1. Introduction
In one organism, a colony in hydroids [
1], along with the normal forms of shoot structure, various deviations from the norm can sometimes be found. They are of great interest when studying the mechanisms of morphogenesis. They offer potential resources of variability in morphogenesis—the realization of parallel ways of development of the organism.
The phenomenon of “morphogenetic polyvariance” was described in hydroids for the first time 45 years ago in
Dynamena pumila (L., 1758), belonging to the Sertulariidae family of the order Leptothecata [
2]. Since that time, the study of this phenomenon has continued, but never before the results of the work were published in English.
Morphogenetic polyvariance is the ability of an organism to produce some alternative phenotypes besides the normal (canonical). Alternative phenotypes are called “morphovariations”.
The majority of species have more than one canonical phenotype; for example, hydroids have polyps (or colonial stage of development) and medusa phenotypes. A lot of species of insects’ life cycles include the stages of the worm, bunch, and winged forms. Among flatworms, there are some examples of complex life cycles with more than two phenotypes.
Between canonic and alternative phenotypes, there is a principal difference. Canonic phenotypes manifest a natural regularity. Each phenotype arises at a definite stage of individual development.
Morphogenetic polyvariance emerges in a fundamentally different way. First of all, morphovariations arise: (1) occasionally (stochastically), and (2) without obvious connection with exogenous factors.
Morphogenetic polyvariance is not easy to recognize in a unitary organism. Comparing individuals of the same species, it is difficult to determine to what extent the detected deviation is a simple consequence of population variability or genetic mutation.
It is much easier to detect and study morphogenetic polyvariance in modular organisms, which include colonial invertebrates, many plants and fungi [
3,
4,
5,
6]. Modular organisms are characterized by reproduction of the same form (module—zooid, leaf, branch, etc.) many times. Among the many stereotypical modules, there are abnormal ones, but all of them, both normal and abnormal modules, are implemented according to the same genetic program, based on a single genotype of a given organism.
Nevertheless, among the whole set of modular organisms, only in one species, D. pumila, the phenomenon of morphogenetic polyvariance has been studied in sufficient detail. This is due to two circumstances. First, D. pumila forms dense populations at the boundary between the littoral and the upper sublittoral, which makes it easy to collect colonies in any quantity, in any season and from different habitats. Secondly, any deviations from the norm in the structure of D. pumila shoots are easy to recognize.
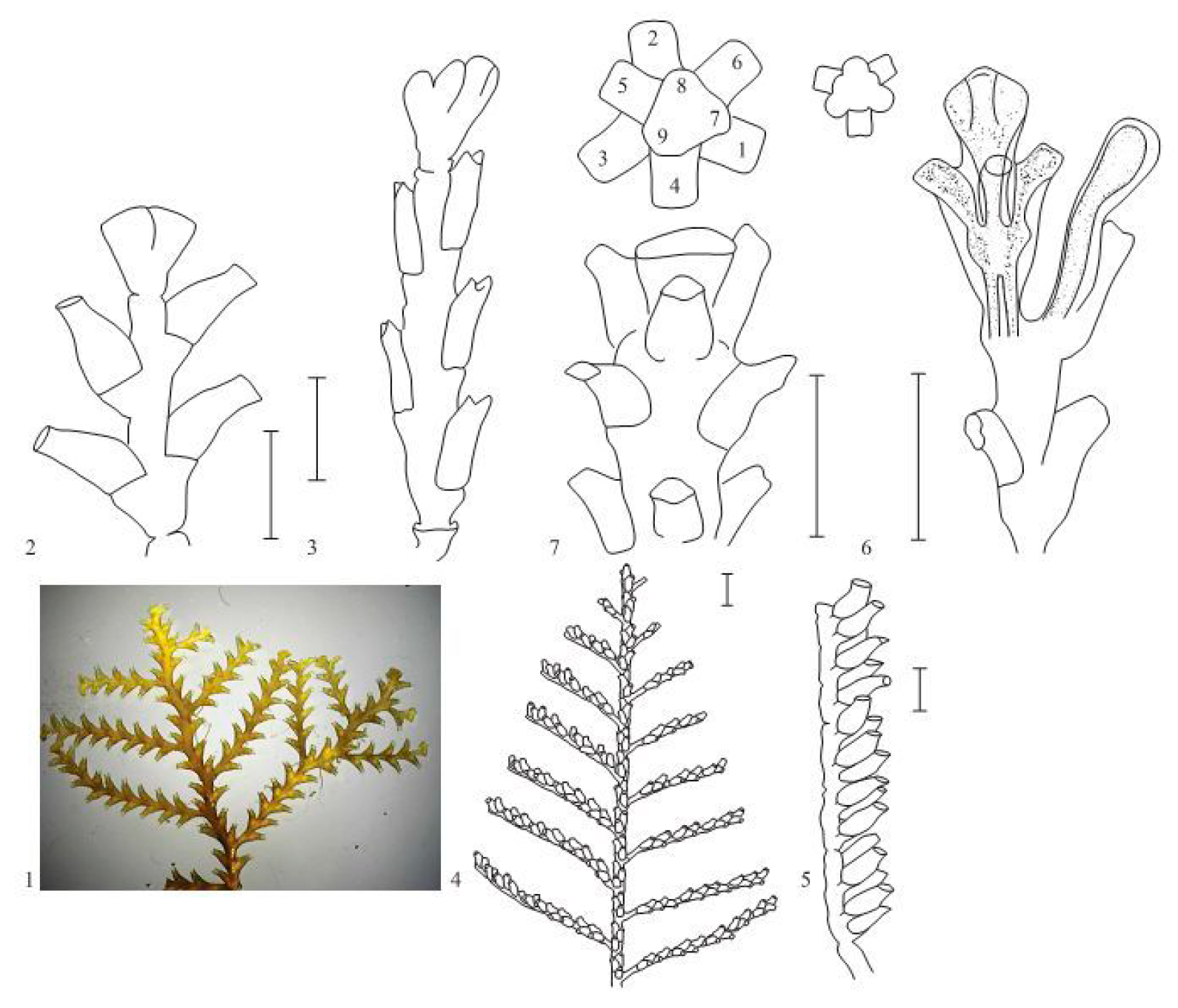
Among all Leptothecata, D. pumila is distinguished by the particularly symmetrical form of its shoots, in which the hydrants (and their perisarcal sheaths—hydrotheca) are located opposite one another in two rows. The slightest distortion in the formation is immediately noticeable by the deviation of the shape of the shoot from the ideally symmetric.
This is why morphogenetic polyvariance was found easily in D. pumila. For this species, research methods were developed based mainly on the classification of morphovariations and their quantitative accounting.
By summarizing all the results obtained over many years of research, for the first time in this article the phenomenon of morphogenetic polyvariance will be described, focusing on the following issues:
- (a)
How diverse are morphological aberrations?
- (b)
Are they deformities?
- (c)
Is there a limit to deviations from the norm of the structure?
- (d)
Are there any regular differences between morphovariations in terms of their frequency of occurrence?
- (e)
How often do morphological aberrations appear in the colony?
