
Soil Bacterial and Fungal Composition and Diversity Responses to Seasonal Deer Grazing in a Subalpine Meadow



Abstract
:1. Introduction
2. Materials and Methods
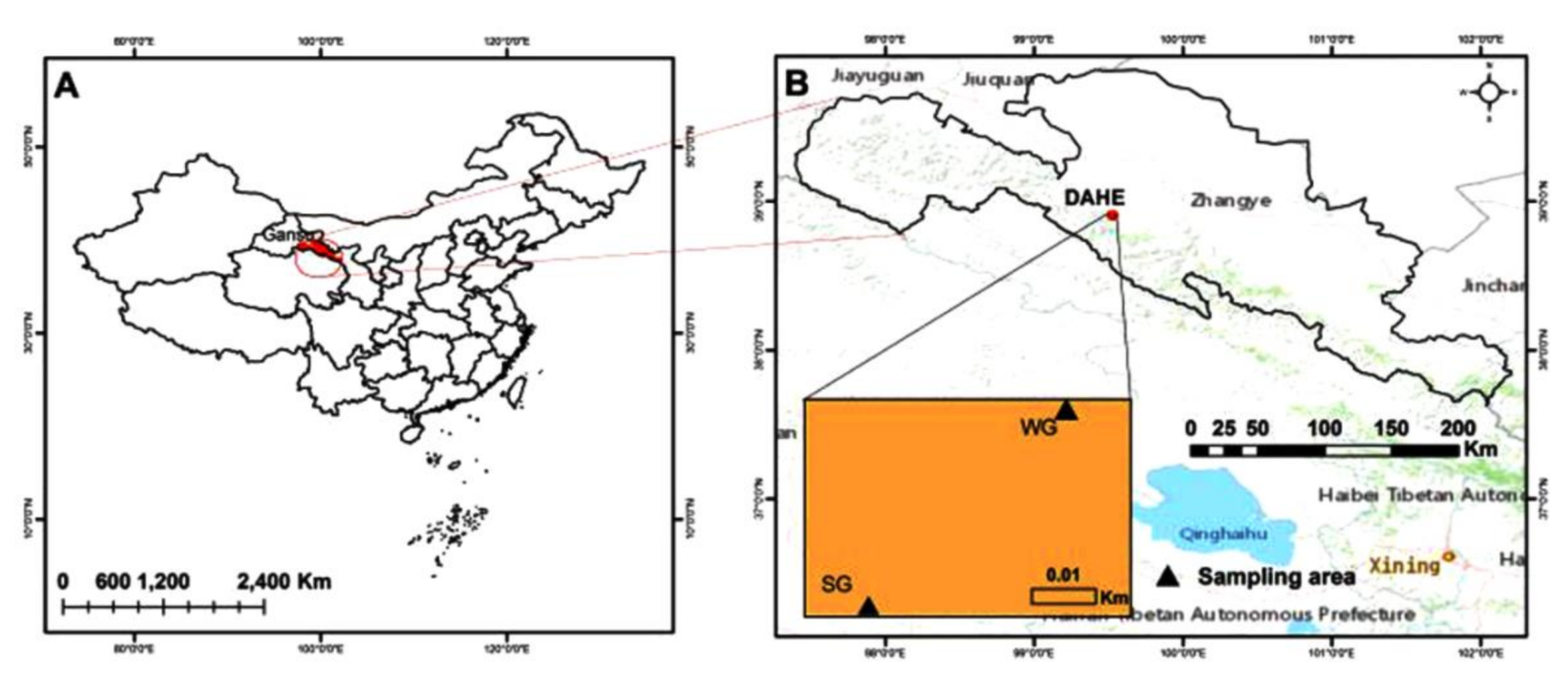
2.1. Research Site
2.2. Sample Collection and Processing
2.3. Soil Physicochemical Analysis
2.4. Microbial Community Analysis
2.5. Statistical Analysis
2.6. Nucleotide Sequence Accession Numbers
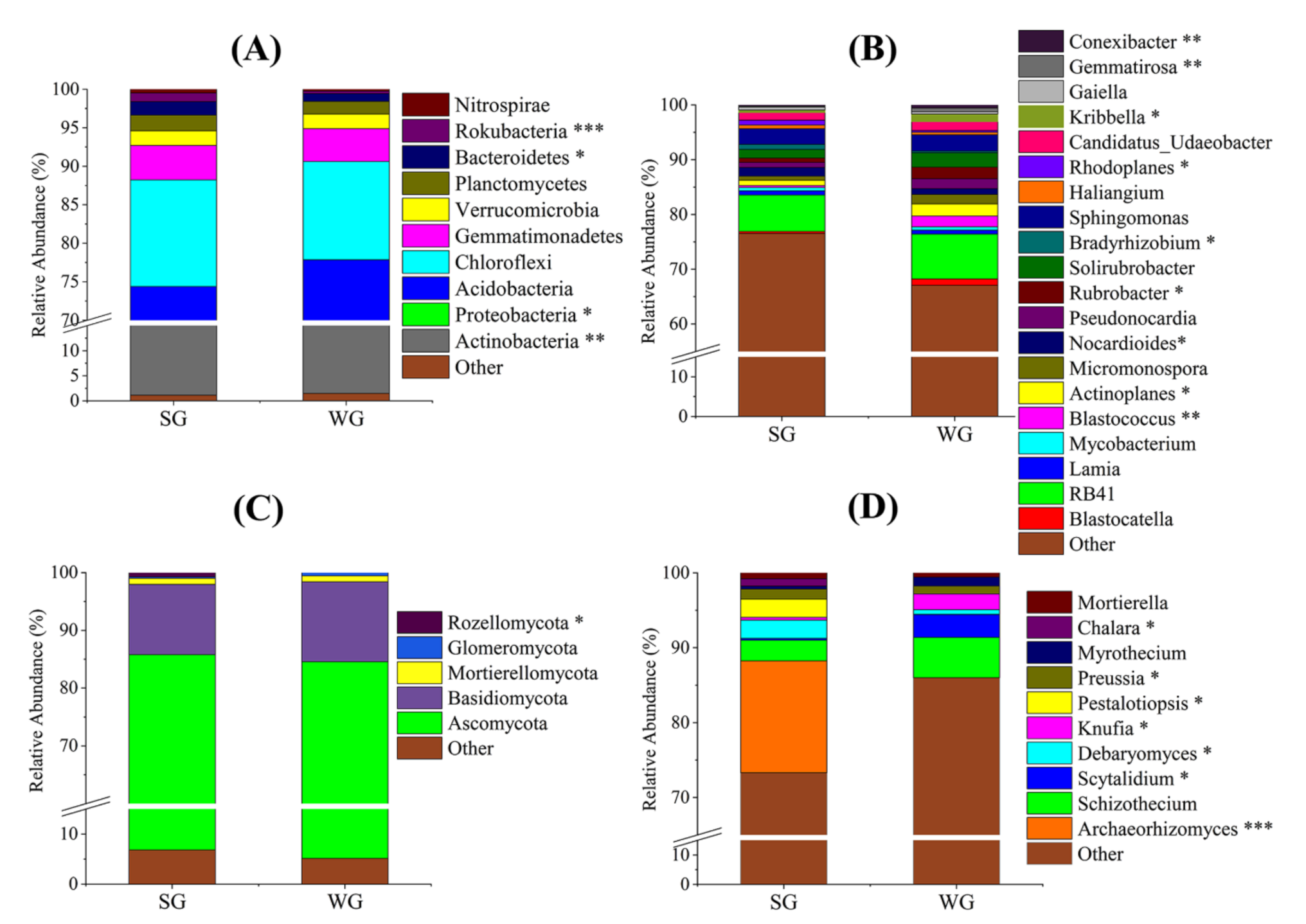
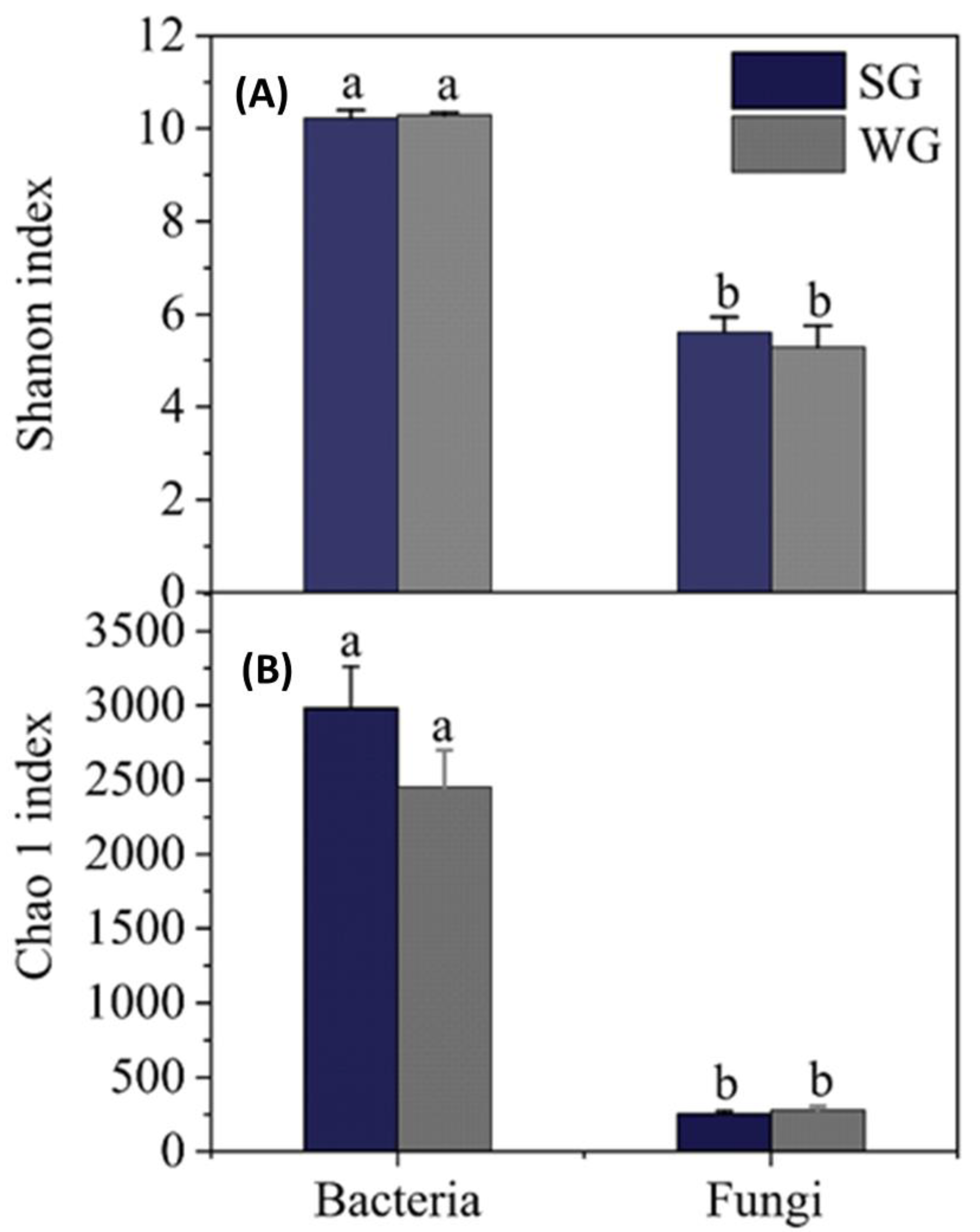
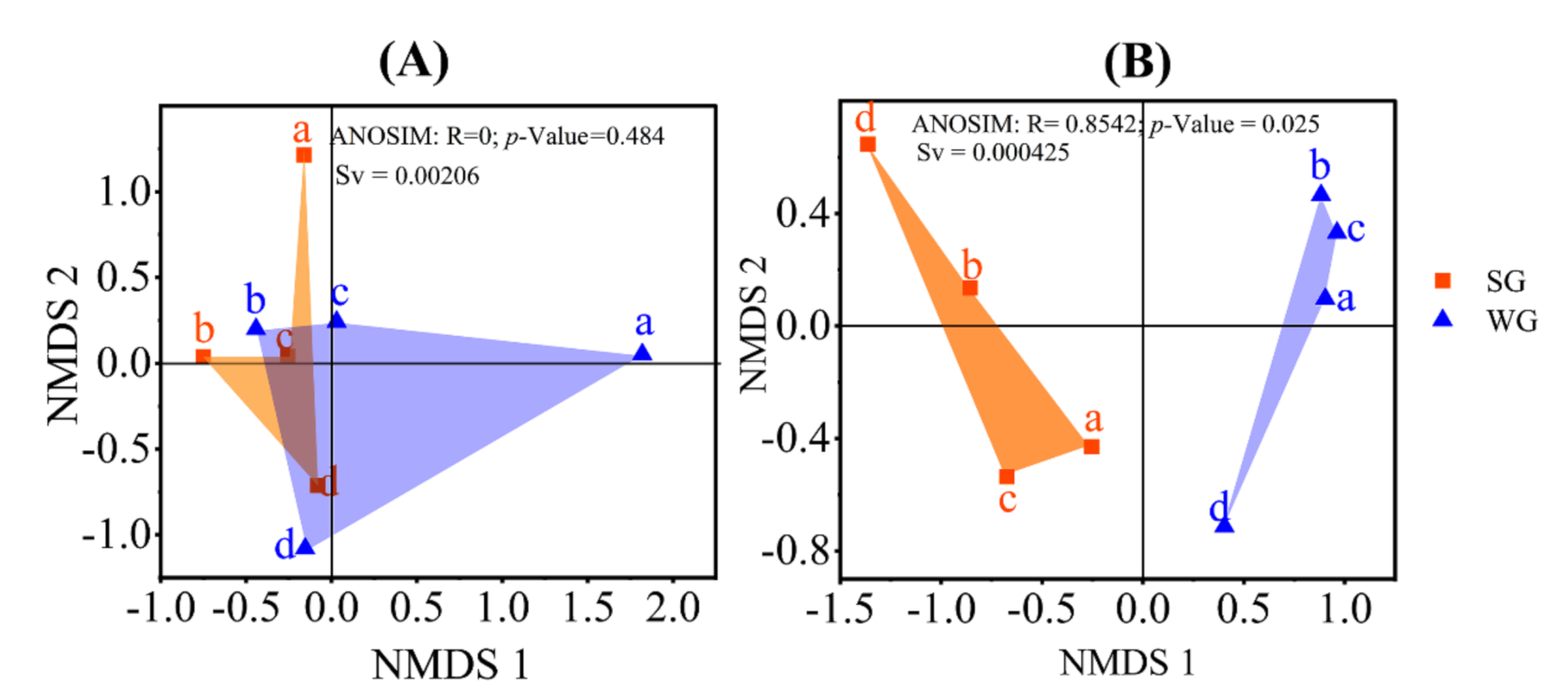
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgements
Conflicts of Interest
References
- Franzluebbers, A.J.; Paine, L.K.; Winsten, J.R.; Krome, M.; Sanderson, M.A.; Ogles, K. Well-Managed Grazing Systems: A Forgotten Hero of Conservation. J. Soil Water Conserv. 2012, 67, 100A–104A. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.; Gourley, C.; Rotz, C.A.; Weaver, D.M. Nitrogen Use Efficiency: A Potential Performance Indicator and Policy Tool for Dairy Farms. J. Environ. Sci. 2010, 13, 217–228. [Google Scholar] [CrossRef]
- Enri, S.R.; Probo, M.; Farruggia, A.; Lanore, L.; Blanchetete, A.; Dumont, B. A Biodiversity-Friendly Rotational Grazing System Enhancing Flower-Visiting Insect Assemblages While Maintaining Animal and Grassland Productivity. Agric. Ecosyst. Environ. 2017, 241, 1–10. [Google Scholar] [CrossRef]
- García, R.R.; Celaya, R.; García, U. Goat Grazing, Its Interactions with Other Herbivores and Biodiversity Conservation Issues. Small Rumin. Res. 2012, 107, 49–64. [Google Scholar] [CrossRef] [Green Version]
- Bilotta, G.; Brazier, R. The Impacts of Grazing Animals on the Quality of Soils, Vegetation, and Surface Waters in Intensively Managed Grasslands. Adv. Agron. 2007, 94, 237–280. [Google Scholar]
- Greenwood, K. Grazing Effects on Soil Physical Properties and the Consequences for Pastures: A Review. Aust. J. Exp. Agric. 2001, 41, 1231–1250. [Google Scholar] [CrossRef]
- O’Connor, K. Comments on Dr Floate’s Paper on Grazing Effect by Large Herbivores. Ecol. Bull. 1981, 33, 707–714. [Google Scholar]
- Miao, F.; Guo, Z.; Xue, R.; Wang, X. Effects of Grazing and Precipitation on Herbage Biomass, Herbage Nutritive Value, and Yak Performance in an Alpine Meadow on the Qingha Tibetan Plateau. PLoS ONE 2015, 10, e0127275. [Google Scholar] [CrossRef]
- Soder, K.; Rook, A.; Sanderson, M. Interaction of Plant Species Diversity on Grazing Behavior and Performance of Livestock Grazing Temperate Region Pastures. Crop. Sci. 2007, 47, 416–425. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, J.B.; Thorpe, A.S. Livestock Exclusion and Belowground Ecosystem Responses in Riparian Meadows of Eastern Oregon. Ecol. Appl. 2004, 14, 1671–1679. [Google Scholar] [CrossRef]
