, 107274. [Google Scholar] [CrossRef]
- Liu, J.Q.; Sun, Y.S.; Ge, X.J.; Gao, L.M.; Qiu, Y.X. Phylogeographic studies of plants in China: Advances in the past and directions in the future. J. Syst. Evol. 2012, 50, 267–275. [Google Scholar] [CrossRef]
- Qiu, Y.X.; Fu, C.X.; Comes, H.P. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogenetics Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef]
- Zhang, K.L.; Yao, L.J.; Meng, J.S.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Kebede, A.S.; Nicholls, R.; Allan, A.; Arto, I.; Cazcarro, I.; Fernandes, J.A.; Hill, C.T.; Hutton, C.; Kay, S.; Lázár, A.N.; et al. Applying the global RCP-SSP-SPA scenario framework at sub-national scale: A multi-scale and participatory scenario approach. Sci. Total Environ. 2018, 635, 659–672. [Google Scholar] [CrossRef]
- Wu, T.; Lu, Y.; Fang, Y.; Xin, X.; Li, L.; Li, W.; Jie, W.; Zhang, J.; Liu, Y.; Zhang, L.; et al. The Beijing Climate Center Climate System Model (BCC-CSM): The main progress from CMIP5 to CMIP6. Geosci. Model Dev. 2019, 12, 1573–1600. [Google Scholar] [CrossRef] [Green Version]
- Booth, T.H. Why understanding the pioneering and continuing contributions of BIOCLIM to species distribution modelling is important. Austral Ecol. 2018, 43, 852–860. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Li, G.; Qin, W.; Zhang, J.; Lin, G.; Cai, Z.; Gao, H.; Zhang, T. Setting priority conservation areas of wild Tibetan gazelle (Procapra picticaudata) in China’s first national park. Glob. Ecol. Conserv. 2019, 20, e00725. [Google Scholar] [CrossRef]
- Zeng, Y.; Low, B.W.; Yeo, D.C.J. Novel methods to select environmental variables in MaxEnt: A case study using invasive crayfish. Ecol. Model. 2016, 341, 5–13. [Google Scholar] [CrossRef]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Zhu, C.; Qu, Y.; Wang, H. Predicting the potential distribution of an invasive species, Erigeron canadensis L., in China with a maximum entropy model. Glob. Ecol. Conserv. 2020, 21, e00822. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Jose, S.; Nameer, P.O. The expanding distribution of the Indian Peafowl (Pavo cristatus) as an indicator of changing climate in Kerala, southern India: A modelling study using MaxEnt. Ecol. Indic. 2020, 110, 105930. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rodder, D.; Secondi, J. Mapping Species Distributions with MAXENT Using a Geographically Biased Sample of Presence Data: A Performance Assessment of Methods for Correcting Sampling Bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Peng, J.; Gou, X.; Chen, F.; Li, J.; Liu, P.; Zhang, Y.; Fang, K. Difference in tree growth responses to climate at the upper treeline: Qilian Juniper in the Anyemaqen Mountains. J. Integr. Plant Biol. 2008, 50, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Feng, J.; Chen, C.; Chen, J.; Long, T.; Li, J.; Zang, R.; Li, J. Differential Responses to Climate and Land-Use Changes in Threatened Chinese Taxus Species. Forests 2019, 10, 766. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Guo, Y.; Wei, H.; Ran, Q.; Liu, J.; Zhang, Q.; Gu, W. Potential distribution of Notopterygium incisum Ting ex H. T. Chang and its predicted responses to climate change based on a comprehensive habitat suitability model. Ecol. Evol. 2020, 10, 3004–3016. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.J.; Wang, Y.H.; Webb, A.A.; Li, Z.Y.; Tian, X.; Han, Z.T.; Wang, S.; Yu, P. Influence of climatic and geographic factors on the spatial distribution of Qinghai spruce forests in the dryland Qilian Mountains of Northwest China. Sci. Total Environ. 2018, 612, 1007–1017. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, P.; Wang, Y.; Wang, S.; Liu, X. Biomass Change of Middle Aged Forest of Qinghai Spruce along an Altitudinal Gradient on the North Slope of Qilian Mountains. Sci. Silvae Sin. 2015, 51, 4–10. [Google Scholar] [CrossRef]
