
Fawn Counts and Adult Female Site Use Are Mismatched Indicators of Habitat Quality in an Endangered Deer


Abstract
:1. Introduction
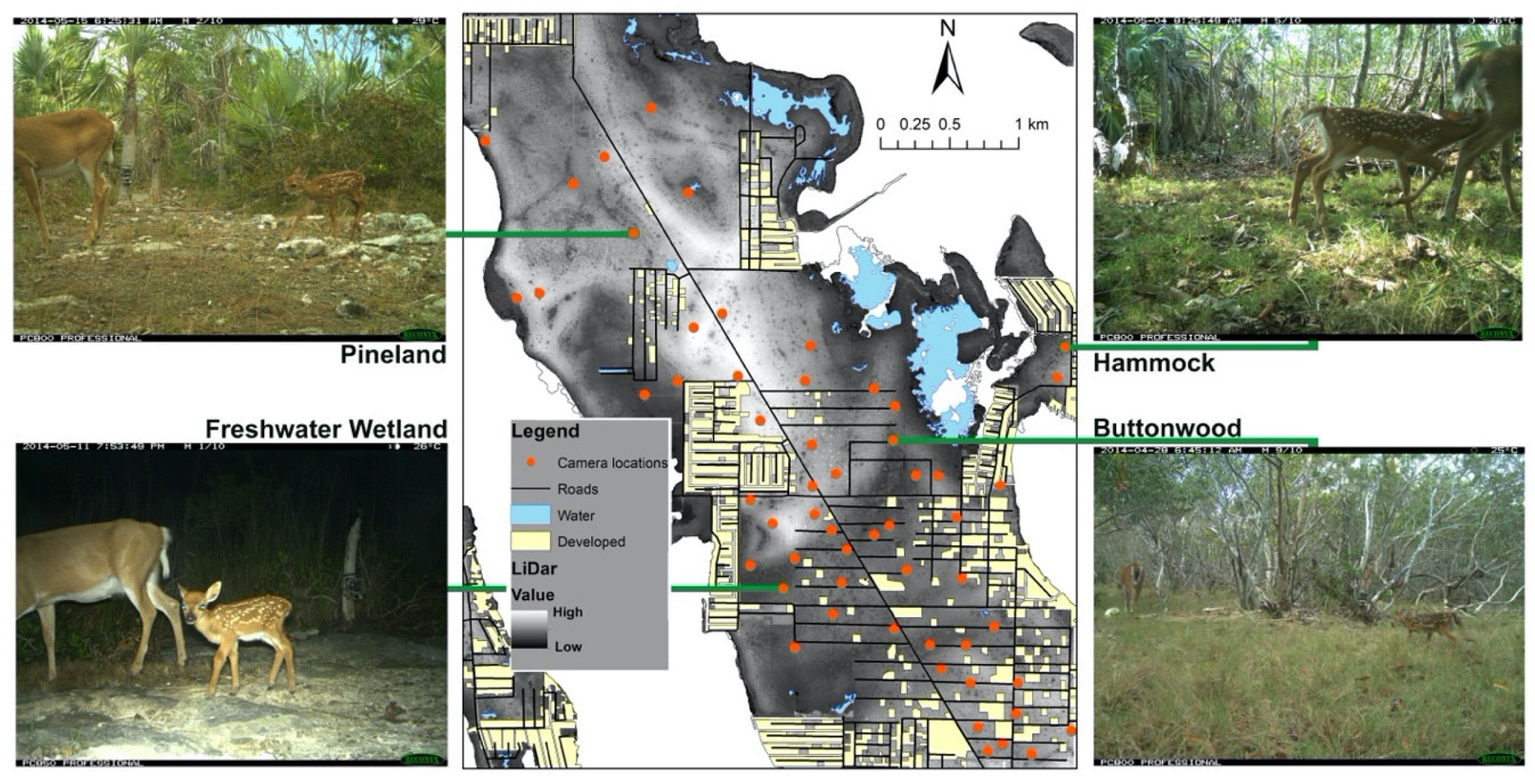
2. Materials and Methods
3. Results
3.1. Fawn Identification
3.2. Fawn Models
3.3. Doe Models
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ripple, W.J.; Newsome, T.M.; Wolf, C.; Dirzo, R.; Everatt, K.T.; Galetti, M.; Macdonald, D.W. Collapse of the world’s largest herbivores. Sci. Adv. 2015, 1, e1400103. [Google Scholar] [CrossRef] [Green Version]
- Schipper, J.; Chanson, J.S.; Chiozza, F.; Cox, N.A.; Hoffmann, M.; Katariya, V.; Lamoreux, J.; Rodrigues, A.S.L.; Stuart, S.N.; Temple, H.J.; et al. The Status of the World’s Land and Marine Mammals: Diversity, Threat, and Knowledge. Science 2008, 322, 225–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieger, D.J. Economic Value of Forest Ecosystem Services: A Review; The Wilderness Society: Washington, DC, USA, 2001. [Google Scholar]
- Milner, J.M.; Bonenfant, C.; Mysterud, A.; Gaillard, J.-M.; Csányi, S.; Stenseth, N.C. Temporal and spatial development of red deer harvesting in Europe: Biological and cultural factors. J. Appl. Ecol. 2006, 43, 721–734. [Google Scholar] [CrossRef]
- Jacobson, H.A.; Kroll, J.C.; Browning, R.W.; Koerth, B.H.; Conway, M.H. Infrared-triggered cameras for Censusing White-Tailed Deer. Wildl. Soc. Bull. 1997, 25, 547–556. [Google Scholar]
- Smart, J.C.R.; Ward, A.I.; White, P.C.L. Monitoring woodland deer populations in the UK: An imprecise science. Mammal Rev. 2004, 34, 99–114. [Google Scholar] [CrossRef]
- Lopez, R.R.; Silvy, N.J.; Wilkins, R.N.; Frank, P.A.; Peterson, M.J.; Peterson, M.N. Habitat-Use Patterns of Florida Key deer: Implications of Urban Development. J. Wildl. Manag. 2004, 68, 900–908. [Google Scholar]
- Lopez, R.R.; Harveson, P.M.; Peterson, M.N.; Silvy, N.J.; Frank, P.A. Changes in ranges of Florida Key deer—Does population density matter? Wildl. Soc. Bull. 2005, 33, 343–348. [Google Scholar] [CrossRef]
- Parker, I.D.; Lopez, R.R.; Silvy, N.J.; Davis, D.S.; Owen, C.B. Long-term effectiveness of US 1 crossing project in reducing florida key deer mortality. Wildl. Soc. Bull. 2011, 35, 296–302. [Google Scholar] [CrossRef]
