
On the Systematics and Biodiversity of the Opheliidae and Scalibregmatidae



Abstract
:1. Introduction
2. Methods
3. Results
3.1. Opheliidae Malmgren, 1867
3.1.1. Systematics
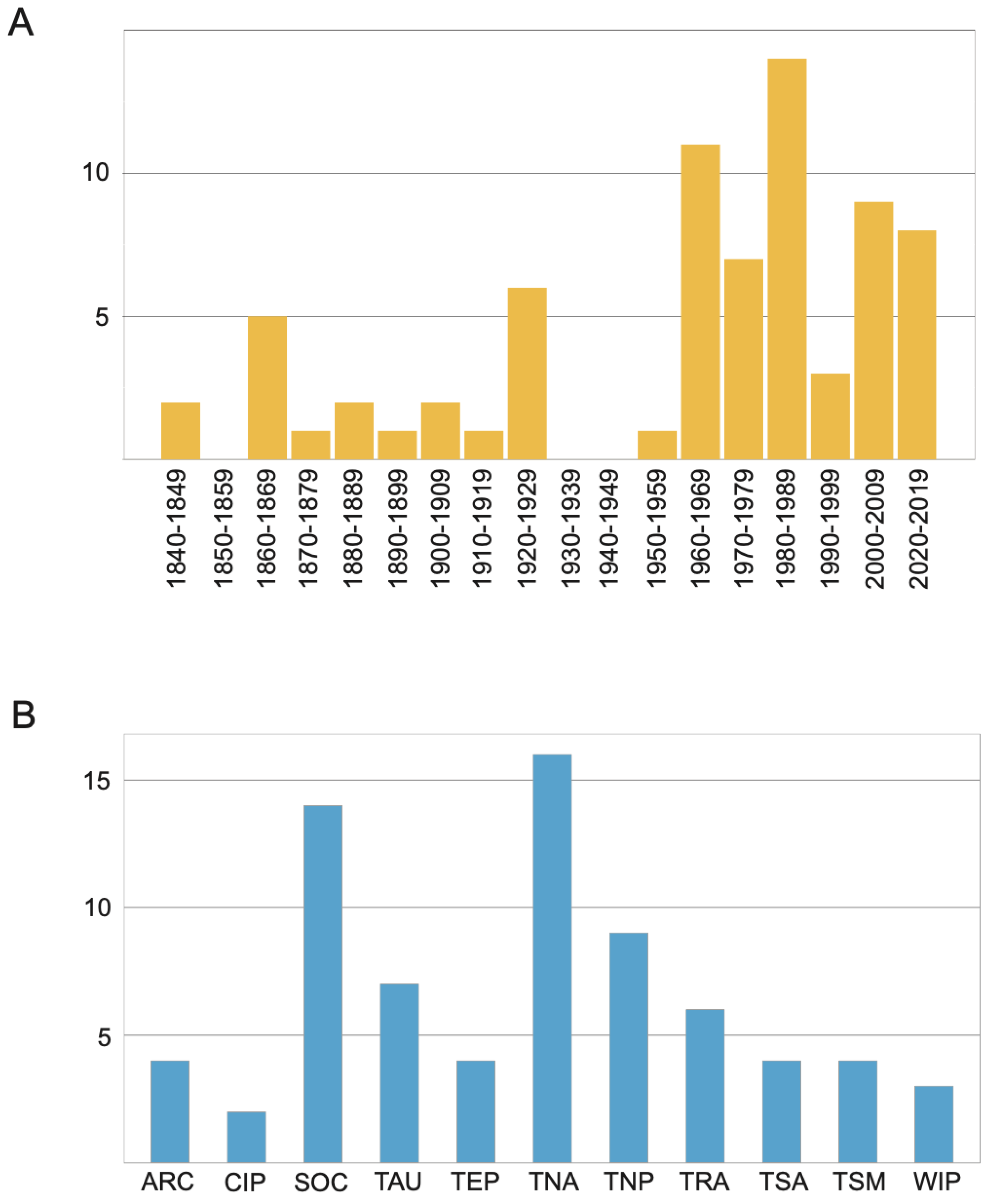
3.1.2. Taxonomic History
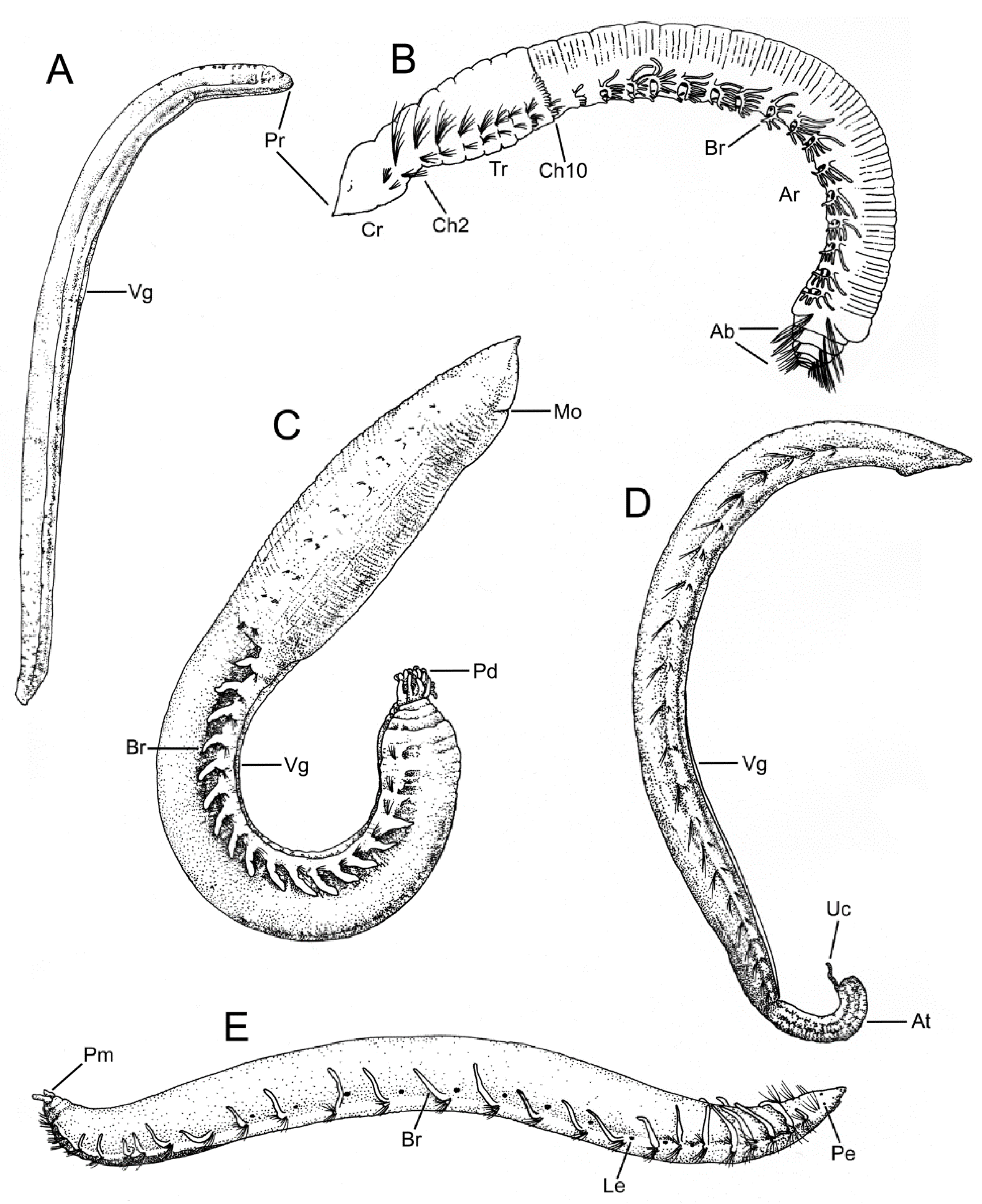
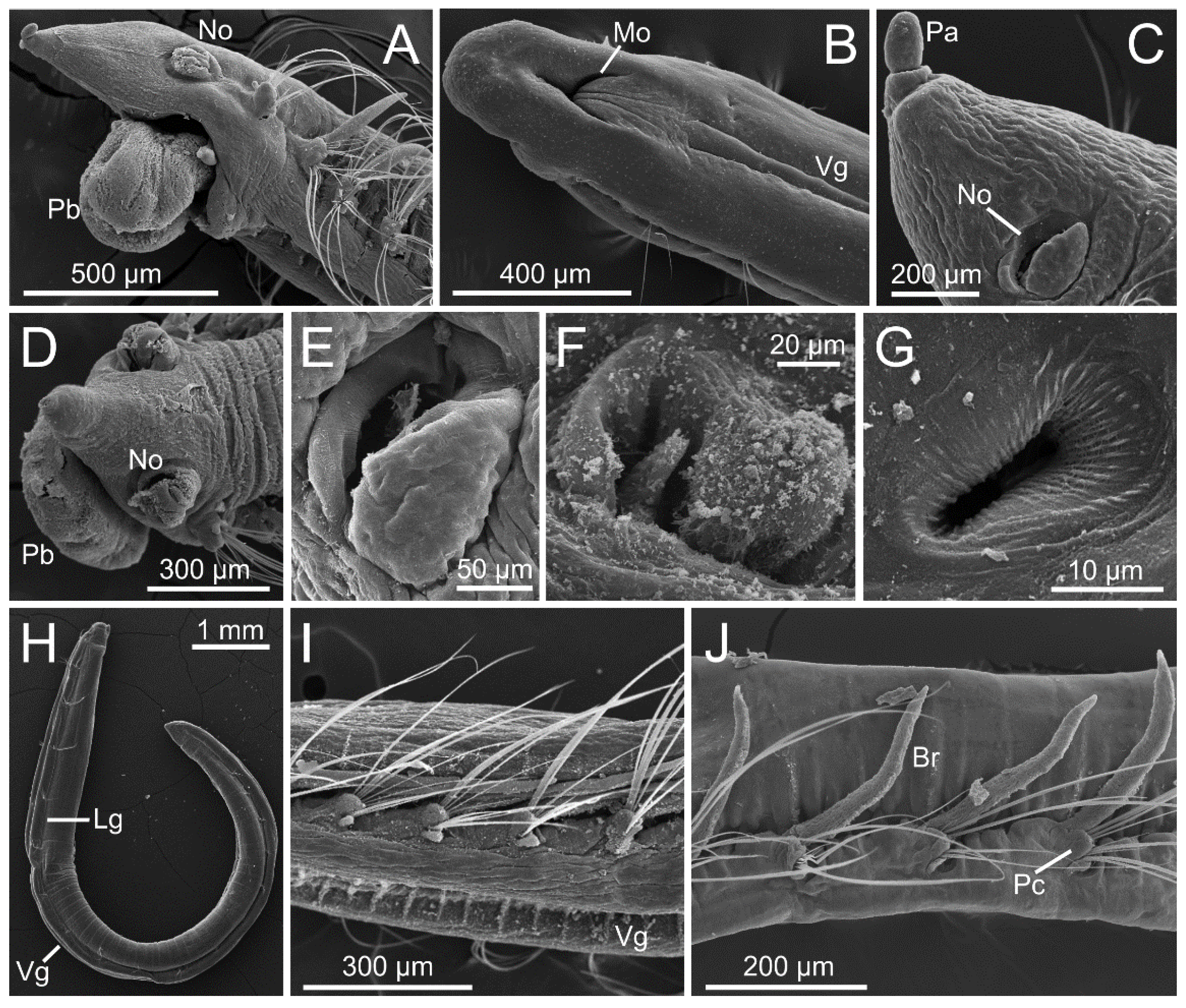
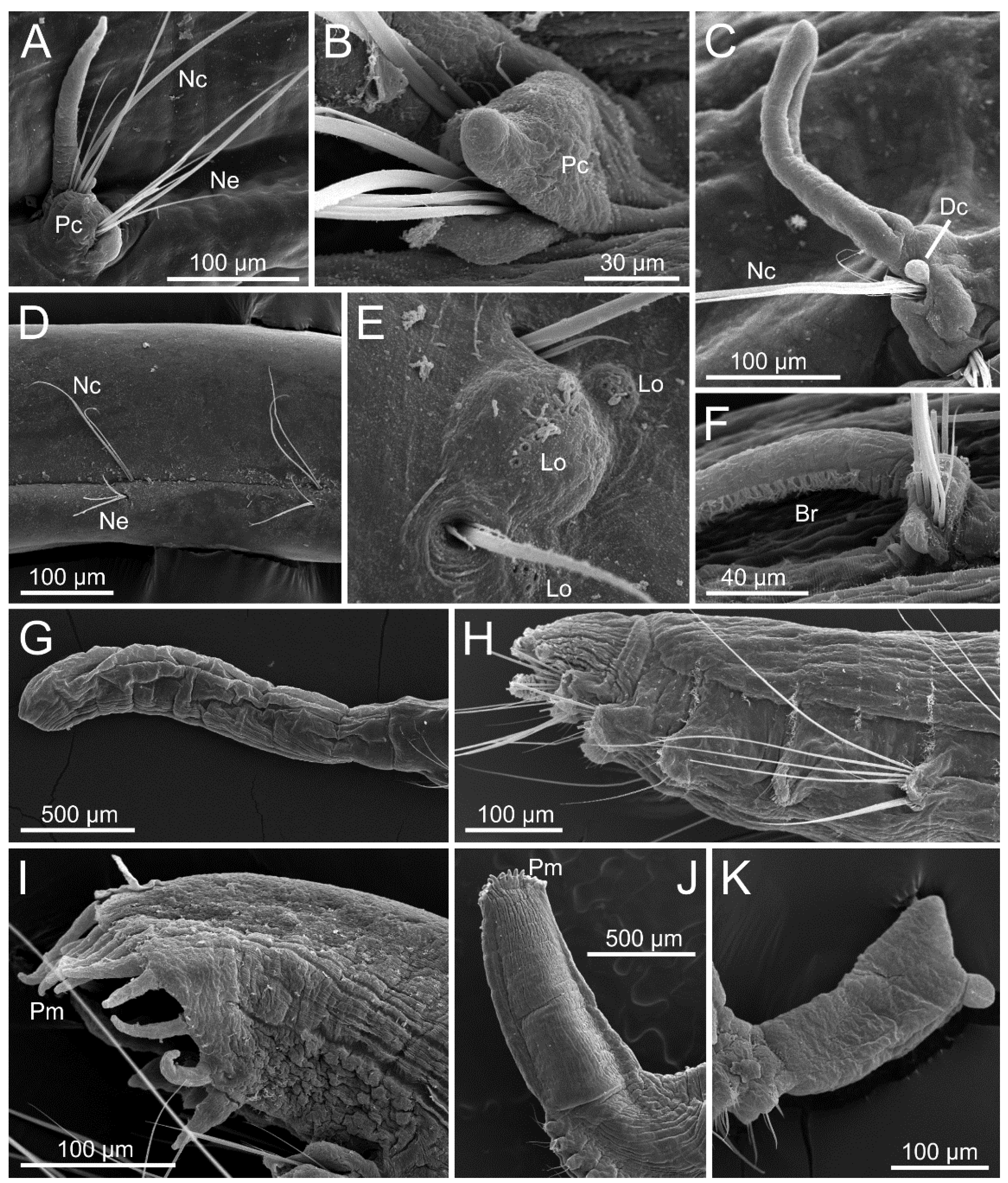
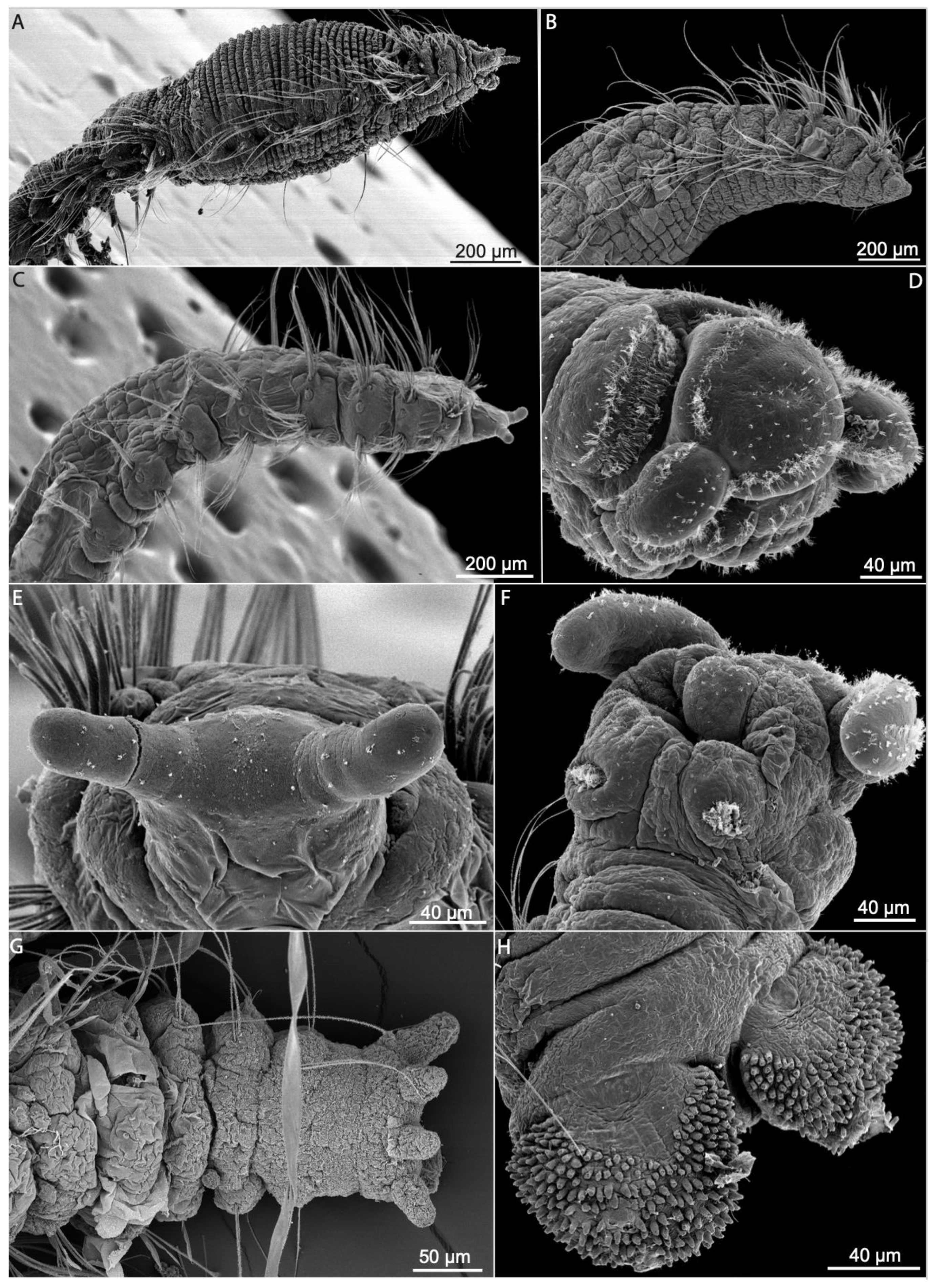
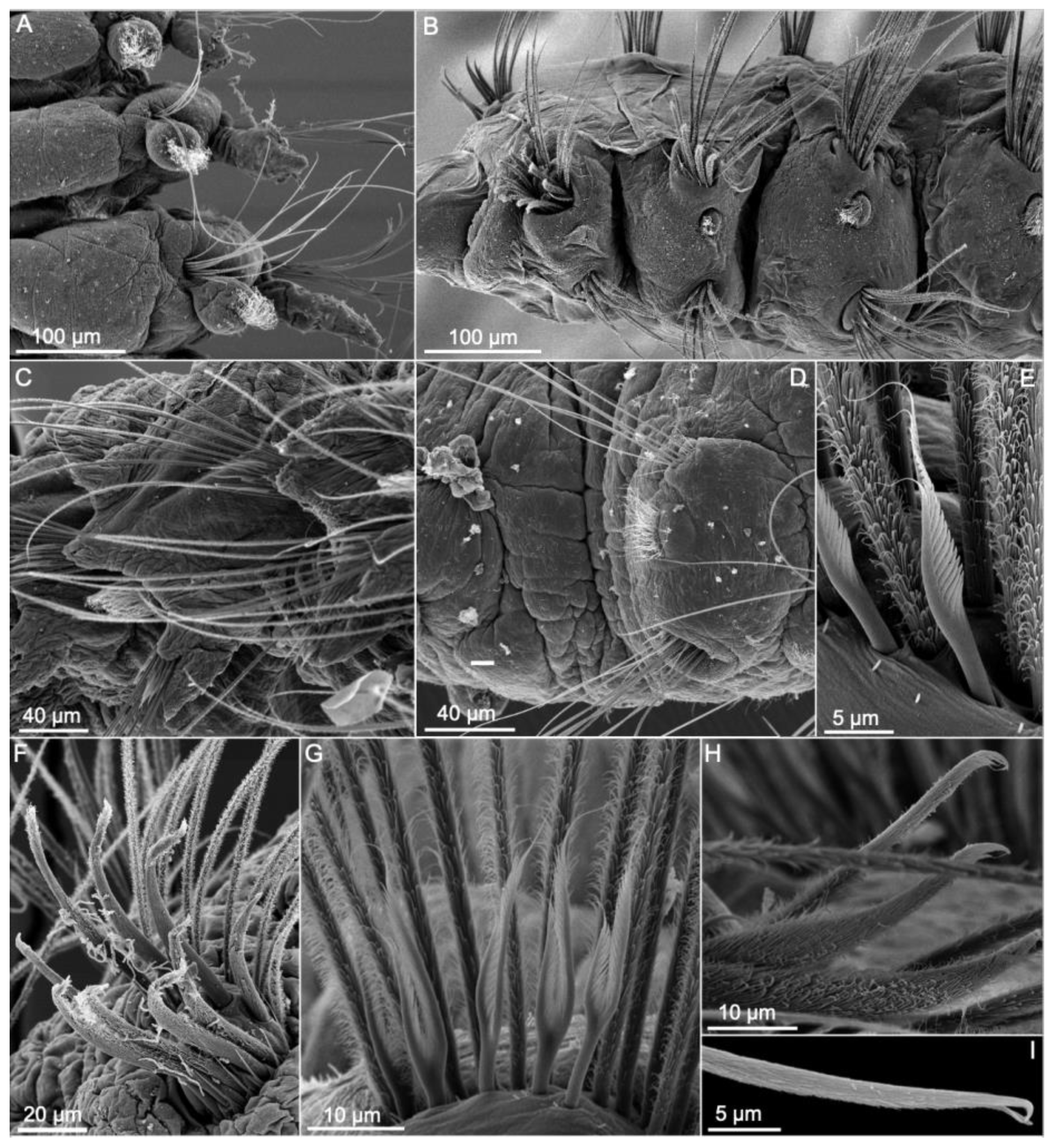
3.1.3. Taxonomic Characters and External Morphology
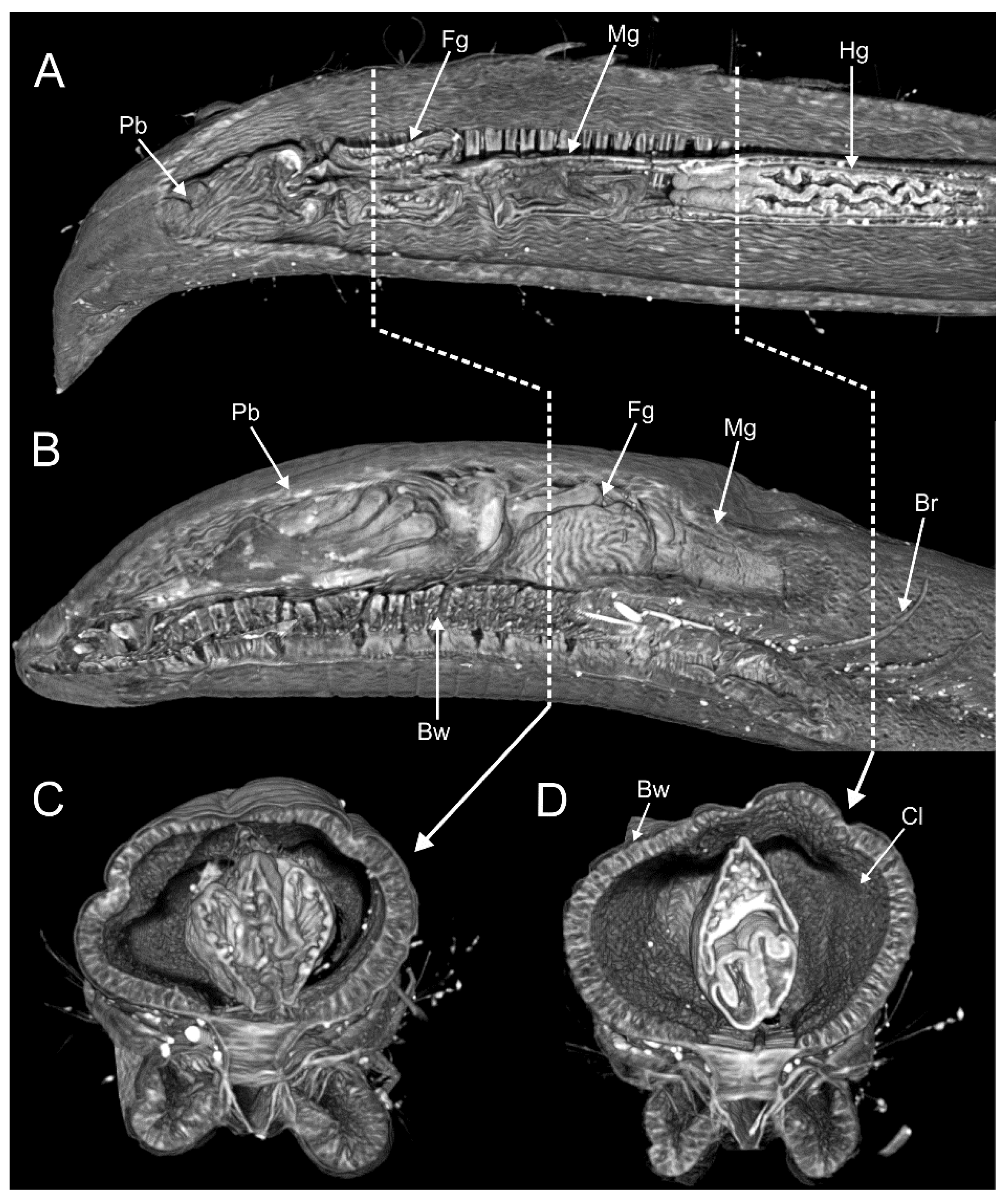
3.1.4. Internal Morphology
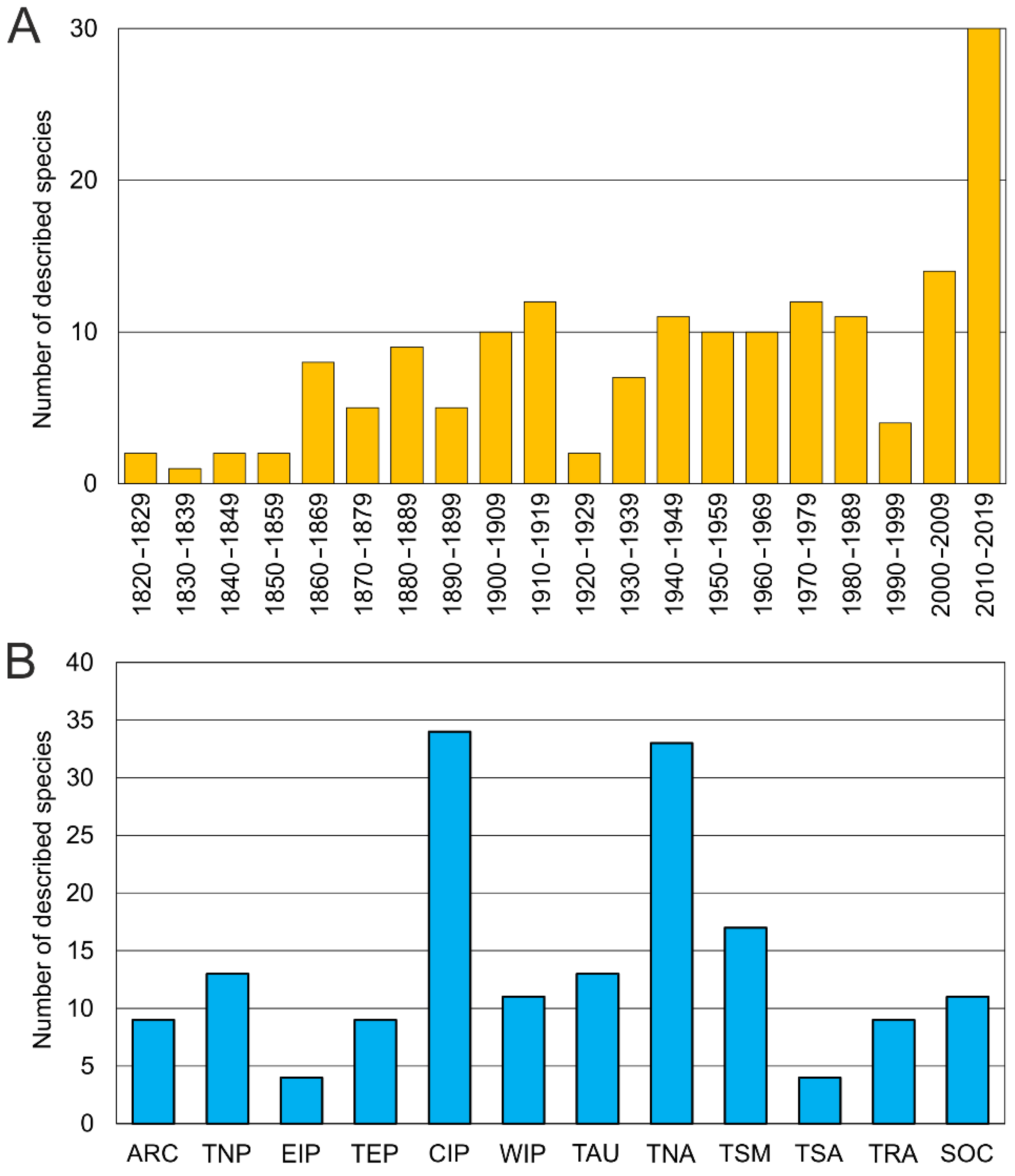
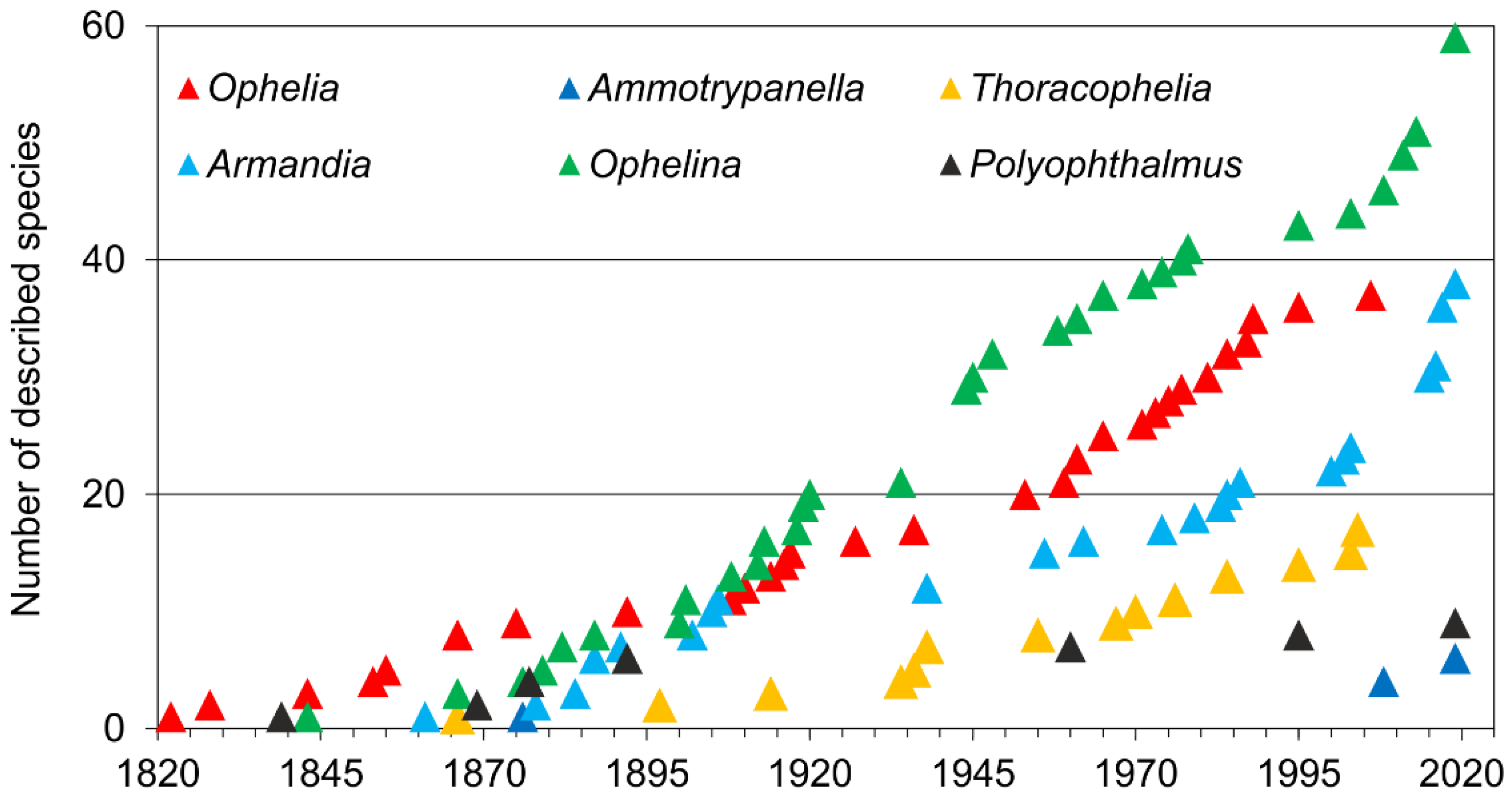
3.1.5. Species Diversity and Distribution
3.1.6. Biology and Ecology
3.2. Scalibregmatidae Malmgren, 1867
3.2.1. Systematics
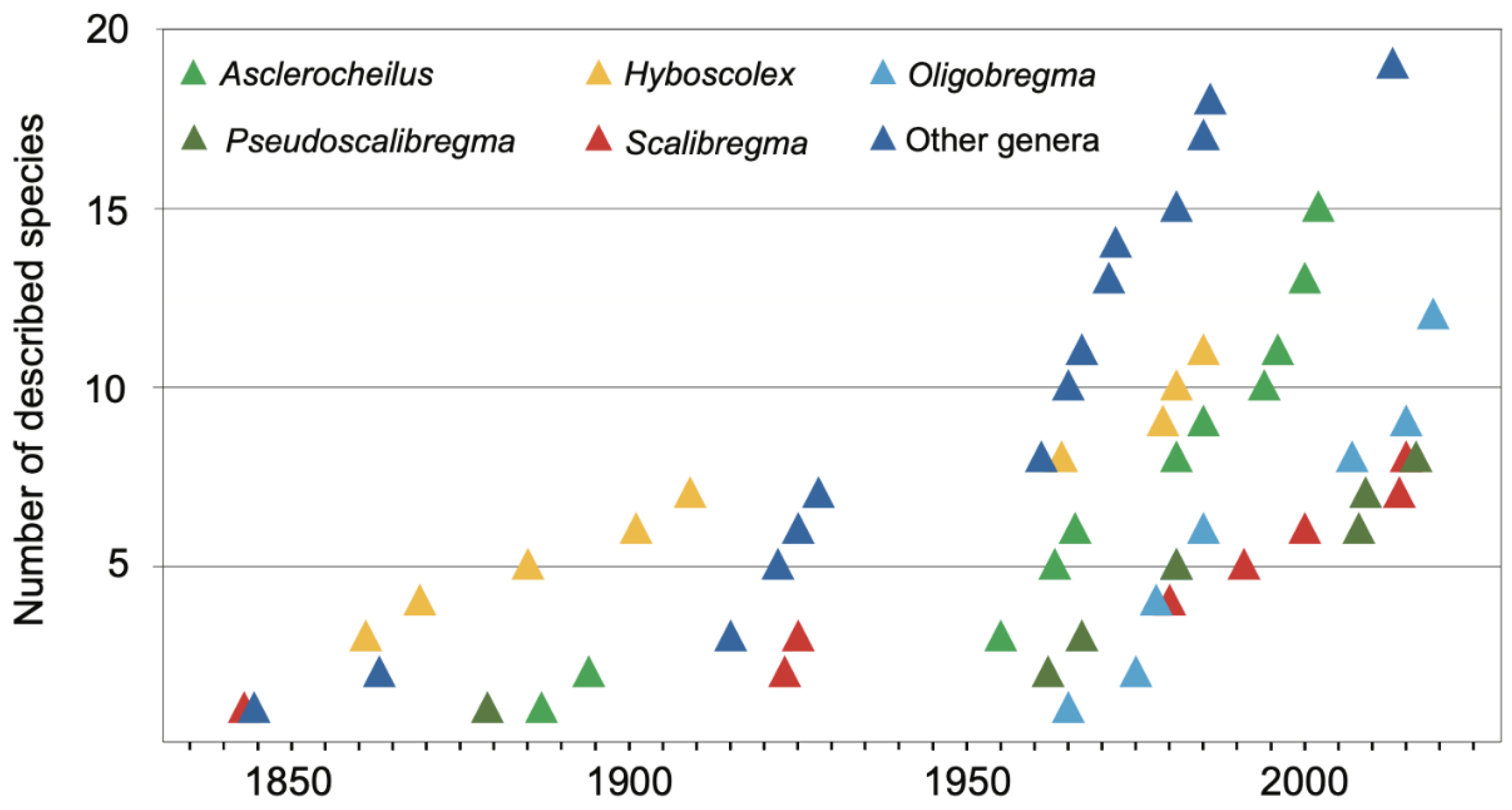
3.2.2. Taxonomic History
3.2.3. Taxonomic Characters and External Morphology
3.2.4. Internal Morphology
3.2.5. Species Diversity and Distribution
3.2.6. Biology and Ecology
4. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Type Locality | Realms | Depth (m) |
|---|---|---|---|
| Ammotrypanella arctica (McIntosh, 1879) | North Atlantic Ocean | Temperate Northern Atlantic | 2014–5023 |
| Ammotrypanella cirrosa Schüller, 2008 | Antarctic Peninsula; Weddell Sea | Southern Ocean | 2014–4817 |
| Ammotrypanella keenani Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 404–4302 |
| Ammotrypanella kersteni Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 4128 |
| Ammotrypanella mcintoshi Schüller, 2008 | off South Africa | Southern Ocean | 1047–4720 |
| Ammotrypanella princessa Schüller, 2008 | Antarctic Peninsula, Weddell Sea | Southern Ocean | 2014–4720 |
| Armandia agilis (Andrews, 1891) | North Carolina, USA | Temperate Northern Atlantic | N.d. |
| Armandia amakusaensis Saito, Tamaki and Imajima, 2000 | Western Kyushu, Japan | Temperate Northern Pacific | Intertidal |
| Armandia andamana Eibye-Jacobsen, 2002 | Andaman Sea | Central Indo-Pacific | 42–63 |
| Armandia bifida Parapar and Moreira, 2015 | Lizard Island, Australia | Central Indo-Pacific | Intertidal |
| Armandia bilobata Hartmann-Schröder, 1986 | South Australia | Temperate Australasia | Intertidal |
| Armandia bipapillata Hartmann-Schröder, 1974 | Inhambane, Mozambique | Western Indo-Pacific | Intertidal |
| Armandia brevis (Moore, 1906) | Icy Cape, Alaska, USA | Arctic | Intertidal to shallow subtidal |
| Armandia broomensis Hartmann-Schröder, 1979 | Broome, Australia | Central Indo-Pacific | Intertidal |
| Armandia buccina Moreira and Parapar, 2017 | Lizard Island, Australia | Central Indo-Pacific | 0–12 |
| Armandia casuarina Moreira and Parapar, 2017 | Lizard Island, Australia | Central Indo-Pacific | Intertidal |
| Armandia circumpapillata Magalhães, Rizzo and Bailey-Brock, 2019 | Oahu, Hawaii | Eastern Indo-Pacific | Shallow subtidal to 82 m |
| Armandia cirrhosa Filippi, 1861 | Cagliari, Sardinia, Mediterranean | Temperate Northern Atlantic | Shallow subtidal |
| Armandia dolio Parapar and Moreira, 2015 | Lizard Island, Australia | Central Indo-Pacific | 5–14 |
| Armandia exigua Kükenthal, 1887 | China | Central Indo-Pacific | 15 |
| Armandia filibranchia Parapar and Moreira, 2015 | Lizard Island, Australia | Central Indo-Pacific | 9 |
| Armandia garretti Magalhães, Rizzo and Bailey-Brock, 2019 | Pearl Harbor, Hawaii | Eastern Indo-Pacific | 0–20 |
| Armandia hossfeldi Hartmann-Schröder, 1956 | Brazil | Temperate South America | 9.5–14 |
| Armandia ilhabelae Hartmann-Schröder, 1956 | Santos, Brazil | Temperate South America | Intertidal |
| Armandia intermedia Fauvel, 1902 | River Casamance estuary, Senegal | Tropical Atlantic | Intertidal to subtidal |
| Armandia laminosa Parapar and Moreira, 2015 | Lizard Island, Australia | Central Indo-Pacific | 0–15 |
| Armandia lanceolata Willey, 1905 | South of Manaar Island, Sri Lanka | Western Indo-Pacific | 15–16 |
| Armandia leptocirris (Grube, 1878) | Philippines | Central Indo-Pacific | 0–18 |
| Armandia loboi Elías and Bremec, 2003 | off Mar del Plata, Argentina | Temperate South America | 5–13 |
| Armandia maculata (Webster, 1884) | Bermuda | Tropical Atlantic | 9–38 |
| Armandia mariacapae Moreira and Parapar, 2017 | Lizard Island, Australia | Central Indo-Pacific | 0–12 |
| Armandia melanura Gravier, 1905 | Djibouti, Gulf of Aden | Western Indo-Pacific | Intertidal |
| Armandia nonpapillata Jones, 1962 | Kingston Harbour, Jamaica | Tropical Atlantic | N.d. |
| Armandia opisthoculata Moreira and Parapar, 2017 | Lizard Island, Australia | Central Indo-Pacific | 5–15 |
| Armandia paraintermedia Parapar and Moreira, 2015 | Lizard Island, Australia | Central Indo-Pacific | 0–24 |
| Armandia parva Moreira and Parapar, 2017 | Lizard Island, Australia | Central Indo-Pacific | 0–15 |
| Armandia polyophthalma Kükenthal, 1887 | Gulf of Naples, Italy | Temperate Northern Atlantic | 0–20 |
| Armandia salvadoriana Hartmann-Schröder, 1956 | El Salvador | Tropical Eastern Pacific | Intertidal |
| Armandia sampadae Gopal, Jaleel, Parameswaran and Vijayan, 2016 | Andaman Islands | Western Indo-Pacific | 52–57 |
| Armandia secundariopapillata Hartmann-Schröder, 1984 | SW Australia | Temperate Australasia | Intertidal |