2. Materials and Methods
2.1. Characteristics of the Studied Species
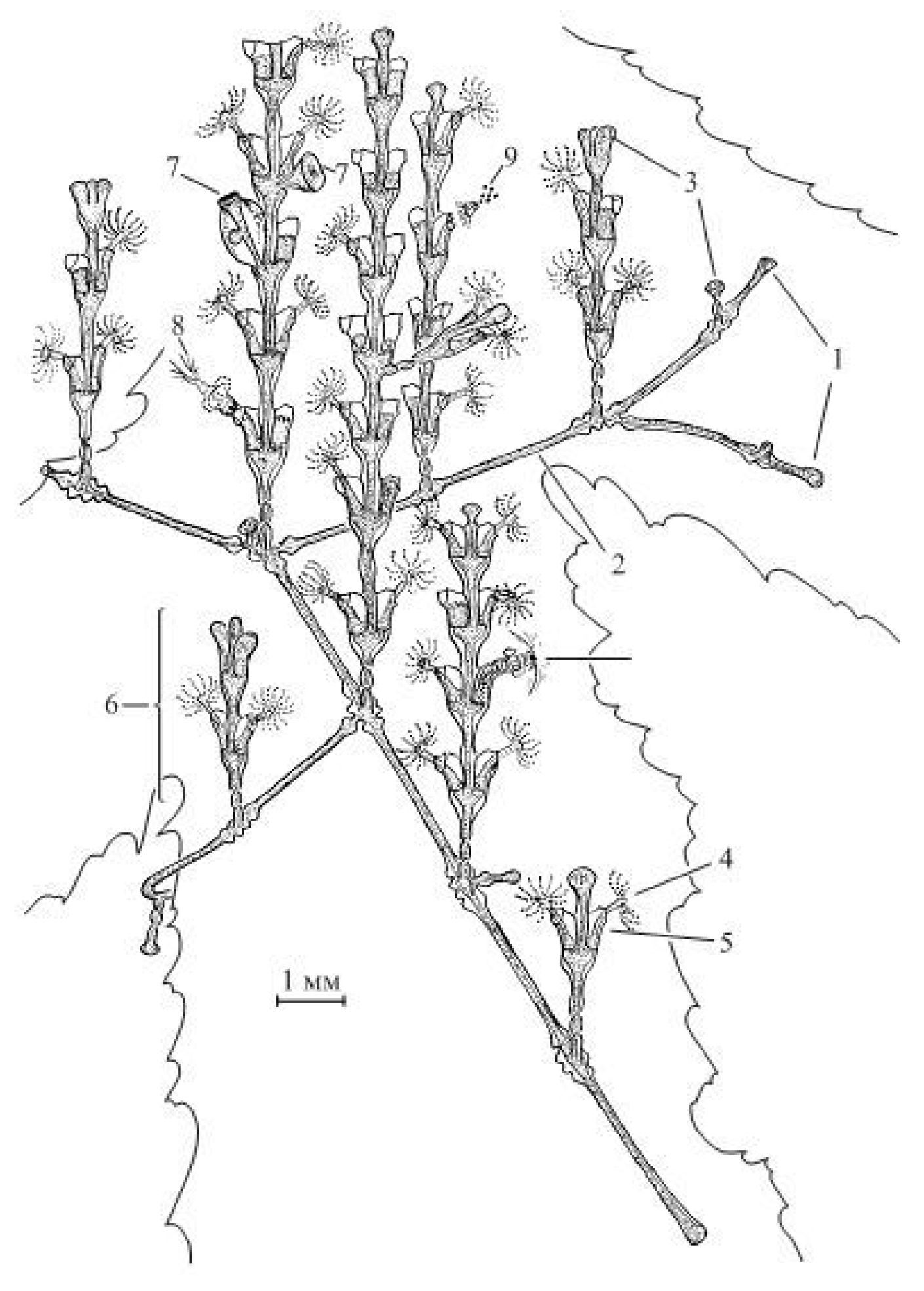
The colonial hydroid
D. pumila consists of many shoots, arising from a smooth, tubular stolon. The hydrants inside hydrothecae are located on the shoot stem directly opposite in two rows, connected by the common coenosarc [
7,
8,
9,
10]. All this is covered by an external chitinous perisarc, whose shape is quite complex and species-specific (
Figure 1). The shoot and its lateral branches are arranged in the same way, differing neither in the size and shape of the hydrotheca, nor in their mutual arrangement.
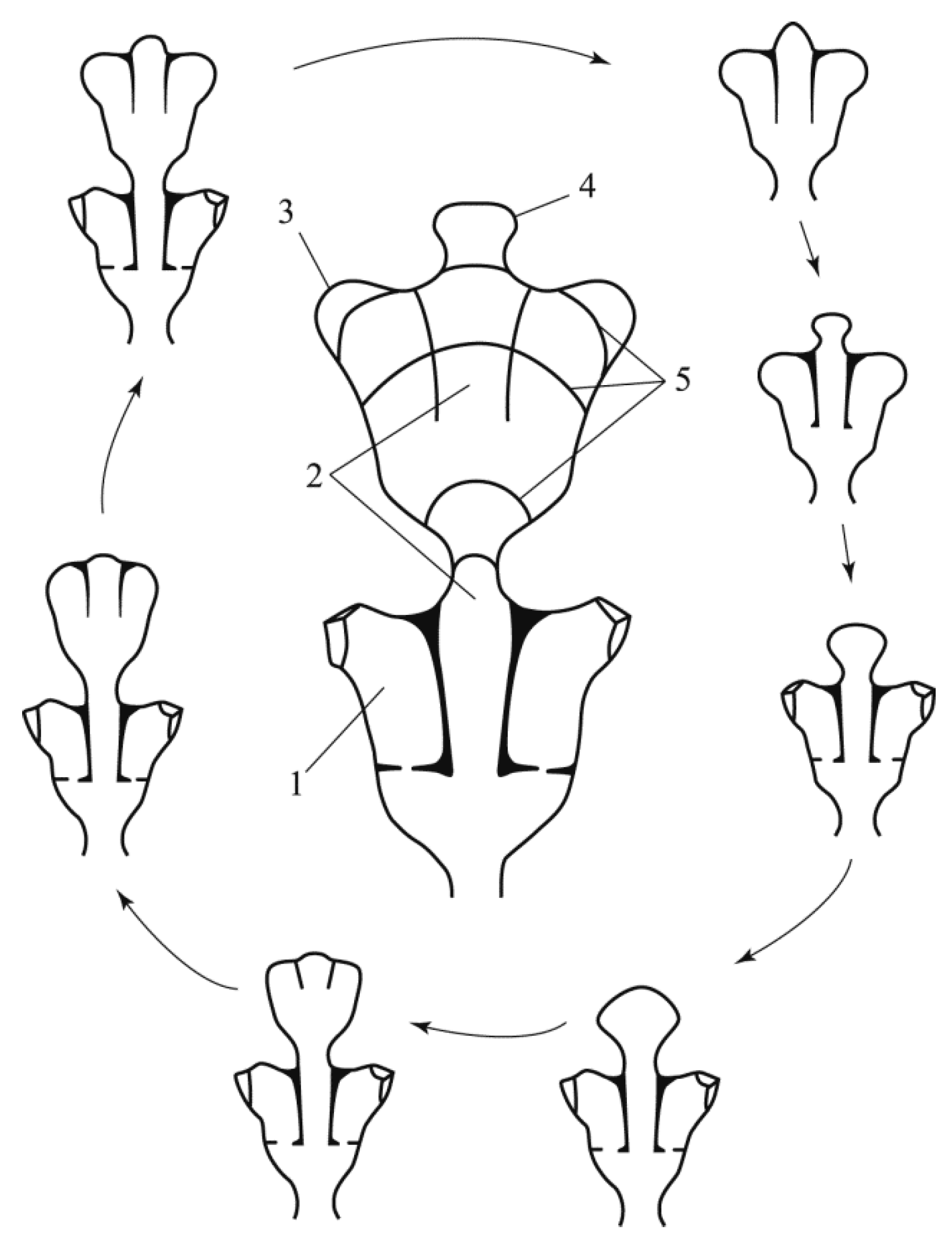
Repeated (cyclic) morphogenesis [
11] means that in the process of differentiation, in addition to the parts characteristic of a given shoot module, a new primordium is also formed, from which the formation of the next module will begin (
Figure 2). That is, cyclic morphogenesis is characterized by the formation of a rudiment at the end of a series of transformations of the same size and shape as the one from which the growth of the next module began.