- Parker, I.D.; Parker, J.; Montalvo, A.E.; Lopez, R.R.; Silvy, N.J.; Lund, A.A.; Barham, E.; Finn, D.S.; Crawford, M. Florida Key Deer Presence on Outer Islands Following New World Screwworm and Hurricane Irma. Southeast. Nat. 2020, 19, 20. [Google Scholar] [CrossRef]
- Lopez, R.R.; Silvy, N.J.; Pierce, B.L.; Frank, P.A.; Wilson, M.T.; Burke, K.M. Population Density of the Endangered Florida Key Deer. J. Wildl. Manag. 2004, 68, 570–575. [Google Scholar] [CrossRef]
- Villanova, V.L.; Hughes, P.T.; Hoffman, E.A. Combining genetic structure and demographic analyses to estimate persistence in endangered Key deer (Odocoileus virginianus clavium). Conserv. Genet. 2017, 18, 1061–1076. [Google Scholar] [CrossRef]
- Kays, R.; Arbogast, B.S.; Baker-Whatton, M.; Beirne, C.; Boone, H.M.; Bowler, M.; Burneo, S.F.; Cove, M.V.; Ding, P.; Espinosa, S.; et al. An empirical evaluation of camera trap study design: How many, how long and when? Methods Ecol. Evol. 2020, 11, 700–713. [Google Scholar] [CrossRef]
- O’Connell, A.F.; Nichols, J.D.; Karanth, K.U. (Eds.) Camera Traps in Animal Ecology: Methods and Analyses; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Zheng, X.; Owen, M.A.; Nie, Y.; Hu, Y.; Swaisgood, R.R.; Yan, L.; Wei, F. Individual identification of wild giant pandas from camera trap photos—A systematic and hierarchical approach. J. Zool. 2016, 300, 247–256. [Google Scholar] [CrossRef]
- Parsons, A.W.; Forrester, T.; McShea, W.J.; Baker-Whatton, M.C.; Millspaugh, J.J.; Kays, R. Do occupancy or detection rates from camera traps reflect deer density? J. Mammal. 2017, 98, 1547–1557. [Google Scholar] [CrossRef]
- Chandler, R.B.; Engebretsen, K.; Cherry, M.J.; Garrison, E.P.; Miller, K.V. Estimating recruitment from capture–recapture data by modelling spatio-temporal variation in birth and age-specific survival rates. Methods Ecol. Evol. 2018, 9, 2115–2130. [Google Scholar] [CrossRef]
- USFWS. Multi-Species Recovery Plan for South Florida; US Fish and Wildlife Service Report; USFWS: Vero Beach, FL, USA, 1999.