- Kobayashi, T.; Hori, Y. Effects of Trampling and Vegetation Removal on Species Diversity and Micro-Environment under Different Shade Conditions. J. Veg. Sci. 1997, 8, 873–880. [Google Scholar] [CrossRef]
- Hou, F.J.; Nan, Z.; Xie, Y.; Li, X.; Lin, H. Integrated Crop-Livestock Production Systems in China. Rangel. J. 2008, 30, 221–231. [Google Scholar] [CrossRef]
- Taboada, M.A.; Rubio, G.; Enrique, J. Grazing Impacts on Soil Physical, Chemical, and Ecological Properties in Forage Production Systems. In Soil Management: Building a Stable Base for Agriculture; Hatfield, J.L., Thomas, J.S., Eds.; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 2011; pp. 301–320. [Google Scholar]
- Porensky, L.M.; Derner, J.D.; Augustine, D.J.; Milchunas, D.G. Plant Community Composition after 75 Yr of Sustained Grazing Intensity Treatments in Shortgrass Steppe. Rangel. Ecol. Manag. 2017, 70, 456–464. [Google Scholar] [CrossRef]
- Rahmanian, S.; Hejda, M.; Ejtehadi, H.; Farzam, M.; Memariani, F.; Pyšek, P. Effects of Livestock Grazing on Soil, Plant Functional Diversity, and Ecological Traits Vary between Regions with Different Climates in Northeastern Iran. Ecol. Evol. 2019, 9, 8225–8237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalho, I.O.; Rezende, C.P.; Pereira, J.M.; Macedo, R.O.; Santos, C.A.; Monteiro, R.C.; Boddey, R.M. Deposition and Decomposition of Litter in Periods of Grazing and Rest of a Tropical Pasture under Rotational Grazing. Cienc. Rural 2019, 49. [Google Scholar] [CrossRef]
- Sun, Y.; He, X.Z.; Hou, F.J.; Wang, Z.; Chang, S. Grazing Increases Litter Decomposition Rate but Decreases Nitrogen Release Rate in an Alpine Meadow. Biogeosciences 2018, 15, 4233–4243. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.; Abbadie, L.; Clays-Josserand, A.; Degrange, V.; Grayston, S.; Loiseau, P. Effects of Grazing on Microbial Functional Groups Involved in Soil N Dynamics. Ecol. Monogr. 2005, 75, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Gou, Y.; Nan, Z.; Hou, F.J. Diversity and Structure of a Bacterial Community in Grassland Soils Disturbed by Sheep Grazing in the Loess Plateau of Northwestern China. Genet. Mol. 2015, 14, 16987–16999. [Google Scholar] [CrossRef]
- Qi, S.; Zheng, H.; Lin, Q.; Li, G.; Xi, Z.; Zhao, X. Effects of Livestock Grazing Intensity on Soil Biota in a Semiarid Steppe of Inner Mongolia. Plant Soil 2011, 340, 117–126. [Google Scholar] [CrossRef]
