
Alternative Development Strategies of Clinostomum chabaudi (Digenea) Metacercariae in Frog Hosts (Hyperolius spp.)




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
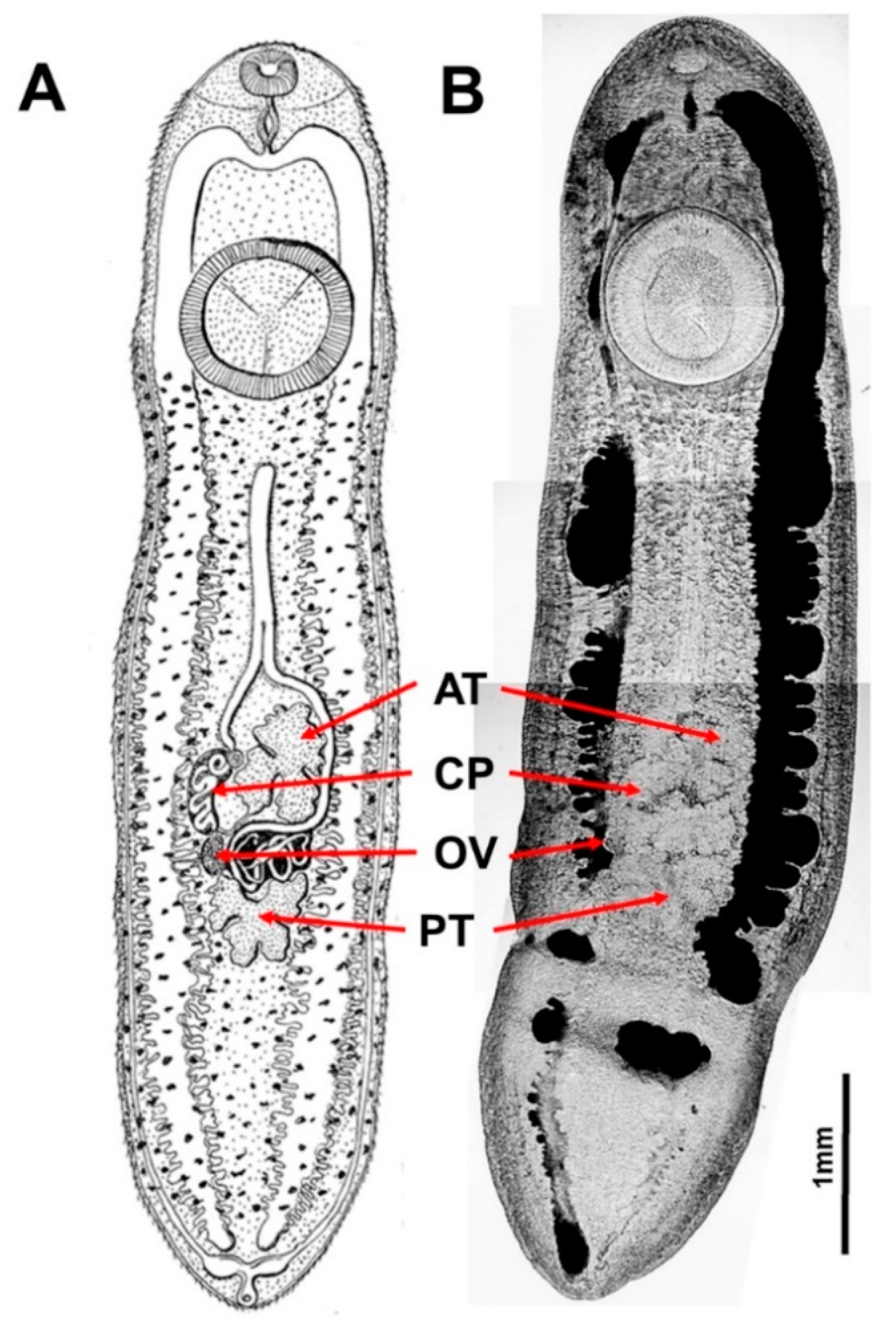
2.2. Morphological Examination of Metacercariae
2.3. Morphological and Demographic Traits of Frog Hosts
2.4. Statistical Procedures
2.5. Nucleic Acid Extraction and Polymerase Chain Reactions
2.6. Sequencing and Phylogenetic Analysis
3. Results
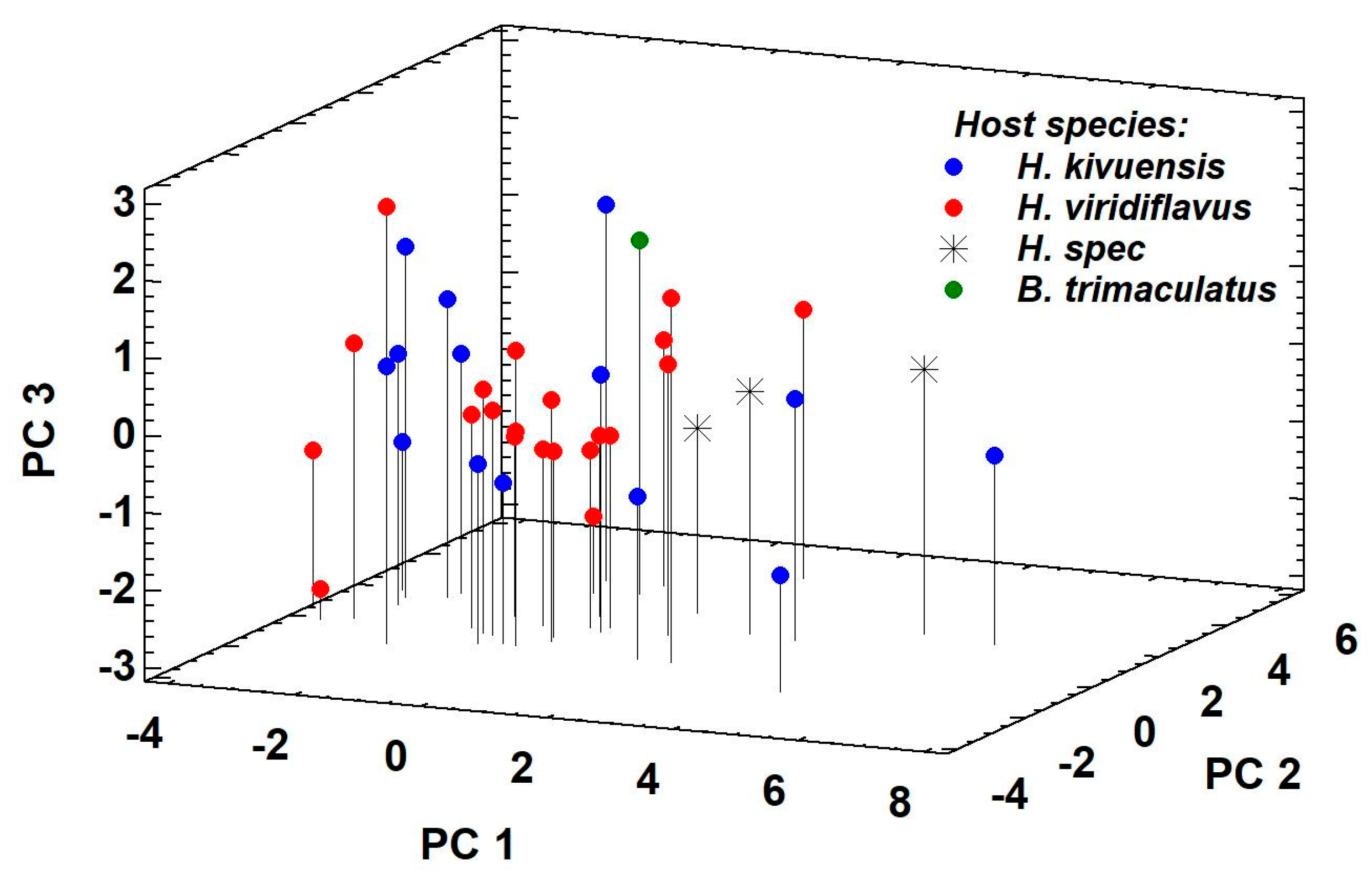
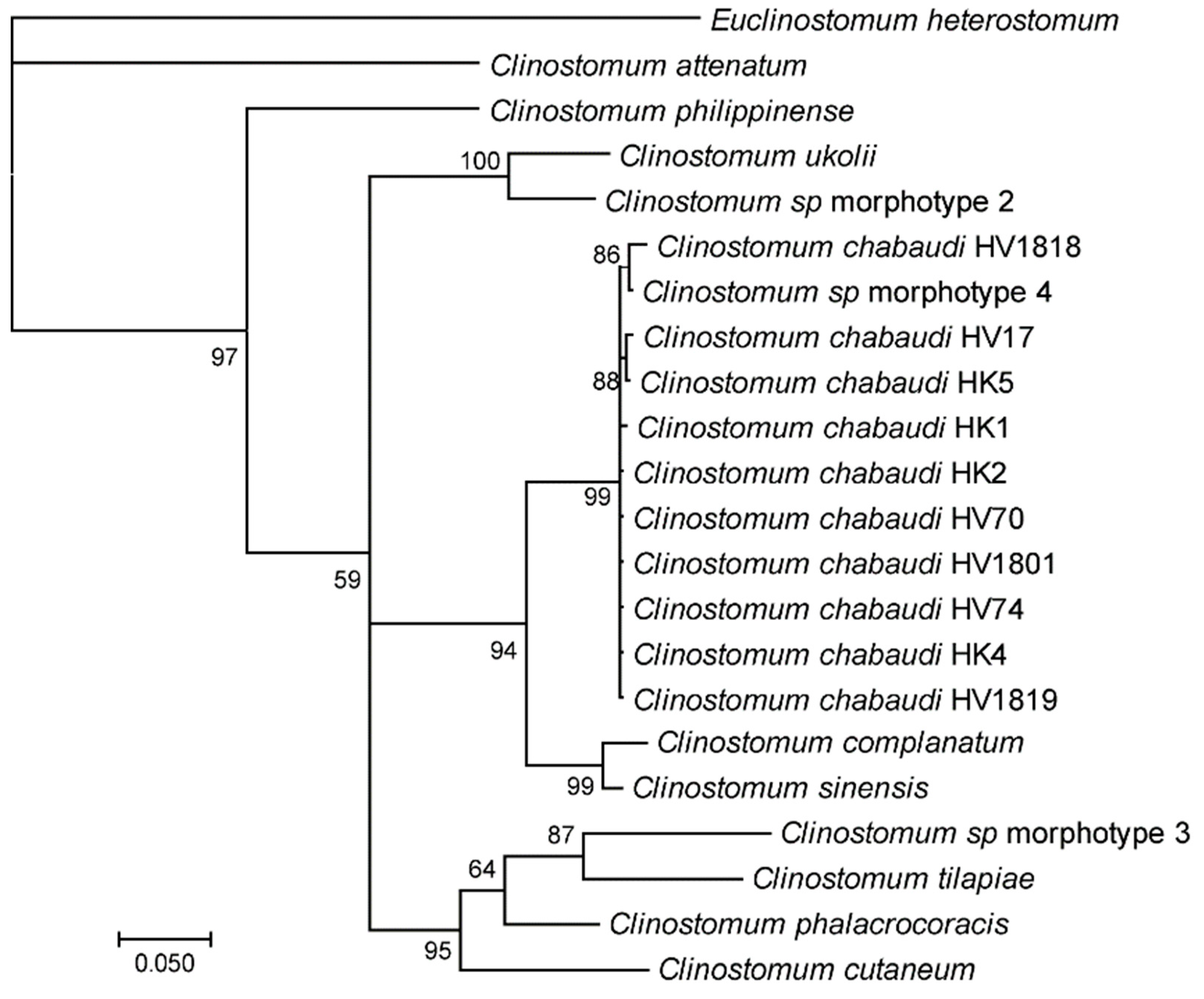
3.1. Morphological and Molecular Identification of Clinostomum Metacercaria
3.2. Infestation Ecology of C. chabaudi Metacercaria
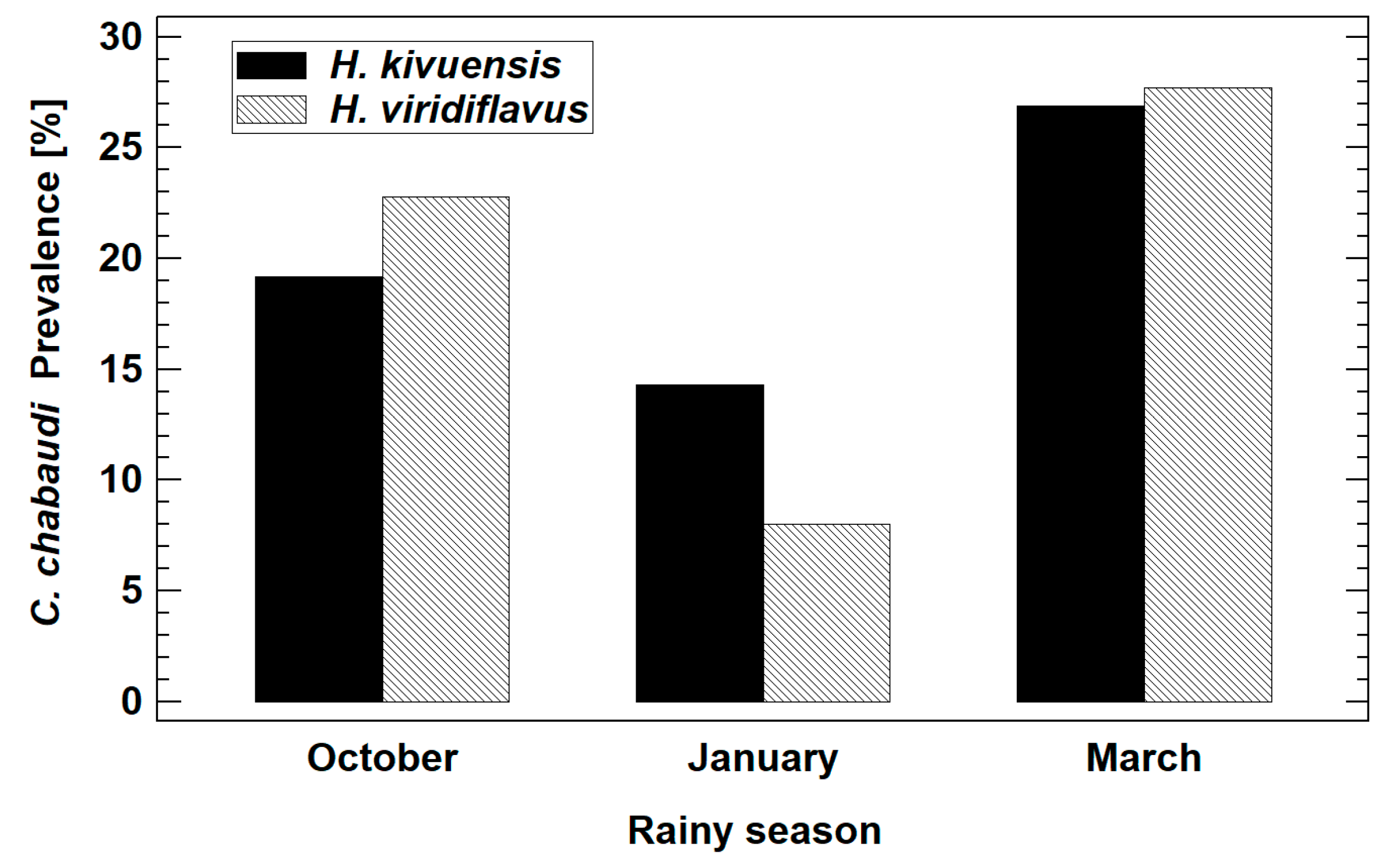
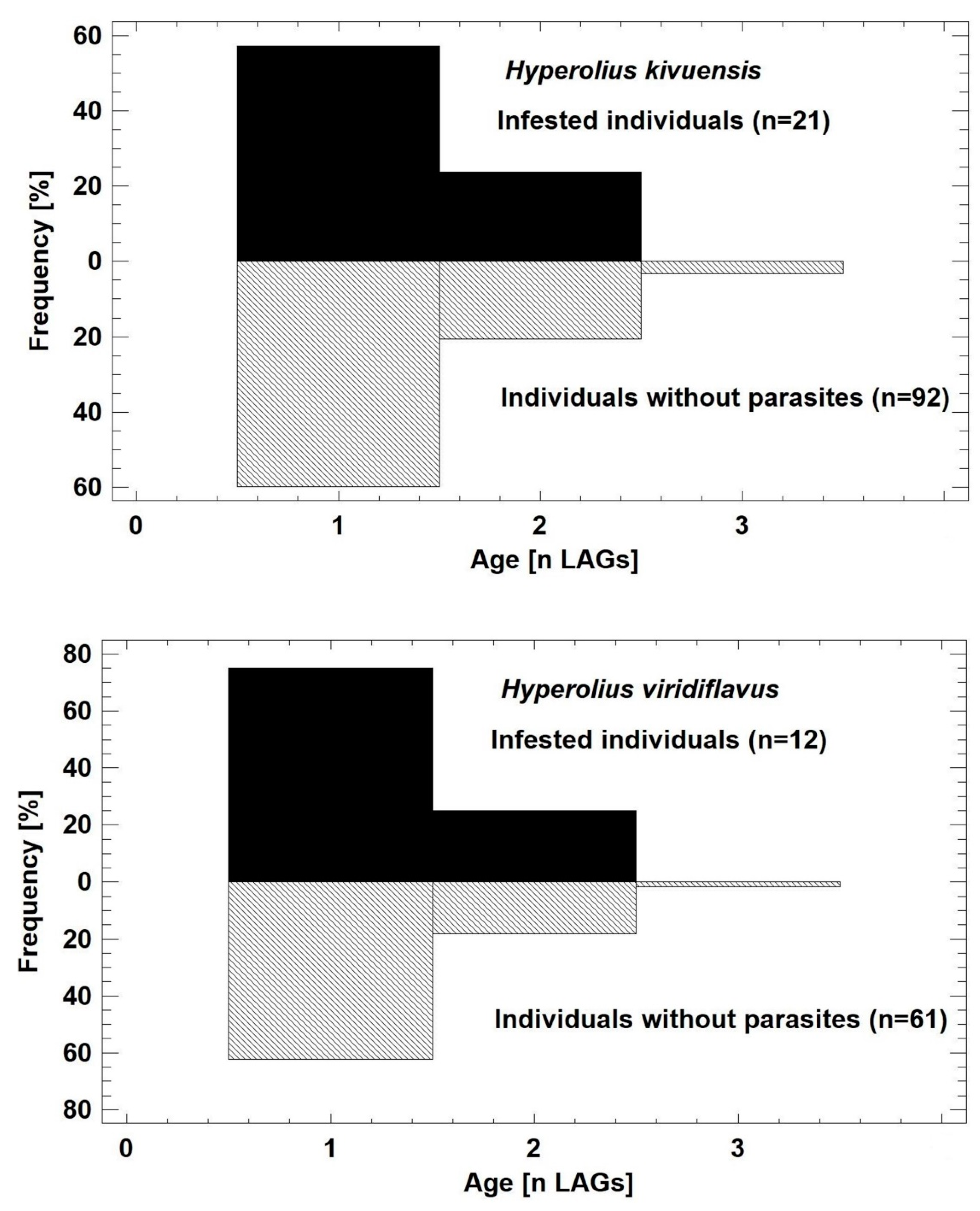
3.2.1. Prevalence and Infestation Intensity
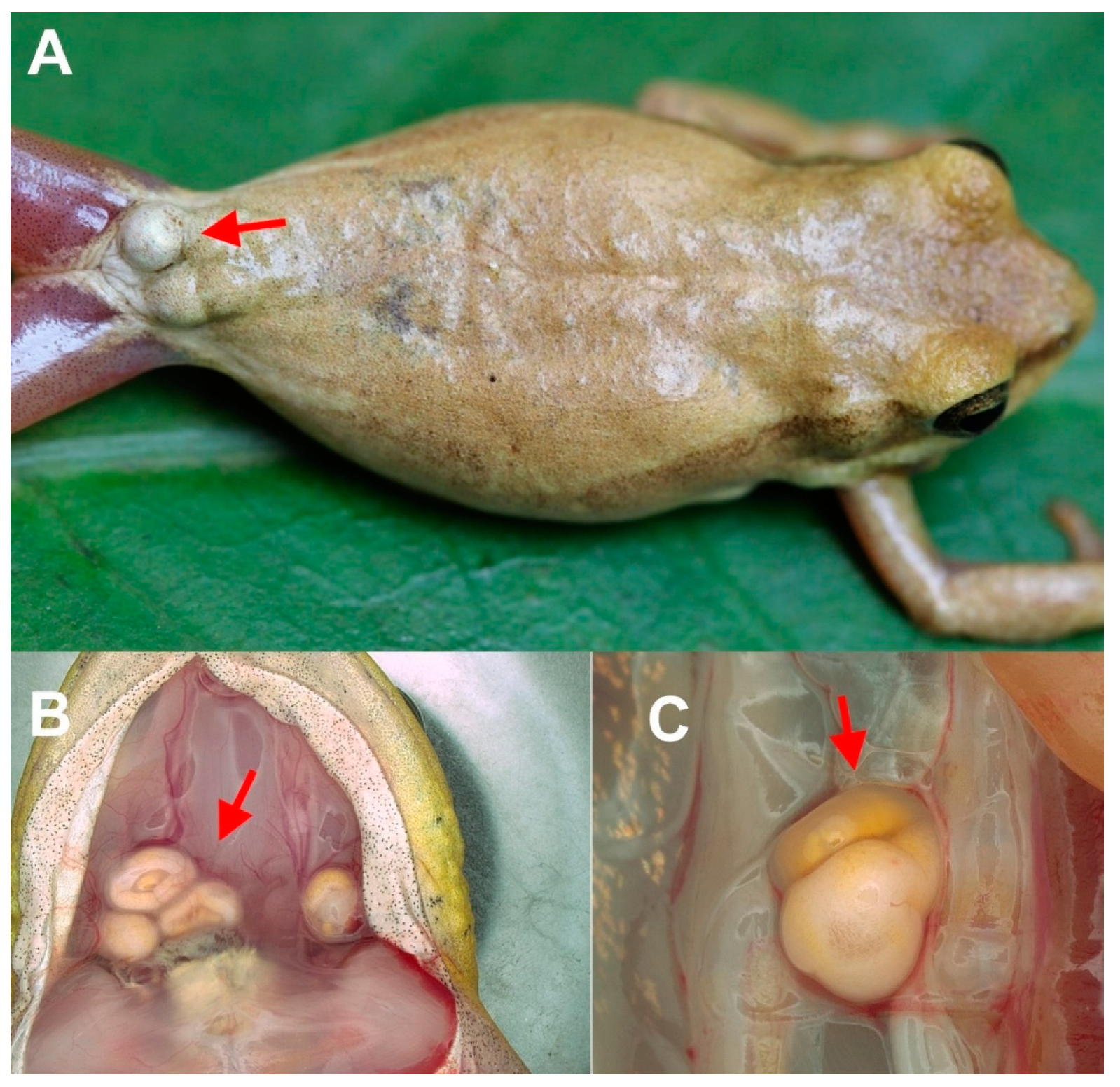
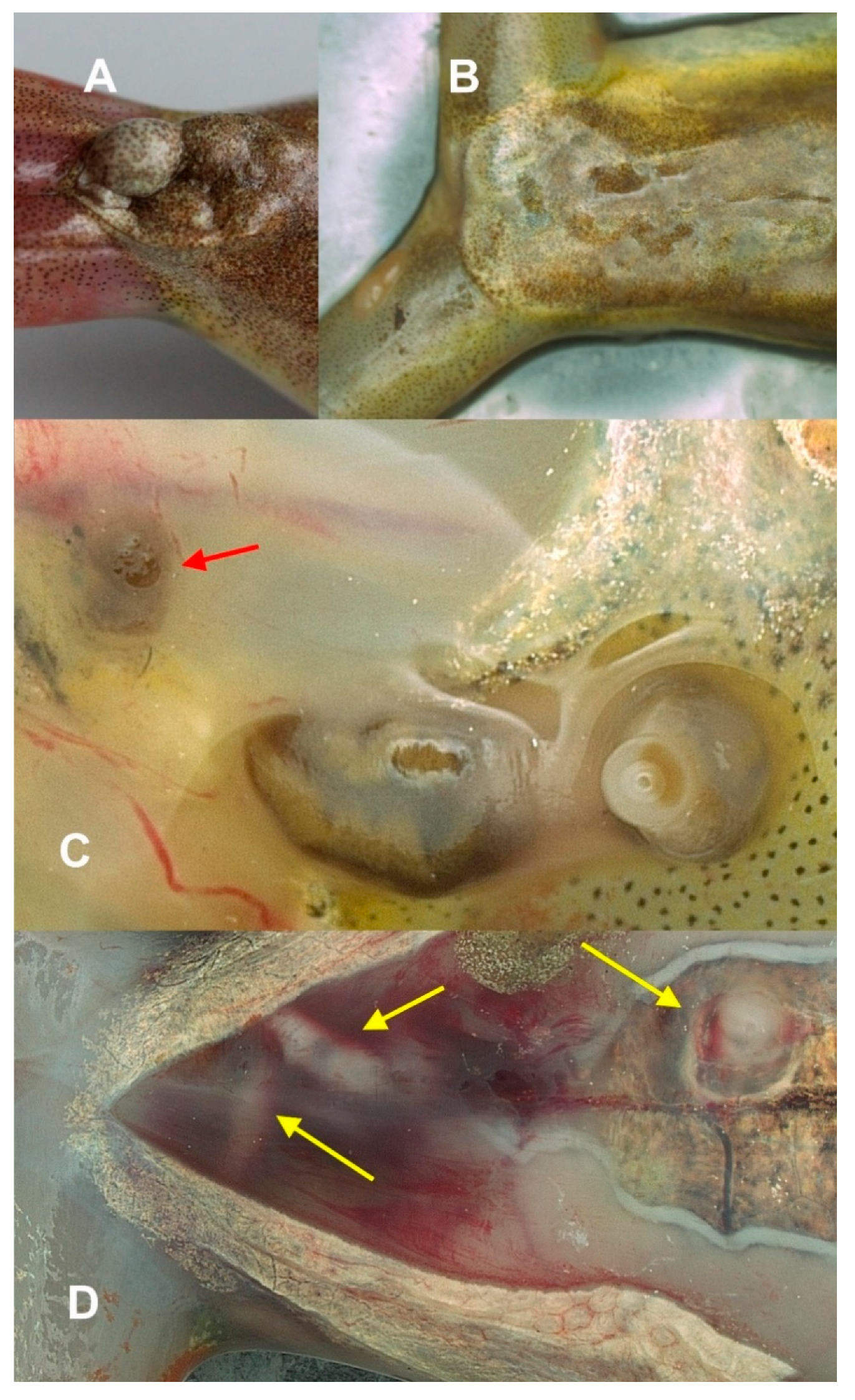
3.2.2. Metacercaria Location in the Host
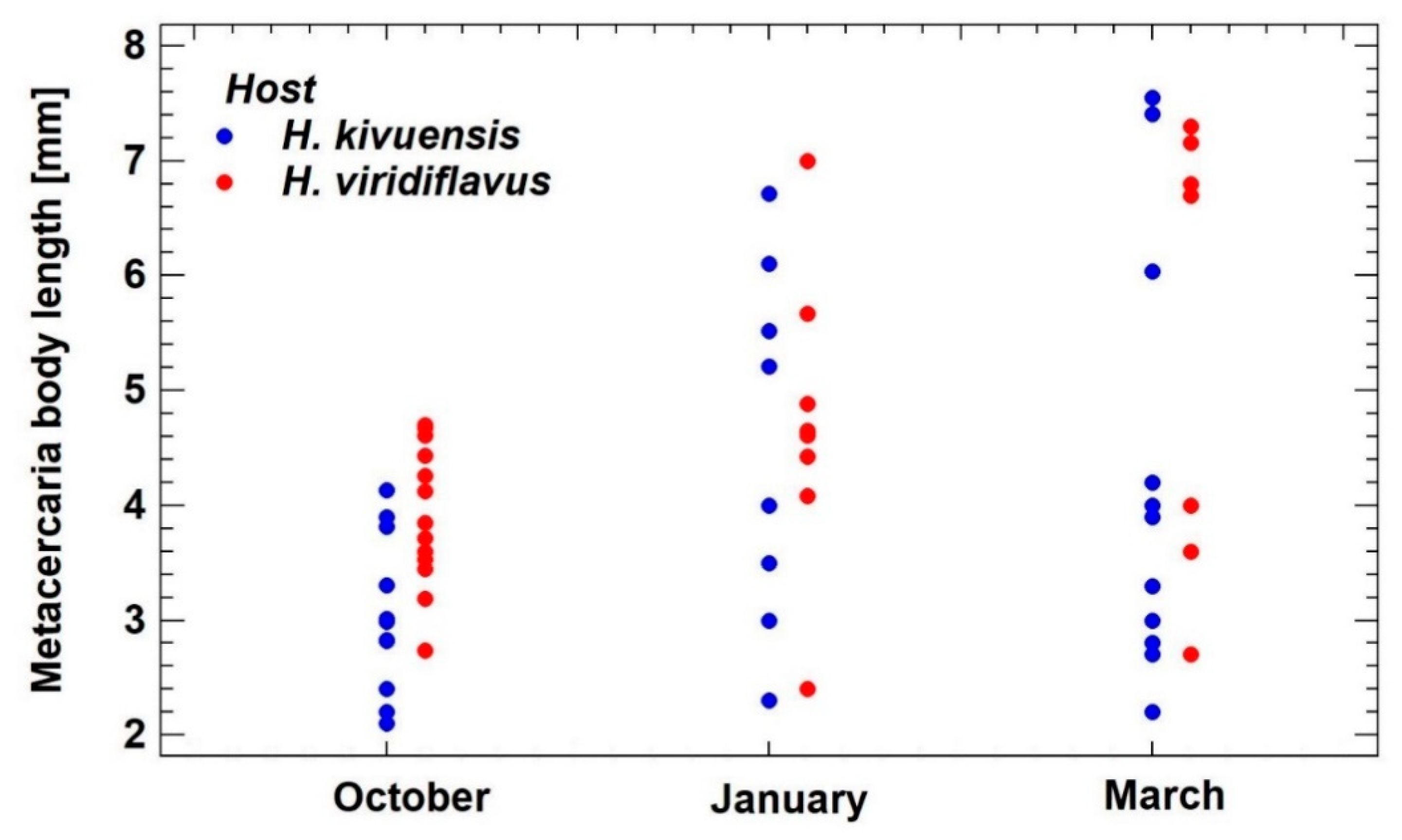
3.2.3. Metacercaria Growth
3.3. Morphological and Demographic Traits of Frog Hosts
4. Discussion
4.1. Taxonomic Identification of Clinostomum Metacercariae Infesting Reed Frogs in Rwanda
4.2. Alternative Metacercaria Life-Histories in Reed Frogs
4.3. Metacercaria Impact on the Life-History of Reed Frogs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kanev, I.; Radev, V.; Fried, B. Family Clinostomidae Lühe, 1901. In Keys to the Trematoda; Gibson, D.I., Jones, A., Bray, R.A., Eds.; CAB International and the Natural History Museum: Wallingford, UK, 2002; Volume 1, pp. 113–120. [Google Scholar]
- Lo, C.F.; Huber, F.; Kou, G.H.; Lo, C.J. Studies of Clinostomum complanatum (RUD., 1819). Fish Pathol. 1981, 15, 219–227. [Google Scholar] [CrossRef]
- Aohagi, Y.; Shibahara, T.; Machida, N.; Yamaga, Y.; Kagota, K.; Hayashi, T. Natural infections of Clinostomum complanatum (Trematoda, Clinostomatidae) in wild herons and egrets, Tottori prefecture, Japan. J. Wildl. Dis. 1992, 28, 470–471. [Google Scholar] [CrossRef] [Green Version]
- Dias, M.L.; Eiras, J.C.; Machado, M.H.; Souza, G.T.; Pavanelli, G.C. The life cycle of Clinostomum complanatum Rudolphi, 1814 (Digenea, Clinostomidae) on the floodplain of the high Parana river, Brazil. Parasitol. Res. 2003, 89, 506–508. [Google Scholar] [CrossRef]