The escape modules of D. pumila have highly stable, accurate and rich characteristic features such as the shape of the opening of the hydrothecae; the aperture at the base of hydrotheca; the defined length ratio of adjacent and distant parts of the hydrotheca; the strict alternation of wide and narrow parts of the shoot. Therefore, any deviations from the norm are clearly distinguishable when studying shoots under a stereomicroscope.
The cyclical morphogenesis in D. pumila makes it possible to track the process of the multiple formations of stereotypical colony modules and compare them with each other, helping determine the nature and degree of environmental influence on the development of the organism, or setting the limit of endogenous phenotypic variability.
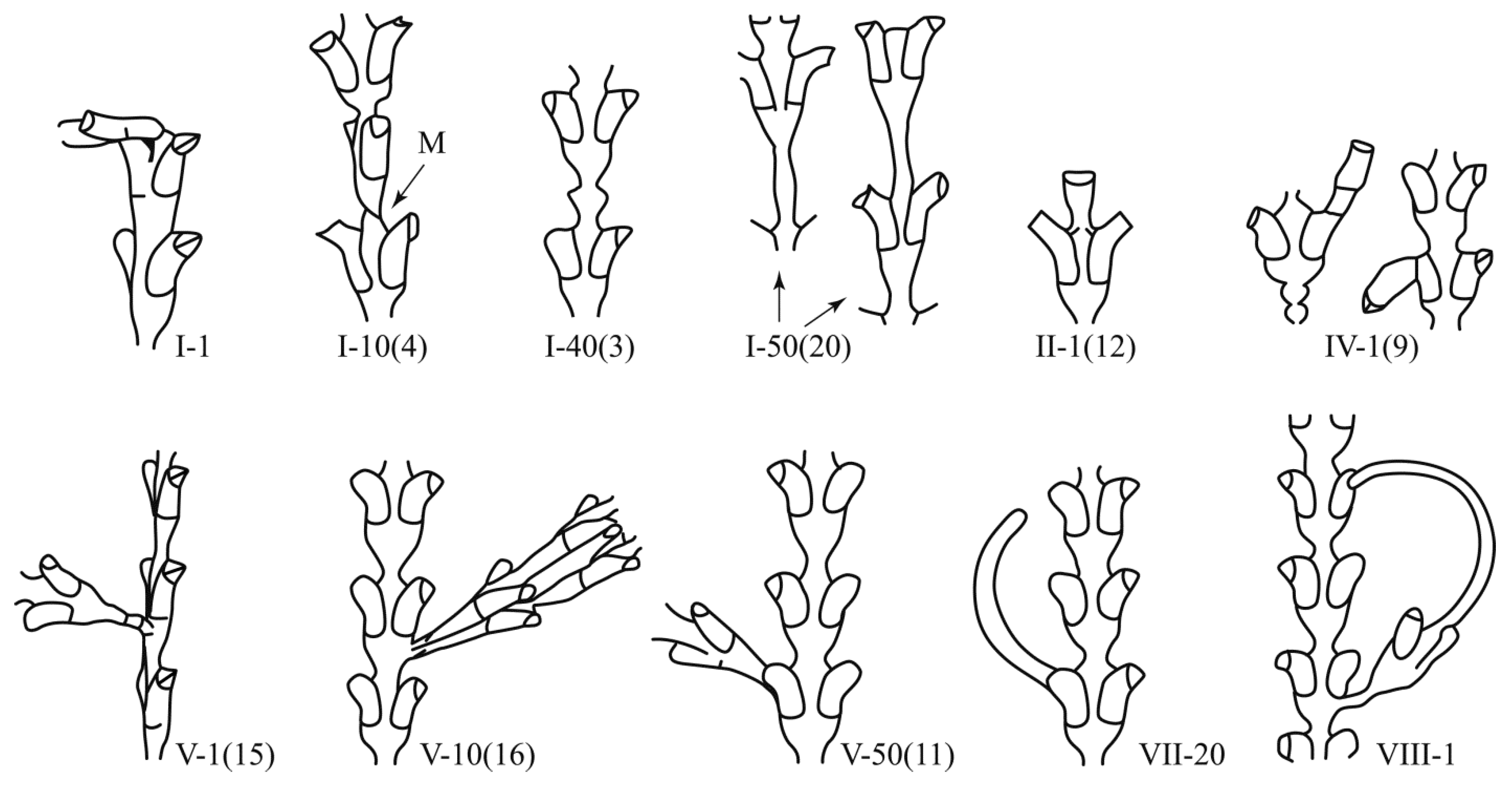
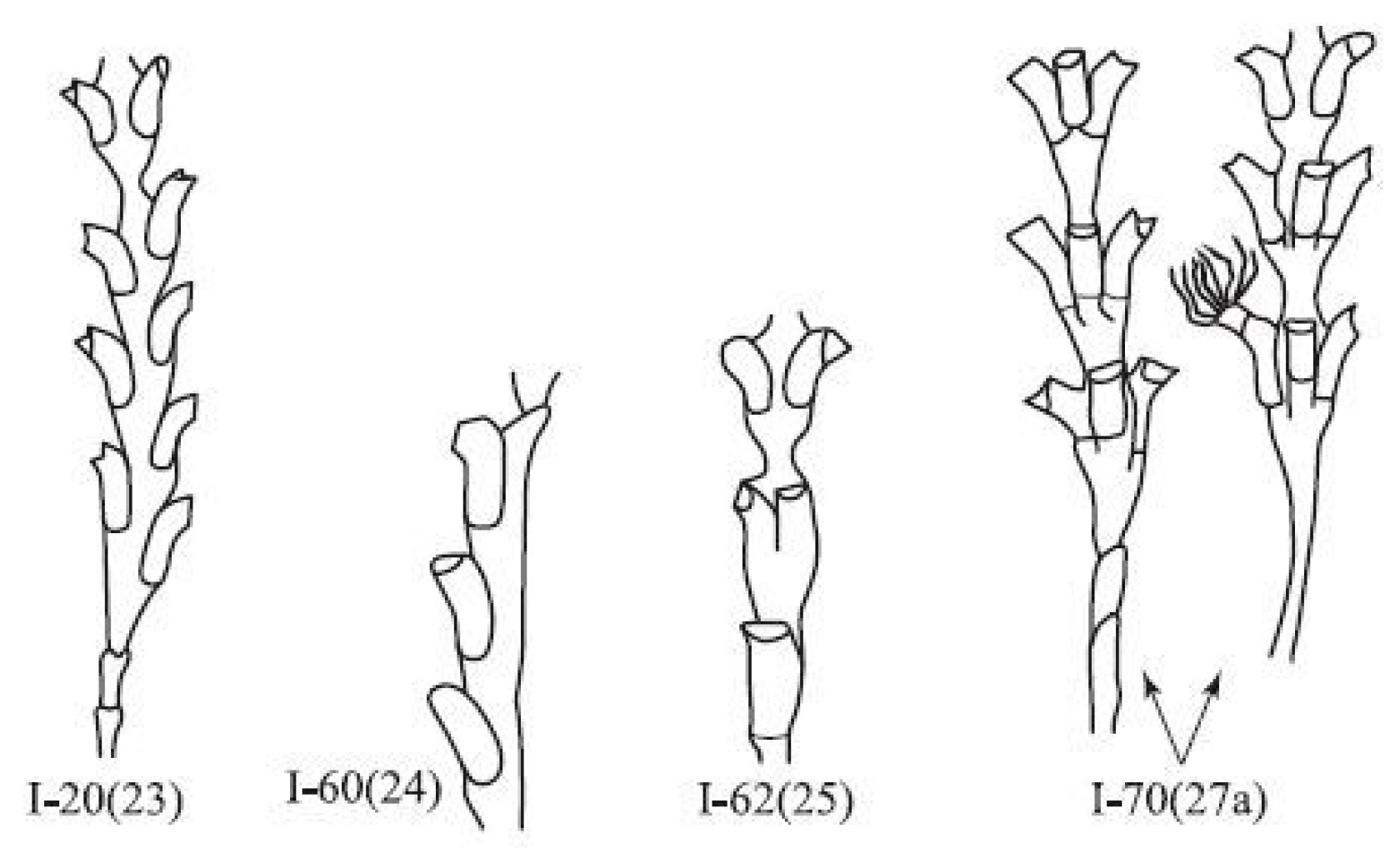
Deviations from the usual (canonical) form of the module are called aberrant or anomalous modules. All anomalies must be subdivided into those that are capable of performing their vital functions, and others that are not capable of this. The latter are essentially aberrant. Aberrant modules are very diverse, and therefore difficult to classify, while viable anomalous modules have characteristic forms and their morphogenesis has clear signs of stability. Therefore, such anomalous modules are called morphovariations. Some of the morphovariations turn out to be so similar to the modules of shoots of hydroids from