- Jangid, K.; Williams, M.A.; Franzluebbers, A.J.; Sanderlin, J.S.; Reeves, J.H.; Jenkins, M.B. Relative Impacts of Land-Use, Management Intensity and Fertilization Upon Soil Microbial Community Structure in Agricultural Systems. Soil Biol. Biochem. 2008, 40, 2843–2853. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.; Hao, Y.; Wang, Y. Intermediate Grazing Intensities by Sheep Increase Soil Bacterial Diversities in an Inner Mongolian Steppe. Biol. Fertil. Soils 2010, 46, 817–824. [Google Scholar] [CrossRef] [Green Version]
- Xun, W.; Yan, R.; Ren, Y.; Jin, D.; Xiong, W.; Zhang, G. Grazing-Induced Microbiome Alterations Drive Soil Organic Carbon Turnover and Productivity in Meadow Steppe. Microbiome 2018, 6, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, T.; Du, W.; Yuan, X.; Yang, Z.; Liu, D.; Wang, D.; Yu, L. Impacts of Grazing Intensity and Plant Community Composition on Soil Bacterial Community Diversity in a Steppe Grassland. PLoS ONE 2016, 11, e0159680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, G.; Song, Z.; Wang, J.; Guo, L. Interactions of Soil Bacteria and Fungi with Plants During Long-Term Grazing Exclusion in Semiarid Grasslands. Soil Biol. Biochem. 2018, 124, 47–58. [Google Scholar] [CrossRef]
- Xiao, X.; Li, D.C.; An, Y.F.; Ma, Z.W.; Wu, Z.; Peng, Z.C.; Hou, F.J. Plant community characteristics of grazing grassland in a deer farm in summer. Pratacultural Sci. 2019, 36, 1693–1705. [Google Scholar]
- Flora of China. Available online: http://www.efloras.org/florataxon.aspx?flora_id=2&taxon_id (accessed on 9 February 2021).
- Ma, Z.W.; Wang, H.; Yu, Y.W.; Chang, S.H.; Zhou, Q.P.; Hou, F.J. Short-term effect of mowing winter pastures of Gansu wapiti on forage regeneration. Pratacultural Sci. 2018, 35, 157–164. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis: Part 3 Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 1996; Volume 5, pp. 961–1010. [Google Scholar]
- Kang, B.; Bowatte, S.; Hou, F.J. Soil Microbial Communities and Their Relationships to Soil Properties at Different Depths in an Alpine Meadow and Desert Grassland in the Qilian Mountain Range of China. J. Arid Environ. 2020, 184, 104316. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar] [CrossRef]
- Shannon, C. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped Blast and Psi-Blast: A New Generation of Protein Database Search Programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. A Language and Environment for Statistical Computing [Computer Software Manual]; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Liu, J.; Feng, C.; Wang, D.; Wang, L.; Wilsey, B.J.; Zhong, Z.J. Impacts of Grazing by Different Large Herbivores in Grassland Depend on Plant Species Diversity. J. Appl. Ecol. 2015, 52, 1053–1062. [Google Scholar] [CrossRef]
