
Reappraisal on the Phylogenetic Relationships of the Enigmatic Flightless Bird (Brontornis burmeisteri) Moreno and Mercerat, 1891

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nomenclature
2.2. Phylogenetic Analysis
2.3. Institutional Abbreviations
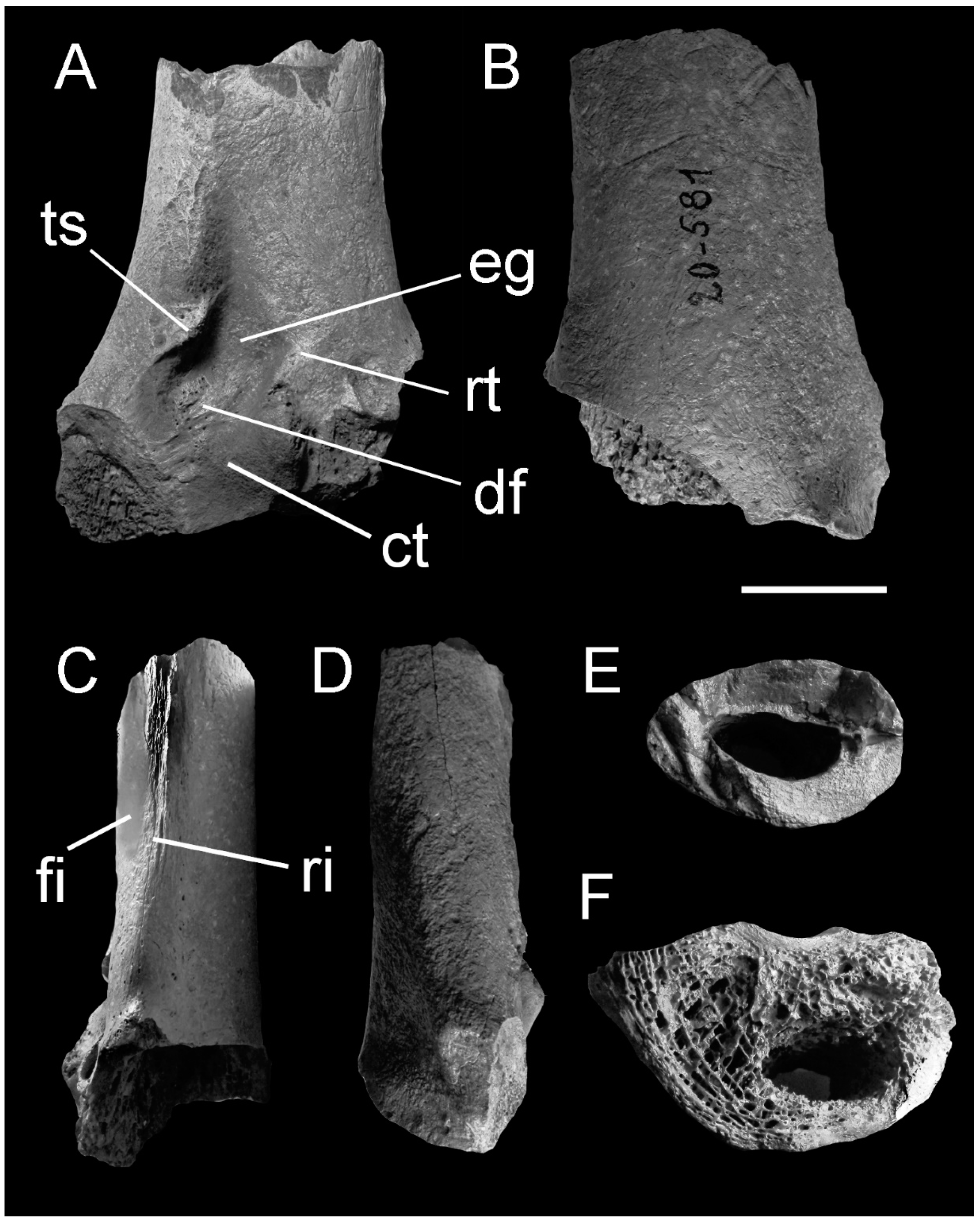
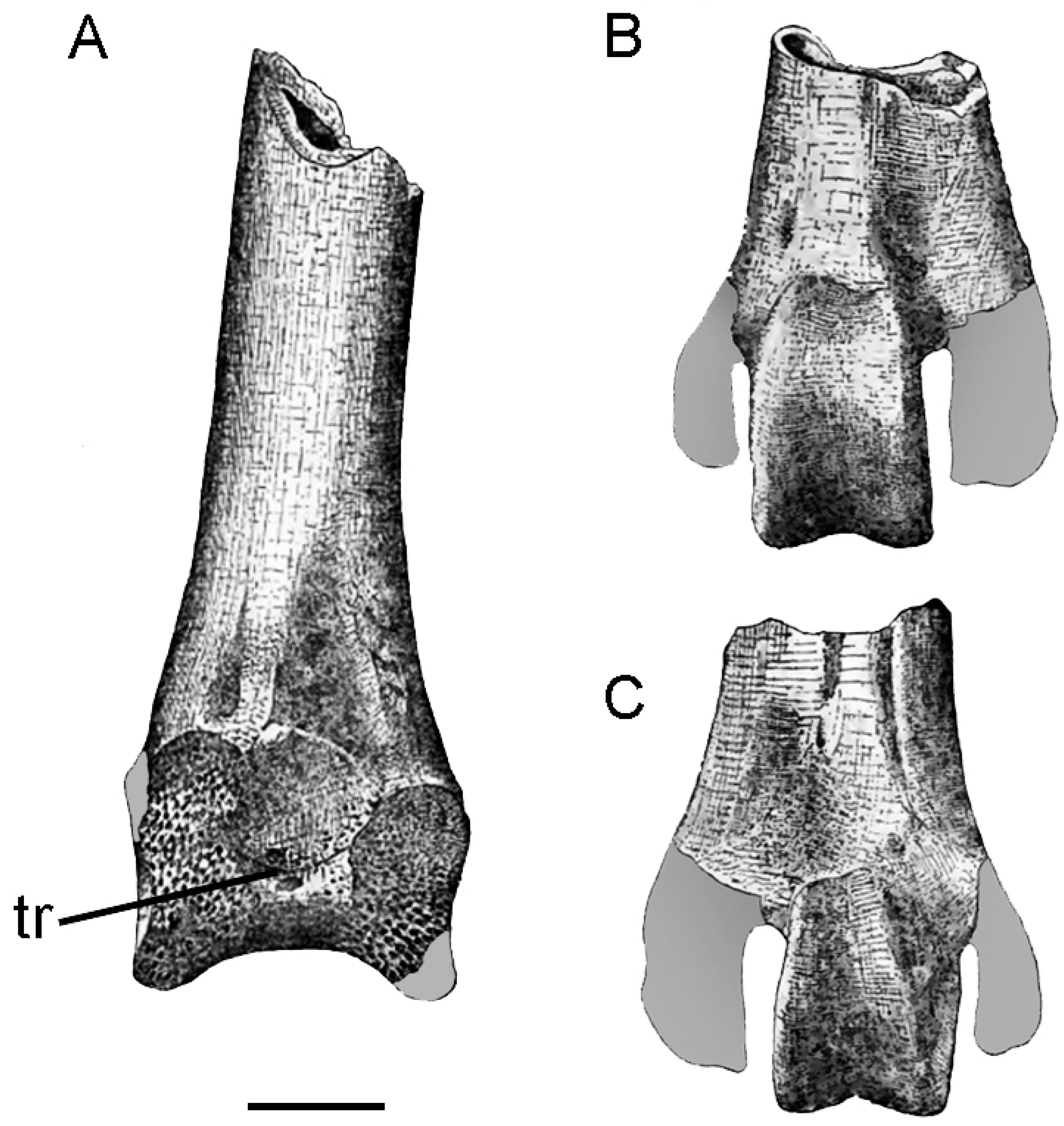
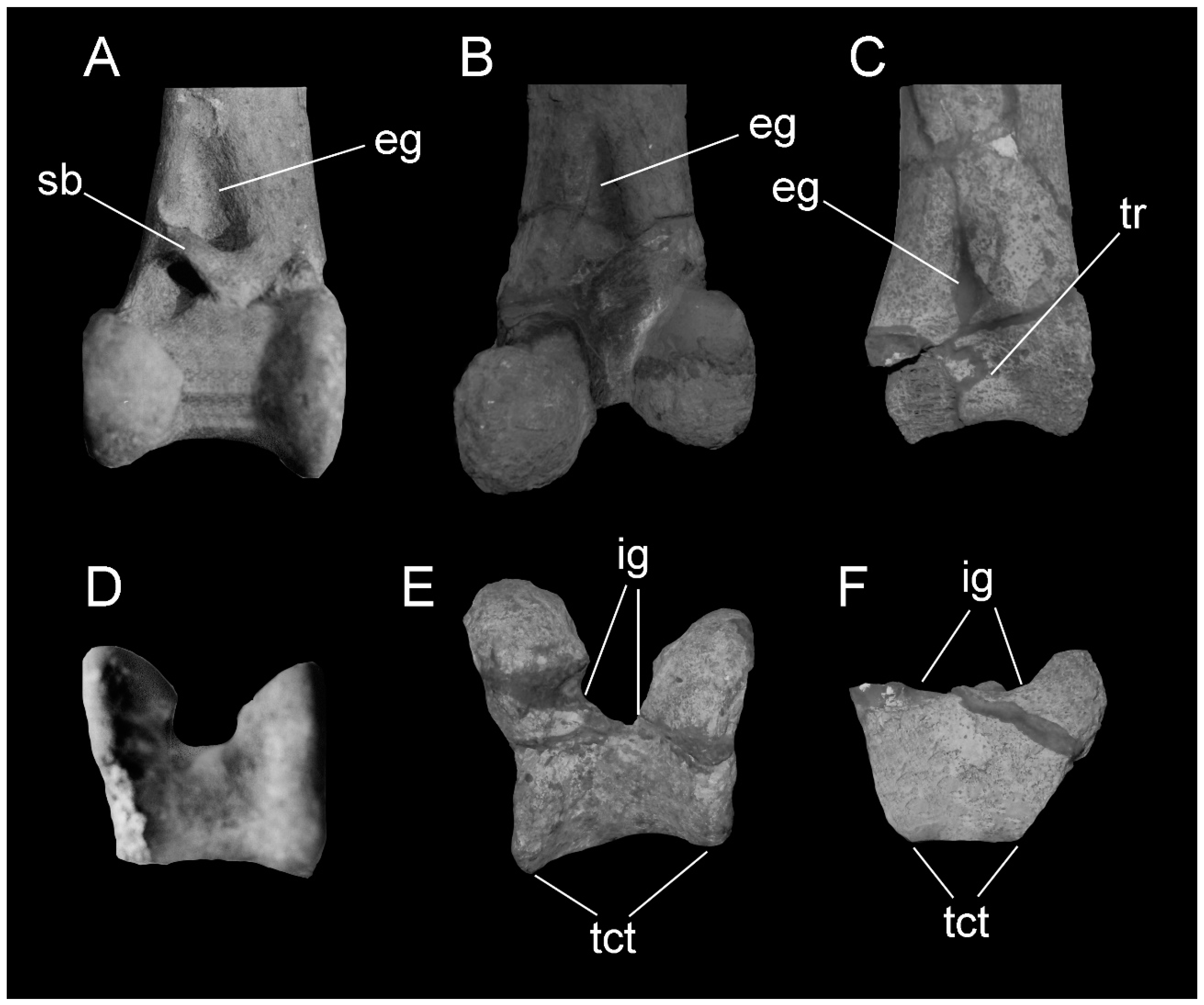
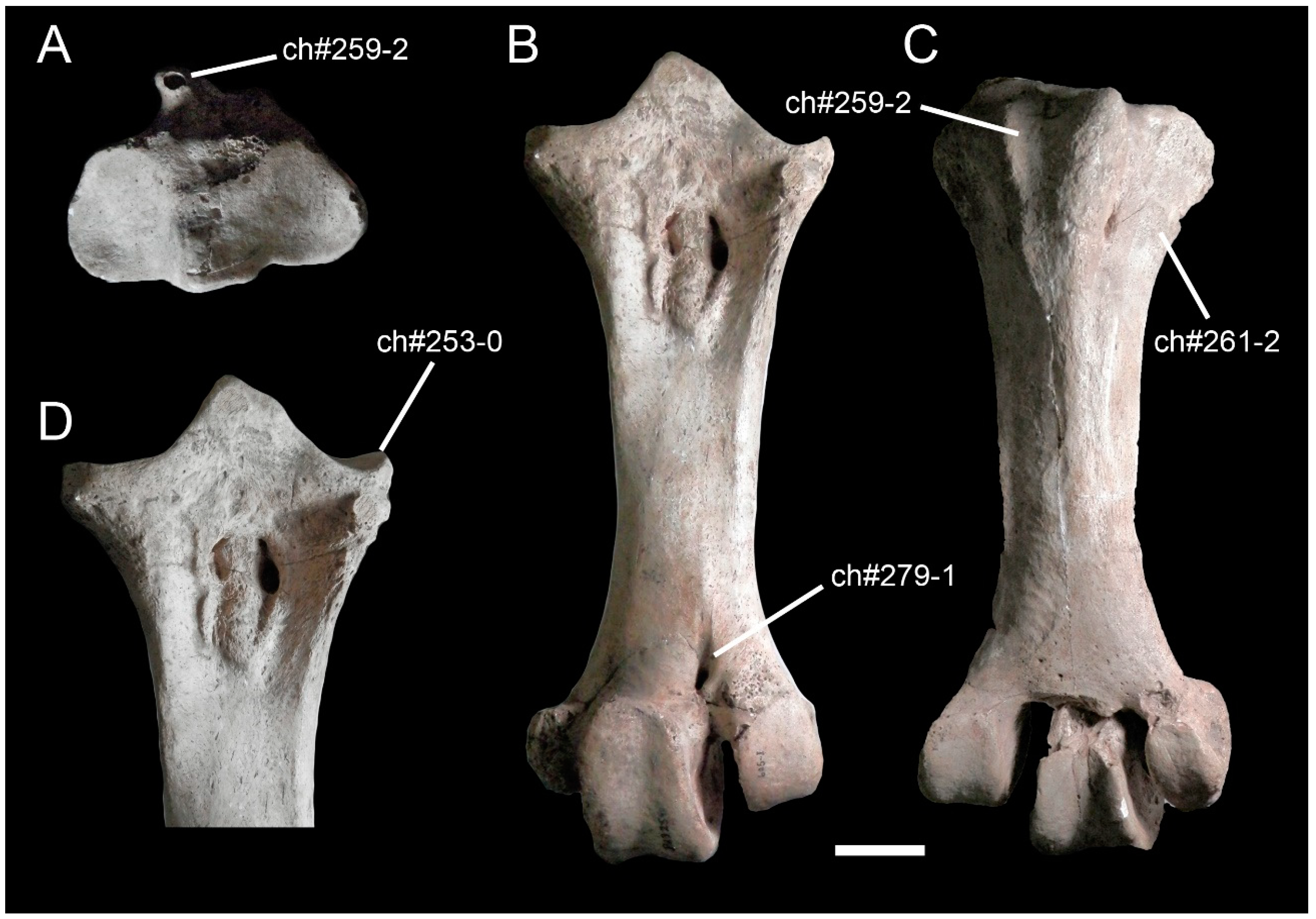
3. Description
4. Discussion
4.1. Comments on the Genus Liornis Ameghino, 1895
4.2. The Quadrate Bone Referred to Brontornis burmeisteri Moreno and Mercerat, 1891
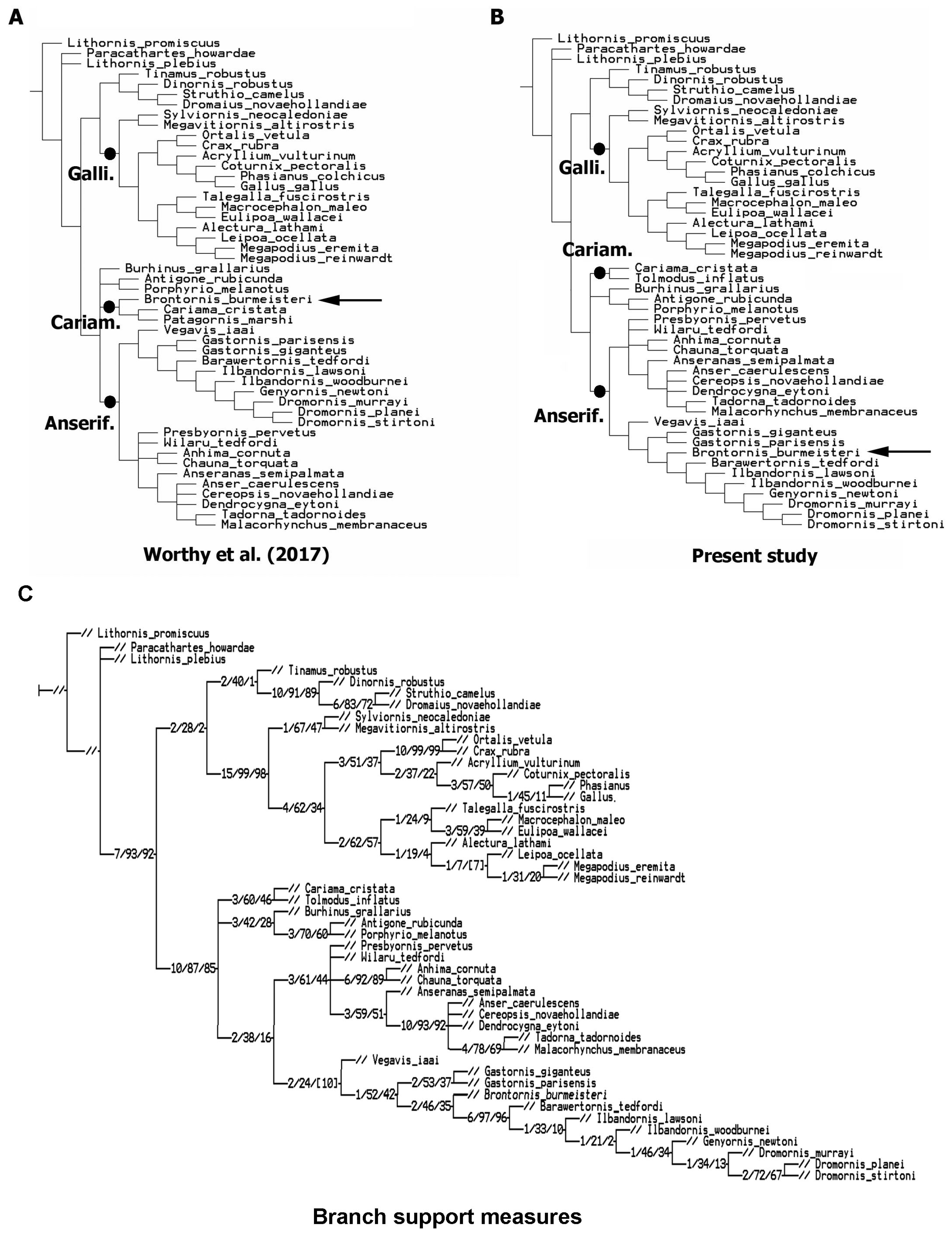
4.3. Phylogenetic Results
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Tonni, E.P. The present state of knowledge of the Cenozoic birds of Argentina. Contrib. Sci. 1980, 330, 105–114. [Google Scholar]
- Alvarenga, H.M.; Höfling, E. Systematic revision of the Phorusrhacidae (Aves: Ralliformes). Pap. Avulsos Zool. 2003, 43, 55–91. [Google Scholar] [CrossRef] [Green Version]
- Moreno, F.P.; Mercerat, A. Paleontología Argentina I. Catálogo Pájaros Rep. Argent. Conserv. Mus. Plata 1891, 1, 8–71. [Google Scholar]
- Angst, D.; Chinsamy, A. Ecological implications of the revised locomotory habits of the giant extinct South American birds (Phorusrhacidae and Brontornithidae). Contrib. Mus. Argent. Cienc. Nat. 2017, 7, 17–38. [Google Scholar]