- Caffara, M.; Davidovich, N.; Falk, R.; Smirnov, M.; Ofek, T.; Cummings, D.; Gustinelli, A.; Fioravanti, M.L. Redescription of Clinostomum phalacrocoracis metacercariae (Digenea: Clinostomidae) in cichlids from Lake Kinneret, Israel. Parasite. 2014, 21. [Google Scholar] [CrossRef] [Green Version]
- Caffara, M.; Locke, S.A.; Gustinelli, A.; Marcogliese, D.J.; Fioravanti, M.L. Morphological and molecular differentiation of Clinostomum complanatum and Clinostomum marginatum (Digenea: Clinostomidae) metacercariae and adults. J. Parasitol. 2011, 97, 884–891. [Google Scholar] [CrossRef]
- Caffara, M.; Locke, S.A.; Echi, P.C.; Halajian, A.; Benini, D.; Luus-Powell, W.J.; Tavakol, S.; Fioravanti, M.L. A morphological and molecular study of Clinostomid metacercariae from African fish with a redescription of Clinostomum tilapiae. Parasitol. 2017, 144, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Locke, S.A.; Caffara, M.; Marcogliese, D.J.; Fioravanti, M.L. A large-scale molecular survey of Clinostomum (Digenea, Clinostomidae). Zool. Scr. 2015, 44, 203–217. [Google Scholar] [CrossRef]
- Caffara, M.; Locke, S.A.; Echi, P.C.; Halajian, A.; Luus-Powell, W.J.; Benini, D.; Tedesco, P.; Fioravanti, M.L. A new species of Clinostomum Leidy, 1856 based on molecular and morphological analysis of metacercariae from African siluriform fishes. Parasitol. Res. 2020, 119, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Locke, S.A.; Caffara, M.; Barčák, D.; Sonko, P.; Tedesco, P.; Fioravanti, M.L.; Li, W. A new species of Clinostomum Leidy, 1856 in East Asia based on genomic and morphological data. Parasitol. Res. 2019, 118, 3253–3265. [Google Scholar] [CrossRef] [PubMed]
- Vercammen-Grandjean, P.H. Les Trématodes du Lac Kivu sud (Vermes). Annales Musée Royale de l’Afrique Centrale 1960, 5, 1–171. [Google Scholar]
- Bush, A.O.; Fernández, J.C.; Esch, G.W.; Seed, J.R.; Ndez, J.C.F. Parasitism: The Diversity and Ecology Animal Parasites; University Press: Cambridge, UK, 2001. [Google Scholar]
- Nicola, D.N.; Affonso, I.P.; Takemoto, R.M. Behavioral changes in host foraging: Experiments with Clinostomum (Trematoda, Digenea) parasitizing Loricariichthys platymetopon (Loricariidae). Exp. Parasitol. 2020, 216. [Google Scholar] [CrossRef] [PubMed]
- Hazen, T.C.; Esch, G.W. Observations on ecology of Clinostomum marginatum in largemouth bass (Micropterus salmoides). J. Fish Biol. 1978, 12, 411–420. [Google Scholar] [CrossRef]