- He, Y.; Xu, M.; Qi, Y.; Dong, Y.; He, X.; Li, J.; Sun, L. Differential Responses of Soil Microbial Community to Four-Decade Long Grazing and Cultivation in a Semi-Arid Grassland. Sustainability 2017, 9, 128. [Google Scholar] [CrossRef]
- Metzger, K.; Coughenour, M.; Reich, R.; Boone, R. Effects of Seasonal Grazing on Plant Species Diversity and Vegetation Structure in a Semi-Arid Ecosystem. J. Arid Environ. 2005, 61, 147–160. [Google Scholar] [CrossRef]
- Pulungan, M.A.; Suzuki, S.; Gavina, M.K.A.; Tubay, J.M.; Ito, H.; Nii, M.; Shiyomi, M. Grazing Enhances Species Diversity in Grassland Communities. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Hou, F.J. Grazing Intensity and Soil Depth Effects on Soil Properties in Alpine Meadow Pastures of Qilian Mountain in Northwest China. Acta Agric. Scand. B Soil Plant Sci. 2015, 65, 222–232. [Google Scholar] [CrossRef] [Green Version]
- Ingram, L.; Stahl, P.; Schuman, G.; Buyer, J.; Vance, G.; Ganjegunte, G.; Derner, J. Grazing Impacts on Soil Carbon and Microbial Communities in a Mixed-Grass Ecosystem. Soil Sci. Soc. Am. J. 2008, 72, 939–948. [Google Scholar] [CrossRef]
- Davis, K.E.; Sangwan, P.; Peter, H.J. Acidobacteria, Rubrobacteridae, and Chloroflexi Are Abundant among Very Slow-Growing and Mini-Colony-Forming Soil Bacteria. Environ. Microbiol. 2011, 13, 798–805. [Google Scholar] [CrossRef]
- Deng, J.; Yin, Y.; Zhu, W.; Zhou, Y. Variations in Soil Bacterial Community Diversity and Structures among Different Revegetation Types in the Baishilazi Nature Reserve. Front. Microbiol. 2018, 9, 2874. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.; Dong, Y.; An, S. Bacterial Community Responses to Soils Along a Latitudinal and Vegetation Gradient on the Loess Plateau, China. PLoS ONE 2016, 11, e0152894. [Google Scholar] [CrossRef] [Green Version]
- Belov, A.A.; Cheptsov, V.S.; Vorobyova, E.A. Soil Bacterial Communities of Sahara and Gibson Deserts: Physiological and Taxonomical Characteristics. AIMS Microbiol. 2018, 4, 685. [Google Scholar] [CrossRef] [PubMed]
- Mohammadipanah, F.; Wink, J. Actinobacteria from Arid and Desert Habitats: Diversity and Biological Activity. Front. Microbiol. 2016, 6, 1541. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Chen, B.; Luo, S.; Ma, B.; Qi, W.; Cao, C.; Du, G. Increasing Soil pH Enhances the Network Interactions among Bacterial and Archaeal Microbiota in Alpine Grasslands of the Tibetan Plateau. Prepr. Res. Sq. 2019, PPR149373. [Google Scholar] [CrossRef] [Green Version]