- Angst, D.; Buffetaut, E.; Lecuyer, C.; Amiot, R. A new method for estimating locomotion type in large ground birds. Palaeontology 2015, 59, 217–223. [Google Scholar] [CrossRef]
- Kraglievich, L. Una gigantesca ave fósil del Uruguay, Devincenzia gallinali n. gén. n. sp., tipo de una nueva familia Devincenziidae del orden Stereornithes. Anal. Mus. Hist. Nat. Montev. 1932, 2, 323–353. [Google Scholar]
- Tonni, E. El rol ecológico de algunas aves fororracoideas. Ameghiniana 1977, 14, 316–317. [Google Scholar]
- Agnolín, F.L. Brontornis burmeisteri Moreno & Mercerat, un Anseriformes (Aves) gigante del Mioceno medio de Patagonia, Argentina. Rev. Mus. Argent. Cienc. Nat. 2007, 9, 15–25. [Google Scholar]
- Ameghino, F. Sur les oiseaux fossiles de Patagonie. Bol. Inst. Geogr. Argent. 1895, 15, 501–602. [Google Scholar]
- de Sáez, M.D. Las aves corredoras fósiles del Santacrucense. Anal. Soc. Cient. Argent. 1927, 103, 145–160. [Google Scholar]
- Kraglievich, L. Contribución al conocimiento de las aves fósiles de la época arauco-entrerriana. Physis 1931, 10, 304–315. [Google Scholar]
- Brodkorb, P. Catalogue of fossil birds. Part III (Gruiformes, Ichthyornithiformes, Charadriiformes). Bull. Flo. State Mus. 1967, 11, 99–220. [Google Scholar]
- Lambrecht, K. Handbuch der Palaeornithologie; Gebrüder Borntraeger: Berlin, Germany, 1933; 988p. [Google Scholar]
- Patterson, B.; Kraglievich, J.L. Sistemática y nomenclatura de las Aves Fororracoideas del Plioceno Argentino. Publ. Mus. Cienc. Nat. Tradic. Mar. Plata 1960, 1, 1–52. [Google Scholar]