- Silva-Souza, A.T.; Ludwig, G. Parasitism of Cichlasoma paranaense Kullander, 1983 and Gymnotus carapo Linnaeus, 1814 by Clinostomum complanatum (Rudolphi, 1814) Metacercariae in the Taquari river. Braz. J. Biol. 2005, 65, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Li, B.F.; Liu, X.-H.; Ge, H.-L.; Xie, C.-Y.; Cai, R.-Y.; Hu, Z.-C.; Zhang, Y.-G.; Wang, Z.-J. The discovery of Clinostomum complanatum metacercariae in farmed Chinese sucker, Myxocyprinus Asiaticus. Aquac. 2018, 495, 273–280. [Google Scholar] [CrossRef]
- Fedorčák, J.; Šmiga, Ľ.; Kutsokon, I.; Kolarčik, V.; Koščová, L.; Oros, M.; Koščo, J. Parasitic infection of Cobitis elongatoides Băcescu & Mayer, 1969 by zoonotic metacercariae Clinostomum complanatum (Rudolphi, 1814). J. Fish Dis. 2019, 42, 1677–1685. [Google Scholar] [CrossRef]
- Lo, C.-F.; Chen, S.-C.; Wang, C.-H. The study of Clinostomum complanatum (Rud., 1814) V. The influences of metacercaria of Clinostomum complanatum on fish. Fish Pathol. 1985, 20, 305–312. [Google Scholar] [CrossRef]
- Shareef, P.; Abidi, S. Incidence and histopathology of encysted progenetic metacercaria of Clinostomum complanatum (Digenea: Clinostomidae) in Channa punctatus and its development in experimental host. Asian Pac. J. Trop. Biomed. 2012, 2, 421–426. [Google Scholar] [CrossRef] [Green Version]
- de Lima, H.L.; Stefani, L.M.; Pedron, F.D.A.; Baldissera, M.D.; da Silva, A.S. Pro-Inflammatory Cytokines in the Serum of Silver Catfish (Rhamdia quelen) Naturally Infected by Clinostomum complanatum: A Preliminary Study. J. Parasitol. 2014, 100, 142–143. [Google Scholar] [CrossRef]
- Montes, M.M.; Plaul, S.E.; Croci, Y.; Waldbillig, M.; Ferrari, W.; Topa, E.; Martorelli, S.R. Pathology associated with three new Clinostomum metacercariae from Argentina with morphological and DNA barcode identification. J. Helminthol 2020, 94, e148. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, J.M.; Sarabia, D.O.; Lopez, R.C.; Lopez, J.F. Helmintos del pez Dormitator maculatus (Osteichthyes: Eleotridae) de Alvarado, Veracruz, México. Rev. De Biol. Trop. 2004, 52, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Kaur, P.; Qureshi, T.; Shrivastav, R.; Manohar, S.; Bilal, A. Histopathological and haematological investigations on Nandus nandus (Ham.) parasitized by metacercariae of Clinostomum complanatum (Rudolphi, 1819). Int. J. Environ. Sci. 2012, 2, 1324–1330. [Google Scholar]