| Armandia sinaitica Amoureux, 1983 | Gulf of Akaba, Red Sea | Western Indo-Pacific | Intertidal |
| Armandia simodaensis Takahashi, 1938 | Japan | Temperate Northern Pacific | N.d. |
| Armandia tubulata Parapar and Moreira, 2015 | Lizard Island, Australia | Central Indo-Pacific | 1–10 |
| Armandia weissenbornii Kükenthal, 1887 | Perim Island, Red Sea | Western Indo-Pacific | N.d. |
| Ophelia africana Tebble, 1953 | Table Bay, South Africa | Temperate Southern Africa | Intertidal to shallow subtidal |
| Ophelia agulhana Day, 1961 | False Bay, South Africa | Temperate Southern Africa | Intertidal to shallow subtidal |
| Ophelia algida Maciolek and Blake, 2006 | Off Macquarie Island, Southern Ocean | Southern Ocean | 112–124 |
| Ophelia amoureuxi Bellan and Costa, 1988 | Côte d’Azur | Temperate Northern Atlantic | 90–95 |
| Ophelia anomala Day, 1961 | False Bay, South Africa | Temperate Southern Africa | 15–80 |
| Ophelia ashworthi Fauvel, 1917 | Gulf of St Vincent and Spencer | Temperate Australasia | Subtidal |
| Ophelia assimilis Tebble, 1953 | Central California | Temperate Northern Pacific | Intertidal to shallow subtidal |
| Ophelia barquii Fauvel, 1927 | Agay, Var, France | Temperate Northern Atlantic | Intertidal |
| Ophelia bicornis Savigny, 1822 | La Rochelle, France | Temperate Northern Atlantic | Intertidal to subtidal |
| Ophelia bipartita Monro, 1936 | S Chile | Temperate South America | 35 |
| Ophelia borealis Quatrefages, 1866 | Greenland | Arctic | N.d. |
| Ophelia bulbibranchiata Hartmann-Schröder and Parker, 1995 | Pearson Island, Australia | Temperate Australasia | not recorded |
| Ophelia capensis Kirkegaard, 1959 | Table Bay, South Africa | Temperate Southern Africa | 50–80 |
| Ophelia celtica Amoureux and Dauvin, 1981 | Atlantic France | Temperate Northern Atlantic | <100 |
| Ophelia dannevigi Benham, 1916 | St. Francis Island, Australia | Temperate Australasia | ~60 |
| Ophelia denticulata Verrill, 1875 | Maine, USA | Temperate Northern Atlantic | N.d. |
| Ophelia elongata Hutchings and Murray, 1984 | Burwood Beach, Australia | Temperate Australasia | Subtidal |
| Ophelia formosa (Kinberg, 1866) | La Plata, Argentina (?) | Temperate South America | N.d. |
| Ophelia glabra Stimpson, 1853 | East Canada | Temperate Northern Atlantic | N.d. |
| Ophelia kirkegaardi Intes and Le Loeuff, 1977 | Off Abidjan, Ivory Coast | Tropical Atlantic | 20–40 |
| Ophelia koloana Gibbs, 1971 | Solomon Islands | Central Indo-Pacific | 2 |
| Ophelia laubieri Bellan and Costa, 1988 | Estuârio do Sado, Portugal | Temperate Northern Atlantic | Low intertidal to subtidal |
| Ophelia limacina (Rathke, 1843) | Norway | Temperate Northern Atlantic | 0–500 |
| Ophelia magna (Treadwell, 1914) | California, USA | Temperate Northern Pacific | N.d. |
| Ophelia multibranchia Hutchings and Murray, 1984 | Botany Bay, Australia | Temperate Australasia | 3–4 |
| Ophelia neglecta Schneider, 1892 | France | Temperate Northern Atlantic | Intertidal to subtidal |
| Ophelia peresi Bellan and Picard, 1965 | Tuléar, Madagascar | Western Indo-Pacific | 15–20 |
| Ophelia praetiosa (Kinberg, 1866) | Cabo Vírgenes, Argentina | Temperate South America | 13–95 |
| Ophelia profunda Hartman, 1965 | Off New England, USA | Temperate Northern Atlantic | 1000–1700 |
| Ophelia pulchella Tebble, 1953 | Southern California, USA | Temperate Northern Pacific | Subtidal |
| Ophelia radiata (Delle Chiaje, 1828) | Gulf of Naples, Italy | Temperate Northern Atlantic | Intertidal to subtidal |
| Ophelia rathkei McIntosh, 1908 | UK | Temperate Northern Atlantic | Intertidal to subtidal |
| Ophelia roscoffensis Augener, 1910 | Roscoff, France | Temperate Northern Atlantic | Subtidal |
| Ophelia rullieri Bellan, 1975 | Gaspésie, Quebec | Temperate Northern Atlantic | Intertidal to shallow subtidal |
| Ophelia simplex Leidy, 1855 | Rhode Island, USA | Temperate Northern Atlantic | N.d. |
| Ophelia translucens (Katzmann, 1973) | Zlarin, Croatia | Temperate Northern Atlantic | 20–40 |
| Ophelia verrilli Riser, 1987 | New England, USA | Temperate Northern Atlantic | Intertidal to shallow subtidal |
| Ophelina abranchiata Støp-Bowitz, 1948 | Greenland | Arctic | 90–4500 |
| Ophelina acuminata Örsted, 1843 | Øresund Strait, Hveen Island, Sweden | Temperate Northern Atlantic | Shelf and slope |
| Ophelina adamantea (Kinberg, 1866) | Rio de Janeiro, Brazil | Temperate South America | N.d. |
| Ophelina alata Elías, Bremec, Lana and Orensanz, 2003 | SE Brazil | Temperate South America | Subtidal |
| Ophelina ammotrypanella Schüller, 2008 | Antarctic peninsula, Weddell Sea | Southern Ocean | 1970–3050 |
| Ophelina aulogastrella (Hartman and Fauchald, 1971) | off New England, USA | Temperate Northern Atlantic | 196–5023 |
| Ophelina basicirra Parapar, Moreira and Helgason, 2011 | NW Iceland | Temperate Northern Atlantic | 23–2298 |
| Ophelina bimensis (Caullery, 1944) | Indonesia | Central Indo-Pacific | N.d. |
| Ophelina bowitzi Parapar, Moreira and Helgason, 2011 | Southern Iceland | Temperate Northern Atlantic | 1897–2709 |
| Ophelina brasiliensis Hansen, 1882 | Brazil | Temperate South America | Subtidal |