- Cove, M.V.; Gardner, B.; Simons, T.R.; O’Connell, A.F. Co-occurrence dynamics of endangered Lower Keys marsh rabbits and free-ranging domestic cats: Prey responses to an exotic predator removal program. Ecol. Evol. 2018, 8, 4042–4052. [Google Scholar] [CrossRef] [Green Version]
- Cove, M.V.; Gardner, B.; Simons, T.R.; Kays, R.; O’Connell, A.F. Free-ranging cats (Felis catus) on public lands: Estimating density, movement, activity and diet. Biol. Invasions 2018, 20, 333–344. [Google Scholar] [CrossRef]
- Bolger, D.T.; Morrison, T.A.; Vance, B.; Lee, D.; Farid, H. A computer-assisted system for photographic mark-recapture analysis. Methods Ecol. Evol. 2012, 3, 813–822. [Google Scholar] [CrossRef]
- Saha, S.; Bradley, K.; Ross, M.S.; Hughes, P.; Wilmers, T.; Ruiz, P.L.; Bergh, C. Hurricane effects on subtropical pine rocklands of the Florida Keys. Clim. Chang. 2011, 107, 169–184. [Google Scholar] [CrossRef]
- Peterson, M.N.; Lopez, R.R.; Frank, P.A.; Porter, B.A.; Silvy, N.J. Key deer fawn response to urbanization: Is sustainable development possible? Wildl. Soc. Bull. 2004, 32, 493–499. [Google Scholar] [CrossRef]
- Royle, J.A.; Dorazio, R.M. Hierarchical Modeling and Inference in Ecology: The Analysis of Data from Populations, Metapopulations and Communities; Elsevier: London, UK, 2008. [Google Scholar]
- Keever, A.C.; McGowan, C.P.; Ditchkoff, S.S.; Acker, P.K.; Grand, J.B.; Newbolt, C.H. Efficacy of N-mixture models for surveying and monitoring white-tailed deer populations. Mammal Res. 2017, 62, 413–422. [Google Scholar] [CrossRef]
- Burnham, K.; Anderson, D.R. Model Selection and Multi-Model Inference, 2nd ed.; Springer: New York, NY, USA, 2002; p. 63. [Google Scholar]
- Fiske, I.; Chandler, R. Unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef] [Green Version]
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Chitwood, M.C.; Lashley, M.A.; Kilgo, J.C.; Cherry, M.J.; Conner, L.M.; Vukovich, M.; Ray, H.S.; Ruth, C.; Warren, R.J.; DePerno, C.S.; et al. Are camera surveys useful for assessing recruitment in white-tailed deer? Wildl. Biol. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Norouzzadeh, M.S.; Nguyen, A.; Kosmala, M.; Swanson, A.; Palmer, M.S.; Packer, C.; Clune, J. Automatically identifying, counting, and describing wild animals in camera-trap images with deep learning. Proc. Natl. Acad. Sci. USA 2018, 115, E5716–E5725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huegel, C.N.; Gladfelter, H.L.; Dahlgren, R.B. Bedsite Selection by White-Tailed Deer Fawns in Iowa. J. Wildl. Manag. 1986, 50, 474. [Google Scholar] [CrossRef]
- Chitwood, M.C.; Lashley, M.A.; Kilgo, J.C.; Pollock, K.H.; Moorman, C.E.; DePerno, C.S. Do biological and bedsite characteristics influence survival of neonatal white-tailed deer? PLoS ONE 2015, 10, E0119070. [Google Scholar] [CrossRef]