- Mohanta, T.K.; Bae, H. The Diversity of Fungal Genome. Biol. Proced. Online 2015, 17, 8. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Guo, S.; Huang, M.; Thorsten, L.H.; Wei, J. Ascomycota Has a Faster Evolutionary Rate and Higher Spe-cies Diversity Than Basidiomycota. Sci. China Life Sci. 2010, 53, 1163–1169. [Google Scholar] [CrossRef]
- Malik, A.A.; Chowdhury, S.; Schlager, V.; Oliver, A.; Puissant, J.; Vazquez, P.G.; Gleixner, G. Soil Fungal: Bacterial Ratios Are Linked to Altered Carbon Cycling. Front. Microbiol. 2016, 7, 1247. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Zhang, X.; Wang, X.; Fu, S.; Wu, S.; Lu, X. Soil Bacteria and Fungi Respond Differently to Plant Diversity and Plant Family Composition During the Secondary Succession of Abandoned Farmland on the Loess Plateau, China. Plant Soil. 2020, 448, 183–200. [Google Scholar] [CrossRef]
- Prober, S.M.; Leff, J.W.; Bates, S.T.; Borer, E.T.; Firn, J.; Harpole, W.S.; Bakker, J.D. Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide. Ecol. Lett. 2015, 18, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Ulrich, W. Increasing Aridity Reduces Soil Microbial Diversity and Abundance in Global Drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; He, T.; Gen, S.; Zhang, X.Q.; Wang, X.; Jiang, D.; Zhang, W. Soil properties and agricultural practices shape microbial communities in flooded and rainfed croplands. Appl. Soil Ecol. 2020, 147, 103449. [Google Scholar] [CrossRef]
- O’Brien, F.J.M.; Almaraz, M.; Foster, M.; Hill, A.; Huber, D.; King, E.; Miller, V. Soil Salinity and Ph Drive Soil Bacterial Community Composition and Diversity Along a Lateritic Slope in the Avon River Critical Zone Observatory, Western Australia. Front. Microbiol. 2019, 10, 1486. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhou, X.; Guo, D.; Zhao, J.; Yan, L.; Feng, G.; Zhao, L. Soil pH is the Primary Factor Driving the Distribution and Function of Microorganisms in Farmland Soils in Northeastern China. Ann. Microbiol. 2019, 69, 1461–1473. [Google Scholar] [CrossRef] [Green Version]
- Xue, P.P.; Carrillo, Y.; Pino, V.; Minasny, B.; McBratney, A.B. Soil Properties Drive Microbial Community Structure in a Large Scale Transect in South Eastern Australia. Sci. Rep. 2018, 8, 11725. [Google Scholar] [CrossRef] [Green Version]
- Urbina, H.; Breed, M.F.; Zhao, W.; Gurrala, K.L.; Andersson, S.G.; Ågren, J.; Rosling, A. Specificity in Arabidopsis Thaliana Recruitment of Root Fungal Communities from Soil and Rhizosphere. Fungal Biol. 2018, 122, 231–240. [Google Scholar] [CrossRef]