- Agnolin, F.L. Sistemática y Filogenia de las aves Fororracoideas (Gruiformes, Cariamae); de Azara, F., Mazzini, V., Eds.; Fundación de Historia Natural: Buenos Aires, Argentina, 2009; p. 82. Available online: https://www.fundacionazara.org.ar/img/libros/sistematica-y-filogenia-de-aves-fororracoideas-ok.pdf (accessed on 19 January 2021).
- Agnolin, F. La posición sistemática de Hermosiornis (Aves, Phororhacoidea) y sus implicancias filogenéticas. Rev. Mus. Argent. Cienc. Nat. 2013, 15, 39–60. [Google Scholar] [CrossRef] [Green Version]
- Angst, D.; Buffetaut, É. Palaeobiology of Giant Flightless Birds; Elsevier: Amsterdam, The Netherlands; ISTE Press: Great Britain, UK, 2017; 282p. [Google Scholar]
- Buffetaut, E. Tertiary ground birds from Patagonia (Argentina) in the Tournouër collection of the Muséum National d’Histoire Naturelle, Paris. Bull. Soc. Géol. Franc. 2014, 185, 207–214. [Google Scholar] [CrossRef]
- Buffetaut, E. A reassessment of the giant birds Liornis floweri Ameghino, 1895 and Callornis giganteus Ameghino, 1895, from the Santacrucian (late Early Miocene) of Argentina. Palaeovertebrata 2016, 40, 3. [Google Scholar] [CrossRef]
- Mayr, G. Paleogene Fossil Birds; Springer International Publishing: Berlin/Heidelberg, Germany, 2009; 262p. [Google Scholar]