- Kalantan, A.M.N.; Arfin, M.; Nizami, W.A. Histochemical nature and origin of the metacercarial cyst of Clinostomum complanatum (Trematoda: Digenea). Trans. the Am. Microsc. Soc. 1986, 105, 290–295. [Google Scholar] [CrossRef]
- Lo, C.F.; Wang, C.H.; Ogawa, K.; Wakabayashi, H. The mechanism of the metacercarial excystment of Clinostomum complanatum (Rud 1814). Fish Pathol. 1987, 22, 227–235. [Google Scholar] [CrossRef]
- Szalai, A.J.; Dick, T.A. Helminths of stocked rainbow-trout (Salmo gairdneri) with special reference to Clinostomum complanatum. J. Wildl. Dis. 1988, 24, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, S.; Kabayama, T.; Kondo, M.; Takahashi, Y. Mass Mortalities of Goldfish Carassius auratus Infected with Clinostomum Metacercariae, Associated with Elevated Water Temperature. Fish Pathol. 2018, 53, 44–47. [Google Scholar] [CrossRef] [Green Version]
- Caffara, M.; Bruni, G.; Paoletti, C.; Gustinelli, A.; Fioravanti, M.L. Metacercariae of Clinostomum complanatum (Trematoda: Digenea) in European newts Triturus carnifex and Lissotriton vulgaris (Caudata: Salamandridae). J. Helminthol 2014, 88, 278–285. [Google Scholar] [CrossRef]
- Etges, F.J. Clinostomum attenuatum (Digenea) from the Eye of Bufo marinus. J. Parasitol. 1991, 77, 634–635. [Google Scholar] [CrossRef]
- Perpiñán, D.; Garner, M.M.; Trupkiewicz, J.G.; Malarchik, J.; Armstrong, D.L.; Lucio-Forster, A.; Bowman, D.D. Scoliosis in a tiger salamander (Ambystoma tigrinum) associated with encysted digenetic trematodes of the genus Clinostomum. J. Wildl. Dis. 2010, 46, 579–584. [Google Scholar] [CrossRef]
- Bruni, G.; Angelini, C. No short term effect of Clinostomum complanatum (Trematoda: Digenea: Clinostomatidae) on survival of Triturus carnifex (Amphibia: Urodela: Salamandridae). Acta Herpetol 2016, 11, 75–79. [Google Scholar] [CrossRef]
- Sinsch, U.; Lümkemann, K.; Rosar, K.; Schwarz, C.; Dehling, J.M. Acoustic niche partitioning in an anuran community inhabiting an Afromontane wetland (Butare, Rwanda). Afr. Zool. 2012, 47, 60–73. [Google Scholar] [CrossRef]