- Chitwood, M.C.; Lashley, M.A.; Moorman, C.E.; Deperno, C.S. Setting an evolutionary trap: Could the hider strategy be maladaptive for white-tailed deer? J. Ethol. 2017, 35, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cove, M.V.; Maurer, A.S.; O’Connell, A.F. Camera traps reveal an apparent mutualism between a common mesocarnivore and an endangered ungulate. Mamm. Biol. 2017, 87, 143–145. [Google Scholar] [CrossRef]
- Gulsby, W.D.; Kilgo, J.C.; Vukovich, M.; Martin, J.A. Landscape heterogeneity reduces coyote predation on white-tailed deer fawns. J. Wildl. Manag. 2017, 81, 601–609. [Google Scholar] [CrossRef]
- Nardone, A.; Ronchi, B.; Lacetera, N.; Ranieri, M.; Bernabucci, U. Effects of climate changes on animal production and sustainability of livestock systems. Livest. Sci. 2010, 130, 57–69. [Google Scholar] [CrossRef]
- Taillie, P.J.; Jolly, S.R.; Bobay, L.R.; Sneckenberger, S.; Mccleery, R.A. Habitat use across multiple scales suggests resilience to rising seas for endangered island endemic compared to sympatric invasive species. Anim. Conserv. 2020. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Model | K | AICc | ΔAICc | ω | LL | Estimated β Coefficients (SE) |
|---|---|---|---|---|---|---|
| elevation | 4 | 296.46 | 0 | 0.23 | −143.84 | βe = −0.956 (0.351) |
| anthropogenic | 5 | 296.95 | 0.49 | 0.18 | −142.88 | βt = −0.601 (0.351) βd = 0.002 (0.001) |
| development | 4 | 297.34 | 0.88 | 0.15 | −144.28 | βd = 0.002 (0.001) |
| null NB | 3 | 298.29 | 1.82 | 0.09 | −145.91 | − |
| trail | 4 | 299.65 | 3.19 | 0.05 | −145.43 | βt = −0.335 (0.341) |
| road | 4 | 300.5 | 4.04 | 0.03 | −145.86 | βr = 0.000 (0.001) |
| Model | K | AICc | ΔAICc | ω | LL | Estimated β Coefficients (SE) |
|---|---|---|---|---|---|---|
| elevation | 5 | 798.10 | 0 | 0.37 | −393.45 | βe = 0.456 (0.256) |
| null NB | 4 | 798.89 | 0.79 | 0.25 | −395.05 | − |
| road | 5 | 799.54 | 1.44 | 0.18 | −394.17 | βr = 0.001 (0.001) |
| development | 5 | 800.71 | 2.61 | 0.1 | −394.76 | βd = 0.001 (0.000) |
| trail | 5 | 801.10 | 3.00 | 0.08 | −394.95 | βt = 0.110 (0.237) |
| anthropogenic | 6 | 803.21 | 5.11 | 0.03 | −394.75 | βt = 0.009 (0.256) βd = 0.001 (0.001) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacobs, J.G.; Lashley, M.A.; Cove, M.V. Fawn Counts and Adult Female Site Use Are Mismatched Indicators of Habitat Quality in an Endangered Deer. Diversity 2021, 13, 92. https://doi.org/10.3390/d13020092
Jacobs JG, Lashley MA, Cove MV. Fawn Counts and Adult Female Site Use Are Mismatched Indicators of Habitat Quality in an Endangered Deer. Diversity. 2021; 13(2):92. https://doi.org/10.3390/d13020092
Chicago/Turabian StyleJacobs, Julia G., Marcus A. Lashley, and Michael V. Cove. 2021. "Fawn Counts and Adult Female Site Use Are Mismatched Indicators of Habitat Quality in an Endangered Deer" Diversity 13, no. 2: 92. https://doi.org/10.3390/d13020092
APA StyleJacobs, J. G., Lashley, M. A., & Cove, M. V. (2021). Fawn Counts and Adult Female Site Use Are Mismatched Indicators of Habitat Quality in an Endangered Deer. Diversity, 13(2), 92. https://doi.org/10.3390/d13020092