other genera that it would be correct to call them morphotypes. If the morphovariations are rarely repeated in sequentially formed modules, then the morphotypes are often repeated, thus demonstrating certain morphogenetic stability.
2.2. Collection of Material
The material was collected near the White Sea biological station of Moscow State University in Kandalaksha Bay. To determine the degree of homogeneity in the D. pumila population, several samples were taken at a distance of 40 km from Sidorov Island near Chupa Bay, as well as at 840 km out into the Barents Sea, near the station of the Murmansk Marine Biological Institute in the village of Dalnie Zelentsyi. Colonies of hydroids together with algae on which they lived were collected in summer 1992, 1993, 1994, 2000, 2005.
2.3. Methodology for Accounting for the Composition and Frequency of Occurrence of Deformities, Morphogenetic Variations and Morphotypes
To determine the composition and frequency of occurrence of alternative morphovariations in D. pumila shoots, the following parameters were taken into account:
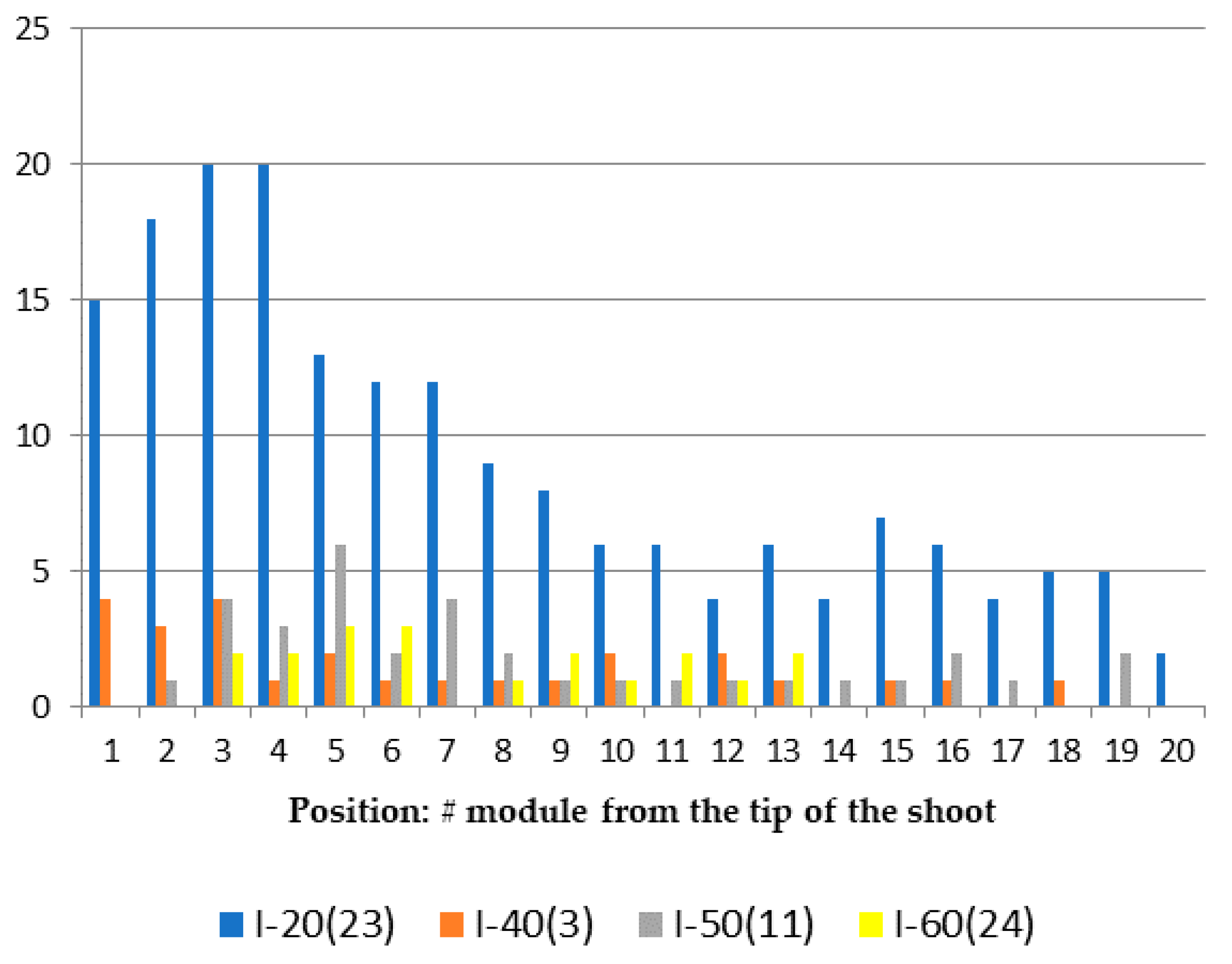
(1) The number of modules in the shoot stem; (2) the number of modules in each of the branches; (3) morphovariations in the structure of the shoot modules, identifying them according to the classifications published earlier (Marfenin, 1995) or adding new ones, if necessary; (4) the places of the unusual modules on the branch, counting from the apex.