- Tambussi, C.P. Palaeoenvironmental and faunal inferences based on the avian fossil record of Patagonia and Pampa: What works and what does not. Biol. J. Linn. Soc. 2011, 103, 458–474. [Google Scholar] [CrossRef] [Green Version]
- Tambussi, C.P.; Degrange, F. South American and Antarctic Continental Cenozoic Birds. In Paleobiogeographical Affinities and Disparities; Springer: Dordrecht, Switzerlamd, 2013; 114p. [Google Scholar]
- Alvarenga, H.M.F.; Chiappe, L.; Bertelli, S. Phorusrhacids: The terror birds. In Living Dinosaurs. The Evolutionary History of Modern Birds; Dyke, G., Kaiser, G., Eds.; John Wiley & Sons, Ltd.: London, UK, 2011; pp. 187–208. [Google Scholar]
- Worthy, T.H.; Degrange, F.J.; Handley, W.D.; Lee, M.S. The evolution of giant flightless birds and novel phylogenetic relationships for extinct fowl (Aves, Galloanseres). R. Soc. Open Sci. 2017, 4, 170975. [Google Scholar] [CrossRef] [Green Version]
- Agnolín, F.L. Posición sistemática de algunas aves fororracoideas (Gruiformes; Cariamae) Argentinas. Rev. Mus. Argent. Cienc. Nat. 2006, 8, 27–33. [Google Scholar]
- Buffetaut, E. Phororhacoidea or Phorusrhacoidea? A note on the nomenclature of the “terror birds”. Ann. Paléontol. 2013, 99, 157–161. [Google Scholar] [CrossRef]
- Baumel, J.J.; Witmer, L.M. Osteology. In Handbook of Avian Anatomy: Nomina Anatomica Avium; Baumel, J.J., King, A.S., Brazile, J.E., Evans, H.E., Van den Berge, J.C., Eds.; Nuttal Ornithological Club: Cambridge, MA, USA, 1993; pp. 45–132. [Google Scholar]