| Ophelina brattegardi Kongsrud, Bakken and Oug, 2011 | off East Greenland | Arctic | 1600 |
| Ophelina breviata (Ehlers, 1913) | Wilhelm II Coast, Antarctic Ocean | Southern Ocean | 20–3000 |
| Ophelina brevibranchiata (Caullery, 1944) | Indonesia | Central Indo-Pacific | N.d. |
| Ophelina buitendijki (Horst, 1919) | Java Sea | Central Indo-Pacific | N.d. |
| Ophelina chaetifera (Hartman, 1965) | off New England, USA | Temperate Northern Atlantic | 1330–5007 |
| Ophelina cordiformis (Caullery, 1944) | Indonesia | Central Indo-Pacific | N.d. |
| Ophelina curli Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 4026 |
| Ophelina cylindricaudata (Hansen, 1879) | off West Norway | Temperate Northern Atlantic | 7–4663 |
| Ophelina cyprophilia Neave and Glasby, 2013 | Darwin Harbour, Australia | Central Indo-Pacific | 0–10 |
| Ophelina delapidans (Kinberg, 1866) | Valparaiso, Chile | Temperate South America | Shallow subtidal |
| Ophelina dubia (Caullery, 1944) | Indonesia | Central Indo-Pacific | N.d. |
| Ophelina ehlersi (Horst, 1919) | Jedan, Aroe Isles, Indonesia | Central Indo-Pacific | N.d. |
| Ophelina fauveli (Caullery, 1944) | Indonesia | Central Indo-Pacific | 10–21 |
| Ophelina ganae Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 4076–4302 |
| Ophelina gaucha Elías, Bremec, Lana and Orensanz, 2003 | SE Brazil | Temperate South America | 0–18 |
| Ophelina hachaensis Augener, 1934 | Riohacha, Colombia | Tropical Atlantic | 6 |
| Ophelina helgolandiae Augener, 1912 | Spitsbergen, Norway | Arctic | 562–2710 |
| Ophelina gigantea (Rullier, 1965) | Moreton Bay, Australia | Temperate Australasia | N.d. |
| Ophelina grandis (Pillai, 1961) | Tambalagam Bay, Sri Lanka | Western Indo-Pacific | 1.8–7.3 |
| Ophelina groenlandica Støp-Bowitz, 1948 | East Greenland | Arctic | Shelf and slope |
| Ophelina gymnopyge (Ehlers, 1908) | Kerguelen Islands | Southern Ocean | 13–199 |
| Ophelina jeffreysi (McIntosh in Jeffreys, 1876) | Labrador Sea, North Atlantic Ocean | Arctic | 1066–3200 |
| Ophelina juhazi Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 4100 |
| Ophelina kampeni (Horst, 1919) | Jedan, Aroe Isles, Indonesia | Central Indo-Pacific | N.d. |
| Ophelina kinbergii Hansen, 1882 | Río de Janeiro, Brazil | Temperate South America | Subtidal |
| Ophelina kohni Magalhães, Rizzo and Bailey-Brock, 2019 | Guam | Central Indo-Pacific | Shallow subtidal |
| Ophelina kuekenthali (McIntosh, 1908) | North Atlantic Ocean | Temperate Northern Atlantic | ~1400 |
| Ophelina langii (Kükenthal, 1887) | Philippines | Central Indo-Pacific | 20 |
| Ophelina longicaudata (Caullery, 1944) | Indonesia | Central Indo-Pacific | N.d. |
| Ophelina longicephala Hartmann-Schröder, 1977 | Off Portugal | Temperate Northern Atlantic | 77 |
| Ophelina longicirrata Hartmann-Schröder and Parker, 1995 | South Australia | Temperate Australasia | N.d. |
| Ophelina manana Magalhães, Rizzo and Bailey-Brock, 2019 | Oahu, Hawaii | Eastern Indo-Pacific | 400–500 |
| Ophelina martinezarbizui Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 4026–4425 |
| Ophelina meyerae Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 4302 |
| Ophelina minima Hartmann-Schröder, 1974 | Skagerrak | Temperate Northern Atlantic | 230–645 |
| Ophelina modesta Støp-Bowitz, 1958 | Oslo, Norway | Temperate Northern Atlantic | 100–200 |
| Ophelina nematoides (Ehlers, 1913) | Antarctic Ocean | Southern Ocean | 246–2725 |
| Ophelina norvegica Støp-Bowitz, 1945 | East Norway | Temperate Northern Atlantic | Subtidal |
| Ophelina nunnallyi Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 4425–4302 |
| Ophelina nybelini (Eliason, 1951) | Azores Islands | Temperate Northern Atlantic | 4540–4600 |
| Ophelina opisthobranchiata Wirén, 1901 | Spitsbergen, Norway | Arctic | 800–3900 |
| Ophelina profunda (Caullery, 1944) | Indonesia | Central Indo-Pacific | N.d. |
| Ophelina pygocirrata (Ehlers, 1920) | Indonesia | Central Indo-Pacific | N.d. |
| Ophelina remigera (Ehlers, 1918) | Aru Islands, Sungi Manumbai, Indonesia | Central Indo-Pacific | N.d. |
| Ophelina robusta Schüller, 2008 | Antarctic Peninsula, Weddell Sea | Southern Ocean | 2668–3050 |
| Ophelina scaphigera (Ehlers, 1900) | Magellan Strait | Temperate South America | 18–3382 |
| Ophelina setigera (Hartman, 1978) | Weddell Sea | Southern Ocean | 3111 |
| Ophelina sibogae (Caullery, 1944) | Java | Central Indo-Pacific | 79–81 |
| Ophelina syringopyge (Ehlers, 1901) | South Georgia | Southern Ocean | 9–876 |
| Ophelina tessellata Neave and Glasby, 2013 | Melville Bay, Australia | Central Indo-Pacific | 0–10 |
| Polyophthalmus australis Grube, 1869 | Cape York, Australia | Central Indo-Pacific | N.d. |
| Polyophthalmus ceylonensis Kükenthal, 1887 | Sri Lanka | Western Indo-Pacific | N.d. |
| Polyophthalmus collaris Michaelsen, 1892 | Sri Lanka | Western Indo-Pacific | N.d. |
| Polyophthalmus longisetosus Michaelsen, 1892 | Sri Lanka | Western Indo-Pacific | N.d. |
| Polyophthalmus mauliola Magalhães, Rizzo and Bailey-Brock, 2019 | Mamala Bay, Hawaii | Eastern Indo-Pacific | 27–56 |
| Polyophthalmus pictus (Dujardin, 1839) | France | Temperate Northern Atlantic | Intertidal |