- Sinsch, U.; Dehling, J.M. Tropical anurans mature early and die young: Evidence from eight Afromontane Hyperolius species and a meta-analysis. PLOS ONE 2017, 12, e0171666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Leon, G.P.P.; Garcia-Varela, M.; Pinacho-Pinacho, C.D.; Sereno-Uribe, A.L.; Poulin, R. Species delimitation in trematodes using DNA sequences: Middle-American Clinostomum as a case study. Parasitol. 2016, 143, 1773–1789. [Google Scholar] [CrossRef] [Green Version]
- Tumushimire, L.; Mindje, M.; Sinsch, U.; Dehling, M.J. Anuran diversity of cultivated wetlands in Rwanda: Melting pot of generalists? Salamandra 2020, 56, 99–112. [Google Scholar]
- Sinsch, U.; Dehling, J.M.; Scheid, P.; Balczun, C. A new African species of parasitic Dero (Annelida, Clitellata, Naididae) in the urinary tract of reed frogs. Parasitol. Res. 2019, 118, 3359–3370. [Google Scholar] [CrossRef] [PubMed]
- Sinsch, U.; Dehling, J.M.; Scheid, P.; Balczun, C. Molecular Diversity of Nematode Parasites in Afrotropical Reed Frogs (Hyperolius spp.). Diversity 2020, 12, 265. [Google Scholar] [CrossRef]
- Bush, A.; Lafferty, K.; Lotz, J.; Shostak, A. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Sinsch, U. Review: Skeletochronological assessment of demographic life-history traits in amphibians. Herpetol. J 2015, 25, 5–13. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinform 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar] [CrossRef]
- Moszczynska, A.; Locke, S.A.; McLaughlin, J.D.; Marcogliese, D.J.; Crease, T.J. Development of primers for the mitochondrial cytochrome c oxidase I gene in digenetic trematodes (Platyhelminthes) illustrates the challenge of barcoding parasitic helminths. Mol. Ecol. Resour. 2009, 9, 75–82. [Google Scholar] [CrossRef]
- Siddiqui, A.; Nizami, W. Seasonal population dynamics of the metacercariae of Clinostomum complanatum (Trematoda: Digenea) in relation to sex of the host. Riv. di Parassitol. 1982, 43, 275–279. [Google Scholar]
- Kalantan, A.; Arfin, M.; Nizami, W.A. Seasonal incidence and pathogenicity of the metacercariae of Clinostomum complanatum in Aphanius dispar. Jpn. J. Parasitol. 1987, 36, 17–23. [Google Scholar]
- Tabaro, S.R.; Mutanga, O.; Rugege, D.; Micha, J.C. Optimum rabbit density over fish ponds to optimise Nile tilapia production in an integrated rabbit–fish system in Rwanda. Afr. J. Aquat. Sci. 2012, 37, 165–174. [Google Scholar] [CrossRef]