The unusually shaped shoot modules were drawn and photographed. All anomalies were selected and stored separately at the Department of Invertebrate Zoology, Lomonosov Moscow State University (Moscow, Russia).
In total, 40 samples were examined, including 280,000 modules in the White Sea population and 140,000 modules in the Barents Sea population of D. pumila. These included 11,000 and 4000 shoots, respectively.
For each sample, it was possible to indicate:
the cumulative proportion (%) of all morphovariations, hereinafter referred to as the cumulative frequency of occurrence of morphovariations (CFOM);
the absolute value of the occurrence of each morphovariation (FOM) in % of the total number of studied modules.
Roman numerals are the alleged reasons for the aberration of the module and its location, namely:
Group I: Deviations in the morphogenesis of the shoot stem without stopping its further growth;
Group II: Deviations in the morphogenesis of the shoot tip, preventing its further growth;
Group III: Juvenile ugly shoots;
Group IV: Deviations from the norm of the structure of the hydrotheca;
Group V: Deviations from the norm of the location of the lateral branches of the shoot;
Group VI: Deviations from the norm of the shape and position of the gonotheca;
Group VII: Deviations from the norm in the position of stolons in the colony;
Group VIII: Deviations in shoot morphogenesis, possibly caused by damage to it, as well as incomprehensible abnormalities of unknown origin.
Arabic numerals are the registration numbers of the morphovariations within the groups.
Arabic numerals in brackets are the ordinal numbers in the first classification [
2], in which there were only 27 morphovariations.
2.4. Valid Sample Size
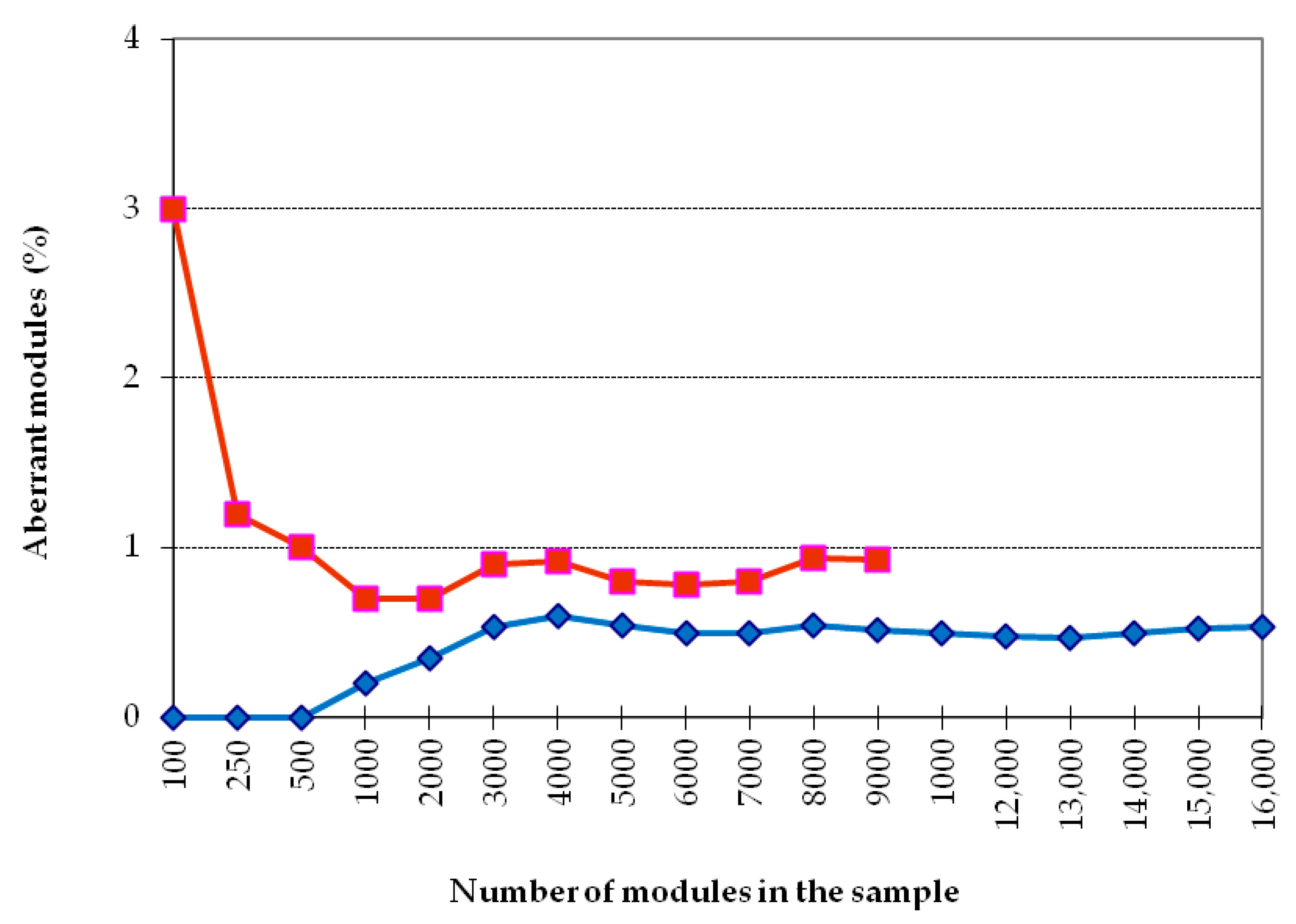
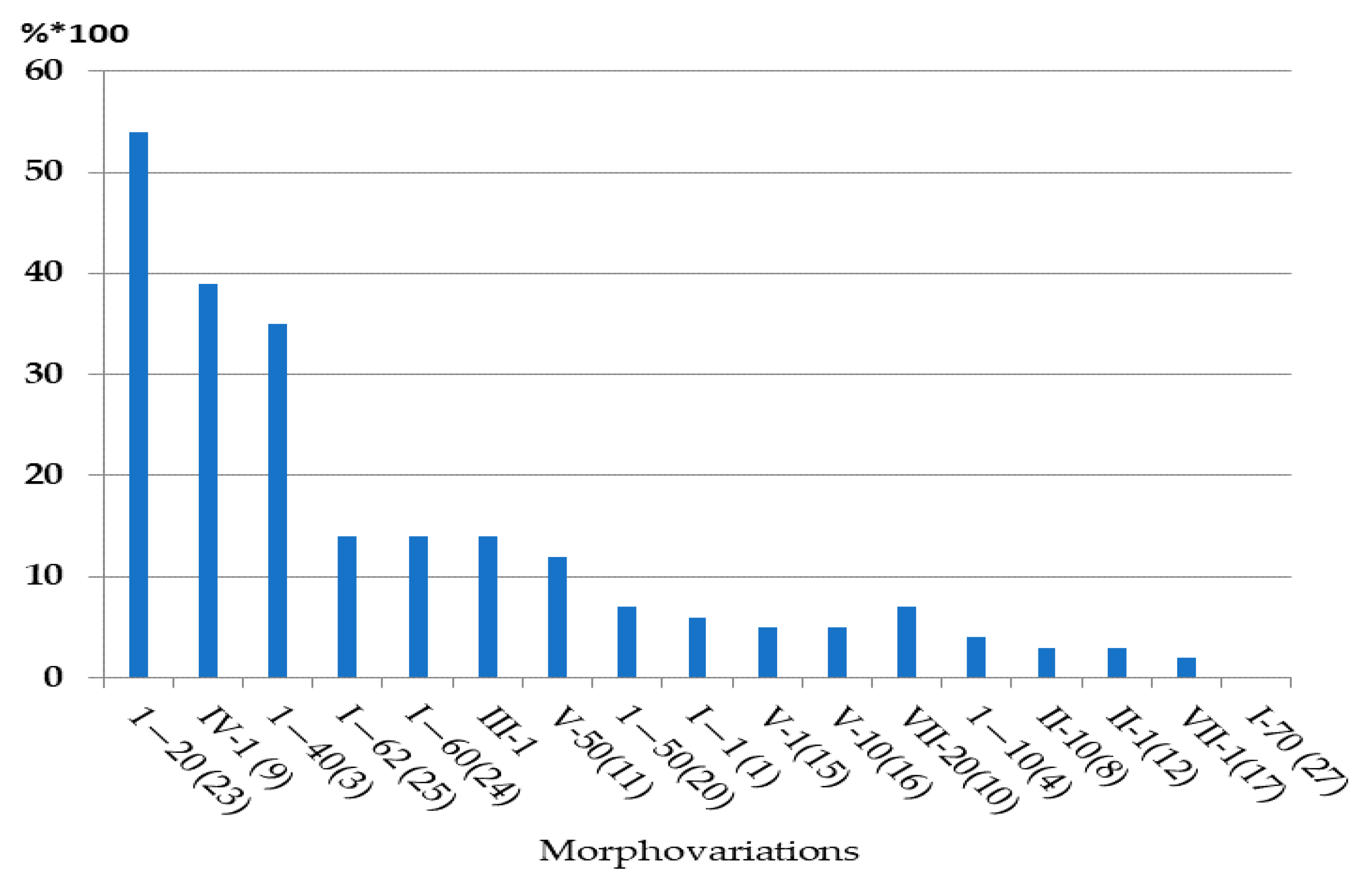
First of all, it was necessary to establish the size of a reliable sample for the analysis of the cumulative frequency of occurrence of morphovariations (CFOM).