| Polyophthalmus qingdaoensis Purschke, Ding and Müller, 1995 | Qingdao, Yellow Sea | Temperate Northern Pacific | N.d. |
| Polyophthalmus striatus Kükenthal, 1887 | Hong Kong | Central Indo-Pacific | N.d. |
| Polyophthalmus translucens Hartman, 1960 | Southern California, USA | Temperate Northern Pacific | 914 |
| Thoracophelia arctica (Grube, 1866) | Arctic Ocean | Arctic | N.d. |
| Thoracophelia bibrancha (Hutchings and Murray, 1984) | Merimbula, Australia | Temperate Australasia | Intertidal |
| Thoracophelia dillonensis (Hartman, 1938) | Dillon Beach, California, USA | Temperate Northern Pacific | Intertidal |
| Thoracophelia ezoensis Okuda, 1936 | Hokkaido, Japan | Temperate Northern Pacific | N.d. |
| Thoracophelia flabellifera Ziegelmeier, 1955 | German Bight, North Sea | Temperate Northern Atlantic | 13 |
| Thoracophelia furcifera Ehlers, 1897 | Punta Arenas, Magellan Strait | Temperate South America | Intertidal |
| Thoracophelia heterocirra (Rozbaczylo and Zamorano, 1970) | El Tabo, Chile | Temperate South America | Intertidal |
| Thoracophelia japonica (Misaka and Sato, 2003) | Oura Bay, Japan | Temperate Northern Pacific | 0–16 |
| Thoracophelia longiseta (Hutchings and Murray, 1984) | Ocean Beach, Australia | Temperate Australasia | Intertidal |
| Thoracophelia mammillata (Santos, Nonato and Petersen, 2004) | Rio de Janeiro, Brazil | Temperate South America | 22–45 |
| Thoracophelia mucronata (Treadwell, 1914) | La Jolla, Southern California | Temperate Northern Pacific | Intertidal |
| Thoracophelia otagoensis (Probert, 1976) | Otago Peninsula, New Zealand | Temperate Australasia | Intertidal |
| Thoracophelia papillata (Santos, Nonato and Petersen, 2004) | Abaís beach, Brazil | Tropical Atlantic | Intertidal |
| Thoracophelia profunda (Hartman, 1967) | Cape Horn, Chile | Temperate South America | 4008 |
| Thoracophelia yasudai Okuda, 1934 | Kainawa, Japan | Temperate Northern Pacific | Intertidal |
| Thoracophelia williamsi (Hartman, 1938) | Dillon Beach, California, USA | Temperate Northern Pacific | Intertidal |
| Thoracophelia zeidleri (Hartmann-Schröder and Parker, 1995) | Haystack Beach, Australia | Temperate Australasia | Intertidal |
| Species | Type Locality | Realms | Depth (m) |
|---|---|---|---|
| Asclerocheilus acirratus (Hartman, 1966) | White Cove, Southern California, USA | Temperate Northern Pacific | 0–3 |
| Asclerocheilus ashworthi Blake, 1981 | Elephant Island, Antarctica | Southern Ocean | 223–397 |
| Asclerocheilus beringianus Uschakov, 1955 | Bering Sea | Arctic | 986–2005 |
| Asclerocheilus californicus Hartman, 1963 | Santa Monica, Redondo and San Pedro valley, California, USA | Temperate Northern Pacific | 542–890 |
| Asclerocheilus capensis Day, 1963 | South Africa | Temperate Southern Africa | 9–26 |
| Asclerocheilus elisabethae Eibye-Jacobsen, 2002 | Thailand, Andaman Sea | Western Indo-Pacific | 70–76 |
| Asclerocheilus glabrus (Ehlers, 1887) | Cuba | Tropical Atlantic | 320 |
| Asclerocheilus intermedius (Saint-Joseph, 1894) | Dinard, France | Temperate Northern Atlantic | 96–1830 |
| Asclerocheilus kudenovi Blake, 2000 | Point Arguello, California, USA | Temperate Northern Pacific | 91.5–123 |
| Asclerocheilus mexicanus Kudenov, 1985 | Florida, Gulf of Mexico | Tropical Atlantic | 2.4–76.2 |
| Asclerocheilus shanei Hartmann-Schröder, 1994 | Scamander, Tasmania | Temperate Australia | 122 |
| Asclerocheilus shanonae Eibye-Jacobsen, 2002 | Thailand, Andaman Sea | Western Indo-Pacific | 70–76 |
| Asclerocheilus tasmanicus Kirkegaard, 1996 | Tasman Sea, W of New Zealand | Temperate Australia | 3710–3830 |
| Asclerocheilus tropicus Blake, 1981 | NW off Guayaquil, Ecuador | Tropical Eastern Pacific | 3–10 |
| Asclerocheilus victoriensis Blake, 2000 | Victoria, Australia | Temperate Australia | 6–22 |
| Axiokebuita cavernicola Martínez, Di Domenico and Worsaae, 2013 | Los Cerebros cave, Tenerife, Canary Islands | Temperate Northern Atlantic | 8–15 |
| Axiokebuita minuta (Hartman, 1967) | Antarctica | Southern Ocean | 180–3685 |
| Cryptosclerocheilus baffinensis Blake, 1972 | Southern Baffin Bay | Arctic | 1830 |
| Hyboscolex dicranochaetus (Schmarda, 1861) | Table Bay, Cape New Hope, South Africa | Temperate Southern Africa | 0.5–19.8 |
| Hyboscolex equatorialis Blake, 1981 | NW of Guayaquil, Ecuador | Temperate Southern America | 8–9 |
| Hyboscolex homochaetus (Schmarda, 1861) | New Zealand | Temperate Southern Africa | N.d. |
| Hyboscolex longisetus Schmarda, 1861 | Table Bay, Cape New Hope, South Africa | Temperate Southern Africa | 9–110 |
| Hyboscolex oculatus (Ehlers, 1901) | Tumbes Peninsula, near Talcahuano, Chile | Temperate Southern America | ca. 0.5–10 |
| Hyboscolex pacificus (Moore, 1909) | Santa Monica, California, USA | Temperate Northern Pacific | 200 |
| Hyboscolex quadricincta Kudenov, 1985 | Florida, Gulf of Mexico | Tropical Atlantic | 0.6–31 |
| Hyboscolex reticulatus (McIntosh, 1885) | Queen Charlotte Sound, New Zealand | Temperate Australia | 2011 |
| Hyboscolex verrucosus Hartmann-Schröder, 1979 | Port Hedland, Western Australia | Central Indo-Pacific | 0–5 |
| Lipobranchius jeffreysii McIntosh, 1869 | Hebrides and Shetland Islands | Temperate Northern Atlantic | 22–1194 |