- Zinoviev, A.V. Notes on pelvic and hindlimb myology and syndesmology of Emeus crassus and Dinornis robustus (Aves: Dinornithiformes). In Proceedings of the 8th International Meeting of the Society of Avian Paleontology and Evolution, Vienna, Austria, 11–16 June 2012; Verlag Naturhistorisches Museum Wien: Wien, Austria, 2013; Volume 2013, pp. 253–278. [Google Scholar]
- Goloboff, P.A.; Catalano, S.A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Hospitaleche, C.A.; Tambussi, C.P.; Reguero, M. Catálogo de los Tipos de aves fósiles del Museo de La Plata. Serie Técnica y Didáctica del Museo de La Plata 2001, 41, 1–28. [Google Scholar]
- Ono, K. A bony-toothed bird from the Middle Miocene, Chichibu Basin, Japan. Bull. Nat. Sci. Mus. 1989, 15, 33–38. [Google Scholar]
- Alvarenga, H.M. Paraphysornis novo gênero para Physornis brasiliensis Alvarenga, 1982 (Aves: Phorusrhacidae). An. Acad. Bras. Ciênc. 1993, 65, 403–406. [Google Scholar]
- Buffetaut, E. A brontornithid from the Deseadan (Oligocene) of Bolivia. Contrib. Mus. Argent. Cienc. Nat. 2017, 7, 39–47. [Google Scholar]
- McGowan, C. Tarsal development in birds: Evidence for homology with the theropod condition. J. Zool. 1985, 206, 53–67. [Google Scholar] [CrossRef]
- Agnolín, F.L.; Martinelli, A.G. Fossil birds from the Late Cretaceous Los Alamitos Formation, Río Negro Province, Argentina. J. South Am. Earth Sci. 2009, 27, 42–49. [Google Scholar] [CrossRef]