Figure 3 shows the dependence of CFOM on the sample size, divided into two portions according to the size of shoots (<20 and >20 modules). The same calculations were carried out for a number of other samples. CFOM stabilization occurred in all cases after the sample reached approximately 2000–3000 modules. Stabilization was not necessarily accompanied by a gradual increase in the CFOM value as the sample increased; there were examples of gradual decreases in CFOM to a stable level. Thus, to determine the long exponent, the CFOM was limited to the processing of the modules of 3000 shoot’s modules of
D. pumila. Since the cumulative frequency of occurrence mainly comprises several dominant morphovariations, a separate analysis of the most widespread varieties of deviation in the structure of the shoots using the same sample size is reliable.
However, for the analysis of the occurrence of more rare morphovariations, the established sample is insufficient. Having increased the sample to 10,000 modules, we created conditions for the comparative assessment of not only the indicator of the total frequency of occurrence of all abnormal modules but also the occurrence of most of the usual morphovariations individually. Thus, a sample of 10,000 modules was taken as a standard.
Only in some cases, lacking material, I used samples with less than 10,000 modules—we used as little as 3000, and then only for analyzing the indicator of the cumulative frequency of occurrence of all morphovariations (CFOM).
4. Discussion
Morphogenetic polyvariance is of great use for understanding the mechanisms of morphogenesis. When abnormalities are found in single organisms, there is usually no doubt that they are the result of genetic mutations. Rare deviations from the norm in a colonial organism are perceived in a fundamentally different way. The abnormal module is preceded by completely normal ones, and after it, the same normal modules occur—in our case, the normal segments of the shoots of the D. pumila colony. Therefore, morphogenetic aberrations cannot be the result of genetic mutations, but at the same time, they do not adhere to the usual ideas about morphological variability. At least six types of morphological variability in organisms are known:
Sexual dimorphism;
Polymorphism—the presence of several variants in the zooid’s morphology in the colony (Hydrozoa, Bryozoa);
Sequential change in phenotypes in the life cycle—for example, in the classes Hydrozoa and Scyphozoa, the formation of a medusoid stage after a polypoid one may occur. In many parasitic organisms, we see phenotype changes during the transition from one stage of the life cycle to the next; in the Insecta class, individual development usually includes metamorphosis. There are many similar examples. Moreover, ontogeny in any species can be represented by a series of phenotypes;
Intrapopulation morphological variability— this reflects the breadth of variations in morphological characteristics that accumulate in the gene pool of a species during the crossing and exchanging of genetic information;
Phenotypic plasticity—a change in the shape of an organism or its individual parts in response to a change in the habitat;
Deformities—deviations in the shape and structure of the body from the norm as a result of genetic mutations, or other causes. Depending on the nature of the mutations, deformities can be inherited or appear in the same generation. In the latter case, one speaks of somatic mutations.
Sexual dimorphism and polymorphism are obligatory and stable manifestations of phenotypic polyvariance fixed in the genotype. Additionally, successive changes in the phenotypes in the life cycle are fixed genetically. In hydroids, this is expressed firstly in a change in polypoid and medusoid generations of many species, and secondly, in the presence of several zooids, such as gastrozooids, gonozooids, dactilozooids, etc. Morphogenetic polyvariance, such as that in
D. pumila, does not correspond to the natural manifestation of polymorphism, since morphovariations arise out of connection with the stages of development of the colony or its life cycle [
13,
14,
15].
The intrapopulation variability of morphological characteristics manifests only when comparing organisms with one another. In each individual organism, the phenotype is predetermined by the individual characteristics of the genotype. This does not correspond to cases of aberrant modules being detected in one colony.
Phenotypic plasticity is a consequence of the broad reaction rate of a species. Morphological variations generated by phenotypic plasticity always occur in response to external factors. Phenotypic plasticity has always attracted the attention of researchers, and there is a huge number of publications on this subject.
The phenomenon of morphogenetic plasticity, which we found in the colonial hydroid D. pumila in the form of a deviation from the norm in the structures of shoot modules, cannot be attributed to either polymorphism or phenotypic plasticity. The results of our study of morphovariations in D. pumila indicate that anomalies appear regardless of the state of the environment; they are not sufficiently genetically fixed to manifest in each individual at a certain stage of its individual development. At the same time, morphological abnormalities occur rarely, but with a certain stable frequency. Among the variations in the structure of the module, there are unconditional deformities, but most of the variations are distinguished by a harmonious structure, and in some cases the morphovariations are similar to the phenotypes of the shoot modules of other genera of the same family (Sertulariidae).
Therefore, morphogenetic plasticity is not reducible to the known forms of morphological variations but is an independent phenomenon. Its main feature is that morphogenesis occurs only in some of the many simultaneously forming modules of one organism, while in all the others, morphogenesis proceeds according to the usual program.
To date, it has been established that the presence of alternative morphovariations in
D. pumila is not limited to samples collected in the White and Barents Seas, i.e., on the borders of the range of this species. The same morphovariations were also found in colonies collected in the North Sea near Helgoland [
16].
After examining samples that include several hundreds of thousands of modules and developing a more complete classification of morphovariations, I came to two general conclusions:
Despite the dozens of described morphogeneses, the real diversity of deviations in the morphogenesis of modules of the D. pumila shoot is very small; and
most morphovariations derive from the morphogenesis typical of this type.