- Brown, D.S. Freshwater Snails of Africa and Their Medical Importance; Taylor and Francis Ltd.: London, UK, 1980. [Google Scholar]
- Isabwe, A.; Ruberanziza, E.; Mupfasoni, D.; Ruxin, J.; Clerinx, J.; White, P. Potential for transmission of schistosomiasis in Kayonza district. Rwanda Med J. 2012, 69, 14–19. [Google Scholar]
- Osborn, H.L. On the structure of Clinostomum marginatum, a trematode parasite of the frog, bass and heron. J. Morphol. 1912, 23, 189–229. [Google Scholar] [CrossRef] [Green Version]
- Sinsch, U.; Kaschek, J.; Wiebe, J. Heavy metacercariae infestation (Parastrigea robusta) promotes the decline of a smooth newt population (Lissotriton vulgaris). Salamandra 2018, 54, 210–221. [Google Scholar]
- Acosta, A.A.; Caffara, M.; Fioravanti, M.L.; Utsunomia, R.; Zago, A.C.; Franceschini, L.; da Silva, R.J. Morphological and Molecular Characterization of Clinostomum detruncatum (Trematoda: Clinostomidae) Metacercariae Infecting Synbranchus marmoratus. J. Parasitol. 2016, 102, 151–156. [Google Scholar] [CrossRef]
- Agarwal, S.M. Studies on the morphology, systematics and life history of Clinostomum giganticum n.sp. (Trematoda: Clinostomatidae). Indian J. Helminthol 1960, 11, 75–115. [Google Scholar]
- Ukoli, F.M.A. On Clinostomum tilapiae n. sp., and C. phalacrocoracis Dubois, 1931 from Ghana, and a Discussion of the Systematics of the Genus Clinostomum Leidy, 1856. J. Helminthol 1966, 40, 187–214. [Google Scholar] [CrossRef]
- Ahmad, F.; Sofi, O.; Sofi, T.A.; Fazili, K.M.; Sheikh, B.A. Morphological, molecular and karyological analysis of Clinostomum schizothoraxi kaw, 1950 (Digenea) from fishes of Kashmir Valley, India. Int. J. Pure Appl. Zool. 2017, 5, 33–44. [Google Scholar]
- Tansatit, T.; Sobhon, P.; Sahaphong, S.; Sangsuriya, P.; Klinsrithong, S. Prevalence and Histopathology of Trichogaster pectoralis Harbouring Metacercaria of Clinostomum piscidium (Southwell and Prashad, 1918) in Central Thailand. Thai J. Vet. Med. 2014, 44, 223–230. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence | Reference |
|---|---|---|---|
| ITS1-5.8S-ITS2 | ITS1 | 5′-TCCGTAGGTGAACCTGCGG-3′ | [42] |
| ITS4 | 5′-TCCTCCGCTTATTGATATGC-3′ | [42] | |
| COI | MplatCOX1dF | 5′-TGTAAAACGACGGCCAGTTTWCITTRGATCATAAG-3′ | [43] |
| MplatCOX1dR | 5′-CAGGAAACAGCTATGACTGAAAYAAYAIIGGATCICCACC-3′ | [43] |