- Ameghino, F. L’âge des formations sédimentaires de Patagonie. Anal. Soc. Cient. Argent. 1901, 51–52, 65–91. [Google Scholar]
- Alvarenga, H.M. Revisao Sistemática das aves Phorusrhacidae. Ph.D. Thesis, Universidade de Sao Paulo, Sao Paulo, Brazil, 1999; 95p. [Google Scholar]
- Bertelli, S.; Chiappe, L.M.; Tambussi, C. A new phorusrhacid (Aves: Cariamae) from the middle Miocene of Patagonia, Argentina. J. Vertebr. Paléontol. 2007, 27, 409–419. [Google Scholar] [CrossRef]
- Agnolín, F.L.; Chafrat, P. New fossil bird remains from the Chichinales Formation (Early Miocene) of northern Patagonia, Argentina. Ann. Paléontol. 2015, 101, 87–94. [Google Scholar] [CrossRef]
- Andrews, C.W. On the Extinct Birds of Patagonia-I. The Skull and Skeleton of Phororhacos inflatus Ameghino. Trans. Zool. Soc. Lond. 2010, 15, 55–86. [Google Scholar] [CrossRef]
- Worthy, T.H.; Handley, W.D.; Archer, M.; Hand, S.J. The extinct flightless mihirungs (Aves, Dromornithidae): Cranial anatomy, a new species, and assessment of Oligo-Miocene lineage diversity. J. Vertebr. Paléontol. 2016, 36, e1031345. [Google Scholar] [CrossRef]
- Elzanowski, A.; Stidham, T.A. Morphology of the quadrate in the Eocene anseriform Presbyornis and extant galloanserine birds. J. Morphol. 2010, 271, 305–323. [Google Scholar] [PubMed]
- Murray, P.F.; Megirian, D. The skull of dromornithid birds: Anatomical evidence for their relationship to Anseriformes. Rec. South Aust. Mus. 1998, 31, 51–97. [Google Scholar]