| Oligobregma aciculatum (Hartman, 1965) | New England, abyssal | Temperate Northern Atlantic | 1925–4825 |
| Oligobregma brasierae Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 4425 |
| Oligobregma collare (Levenstein, 1975) | Drake Passage, Antarctica | Southern Ocean | 3733–3806 |
| Oligobregma lonchochaeta Detinova, 1985 | Reykjanes Ridge, off Iceland | Temperate Northern Atlantic | 2930–2951 |
| Oligobregma mucronata Blake, 2015 | Greenpeace Trough, East Antarctic Peninsula | Southern Ocean | 323–912 |
| Oligobregma notiale Blake, 1981 | Palmer Archipelago, Antarctic Peninsula | Southern Ocean | 18–923 |
| Oligobregma oculata Kudenov and Blake, 1978 | East of Saint Maurice island, New Caledonia | Central Indo-Pacific | 57 |
| Oligobregma pseudocollare Schüller and Hilbig, 2007 | Scotia Sea, Antarctica | Southern Ocean | 2889–2892 |
| Oligobregma quadrispinosa Schüller and Hilbig, 2007 | Scotia Sea, Antarctica | Southern Ocean | 2258–2313 |
| Oligobregma simplex Kudenov and Blake, 1978 | Western Port, Victoria, Australia | Temperate Australia | 11 |
| Oligobregma tani Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 4137 |
| Oligobregma whaleyi Wiklund, Neal, Glover, Drennan, Rabone and Dahlgren, 2019 | Clarion-Clipperton Zone, Pacific Ocean | Tropical Eastern Pacific | 4425 |
| Parasclerocheilus branchiatus Fauvel, 1928 | Shingle Island, Gulf of Manaar, India | Western Indo-Pacific | N.d. |
| Parasclerocheilus capensis Day, 1961 | Langebaan Lagoon, South Africa | Temperate Southern Africa | 26 |
| Polyphysia caulleryi (McIntosh, 1922) | Inland Sea of Japan | Temperate Northern Pacific | 47 |
| Polyphysia crassa (Örsted, 1843) | Denmark | Temperate Northern Atlantic | 0–1755 |
| Polyphysia hystricis (McIntosh, 1922) | Channel slope, Antarctica | Temperate Northern Atlantic | 4701261 |
| Pseudoscalibregma bransfieldium (Hartman, 1967) | E. Bransfield Strait, Antarctica | Southern Ocean | 323–916 |
| Pseudoscalibregma hartmanae Blake, 1981 | Weddell Sea, Antarctica | Southern Ocean | 585 |
| Pseudoscalibregma orientalis Imajima, 2009 | Japan | Temperate Northern Pacific | 373–1005 |
| Pseudoscalibregma pallens Levenstein, 1962 | Kermadec Trench | Temperate Australia | 8928–9174 |
| Pseudoscalibregma palmeri Blake, 2015 | Weddell Sea, Off Lindenberg Island, Antarctica | Southern Ocean | 385–768 |
| Pseudoscalibregma papilia Schüller, 2008 | South Sandwich Islands, Antarctica | Southern Ocean | 2258–2313 |
| Pseudoscalibregma parvum (Hansen, 1878) | North Sea | Temperate Northern Atlantic | 53–1802 |
| Pseudoscalibregma usarpium Blake, 1981 | Ross Sea, Antarctica | Southern Ocean | 2143 |
| Scalibregma australis Blake, 2015 | Greenpeace Trough, East Antarctic Peninsula | Southern Ocean | 12–978 |
| Scalibregma californicum Blake, 2000 | Santa Maria Basin, off Point Sal, California | Temperate Northern Pacific | 90–2710 |
| Scalibregma celticum Mackie, 1991 | Milford Haven, Dyfed, Wales | Temperate Northern Atlantic | 6–21 |
| Scalibregma hanseni Bakken, Oug and Kongsrud, 2014 | Egga, west of Nordland County, Norway | Temperate Northern Atlantic | 765 |
| Scalibregma inflatum Rathke, 1843 | Norway | Temperate Northern Atlantic | 1–3690 |
| Scalibregma robustum Zachs, 1925 | White Sea, Russia | Artic | N.d. |
| Scalibregma stenocerum (Bertelsen and Weston, 1980) | Daytona Beach, Florida | Tropical Atlantic | 17–65 |
| Scalibregma wireni Furreg, 1925 | Kaiser Joseph Fjord, East Greenland | Artic | 3–9 |
| Scalibregmella antennata Hartman and Fauchald, 1971 | Bermuda, abyssal | Temperate Northern Atlantic | 4833–5023 |
| Scalibregmides chilensis Hartmann-Schröder, 1965 | Puerto Aguirre, Chile | Temperate Southern America | 10 |
| Scalibregmides peruanus Blake, 1981 | island near Pucusana, south of Callao, Peru | Temperate Southern America | 0–5 |
| Sclerobregma branchiatum Hartman, 1965 | New England | Temperate Northern Atlantic | 1330–2022 |
| Sclerocheilus antarcticus Ashworth, 1915 | Petermann Island, Antarctica | Southern Ocean | 45–311 |
| Sclerocheilus deriugini Zachs, 1925 | Kola Fjord, Northern Russia | Temperate Northern Atlantic | N.d. |
| Sclerocheilus minutus Grube, 1863 | Nerizine, Mali Losinj, Croatia | Temperate Northern Pacific | N.d. |
| Sclerocheilus unoculus Kudenov, 1985 | Florida, Gulf of Mexico | Tropical Atlantic | 18–37 |
| Speleobregma lanzaroteum Bertelsen, 1986 | La Corona lava tube, Lanzarote, Canary Islands | Temperate Northern Atlantic | 5–25 |
References
- Blake, J.A.; Maciolek, N.J. 7.6.1 Opheliidae Malmgren, 1867. In Handbook of Zoology. Annelida: Pleistoannelida, Sedentaria II; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany, 2020. [Google Scholar]
- Wiklund, H.; Neal, L.; Glover, A.G.; Drennan, R.; Rabone, M.; Dahlgren, T.G. Abyssal fauna of polymetallic nodule exploration areas, eastern Clarion-Clipperton Zone, central Pacific Ocean: Annelida: Capitellidae, Opheliidae, Scalibregmatidae, and Travisiidae. Zookeys 2019, 883, 1–82. [Google Scholar] [CrossRef]
- WoRMS Editorial Board. Register of Marine Species. 2020. Available online: http://www.marinespecies.org/ (accessed on 15 February 2021).