- Zhong, L.; Ma, Y.; Salama, M.S.; Su, Z. Assessment of vegetation dynamics and their response to variations in precipitation and temperature in the Tibetan Plateau. Clim. Chang. 2010, 103, 519–535. [Google Scholar] [CrossRef]
- Jianmeng, F. Spatial patterns of species diversity of seed plants in China and their climatic explanation. Biodivers. Sci. 2008, 16, 470–476. [Google Scholar] [CrossRef]
- Liu, J.; Yang, Y.; Wei, H.; Zhang, Q.; Zhang, X.; Gu, W. Assessing Habitat Suitability of Parasitic Plant Cistanche deserticola in Northwest China under Future Climate Scenarios. Forests 2019, 10, 823. [Google Scholar] [CrossRef] [Green Version]
- Tromp-van Meerveld, H.J.; McDonnell, J.J. On the interrelations between topography, soil depth, soil moisture, transpiration rates and species distribution at the hillslope scale. Adv. Water Resour. 2006, 29, 293–310. [Google Scholar] [CrossRef]
- Feeley, K.J. Distributional migrations, expansions, and contractions of tropical plant species as revealed in dated herbarium records. Glob. Chang. Biol. 2012, 18, 1335–1341. [Google Scholar] [CrossRef]
- Wolf, A.; Zimmerman, N.B.; Anderegg, W.R.L.; Busby, P.E.; Christensen, J. Altitudinal shifts of the native and introduced flora of California in the context of 20th-century warming. Glob. Ecol. Biogeogr. 2016, 25, 418–429. [Google Scholar] [CrossRef]
- He, X.; Burgess, K.S.; Yang, X.F.; Ahrends, A.; Gao, L.M.; Li, D.Z. Upward elevation and northwest range shifts for alpine Meconopsis species in the Himalaya-Hengduan Mountains region. Ecol. Evol. 2019, 9, 4055–4064. [Google Scholar] [CrossRef] [Green Version]
- Turnbull, L.A.; Crawley, M.J.; Rees, M. Are plant populations seed-limited? A review of seed sowing experiments. Oikos 2000, 88, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Frost, G.V.; Epstein, H.E. Tall shrub and tree expansion in Siberian tundra ecotones since the 1960s. Glob. Chang. Biol. 2014, 20, 1264–1277. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.C.; Piao, S.L.; Myneni, R.B.; Huang, M.T.; Zeng, Z.Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and its drivers. Nat. Clim. Chang. 2016, 6, 791. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhuang, Q.; Schaphoff, S.; Sitch, S.; Sokolov, A.; Kicklighter, D.; Melillo, J. Uncertainty analysis of vegetation distribution in the northern high latitudes during the 21st century with a dynamic vegetation model. Ecol. Evol. 2012, 2, 593–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.X.; Miller, P.A.; Smith, B.; Wania, R.; Koenigk, T.; Doscher, R. Tundra shrubification and tree-line advance amplify arctic climate warming: Results from an individual-based dynamic vegetation model. Environ. Res. Lett. 2013, 8, 034023. [Google Scholar] [CrossRef] [Green Version]
- Druel, A.; Ciais, P.; Krinner, G.; Peylin, P. Modeling the Vegetation Dynamics of Northern Shrubs and Mosses in the ORCHIDEE Land Surface Model. J. Adv. Model. Earth Syst. 2019, 11, 2020–2035. [Google Scholar] [CrossRef]
- Holmgren, M.; Lin, C.-Y.; Murillo, J.E.; Nieuwenhuis, A.; Penninkhof, J.; Sanders, N.; Van Bart, T.; Van Veen, H.; Vasander, H.; Vollebregt, M.E.; et al. Positive shrub-tree interactions facilitate woody encroachment in boreal peatlands. J. Ecol. 2015, 103, 58–66. [Google Scholar] [CrossRef]