- Dahl, M.B.; Priemé, A.; Brejnrod, A.; Brusvang, P.; Lund, M.; Nymand, J.; Haugwitz, M.S. Warming, shading and a moth outbreak reduce tundra carbon sink strength dramatically by changing plant cover and soil microbial activity. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Pinto-Figueroa, E.A.; Seddon, E.; Yashiro, E.; Buri, A.; Niculita-Hirzel, H.; Van Der Meer, J.R.; Guisan, A. Archaeorhizo-mycetes Spatial Distribution in Soils Along Wide Elevational and Environmental Gradients Reveal Co-Abundance Patterns with Other Fungal Saprobes and Potential Weathering Capacities. Front. Microbiol. 2019, 10, 656. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Puusepp, R.; Nilsson, R.H.; James, T.Y. Novel Soil-Inhabiting Clades Fill Gaps in the Fungal Tree of Life. Microbiome 2017, 5, 42. [Google Scholar] [CrossRef]
- Siles, J.A.; Cajthaml, T.; Filipova, A.; Minerbi, S.; Margesin, R. Altitudinal, Seasonal and Interannual Shifts in Mi-crobial Communities and Chemical Composition of Soil Organic Matter in Alpine Forest Soils. Soil Biol. Biochem. 2017, 112, 1–13. [Google Scholar] [CrossRef]
- Barnett, S.E.; Youngblut, N.D.; Buckley, D.H. Soil Characteristics and Land-Use Drive Bacterial Community Assembly Patterns. FEMS Microbiol. Ecol. 2020, 96, fiz194. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, K.; Wemheuer, B.; Korolkow, V.; Wemheuer, F.; Nacke, H.; Schöning, I.; Daniel, R. Driving Forces of Soil Bacterial Community Structure, Diversity, and Function in Temperate Grasslands and Forests. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Brandón, M.; Probst, M.; Siles, J.A.; Peintner, U.; Bardelli, T.; Egli, M.; Ascher-Jenull, J. Fungal Communities and Their Association with Nitrogen-Fixing Bacteria Affect Early Decom-position of Norway Spruce Deadwood. Sci. Rep. 2020, 10, 1–11. [Google Scholar]
- Koranda, M.; Kaiser, C.; Fuchslueger, L.; Kitzler, B.; Sessitsch, A.; Zechmeister-Boltenstern, S.; Richter, A. Fungal and Bacterial Utilization of Organic Substrates Depends on Substrate Complexity and N Availability. FEMS Microbiol. Ecol. 2014, 87, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Suyal, D.C.; Yadav, A.; Shouche, Y.; Goel, R. Microbial Diversity and Soil Physicochemical Characteristic of Higher Altitude. PLoS ONE 2019, 14, e0213844. [Google Scholar] [CrossRef] [Green Version]
- Siles, J.A.; Cajthaml, T.; Minerbi, S.; Margesin, R. Effect of Altitude and Season on Microbial Ac-tivity, Abundance and Community Structure in Alpine Forest Soils. FEMS Microbiol. Ecol. 2016, 92, fiw008. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, J.; Chen, C.; Li, J.; Chu, J. Grazing Plays an Important Role in Structuring Alpha and Beta Compo-nents of Taxonomic, Functional, and Phylogenetic Diversity in Semiarid Sandy Land of Northern China. Glob. Ecol. Conserv. 2019, 20, e00790. [Google Scholar] [CrossRef]