- Parapar, J. Familia Opheliidae. In Fauna Ibérica—Annelida Polychaeta III; Parapar, J., Alós, C., Núñez, J., Moreira, J., López, E., Aguirrezabalaga, F., Besteiro, C., Martínez, A., Ramos, M.A., Eds.; Museo Nacional de Ciencias Naturales, CSIC: Madrid, Spain, 2012; Volume 36, pp. 284–332. [Google Scholar]
- Wilson, D.P. The Larval Development of Ophelia bicornis Savigny. J. Mar. Biol. Assoc. UK 1948, 27, 540–553. [Google Scholar] [CrossRef] [Green Version]
- Dales, R.P. The larval development and ecology of Thoracophelia mucronata (Treadwell). Biol. Bull. 1952, 102, 232–242. [Google Scholar] [CrossRef]
- Seike, K. Burrowing behaviour inferred from feeding traces of the opheliid polychaete Euzonus sp. as response to beach morphodynamics. Mar. Biol. 2008, 53, 1199–1206. [Google Scholar] [CrossRef]
- Otegui, M.P.B.; Blankensteyn, A.; Pagliosa, P.R. Population structure, growth and production of Thoracophelia furcifera (Polychaeta: Opheliidae) on a sandy beach in Southern Brazil. Helgol. Mar. Res. 2012, 66, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Dorgan, K.M. Kinematics of burrowing by peristalsis in granular sands. J. Exp. Biol. 2018, 221, jeb167759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giangrande, A.; Gambino, I.; Tundo, M.; Pasqua, M.; Licciano, M.; Fanini, L.; Pinna, M. Reproductive biology of Ophelia barquii (Annelida, Opheliidae) along the Salento Peninsula (Mediterranean Sea, South Italy). Mar. Biodiv. 2020, 50, 3. [Google Scholar] [CrossRef]
- Meador, J.P.; Rice, C.A. Impaired growth in the polychaete Armandia brevis exposed to tributyltin in sediment. Mar. Environ. Res. 2001, 51, 113–129. [Google Scholar] [CrossRef]
- Bat, L.; Şahin, F.; Öztekin, A. Acute toxicity of cadmium on Ophelia bicornis Savigny, 1822. Acta Aquat. Turc. 2019, 15, 289–297. [Google Scholar] [CrossRef]
- Vanreusel, A.; Cosson-Sarradin, N.; Gooday, A.J.; Paterson, G.L.J.; Galéron, J.; Sibuet, M.; Vincx, M. Evidence for episodic recruitment in a small opheliid polychaete species from the abyssal NE Atlantic. Prog. Oceanogr. 2001, 50, 285–301. [Google Scholar] [CrossRef]
- Kongsrud, J.A.; Bakken, T.; Oug, E. Deep-water species of the genus Ophelina (Annelida, Opheliidae) in the Nordic Seas, with the description of Ophelina brattegardi sp. nov. Ital. J. Zool. 2011, 78, 95–111. [Google Scholar] [CrossRef]
- Parapar, J.; Moreira, J.; Helgason, G. Distribution and diversity of Opheliidae (Annelida, Polychaeta) on the continental shelf and slope of Iceland, with a review of the genus Ophelina in Northeast Atlantic waters and description of two new species. Org. Divers. Evol. 2011, 11, 83–105. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.S. The anatomy of the polychaete Ophelia cluthensis McGuire 1935. Proc. R. Soc. Edinb. 1938, 58, 135–160. [Google Scholar] [CrossRef]
- McConnaughey, B.H.; Fox, D.L. The Anatomy and Biology of the Marine Polychaete Thoracophelia mucronata (Treadwell) Opheliidae. Univ. Calif. Publ. Zool. 1949, 47, 319–340. [Google Scholar]
- Hermans, C.O.; Cloney, R.A. Fine structure of the prostomial eyes of Armandia brevis (Polychaeta: Opheliidae). Z. Zellforsch. Mikrosk. Anat. 1966, 72, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Purschke, G.; Ding, Z.; Müller, M.C. Ultrastructural differences as a taxonomic marker: The segmental ocelli of Polyophthalmus pictus and Polyophthalmus qingdaoensis sp. n. Polychaeta, Opheliidae). Zoomorphology 1995, 115, 229–241. [Google Scholar] [CrossRef]
- Tzetlin, A.B.; Filippova, A.V. Muscular system in polychaetes (Annelida). Hydrobiologia 2005, 535, 113–126. [Google Scholar] [CrossRef]
- Law, C.J.; Dorgan, K.M.; Rouse, G.W. Relating divergence in polychaete musculature to different burrowing behaviors: A study using Opheliidae. J. Morphol. 2014, 275, 548–571. [Google Scholar] [CrossRef] [PubMed]
- Misaka, T.; Sato, M. A new species of Euzonus (Polychaeta: Opheliidae) from subtidal zones in Japan. Zool. Sci. 2003, 20, 1171–1177. [Google Scholar] [CrossRef] [Green Version]
- Blake, J.A. 7.6.3 Scalibregmatidae Malmgren, 1867. In Handbook of Zoology. Annelida: Pleistoannelida, Sedentaria II; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany, 2020. [Google Scholar]
- Rouse, G.W.; Pleijel, F. Polychaetes; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Kudenov, J.D. Four new species of Scalibregmatidae (Polychaeta) from the Gulf of Mexico, with comments on the familial placement of Mucibregma Fauchald and Hancock, 1981. Proc. Biol. Soc. Wash. 1985, 98, 332–340. [Google Scholar]
- Martínez, A.; Di Domenico, M.; Worsaae, K. Evolution of cave Axiokebuita and Speleobregma (Scalibregmatidae, Annelida). Zool. Scr. 2013, 42, 623–636. [Google Scholar] [CrossRef]
- Hartman, O.; Fauchald, K. Deep-water benthic polychaetous annelids off New England to Bermuda and other North Atlantic areas. Part II. Allan Hancock Monogr. Mar. Biol. 1971, 6, 1–327. [Google Scholar]
- Ashworth, J.H. Memoirs: The Anatomy of Scalibregma inflatum, Rathke. J. Cell Sci. 1901, 45, 237–309. [Google Scholar]
- Ashworth, J.H. On a New Species of Sclerocheilus, with a revision of the genus. Trans. R. Soc. Edinb. 1915, 50, 405–422. [Google Scholar] [CrossRef] [Green Version]
- Rathke, H. Beiträge Zur Fauna Norwegens; Für die Akademie in Eduard Weber’s Buchhandlng in Bonn: Bonn, Germany, 1843; Volume 1, pp. 1–264. [Google Scholar]
- Blake, J.A. The Scalibregmatidae (Annelida: Polychaeta) from South America and Antarctica collected chiefly during the cruises of the R/V Anton Bruun, R/V Hero and USNS Eltanin. Proc. Biol. Soc. Wash. 1981, 94, 1131–1162. [Google Scholar]
- Pleijel, F.; Fauchald, K. Scalispinigera oculata Hartman, 1967 (Scalibregmatidae: Polychaeta): Senior synonym of Lacydonia antarctica (Lacydoniidae) Hartmann-Schröder & Rosenfeldt, 1988. Proc. Biol. Soc. Wash. 1993, 106, 673–677. [Google Scholar]
- Blake, J.A.; Maciolek, N.J. 7.6.2 Travisiidae Hartmann-Schröder, 1971, new family status. In Handbook of Zoology. Annelida: Pleistoannelida, Sedentaria II; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany, 2020; pp. 302–311. [Google Scholar]
- Mackie, Y. Scalibregma celticum new species (Polychaeta: Scalibregmatidae) from Europe, with a redescription of Scalibregma inflatum Rathke, 1843 and comments on the genus Sclerobregma Hartman, 1965. Bull. Mar. Sci. 1991, 48, 268–276. [Google Scholar]
- Bakken, T.; Oug, E.; Kongsrud, J.A. Occurrence and distribution of Pseudoscalibregma and Scalibregma (Annelida, Scalibregmatidae) in the deep Nordic Seas, with the description of Scalibregma hanseni n. sp. Zootaxa 2013, 3753, 101–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blake, J.A. New species of Scalibregmatidae (Annelida, Polychaeta) from the East Antarctic Peninsula including a description of the ecology and post-larval development of species of Scalibregma and Oligobregma. Zootaxa 2015, 4033, 57–93. [Google Scholar] [CrossRef] [Green Version]
- Schüller, M.; Hilbig, B. Three new species of the genus Oligobregma (Polychaeta, Scalibregmatidae) from the Scotia and Weddell Seas (Antarctica). Zootaxa 2007, 1391, 35–45. [Google Scholar] [CrossRef]
- Kudenov, D.; Blake, J.A. A review of the genera and species of the Scalibregmidae (Polychaeta) with descriptions of one new genus and three new species from Australia. J. Nat. Hist. 1978, 12, 427–444. [Google Scholar] [CrossRef]
- Nogueira, J.M.M. Asclerocheilus tropicus Blake, 1981 (Polychaeta: Scalibregmatidae): Redescription Based on Brazilian specimens. Proc. Biol. Soc. Wash. 2002, 115, 323–332. [Google Scholar]
- Hartmann-Schröder, G. Zur Kenntnis des Sublitorals der Chilenischen Küste unter besonderer Berücksichtigung der Polychaeten und Ostracoden. II Die Polychaeten des Sublitorals. Mitt. Zool. Mus. Hamburg 1965, 62, 59–305. [Google Scholar]
- Dehorne, A.; Dehorne, L. Recherches sur Sclerocheilus minutus (polychète de la famille des Scalibregmides). Morphologie, yeux, néphridie et pavillon. Arch. Zool. Exp. Gen. 1913, 53, 61–137. [Google Scholar]
- Orrhage, L.; Müller, M. Morphology of the Nervous System of Polychaeta (Annelida). Hydrobiologia 2005, 535/536, 79–111. [Google Scholar] [CrossRef]
- Martínez, A.; Di Domenico, M.; Worsaae, K. Gain of palps within a lineage of ancestrally burrowing annelids (Scalibregmatidae). Acta Zool. 2014, 95, 421–429. [Google Scholar] [CrossRef]
- Fauchald, K.; Rouse, G. Polychaete systematics: Past and present. Zool. Scri. 1997, 26, 71–138. [Google Scholar] [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine Ecoregions of the World: A Bioregionalization of Coastal and Shelf Areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Paul, C.; Halanych, K.M.; Tiedemann, R.; Bleidorn, C. Molecules reject an opheliid affinity for Travisia (Annelida). Syst. Biodivers. 2010, 8, 507–512. [Google Scholar] [CrossRef]
- Blake, J.A. Family Opheliidae Malmgren, 1867. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and the Western Santa Barbara Channel. Vol. 7. The Annelida. Part 4: Polychaeta: Flabelligeridae to Sternaspidae; Blake, J.A., Hilbig, B., Scott, P.V., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 2000; Volume 7, pp. 145–168. [Google Scholar]
- Bleidorn, C.; Vogt, L.; Bartolomaeus, T. New Insights into Polychaete Phylogeny (Annelida) Inferred from 18S RDNA Sequences. Mol. Phylogenet. Evol. 2003, 29, 279–288. [Google Scholar] [CrossRef]
- Hall, K.A.; Hutchings, P.A.; Colgan, D.J. Further phylogenetic studies of the Polychaeta using 18S RDNA sequence data. J. Mar. Biol. Asspc. UK 2004, 84, 949–960. [Google Scholar] [CrossRef]
- Belova, P.A.; Zhadan, A.E. Comparative morphology and ultrastructure of the respiratory system in four species of the Opheliidae family. Biol. Bull. 2014, 41, 752–772. [Google Scholar] [CrossRef]
- Weigert, A.; Helm, C.; Meyer, M.; Nickel, B.; Arendt, D.; Hausdorf, B.; Santos, S.R.; Halanych, K.M.; Purschke, G.; Bleidorn, C.; et al. Illuminating the Base of the Annelid Tree Using Transcriptomics. Mol. Biol. Evol. 2014, 31, 1391–1401. [Google Scholar] [CrossRef]
- Andrade, S.C.S.; Novo, M.; Kawauchi, G.Y.; Worsaae, K.; Pleijel, F.; Giribet, G.; Rouse, G.W. Articulating “Archiannelids”: Phylogenomics and Annelid Relationships, with Emphasis on Meiofaunal Taxa. Mol. Biol. Evol. 2015, 32, 2860–2875. [Google Scholar] [CrossRef]