- Druel, A.; Peylin, P.; Krinner, G.; Ciais, P.; Viovy, N.; Peregon, A.; Bastrikov, V.; Kosykh, N.; Mironycheva-Tokareva, N. Towards a more detailed representation of high-latitude vegetation in the global land surface model ORCHIDEE (ORC-HL-VEGv1.0). Geosci. Model Dev. 2017, 10, 4693–4722. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wu, J.; Luo, Y.-Q.; Li, Z.-Y.; Wang, G.-C.; Han, F.-Z. Morphological and biological investigation of two pioneer Ips bark beetles in natural spruce forests in Qinghai Province, northwest China. For. Stud. China 2008, 10, 19–22. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Source | Symbol | Variables | Unit | Important Variables for Modelling |
|---|---|---|---|---|
| WorldClim | Bio1 | Annual mean temperature | °C | |
| Bio2 | Mean diurnal range | °C | ||
| Bio3 | Isothermality (BIO2/BIO7) (×100) | % | ||
| Bio4 | Temperature seasonality (standard deviation ×100) | °C | √ | |
| Bio5 | Max temperature of warmest month | °C | ||
| Bio6 | Min temperature of coldest month | °C | ||
| Bio7 | Temperature annual range (BIO5-BIO6) | °C | √ | |
| Bio8 | Mean temperature of wettest quarter | °C | ||
| Bio9 | Mean temperature of driest quarter | °C | ||
| Bio10 | Mean temperature of warmest quarter | °C | ||
| Bio11 | Mean temperature of coldest quarter | °C | ||
| Bio12 | Annual precipitation | mm | √ | |
| Bio13 | Precipitation of wettest month | mm | ||
| Bio14 | Precipitation of driest month | mm | √ | |
| Bio15 | Precipitation seasonality (coefficient of variation) | 1 | √ | |
| Bio16 | Precipitation of wettest quarter | mm | ||
| Bio17 | Precipitation of driest quarter | mm | ||
| Bio18 | Precipitation of warmest quarter | mm | √ | |
| Bio19 | Precipitation of coldest quarter | mm | √ | |
| DEM | ASL | Elevation | m | √ |
| SLOP | Slope | ° | √ | |
| ASPE | Aspect | ° | √ | |
| HWSD | S_GRAVEL | Subsoil gravel content | %vol | |
| S_SAND | Subsoil sand fraction | %wt | ||
| S_SILT | Subsoil silt fraction | %wt | ||
| S_CLAY | Subsoil clay fraction | %wt | √ | |
| S_USDA_TEX_CLASS | Subsoil USDA texture classification | name | ||
| S_REF_BULK_DENSITY | Subsoil reference bulk density | kg/dm3 | ||
| S_ BULK_DENSITY | Subsoil bulk density | kg/dm3 | √ | |
| S_OC | Subsoil organic carbon | % weight | ||
| S_PH_H2O | Subsoil pH (H2O) | −log(H+) | ||
| S_CEC_CLAY | Subsoil CEC (clay) | cmol/kg | √ | |
| S_CEC_SOIL | Subsoil CEC (soil) | cmol/kg | √ | |
| S_BS | Subsoil base saturation | % | √ | |
| S_TEB | Subsoil TEB | cmol/kg | ||
| S_CACO3 | Subsoil calcium carbonate | % weight | ||
| S_CASO4 | Subsoil gypsum | % weight | √ | |
| S_ESP | Subsoil sodicity (ESP) | % | ||
| S_ECE | Subsoil salinity (Elco) | dS/m |
| Symbol | Variables | Picea crassifolia | Sabina przewalskii | Potentilla parvifolia | |||
|---|---|---|---|---|---|---|---|
| Contribution (%) | Cumulative Percentage (%) | Contribution (%) | Cumulative Percentage (%) | Contribution (%) | Cumulative Percentage (%) | ||
| ASPE | Aspect | 33.9 | 33.9 | 51.7 | 51.7 | 56.1 | 56.1 |
| Bio14 | Precipitation of driest month | 23.1 | 57.0 | 3.4 | 55.1 | 3.5 | 59.6 |
| ASL | Elevation | 20.2 | 77.2 | 26.0 | 81.1 | 15.9 | 75.5 |
| Bio12 | Annual precipitation | 10.7 | 87.9 | 0.0 | 81.1 | 4.9 | 80.4 |
| Bio4 | Temperature seasonality | 6.0 | 93.9 | 0.0 | 81.1 | 0.2 | 80.6 |
| S_CEC_CLAY | Subsoil CEC (clay) | 1.2 | 95.1 | 0.4 | 81.5 | 0.5 | 81.1 |
| S_BULK_DEN | Subsoil bulk density | 0.3 | 95.4 | 0.9 | 82.4 | 2.3 | 83.4 |
| S_CEC_SOIL | Subsoil CEC (soil) | 0.2 | 95.6 | 0.1 | 82.5 | 0.1 | 83.5 |