At the same time, individual morphovariations are fundamentally different from the norm in terms of the structure and order of module formation. They are very much resembling the structure of the shoot in hydroids from other genera, which may be a manifestation of the species’ ability to implement an alternative morphogenetic program.
In general, all deviations from the norm in D. pumila fit into four groups: (a) deformities; (b) morphogenetic abnormalities resulting from growth arrest; (c) morphogenetic failures in the position of the module; (d) alternative morphogenetic programs or morphotypes.
Such forms have no obvious adaptive value but look viable. If deformity occurs at the tip of the shoot, then the next shoot cannot grow. If the hydrant turns out to be aberrant, then it cannot fulfill its food-producing function.
Morphogenetic anomalies that appear after the suspension of shoot growth look quite viable. These include the bulbous expansion of the trunk, a sharp curvature of the trunk, or a change in the orientation of the plane of the trunk, etc. Resumption of growth can occur on a limited surface of the apex, rather than across its entire working surface, which leads to the formation of unusual shapes, such as bulbous expansions and the bending of the trunk. However, there may be other reasons for the latter anomaly. With a chronic lack of food, a distortion in the proportions of the module is possible; for example, the lengthening of its lower thinnest part, which is associated with an increase in the period of expansion of the apex in the morphogenetic cycle (personal observations).
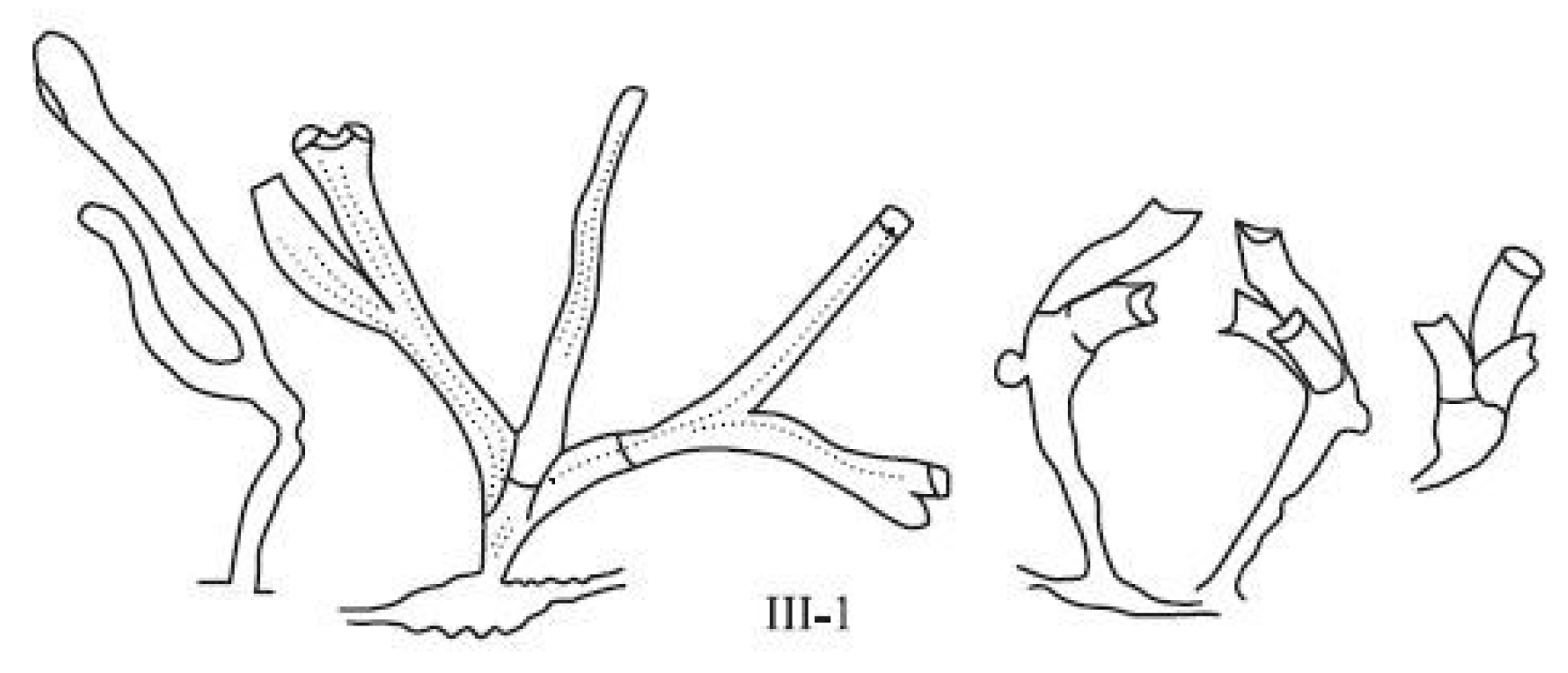
Morphogenetic mistakes include all cases of incorrect body position when its structure is basically normal, for example, an offshoot or stolon grows instead from the hydrothecae (see morphovariation VII-20(10)); the entire growth tip turns into one hydrant (I-1(12)); the lateral branch departs from the trunk not in its frontal plane, but perpendicular to it (V-1(15)), etc. Often, the growth of an organ in the “wrong” place is associated with growth arrest or the process of secondary morphogenesis after hydrant resorption. Therefore, morphogenetic disruptions can be considered special cases of morphogenetic anomalies. However, for the subsequent comparison of the spectra of morphovariations, it is convenient to consider them separately.
Morphotypes are of particular interest, since, firstly, the appearance of alternative morphotypes is associated with a radical change in the shape of the growth apex and the order of its differentiation into rudiments (buds), and, secondly, the transition to a different type of morphogenesis turns out to be temporary even after several cycles, since in the same shoot normal morphogenesis is restored, and so after unusual modules, completely normal ones emerge.
If the appearance of aberrant modules does not lead to the cessation of shoot growth, then subsequent modules on the same shoots are, as a rule, completely normal. This feature is important to the search for mechanisms of morphogenetic polyvariance.
Deviations from the norm in the development of the organism are associated, first of all, with mutations—genetic or somatic. The appearance of alternative phenotypes in a colony of hydroids is difficult to explain via mutations since along with abnormal modules, there are always completely normal ones in the colony. Usually, in a colony, several tens, or even hundreds, of shoot tips grow at the same time. While in one of these the morphogenesis turned out to differ from the canonical one, in all the others it did not differ. Since the entire colony is genetically identical, there is no reason to assume that the main cause of the appearance of abnormal modules is genetic mutations. Somatic mutations are also not easily defined as the main mechanism of morphogenetic polyvariance. In the apical part of the growth apex—in the zone of morphogenetic transformations—mitoses are absent. The zone of mitotic activity, i.e., cell proliferation, is located at the base of the growth apex. Even if we assume that a somatic mutation occurs in any cell of the growth apex, and its multiplication affects the subsequent process of morphogenesis, then, consequently, the rudiment of the next module, isolated on the apical surface of the growth apex, should consist of the same mutant cells, which means that the next module must be aberrant. However, this is very rare.
Thus, the return to normal morphogenesis after the realization of a different morphogenetic program should be taken to indicate the nonmutational causes of the morphogenetic polyvariance—supporting the hypothesis of the epigenetic origin of deviations in the structures of modules.