- Sene-Silva, G. Filogenia de Opheliidae (Annelida: Polychaeta). Ph.D. Thesis, Universidade Federal do Paraná, Curitiba, Brazil, 2007. [Google Scholar]
- Blake, J.A. Revalidation of the genus Thoracophelia Ehlers, 1897, replacing Euzonus Grube, 1866 (Polychaeta: Opheliidae), junior homonym of Euzonus Menge, 1854 (Arthropoda: Diplopoda), together with a literature summary and updated listing of Thoracophelia species. Zootaxa 2011, 2807, 65–68. [Google Scholar] [CrossRef]
- Schüller, M. New Polychaete Species Collected during the Expeditions ANDEEP I, II, and III to the Deep Atlantic Sector of the Southern Ocean in the Austral Summers 2002 and 2005—Ampharetidae, Opheliidae, and Scalibregmatidae. Zootaxa 2008, 1705, 51–68. [Google Scholar] [CrossRef]
- Day, J.H. A Monograph on the Polychaeta of Southern Africa. Part 2, Sedentaria; British Museum (Natural History): London, UK, 1967; Volume 2. [Google Scholar]
- Santos, C.S.G.; Nonato, E.F.; Petersen, M.E. Two New Species of Opheliidae (Annelida: Polychaeta): Euzonus papillatus sp. n. from a Northeastern Brazilian Sandy Beach and Euzonus mammillatus sp. n. from the Continental Shelf of Southeastern Brazil. Zootaxa 2004, 478, 1–12. [Google Scholar] [CrossRef]
- Neave, M.J.; Glasby, C.J. New Species of Ophelina (Annelida: Opheliidae: Ophelininae) from Northern Australia. Org. Divers. Evol. 2013, 13, 331–347. [Google Scholar] [CrossRef]
- Gopal, A.; Jaleel, A.K.U.; Parameswaran, U.V.; Vijayan, A.-K. Armandia sampadae, a new species of polychaete (Opheliidae) from Andaman Sea, Northern Indian Ocean. J. Mar. Biol. Assoc. UK 2016, 96, 1625–1632. [Google Scholar] [CrossRef]
- Magalhães, W.F.; Rizzo, A.E.; Bailey-Brock, J.H. Opheliidae (Annelida: Polychaeta) from the Western Pacific Islands, including five new species. Zootaxa 2019, 4555, 209–235. [Google Scholar] [CrossRef]
- Parapar, J.; Gambi, M.C.; Rouse, G.W. A revision of the deep-sea genus Axiokebuita Pocklington and Fournier, 1987 (Annelida: Scalibregmatidae). Ital. J. Zool. 2011, 78, 148–162. [Google Scholar] [CrossRef] [Green Version]
- Parapar, J.; Moreira, J. Six new species of the genus Armandia Filippi, 1861 (Polychaeta, Opheliidae) from Lizard Island (Great Barrier Reef, Australia). Zootaxa 2015, 4019, 577–603. [Google Scholar] [CrossRef] [Green Version]
- Moreira, J.; Parapar, J. New data on the Opheliidae (Annelida) from Lizard Island (Great Barrier Reef, Australia): Five new species of the genus Armandia Filippi, 1861. Zootaxa 2017, 4290, 483–502. [Google Scholar] [CrossRef]
- Tzetlin, A.; Zhadan, A. Morphological variation of axial non-muscular proboscis types in the Polychaeta. Zoosymposia 2009, 2, 415–427. [Google Scholar] [CrossRef]
- Purschke, G. Ultrastructure of Nuchal Organs in Polychaetes (Annelida)—New Results and Review. Acta Zool. 1997, 78, 123–143. [Google Scholar] [CrossRef]
- Saito, H.; Tamaki, A.; Imajima, M. Description of a new species of Armandia (Polychaeta: Opheliidae) from Western Kyushu, Japan, with character variations. J. Nat. Hist. 2000, 34, 2029–2043. [Google Scholar] [CrossRef]
- Bartolomaeus, T. Different photoreceptors in juvenile Ophelia rathkei (Annelida, Opheliida). Microfauna Mar. 1993, 8, 99–114. [Google Scholar]
- Parapar, J.; Moreira, J. Sobre la presencia del género Ophelina Ørsted, 1843 (Polychaeta, Opheliidae) en el litoral de la península Ibérica. Nova Acta Cient. Compostel. Biol. 2008, 17, 117–134. [Google Scholar]
- Hartmann-Schröder, G. Zur Morphologie der Opheliiden (Polychaeta Sedentaria). Z. Wiss. Zool. 1958, 161, 84–143. [Google Scholar]
- Penry, D.L.; Jumars, P.A. Gut architecture, digestive constraints and feeding ecology of deposit-feeding and carnivorous polychaetes. Oecologia 1990, 82, 1–11. [Google Scholar] [CrossRef]
- Bartolomaeus, T.; Quast, B. Structure and development of nephridia in Annelida and related taxa. Hydrobiologia 2005, 535, 139–165. [Google Scholar] [CrossRef]
- West, D.L. Comparative Ultrastructure of juvenile and adult nuchal organs of an annelid (Polychaeta, Opheliidae). Tissue Cell 1978, 10, 243–257. [Google Scholar] [CrossRef]
- Hermans, C.O. Fine structure of the segmental ocelli of Armandia brevis (Polychaeta: Opheliidae). Z. Zellforsch. Mikrosk. Anat. 1969, 96, 361–371. [Google Scholar] [CrossRef]
- Parapar, J.; Candás, M.; Cunha-Veira, X.; Moreira, J. Exploring annelid anatomy using micro-computed tomography: A taxonomic approach. Zool. Anz. 2017, 270, 19–42. [Google Scholar] [CrossRef]
- Bellan, G.; Dauvin, J.C. Phenetic and biogeographic relationships in Ophelia (Polychaeta, Opheliidae). Bull. Mar. Sci. 1991, 48, 544–558. [Google Scholar]
- Neal, L.; Taboada, S.; Woodall, L.C. Slope-Shelf Faunal Link and Unreported Diversity off Nova Scotia: Evidence from Polychaete Data. Deep Sea Res. Part I Oceanogr. Res. Pap. 2018, 138, 72–84. [Google Scholar] [CrossRef]
- Woodin, S.A. Polychaete Abundance Patterns in a Marine Soft-Sediment Environment: The Importance of Biological Interactions. Ecol. Monog. 1974, 44, 171–187. [Google Scholar] [CrossRef]
- Kuş, S.; Kurt-Şahin, G. Temporal Changes in the Polychaeta (Annelida) Community Associated with Cystoseira Beds of Sinop Peninsula (Southern Black Sea). Turk. J. Fish. Aquat. Sci. 2016, 16, 61–68. [Google Scholar] [CrossRef]
- Hadiyanto, H. Fouling Polychaetes in Tanjung Priok Port of Jakarta, Indonesia. ASEAN J. Sci. Technol. Dev. 2018, 35, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Dorgan, K.M.; Law, C.J.; Rouse, G.W. Meandering worms: Mechanics of undulatory burrowing in muds. Proc. R. Soc. B 2013, 280, 20122948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dafoe, L.T.; Gingras, M.K.; Pemberton, S.G. Determining Euzonus mucronata Burrowing Rates with Application to Ancient Macaronichnus segregatis Trace-makers. Ichnos 2008, 15, 78–90. [Google Scholar] [CrossRef]
- Riser, N.W. Observations on the genus Ophelia (Polychaeta: Opheliidae) with the description of a new species. Ophelia 1987, 28, 11–29. [Google Scholar] [CrossRef]
- Castelli, A.; Lardicci, C.; Castellani, C.; Finocchiaro, C.; Genovesi, S.; Tataranni, M.; Maltagliati, F. Analysis of vertical distribution of two polychaetes (genus Ophelia) in sandy substrate along the Pisan Coast. Biol. Mar. Mediterr. 2006, 13, 160–161. [Google Scholar]
- Tamaki, A. Zonation by size in the Armandia sp. (Polychaeta: Opheliidae) population on an intertidal sand flat. Mar. Ecol. Prog. Ser. 1985, 27, 123–133. [Google Scholar] [CrossRef]
- Fauchald, K.; Jumars, P.A. The diet of worms: A study of polychaete feeding guilds. Oceanogr. Mar. Biol. Ann. Rev. 1979, 17, 193–284. [Google Scholar]
- Kemp, P.F. Direct uptake of detrital carbon by the deposit feeding polychaete Euzonus mucronata (Treadwell). J. Exp. Mar. Biol. Ecol. 1986, 99, 49–61. [Google Scholar] [CrossRef]
- Guérin, J.-P. Modalités d’élevage et description des stades larvaires de Polyophthalmus pictus Dujardin (Annélide Polychète). Vie Milieu 1971, 22, 143–152. [Google Scholar]
- Giangrande, A. Polychaete reproductive patterns, life cycles and life histories: An overview. Oceanogr. Mar. Biol. Ann. Rev. 1997, 35, 323–386. [Google Scholar]
- Hermans, C.O. Metamorphosis in the opheliid polychaete Armandia brevis. In Settlement and Metamorphosis of Marine Invertebrate Larvae; Chia, F., Rice, M.E., Eds.; Elsevier: New York, NY, USA, 1978; pp. 113–126. [Google Scholar]
- Rivain, V. Contribution à L’étude Dynamique et Fonctionnelle des Peuplements de Sables fins du Golfe Normano-Breton. Ph.D Thesis, Université Pierre et Marie Curie, Paris, France, 1983. [Google Scholar]
- Miner, B.G.; Sanford, E.; Strathmann, R.R.; Pernet, B.; Emlet, R.B. Functional and Evolutionary Implications of Opposed Bands, Big Mouths, and Extensive Oral Ciliation in Larval Opheliids and Echiurids (Annelida). Biol. Bull. 1999, 197, 14–25. [Google Scholar] [CrossRef]
- Guérin, J.-P. Le développement larvaire d’Armandia cirrhosa Filippi (Annélide Polychète). Tethys 1972, 4, 969–974. [Google Scholar]
- Parke, S.R. Biological Aspects of Speciation in Three Sympatric Euzonus Species at Dillon Beach, California (Polychaeta: Opheliidae). Master’s Thesis, University of the Pacific, Stockton, CA, USA, 1973. [Google Scholar]
- Wilson, D.P. The role of micro-organisms in the settlement of Ophelia bicornis Savigny. J. Mar. Biol. Assoc. UK 1955, 34, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Bely, A.E. Distribution of segment regeneration ability in the Annelida. Integr. Comp. Biol. 2006, 46, 508–518. [Google Scholar] [CrossRef] [Green Version]
- Conradi, M.; Bandera, M.E.; Marin, I.; Martin, D. Polychaete-parasitizing copepods from the deep-sea Kuril–Kamchatka Trench (Pacific Ocean), with the description of a new Ophelicola species and comments on the currently known annelidicolous Copepods. Deep Sea Res. II 2015, 111, 147–165. [Google Scholar] [CrossRef] [Green Version]
- Hartmann-Schröder, G. Annelida, Borstenwürmer, Polychaeta: Tierwelt Deutschlands 58 (Die Tierwelt Deutschlands und der Angrenzenden Meeresteile); Gustav Fischer: Jena, Germany, 1996; ISBN 978-3-437-35038-2. [Google Scholar]
- Georgieva, Y.G.; Daskalov, G.M.; Klayn, S.L.; Stefanova, K.B.; Stefanova, E.S. Seasonal Diet and Feeding Strategy of Horse Mackerel Trachurus mediterraneus (Steindachner, 1868) (Perciformes: Carangidae) in the South-Western Black Sea. Acta Zool. Bulg. 2019, 71, 201–210. [Google Scholar]
- Kicklighter, C.E.; Hay, M.E. Integrating Prey Defensive Traits: Contrasts of Marine Worms from Temperate and Tropical Habitats. Ecol. Monogr. 2006, 76, 195–215. [Google Scholar] [CrossRef]
- Saes, R.V.S.T.; Moreira, L.B.; Davanso, M.B.; Perina, F.C.; Abess, D.M.S. Developing a protocol whole sediment toxicity testing with the polychaete Armandia agilis. Ecotoxicol. Environ. Contam. 2018, 13, 85–97. [Google Scholar] [CrossRef]
- Malmgren, A.J. Annulata Polychaeta: Spetsbergiae, Groenlandiae, Islandiae et Scandinaviae. Hactenus Cognita; Ex Officina Frenckelliana: Helsinki, Finland, 1867. [Google Scholar]
- Fauchald, K. Polychaetes from intertidal areas in Panama, with a review of previous shallow-water records. Smithson. Contr. Zool. 1977, 221, 1–81. [Google Scholar] [CrossRef] [Green Version]
- Persson, J.; Pleijel, F. On the phylogenetic relationships of Axiokebuita, Travisia and Scalibregmatidae (Polychaeta). Zootaxa 2005, 998, 1–14. [Google Scholar] [CrossRef]