| Trematode Species/Host Specimen | Locus | |
|---|---|---|
| ITS1-5.8S-ITS2 | COX1 | |
| Clinostomum chabaudi | ||
| ex HK1 | MW528862 | MW525123 |
| ex HK2 | MW528861 | MW525125 |
| ex HK4 | MW528859 | MW525129 |
| ex HK 5 | MW528855 | MW525124 |
| ex HV17 | MW528854 | MW525122 |
| ex HV 70 | MW528856 | MW525126 |
| ex HV 74 | MW528857 | MW525128 |
| ex HV 1801 | MW528858 | MW525127 |
| ex HV 1818 | MW528860 | MW525121 |
| ex HV 1819 | MW528863 | MW525130 |
| Morphotype 4 ex B. trimaculatus | KY865644 | KY865661 |
| Palearctic Species: | ||
| Morphotype 2 | KY865645 | KY865662 |
| Morphotype 3 | KY865648 | KY865667 |
| Clinostomum complanatum | KM518259 | KM518253 |
| Clinostomum cutaneum | KP110564 | KP110515 |
| Clinostomum phalacrocoracis | KJ786975 | KJ786967 |
| Clinostomum phillipinese | - | KP110523 |
| Clinostomum sinensis | MK796826 | MK801711 |
| Clinostomum tilapiae | KY649349 | KY649357 |
| Clinostomum ukolii | KY865623 | KY865641 |
| Outgroup Species: | ||
| Clinostomum attenuatum | - | JF718587 |
| Euclinostomum heterostomum | KP721439 | KP721421 |
| Host | H. kivuensis | H. viridiflavus | Hyperolius sp. | Barbus trimaculatus |
|---|---|---|---|---|
| Metacercaria N | 15 | 21 | 3 | 1 |
| Source | This study | This study | [11] | [7] |
| Size features: | ||||
| BL [µm] | 2205–7544 (4496 ± 1778) | 2735–7159 (4331 ± 928) | 4778–7380 (6262 ± 1332) | 3666 |
| BW [µm] | 916–2473 (1460 ± 508) | 982–1838 (1373 ± 204) | 1496–1826 (1677 ± 167) | 1176 |
| BL/BW | 2.4–3.9 (3.1 ± 0.4) | 2.7–4.0 (3.1 ± 0.4) | 3.2–4.0 (3.7 ± 0.4) | 3.1 |
| Sucker features: | ||||
| OSL [µm] | 229–696 (343 ± 123) | 202–390 (295 ± 55) | 328–428 (363 ± 57) * | 256 |
| OSW [µm] | 144–441 (235 ± 94) | 131–335 (224 ± 54) | 328–428 (363 ± 57) * | 287 |
| OSW/BW | 0.09–0.23 (0.16 ± 0.03) | 0.11–0.28 (0.16 ± 0.04) | 0.19–0.23 (0.22 ± 0.02) | 0.24 |
| VSL [µm] | 501–1073 (759 ± 196) | 410–986 (740 ± 134) | 785–1000 (880 ± 110) * | 773 |
| VSW [µm] | 479–1040 (738 ± 200) | 378–942 (682 ± 125) | 785–1000 (880 ± 110) * | 811 |
| VSW/BW | 0.42–0.63 (0.52 ± 0.05) | 0.33–0.59 (0.50 ± 0.05) | 0.50–0.55 (0.52 ± 0.02) | 0.68 |
| VSW/OSW | 2.34–4.94 (3.29 ± 0.63) | 2.14–4.98 (3.14 ± 0.64) | 2.34–2.58 (2.44 ± 0.13) | 2.82 |
| DBS [µm] | 105–549 (375 ± 125) | 168–733 (381 ± 118) | n.a. | 719 |
| Genital features: | ||||