- Bourdon, E. The Pseudo-toothed Birds (Aves, Odontopterygiformes) and their bearing on the early evolution of modern birds. In Living Dinosaurs. The Evolutionary History of Modern Birds; Dyke, G., Kaiser, G., Eds.; John Wiley & Sons, Ltd.: London, UK, 2011; pp. 209–234. [Google Scholar]
- Martin, L.D. The Status of the Late Paleocene Birds Gastornis and Remiornis; Science Series; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1992; Volume 36, pp. 97–108. [Google Scholar]
- Buffetaut, E.; Angst, D. Stratigraphic distribution of large flightless birds in the Palaeogene of Europe and its palaeobiological and palaeogeographical implications. Earth Sci. Rev. 2014, 138, 394–408. [Google Scholar] [CrossRef]
- Mourer-Chauviré, C.; Bourdon, E. The Gastornis (Aves, Gastornithidae) from the Late Paleocene of Louvois (Marne, France). Swiss J. Palaeontol. 2015, 135, 327–341. [Google Scholar] [CrossRef]
- Hellmund, M. Reappraisal of the bone inventory of Gastornis geiselensis (Fischer, 1978) from the Eocene “Geiseltal Fossillagerstätte” (Saxony-Anhalt, Germany). N. Jahrb. Geol. Paläontol. Abh. 2013, 269, 203–220. [Google Scholar] [CrossRef]
- Andors, A. Reappraisal of the Eocene groundbird Diatryma (Aves: Anserimorphae). In Papers in Avian Paleontology Honoring Pierce Brodkorb; Science Series; Campbell, K.E., Ed.; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 1992; Volume 36, pp. 109–125. [Google Scholar]