- Helm, C.; Beckers, P.; Bartolomaeus, T.; Drukewitz, S.H.; Kourtesis, I.; Weigert, A.; Purschke, G.; Worsaae, K.; Struck, T.H.; Bleidorn, C. Convergent evolution of the ladder-like ventral nerve cord in Annelida. Front. Zool. 2018, 15, 36. [Google Scholar] [CrossRef]
- Weigert, A.; Bleidorn, C. Current status of annelid phylogeny. Org. Divers. Evol. 2016, 16, 345–362. [Google Scholar] [CrossRef]
- Örsted, A.S. Annulatorum Danicorum Conspectus. Fasc. I. Maricolæ; Librarie Wahliannae: Copenhagen, Denmark, 1843. [Google Scholar]
- Grube, A.E. Beschreibung Neuer Oder Wenig Bekannter Anneliden. Sechster Beitrag. Arch. Nat. 1863, 29, 37–69. [Google Scholar]
- Saint-Joseph, A.D. Les Annélides Polychètes Des Côtes de Dinard. Troisième Partie. Ann. Sci. Nat. 1849, 17, 1–395. [Google Scholar]
- Hansen, G.A. Annelider Fra Den Norske Nordhavsexpedition i 1876. Nyt Mag. Naturvid. 1879, 24, 1–17. [Google Scholar]
- McIntosh, W.C. On the Structure of the British Nemerteans, and Some New British Annelids. Trans. R. Soc. Edinb. 1869, 25, 305–433. [Google Scholar] [CrossRef] [Green Version]
- Schmarda, L.K. Neue Wirbellose Thiere: Beobachted und Gesammelt auf einer Reise um die Erdr 1853 bis 1857. In Turbellarien, Rotatorien und Anneliden; Erster Band, Zweite Hälfte; Verlag von Wilhelm Engelmann: Leipzig, Germany, 1861. [Google Scholar]
- McIntosh, W.C. Report on the Scientific Results of the Voyage of H.M.S. Challenger during the Years 1873–76, under the Command of Captain George S. Nares and the Late Captain Frank Tourle Thomson: A Summary of the Scientific Results; Printed for H.M.S.O.: London, UK, 1885. [Google Scholar]
- Ehlers, E. Reports on the Results of Dredging, Under the Direction of L.F. Pourtalès, During the Years 1868–1870, and of Alexander Agassiz in the Gulf of Mexico (1877–78) and in the Caribbean Sea (1878–79), in the US Coast. Survey Steamer “Blake” Lieut.-Com. CD Sigsbee, USN, and Commander JR Bartlett, USN, Commanding. XXXI. Report on the Annelids. Mem. Mus. Comp. Zool. Harv. Coll. 1887, 15, 1–335. [Google Scholar]
- Furreg, E. Zur Systematik Der Polychätenfamilie Scalibregmidae. Zool. Jahrb. Abt. Syst. Geogr. Biol. Tiere 1925, 50, 123–190. [Google Scholar]
- Hartman, O. Deep-Water Benthic Polychaetous Annelids off New England to Bermuda and Other North Atlantic Areas. Allan Hancock Found. Publ. Occas. Pap. 1965, 28, 1–384. [Google Scholar]
- Blake, J.A. Two new species of polychaetous annelid worms from Baffin Bay and the Davis Strait. Bull. South. Calif. Acad. Sci. 1972, 71, 127–132. [Google Scholar]
- Day, J.H. The Polychaet [Sic] Fauna of South Africa. Part 6. Sedentary Species Dredged off Cape Coasts with a Few New Records from the Shore. J. Linn. Soc. 1961, 44, 463–560. [Google Scholar] [CrossRef]
- Kirkegaard, J.B. Bathyal and Abyssal Polychaetes (Sedentary Species I). Galathea Rep. 1996, 17, 57–77. [Google Scholar]
- Blake, J. Family Scalibregmatidae Malmgren, 1867. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and the Western Santa Barbara Channel. Vol. 7. The Annelida. Part 4: Polychaeta: Flabelligeridae to Sternaspidae; Blake, J., Hilbig, B., Scott, P., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 2000; Volume 4, pp. 129–144. [Google Scholar]
- Imajima, M. Deep-Sea Benthic Polychaetes off Pacific Coast of the Northern Honshu, Japan. Natl. Mus. Nat. Sci. Monogr. 2009, 39, 39–192. [Google Scholar]
- Imajima, M. Occurrence of Oncoscolex pacificus (Moore), a Species of Family Scalibregmidae (Polychaeta), in Japan. J. Hokkaido Gakugei Univ. 1961, 12, 11–13. [Google Scholar]
- Støp-Bowitz, C. Les Scalibregmiens de Norvége. Medd. Zool. Mus. Oslo 1945, 55, 63–87. [Google Scholar]
- Wesenberg-Lund, E. Lesser Antillean Polychaetes, Chiefly from Brackish Water with a Survey and a Bibliography of Fresh and Brackish–Water Polychaetes. Studies on the Fauna of Curaçao and Other Caribbean Islands 8.1. Stud. Fauna Curacao Caribb. Isl. 1958, 8, 1–41. [Google Scholar]
- Bertelsen, R.D. Speleobregma lanzaroteum, a New Genus and Species of Scalibregmatidae (Polychaeta) from a Marine Cave in the Canary Islands. Proc. Biol. Soc. Wash. 1983, 99, 375–379. [Google Scholar]
- Martínez, A.; Gonzalez, B.C.; Worsaae, K.; Wilkens, H.; Núñez, J.; Oromí, P.; Iliffe, T.M. Guide to the Anchialine Ecosystems of Jameos Del Agua and Túnel de La Atlántida; Medio Ambiente; Cabildo de Lanzarote: Arrecife, Spain, 2016. [CrossRef]
- Martínez, A.; Palmero, A.M.; Brito, M.D.C.; Núñez, J.; Worsaae, K. Anchialine fauna of the Corona lava tube (Lanzarote, Canary Islands): Diversity, endemism and distribution. Mar. Biodiver. 2009, 39, 169–187. [Google Scholar] [CrossRef]
- Martínez, A.; Gonzalez, B.C. Volcanic Anchialine Habitats of Lanzarote. In Cave Ecology. Ecological Studies (Analysis and Synthesis); Moldovan, O.T., Kovac, L., Halse, S., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- García-Valdecasas, A. Estudio Faunístico de La Cueva Submarina “Túnel de La Atlántida”, Jameos Del Agua, Lanzarote. Nat. Hisp. 1985, 27, 1–56. [Google Scholar]
- Martínez, A.; Di Domenico, M.; Leasi, F.; Curini-Galletti, M.; Todaro, M.A.; Zotto, M.D.; Gobert, S.; Artois, T.; Norenburg, J.; Jörger, K.M.; et al. Patterns of Diversity and Endemism of Soft-Bodied Meiofauna in an Oceanic Island, Lanzarote, Canary Islands. Mar. Biodiv. 2019, 49, 2033–2055. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S.; Lunghi, E.; Bilandžija, H.; Cardoso, P.; Grimm, V.; Schmidt, S.I.; Hesselberg, T.; Martínez, A. Collecting Eco-Evolutionary Data in the Dark: Impediments to Subterranean Research and How to Overcome Them. EcoEvorXiv 2020. [Google Scholar] [CrossRef]
- Riera, R.; Monterroso, Ó.; Núñez, J.; Martínez, A. Distribution of meiofaunal abundances in a marine cave complex with secondary openings and freshwater filtrations. Mar. Biodivers. 2018, 48, 203–215. [Google Scholar] [CrossRef]
- Gonzalez, B.C.; Martínez, A.; Borda, E.; Iliffe, T.M.; Fontaneto, D.; Worsaae, K. Genetic spatial structure of an anchialine cave annelid indicates connectivity within–but not between–islands of the Great Bahama Bank. Mol. Phylogenet. Evol. 2017, 109, 259–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorgan, K.M.; Jumars, P.A.; Johnson, B.D.; Boudreau, B.P. Macrofaunal Burrowing: The Medium Is the Message. Oceanogr. Mar. Biol. Ann. Rev. 2006, 44, 5–121. [Google Scholar]
- Elder, H.Y. Direct Peristaltic Progression and the Functional Significance of the Dermal Connective Tissues during Burrowing in the Polychaete Polyphysia crassa (Oersted). J. Exp. Biol. 1973, 58, 637–655. [Google Scholar]
- Hunter, R.D.; Elder, H.L. Burrowing Dynamics and Energy Cost of Transport in the Soft-Bodied Marine Invertebrates Polyphysia crassa and Priapulus caudatus. J. Zool. 1983, 218, 209–222. [Google Scholar] [CrossRef]
- Cahoon, L.B.; Laws, R.A.; Thomas, C.J. Viable diatoms and chlorophylla in continental slope sediments off Cape Hatteras, North Carolina. Deep Sea Res. Part II Top. Stud. Oceanogr. 1994, 41, 767–782. [Google Scholar] [CrossRef]
- Aller, J.Y.; Aller, R.C.; Green, M.A. Benthic faunal assemblages and carbon supply along the continental shelf/shelf break-slope off Cape Hatteras, North Carolina. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 4599–4625. [Google Scholar] [CrossRef]
- Blake, J.A.; Hilbig, B. Dense infaunal assemblages on the continental slope off Cape Hatteras, North Carolina. Deep Sea Res. Part II Top. Stud. Oceanogr. 1994, 41, 875–899. [Google Scholar] [CrossRef]
- Worsaae, K.; Rouse, G.W. Mesonerilla neridae sp. nov. (Nerillidae): First meiofaunal annelid from deep-sea hydrothermal vents. Zoosymposia 2009, 2, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Hartman, O. Quantitative survey of the benthos of San Pedro Basin, southern California. Part II. Final results and conclusions. Allan Hancock Pac. Exped. 1966, 19, 187–455. [Google Scholar]
- Hartmann-Schröder, G.; Hartmann, G. Zur Kenntnis Des Eulitorals Der Australischen Küsten Unter Besonderer Berücksichtigung Der Polychaeten Und Ostracoden (Extract Hyboscolex). Mitt. Hambg. Zool. Mus. Inst. 1979, 76, 75–218. [Google Scholar]
- Hartman, O. Submarine canyons of southern California. Polychaetous Annelids. Allan Hancock Pac. Exped. 1963, 27, 1–93. [Google Scholar]
- Wilkens, H.; Iliffe, T.M.; Oromí, P.; Martínez, A.; Tysall, T.N.; Koenemann, S. The Corona Lava Tube, Lanzarote: Geology, Habitat Diversity and Biogeography. Mar. Biodiv. 2009, 39, 155–167. [Google Scholar] [CrossRef]
- Martínez, A.; Kvindebjerg, K.; Iliffe, T.M.; Worsaae, K. Evolution of cave suspension feeding in Protodrilidae (Annelida). Zool. Scr. 2017, 46, 214–226. [Google Scholar] [CrossRef]
- Worsaae, K.; Gonzalez, B.C.; Kerbl, A.; Nielsen, S.H.; Jørgensen, J.T.; Armenteros, M.; Iliffe, T.M.; Martínez, A. Diversity and evolution of the stygobitic Speleonerilla nom. nov. (Nerillidae, Annelida) with description of three new species from anchialine caves in the Caribbean and Lanzarote. Mar. Biodiv. 2019, 49, 2167–2192. [Google Scholar] [CrossRef]
- Ditlevsen, H. Annelids from the Danmark Expedition. Medd. Grønland Kobenhaven 1911, 45, 411–432. [Google Scholar]
- Clark, R. Pelagic Swarming of Scalibregmidae (Polychaeta). Annu. Rep. Scott. Mar. Biol. Assoc. 1952, 53, 20–22. [Google Scholar]
- Fage, L.; Legendre, R. Pêches Planctoniques à La Lumière, Effectuées à Banyuls-Sur-Mer et à Concarneau. I. Annélides Polychètes. Arch. Zool. Exp. Gen. 1927, 67, 23–222. [Google Scholar]
- Blake, J.A. Life history analysis of five dominant infaunal polychaete species from the continental slope off North Carolina. J. Mar. Biol. Assoc. UK 1993, 73, 123–141. [Google Scholar] [CrossRef]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parapar, J.; Martínez, A.; Moreira, J. On the Systematics and Biodiversity of the Opheliidae and Scalibregmatidae. Diversity 2021, 13, 87. https://doi.org/10.3390/d13020087
Parapar J, Martínez A, Moreira J. On the Systematics and Biodiversity of the Opheliidae and Scalibregmatidae. Diversity. 2021; 13(2):87. https://doi.org/10.3390/d13020087
Chicago/Turabian StyleParapar, Julio, Alejandro Martínez, and Juan Moreira. 2021. "On the Systematics and Biodiversity of the Opheliidae and Scalibregmatidae" Diversity 13, no. 2: 87. https://doi.org/10.3390/d13020087
APA StyleParapar, J., Martínez, A., & Moreira, J. (2021). On the Systematics and Biodiversity of the Opheliidae and Scalibregmatidae. Diversity, 13(2), 87. https://doi.org/10.3390/d13020087