So far, three morphotypes have been discovered that resemble the structures of the shoots of three species belonging to different genera of the Sertulariidae family.
Among all morphovariations, morphotype No. I-20(23) (asymmetric arrangement of hydrothecae) occurs most frequently. This deviation from the norm is very similar to the structures of the shoots in hydroids from the genus
Abietinaria. The hydrothecae are located in two rows, similar to in
D. pumila, but alternately, i.e., with an offset relative to each other in height. In species from the genus
Abietinaria, during the growth of the shoot tip, the primordium of one hydrant is first laid, and then the second primordium is laid on the opposite side of the shoot stem (
Figure 9). Similarly, morphogenesis occurs in
D. pumila when the strictly opposing arrangement of the location of the hydrothecae is deviated from. Asymmetry in the position of the hydrothecae entails the multiplication of this error over and over again during the formation of the following modules. As a result, a significant part of the shoot may be aberrant, i.e., not typical for
D. pumila. Nevertheless, the alternate arrangement of the hydrothecae is usually replaced with an opposite one forming after a narrow constriction (“thin waist”) forms between successive shoot modules. In addition to the position of the hydrothecae, the other distinguishing features of
D. pumila, as a rule, do not change. For example, the opercular apparatus consists of two lids, and not one, as in species from the genus
Abietinaria. The lateral branches are laid not at the growth tips, as in
Abietinaria or
Sertularia suensoni Levinsen, 1913 (
Figure 9), but instead, emerge after some time under the previously formed hydrothecae. The high frequency of occurrence of morphotype No. I-20(23) is due not only to objective differences from the norm, but also to the difficulty of determining the boundary between the norm and deviations from it since such deviations are not discrete, but gradual.
The situation is completely different for morphotype No. I-60(24) (single-row position of the hydrothecae). This is a pronounced discrete anomaly. In this case, there is no difficulty in distinguishing the anomaly from the norm. Several hydrothecae are formed in a row, on only one side of the shoot. The hydrothecae are alternately oriented with their openings facing opposite directions. This structure is typical for the genus
Hydrallmania, in particular for
H. falcata. The morphogenesis of the shoot tip with a single-row structure is fundamentally different from the morphogenesis of a two-row shoot. With a single-row arrangement, the division of the growth apex into two primordia occurs not by a transverse partition, but by an oblique one, which divides the apex into two parts of different sizes, related to each other as 1:2. In morphotype I-20(23), the transverse septum is laid either on one side of the apex or on the other, while in
Hydrallmania, the hydrothecae are all laid on the same side of the shoot apex (
Figure 9), but at alternating angles of tilt (conventionally 45° and 135° relative to the sagittal plane). Notably, in
H. falcata, at the beginning of the growth of a young shoot, a two-row arrangement of hydrothecae was identified [
17,
18,
19]. Consequently, the two-row and single-row phenotypes can be programmed in the genotypes of both
Hydralmania and
Dynamena, although alternative variants are still not species-specific, and occur rarely. The opercular apparatus in the aberrant modules of
D. pumila remains typical of this genus.
Apparently, the three-row arrangement of hydrothecae is also programmed in the
D. pumila genotype. The combination of the extreme rarity of this morphotype with its striking harmony of form is surprising. This is manifested in the fact that an angle of 120° is strictly maintained between the planes of the hydrothecae, and they are not deformed, retaining the shape typical of
D. pumila. Among the species of the Sertulariidae family, we can find some with a three-row arrangement of hydrothecae—if not everywhere, then at least in those with thin branches, such as
Pericladium mirabilis (Verrill, 1873) [
14,
15].
Upon interpreting these facts, we believe that the
D. pumila genotype includes additional information on the construction of a viable “spare” form of shoot module. Such phenotypic plasticity within the same organism can be a reserve of adaptive radiation in biological evolution [
20].
It can be assumed that the phenomenon of morphogenetic polyvariance should also manifest itself in other modular organisms: corals, siphonophores, bryozoans, camptozoas, colonial ascidians, as well as outside the animal kingdom in many plants and fungi. Of these colonial invertebrates, only corals and bryozoans have been the subject of morphological studies. There are few studies on intracolonial morphovariations in the structure of zooids [
21]. The shape variability of corallites is a well-known phenomenon [
22,
23,
24,
25]. The structure of the colony itself in corals varies even more—not only in its size and branching sites but also in other features, such as the relationship between the length and thickness of the branch [
26,
27,
28]. However, this phenomenon is always explainable by one of two mechanisms: it is either a reaction to environmental factors or the effect of positional regulation, namely, a variation in the shape of an organ depending on its location in the body.
It would seem that in plants, this phenomenon should have attracted the attention of researchers. However, phenotypic plasticity is traditionally interpreted as the result of a reaction to a change in the external environment, to which hundreds of articles are devoted (of which I cite only a few to illustrate the diversity of research angles [
29,
30,
31,
32]). Unfortunately, none of the authors of these works have suggested that deviations in the structures of stereotypical modules in plants can occur on their own without connection with environmental factors or genetic changes. We know of one very significant study of leaf morphovariations, published only in Russian almost a century ago [
33].
Indeed, in plants, it is difficult to distinguish changes in the shape of a module, for example, a leaf, resulting from endogenous causes, from those caused by exogenous causes, for example, the intensity or spectral characteristics of light [
34,
35,
36,
37,
38,
39].
Until now, no dependence of the morphovariation frequency in the structure of the shoot module of D. pumila on environmental factors was found. As such, it is important to note that aberrant modules never appear in D. pumila shoots in simultaneous series on parallel-growing shoots, but are instead located “at different levels” from the tips of growth, i.e., form at different times.
The second important feature of morphogenetic polyvariance is the significant consistency in the frequencies of occurrence of different morphogenetic variations. The cumulative frequency of all morphogenetic anomalies together varies within narrow limits, from 0.5 to 4% of the total number of counted modules, but mainly within 0.9–1.5%. The frequencies of occurrence of each morphovariation have also turned out to be limited to narrow intervals. The order of dominance of morphovariations changes little from sample to sample, i.e., the sequence of morphovariation, arranged in decreasing order of their frequency of occurrence within the analyzed sample. Altogether, the CFOA, FOA, and the order of dominance of morphovariations represent a kind of “morphogenetic portrait of the species”, carrying information about its internal morphogenetic potency.
The stability of the frequency of occurrence of the morphovariations found in
D. pumila is similar to that in the morphological abnormalities found in
Drosophila caused by mutation of the Hsp90 protein. In the mutant heterozygous line, in which the action of the protein is suppressed, various deviations from the norm in the structures of various organs arise [
40]. The authors believe that the Hsp90 protein suppresses (buffers) the manifestation of natural morphogenetic variations in the developmental pathways of the organism that remain hidden (Hsp90 buffers the variation affecting the morphogenic pathways that exist in nature, but it is usually silent). With a reduction in the activity of this protein, latent morphogenetic polyvariance occurs, and in the future, under the influence of selection, it can gain a foothold. The frequency of the manifestation of deviations from the norm in the Hsp90 mutant line was 1–2%, and the frequency of occurrence in the offspring indicated genetic fixation. The authors came to the conclusion of the existence of silent genes that predetermine abnormal morphological signs through variations in developmental pathways, using the example of a unitary [
41] rather than a modular organism, in which this phenomenon would be easier to detect. The results of studies on morphovariations in the colonial hydroid
D. pumila confirm this hypothesis.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}