- Matthew, W.D.; Granger, W. The skeleton of Diatryma, a gigantic bird from the Lower Eocene of Wyoming. Bull. Am. Mus. Nat. Hist. 1917, 37, 307–326. [Google Scholar]
- Rich, P.V. The Australian Dromornithidae: A group of large extinct ratites. Los Angel. Cty. Nat. Hist. Mus Contrib. Sci. 1980, 330, 93–103. [Google Scholar]
- Park, T.; Fitzgerald, E.M.G. A late Miocene–early Pliocene Mihirung bird (Aves: Dromornithidae) from Victoria, southeast Australia. Alcheringa Aust. J. Palaeontol. 2012, 36, 419–422. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agnolin, F.L. Reappraisal on the Phylogenetic Relationships of the Enigmatic Flightless Bird (Brontornis burmeisteri) Moreno and Mercerat, 1891. Diversity 2021, 13, 90. https://doi.org/10.3390/d13020090
Agnolin FL. Reappraisal on the Phylogenetic Relationships of the Enigmatic Flightless Bird (Brontornis burmeisteri) Moreno and Mercerat, 1891. Diversity. 2021; 13(2):90. https://doi.org/10.3390/d13020090
Chicago/Turabian StyleAgnolin, Federico L. 2021. "Reappraisal on the Phylogenetic Relationships of the Enigmatic Flightless Bird (Brontornis burmeisteri) Moreno and Mercerat, 1891" Diversity 13, no. 2: 90. https://doi.org/10.3390/d13020090
APA StyleAgnolin, F. L. (2021). Reappraisal on the Phylogenetic Relationships of the Enigmatic Flightless Bird (Brontornis burmeisteri) Moreno and Mercerat, 1891. Diversity, 13(2), 90. https://doi.org/10.3390/d13020090