| Suitable Habitat | The Area of Potential Suitable Habitat for Three Species (km2) | The Area at Current (km2) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2050s | 2070s | |||||||||
| SSP2.6 | SSP4.5 | SSP7.0 | SSP8.5 | SSP2.6 | SSP4.5 | SSP7.0 | SSP8.5 | |||
| P. crassifolia | High | 10,271.02 | 10,171.20 | 8483.41 | 10,813.65 | 6294.73 | 4882.79 | 9386.29 | 4767.55 | 8830.95 |
| Moderately | 33,347.47 | 34,940.89 | 42,712.89 | 30,644.28 | 4143.71 | 34,927.27 | 26,032.80 | 54,663.53 | 32,876.52 | |
| Low | 55,668.94 | 47,247.23 | 57,771.42 | 53,073.74 | 43,502.34 | 58,147.09 | 48,217.26 | 48,614.71 | 57,495.57 | |
| Total | 99,287.43 | 92,359.32 | 108,967.72 | 94,531.67 | 53,940.78 | 97,957.15 | 83,636.35 | 108,045.79 | 99,203.04 | |
| S. przewalskii | High | 625.21 | 1883.79 | 361.15 | 0 | 774.02 | 1046.25 | 232.30 | 120.69 | 194.19 |
| Moderately | 7243.89 | 4968.09 | 3235.84 | 1260.40 | 3468.14 | 4416.38 | 6272.95 | 1838.42 | 3327.49 | |
| Low | 32,359.29 | 18,678.21 | 16,681.90 | 13,747.32 | 26,963.81 | 22,868.65 | 31,810.31 | 18,537.56 | 17,761.72 | |
| Total | 40,228.39 | 25,530.09 | 20,278.89 | 15,007.72 | 31,205.97 | 28,331.28 | 38,315.56 | 20,496.67 | 21,283.40 | |
| P. parvifolia | High | 3047.10 | 991.80 | 2301.20 | 1096.16 | 1153.32 | 3744.90 | 2755.82 | 915.58 | 3167.78 |
| Moderately | 20,949.47 | 13,109.41 | 18,451.36 | 9,434.38 | 14,813.53 | 15,636.56 | 13,672.01 | 19,009.42 | 21,045.65 | |
| Low | 86,588.17 | 83,692.61 | 92,607.04 | 59,658.84 | 81,386.87 | 80,101.97 | 99,566.01 | 99,144.06 | 77,965.92 | |
| Total | 110,584.74 | 97,793.82 | 113,359.60 | 70,189.38 | 97,353.72 | 99,483.43 | 115,993.84 | 119,069.06 | 102,179.35 | |
| Suitable Habitat | The Average Elevation under Different Climate Scenario (m) | Average Elevation at Current (m) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2050s | 2070s | |||||||||
| SSP2.6 | SSP4.5 | SSP7.0 | SSP8.5 | SSP2.6 | SSP4.5 | SSP7.0 | SSP8.5 | |||
| P. crassifolia | High | 2773 | 2834 | 2790 | 2849 | 2961 | 2891 | 2691 | 3043 | 2758 |
| Moderately | 3033 | 3000 | 3124 | 3022 | 2972 | 2964 | 2987 | 3077 | 3031 | |
| Low | 3353 | 3333 | 3374 | 3383 | 3298 | 3335 | 3077 | 3375 | 3359 | |
| S. przewalskii | High | 2597 | 3093 | 3103 | —— | 3090 | 2894 | 2764 | 2649 | 2951 |
| Moderately | 2765 | 3487 | 3305 | 3149 | 3334 | 3224 | 2775 | 2878 | 3175 | |
| Low | 3079 | 3312 | 3130 | 3049 | 3066 | 3081 | 3090 | 2996 | 3083 | |
| P. parvifolia | High | 3712 | 3987 | 3795 | 3552 | 3831 | 3728 | 3640 | 4220 | 3617 |
| Moderately | 3578 | 3597 | 3537 | 3717 | 3822 | 3605 | 3625 | 3586 | 2636 | |
| Low | 3499 | 3539 | 3502 | 3711 | 3587 | 3482 | 3520 | 3629 | 3475 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, H.; Wei, Y.; Wang, W.; Wang, C. The Influence of Climate Change on Three Dominant Alpine Species under Different Scenarios on the Qinghai–Tibetan Plateau. Diversity 2021, 13, 682. https://doi.org/10.3390/d13120682
Hu H, Wei Y, Wang W, Wang C. The Influence of Climate Change on Three Dominant Alpine Species under Different Scenarios on the Qinghai–Tibetan Plateau. Diversity. 2021; 13(12):682. https://doi.org/10.3390/d13120682
Chicago/Turabian StyleHu, Huawei, Yanqiang Wei, Wenying Wang, and Chunya Wang. 2021. "The Influence of Climate Change on Three Dominant Alpine Species under Different Scenarios on the Qinghai–Tibetan Plateau" Diversity 13, no. 12: 682. https://doi.org/10.3390/d13120682
APA StyleHu, H., Wei, Y., Wang, W., & Wang, C. (2021). The Influence of Climate Change on Three Dominant Alpine Species under Different Scenarios on the Qinghai–Tibetan Plateau. Diversity, 13(12), 682. https://doi.org/10.3390/d13120682