- Cao, Y.; Wu, J.; Zhang, X.; Niu, B.; Li, M.; Zhang, Y.; Wang, Z. Dynamic Forage-Livestock Balance Analysis in Alpine Grasslands on the Northern Tibetan Plateau. J. Environ. Manag. 2019, 238, 352–359. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Location ID | Latitude (N) | Longitude (E) | Altitude (m) | Location |
|---|---|---|---|---|
| SG a | 38°54.401′ | 99°31.586′ | 2834 | Dahe, Sunan, Gansu, China |
| SG b | 38°54.332′ | 99°31.526′ | 2866 | Dahe, Sunan, Gansu, China |
| SG c | 38°54.251′ | 99°31.457′ | 2907 | Dahe, Sunan, Gansu, China |
| SG d | 38°54.181′ | 99°31.388′ | 2930 | Dahe, Sunan, Gansu, China |
| WG a | 38°55.587′ | 99°32.528′ | 2745 | Dahe, Sunan, Gansu, China |
| WG b | 38°55.151′ | 99°32.465′ | 2756 | Dahe, Sunan, Gansu, China |
| WG c | 38°54.582′ | 99°32.410′ | 2773 | Dahe, Sunan, Gansu, China |
| WG d | 38°54.468′ | 99°32.262′ | 2802 | Dahe, Sunan, Gansu, China |
| Soil Properties | Summer Grazing | Winter Grazing | p-Value |
|---|---|---|---|
| Total Nitrogen (g kg−1) | 4.6 ± 0.29 | 2.8 ± 0.13 | 0.002 |
| Total Carbon (g kg−1) | 45.3 ± 2.78 | 31.3 ± 0.46 | 0.003 |
| Total Sulfur (g kg−1) | 0.4 ± 0.14 | 0.5 ± 0.02 | 0.401 |
| pH | 7.47 ± 0.120 | 7.59 ± 0.055 | 0.372 |
| Organic Carbon (g kg−1) | 41.4 ± 3.18 | 26.9 ± 1.89 | 0.008 |
| C/N | 9.95 ± 0.040 | 11.14 ± 0.356 | 0.016 |
| Water Content (%) | 13.74 ± 2.43 | 7.16 ± 0.748 | 0.042 |
| Classification | Taxonomy | TN | TC | TS | pH | OC | C/N | SWC |
|---|---|---|---|---|---|---|---|---|
| Fungi | Ascomycota | 0.04 | 0.039 | −0.519 | 0.094 | −0.441 | −0.01 | 0.381 |
| Basidiomycota | 0.126 | 0.112 | 0.728 | −0.387 | 0.519 | −0.342 | −0.19 | |
| Mortierellomycota | −0.135 | −0.091 | −0.252 | 0.522 | 0.149 | 0.772 | −0.284 | |
| Glomeromycota | −0.136 | −0.165 | −0.314 | −0.073 | −0.561 | −0.341 | 0.169 | |
| Rozellomycota | 0.05 | 0.027 | 0.693 | −0.397 | 0.396 | −0.421 | −0.228 | |
| Bacteria | Actinobacteria | −0.097 | −0.098 | −0.619 | 0.211 | −0.558 | 0.08 | 0.252 |
| Proteobacteria | 0.209 | 0.206 | 0.722 | −0.346 | 0.637 | −0.218 | −0.136 | |
| Acidobacteria | −0.37 | −0.372 | 0.256 | 0.166 | 0.109 | 0.18 | −0.663 | |
| Chloroflexi | −0.136 | −0.147 | −0.553 | 0.136 | −0.603 | −0.055 | 0.215 | |
| Gemmatimonadetes | 0.223 | 0.241 | −0.465 | 0.113 | −0.184 | 0.158 | 0.498 | |
| Verrucomicrobia | 0.938 * | 0.941 * | 0.488 | −0.723 | 0.67 | −0.555 | 0.990 ** | |
| Planctomycetes | −0.675 | −0.643 | −0.532 | 0.843 | −0.337 | 0.918 * | −0.79 | |
| Bacteroidetes | 0.174 | 0.201 | 0.388 | 0.003 | 0.605 | 0.267 | −0.143 | |
| Rokubacteria | 0.69 | 0.716 | 0.068 | −0.268 | 0.452 | −0.042 | 0.778 | |
| Nitrospirae | −0.448 | −0.406 | −0.536 | 0.78 | −0.192 | 0.940 * | −0.531 |
| Classification | Taxonomy | TN | TC | TS | pH | OC | C/N | SWC |
|---|---|---|---|---|---|---|---|---|
| Fungi | Ascomycota | −0.866 | −0.724 | 0.776 | −0.726 | 0.375 | 0.913 | −0.779 |
| Basidiomycota | 0.701 | 0.599 | −0.573 | 0.498 | −0.539 | −0.744 | 0.849 | |
| Mortierellomycota | 0.092 | −0.006 | 0.075 | 0.098 | −0.879 | −0.165 | 0.867 | |
| Glomeromycota | 0.103 | −0.201 | −0.128 | 0.787 | −0.315 | −0.237 | 0.354 | |
| Bacteria | Actinobacteria | 0.223 | 0.291 | −0.381 | 0.141 | 0.888 | −0.151 | −0.721 |
| Proteobacteria | −0.394 | −0.666 | 0.396 | 0.4 | −0.532 | 0.249 | 0.312 | |
| Acidobacteria | 0.024 | −0.002 | 0.14 | −0.104 | −0.78 | −0.066 | 0.74 | |
| Chloroflexi | −0.399 | −0.144 | 0.45844 | −0.881 | 0.003 | 0.491 | −0.207 | |
| Gemmatimonadetes | −0.665 | −0.479 | 0.543 | −0.679 | 0.648 | 0.748 | −0.936 * | |
| Verrucomicrobia | 0.351 | 0.101 | −0.221 | 0.639 | −0.872 | −0.48 | 0.994 ** | |
| Planctomycetes | −0.678 | −0.879 | 0.687 | 0.066 | −0.532 | 0.554 | 0.171 | |
| Bacteroidetes | −0.651 | −0.846 | 0.625 | 0.105 | −0.362 | 0.537 | 0.025 | |
| Rokubacteria | 0.211 | 0.391 | −0.104 | −0.41 | −0.153 | −0.137 | 0.24 | |
| Nitrospirae | −0.242 | −0.343 | 0.398 | −0.075 | −0.931 * | 0.155 | 0.753 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turatsinze, A.N.; Kang, B.; Zhu, T.; Hou, F.; Bowatte, S. Soil Bacterial and Fungal Composition and Diversity Responses to Seasonal Deer Grazing in a Subalpine Meadow. Diversity 2021, 13, 84. https://doi.org/10.3390/d13020084
Turatsinze AN, Kang B, Zhu T, Hou F, Bowatte S. Soil Bacterial and Fungal Composition and Diversity Responses to Seasonal Deer Grazing in a Subalpine Meadow. Diversity. 2021; 13(2):84. https://doi.org/10.3390/d13020084
Chicago/Turabian StyleTuratsinze, Andéole Niyongabo, Baotian Kang, Tianqi Zhu, Fujiang Hou, and Saman Bowatte. 2021. "Soil Bacterial and Fungal Composition and Diversity Responses to Seasonal Deer Grazing in a Subalpine Meadow" Diversity 13, no. 2: 84. https://doi.org/10.3390/d13020084
APA StyleTuratsinze, A. N., Kang, B., Zhu, T., Hou, F., & Bowatte, S. (2021). Soil Bacterial and Fungal Composition and Diversity Responses to Seasonal Deer Grazing in a Subalpine Meadow. Diversity, 13(2), 84. https://doi.org/10.3390/d13020084