| ATL [µm] | 226–755 (530 ± 168) | 341–497 (387 ± 48) | 414–612 (546 ± 114) | 447 |
| ATW [µm] | 208–408 (302 ± 66) | 248–426 (306 ± 56) | 342–456 (394 ± 58) | 489 |
| ATW/ATL | 0.39–0.92 (0.61 ± 0.18) | 0.65–0.97 (0.79 ± 0.10) | 0.63–0.83 (0.73 ± 0.10) | 1.09 |
| PTL [µm] | 209–385 (347 ± 63) | 224–377 (281 ± 51) | 314–528 (437 ± 110) | 305 |
| PTW [µm] | 318–708 (549 ± 127) | 360–534 (461 ± 50) | 485–600 (538 ± 58) | 534 |
| PTW/PTL | 1.25–1.85 (1.58 ± 0.19) | 1.34–2.12 (1.68 ± 0.31) | 1.00–1.54 (1.27 ± 0.27) | 1.75 |
| OL [µm] | 97–199 (136 ± 27) | 77–194 (112 ± 67) | 172–214 (195 ± 21) | n.a. |
| OW [µm] | 106–160 (135 ± 55) | 74–161 (108 ± 55) | 86–160 (121 ± 37) | n.a. |
| OW/OL | 0.69–1.24 (0.98 ± 0.28) | 0.65–1.09 (0.96 ± 0.38) | 0.40–0.80 (0.63 ± 0.20) | n.a. |
| CPL [µm] | 251–619 (480 ± 186) | 232–529 (445 ± 156) | 280–428 (378 ± 85) | 374 |
| CPW [µm] | 88–178 (132 ± 42) | 89–166 (127 ± 48) | 128–300 (200 ± 90) | 193 |
| CPL/BL | 0.083–0.095 (0.089 ± 0.006) | 0.063–0.099 (0.091 ± 0.008) | 0.058–0.065 (0.060 ± 0.004) | 0.10 |
| Host Species N Specimens Locality | H. kivuensis 89 Huye | H. kivuensis 26 Musanze | H. viridiflavus 100 Huye | H. viridiflavus 29 Muhanga |
|---|---|---|---|---|
| Number of metacercariae | Ntot = 63 | Ntot = 18 | Ntot = 85 | Ntot = 4 |
| Prevalence [%] | 19.1 | 15.4 | 20.0 | 10.3 |
| Intensity of infection | 3.7 ± 5.8 | 4.5 ± 3.0 | 6.5 ± 4.4 | 1.3 ± 0.6 |
| Range | 1–22 | 2–8 | 1–15 | 1–2 |
| H. kivuensis | H. viridiflavus | |||
|---|---|---|---|---|
| Infested N = 20 | Not Infested N = 81 | Infested N = 12 | Not Infested N = 61 | |
| Log10 (SVL) | 1.441 ± 0.008 | 1.463 ± 0.004 * | 1.443 ± 0.012 | 1.456 ± 0.005 ns |
| Femur length/SVL | 0.429 ± 0.007 | 0.416 ± 0.003 ns | 0.479 ± 0.013 | 0.467 ± 0.006 ns |
| Femur mass/Femur length | 0.404 ± 0.019 | 0.427 ± 0.010 ns | 0.390 ± 0.022 | 0.376 ± 0.010 ns |
| Log10 (Periosteal bone width) | 1.889 ± 0.029 | 1.968 ± 0.014 * | 1.943 ± 0.029 | 1.950 ± 0.013 ns |
| Log10 (Endosteal bone width) | 0.867 ± 0.058 | 1.083 ± 0.028 *** | 0.802 ± 0.141 | 0.825 ± 0.069 ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinsch, U.; Dehling, J.M.; Scheid, P.; Balczun, C. Alternative Development Strategies of Clinostomum chabaudi (Digenea) Metacercariae in Frog Hosts (Hyperolius spp.). Diversity 2021, 13, 93. https://doi.org/10.3390/d13020093
Sinsch U, Dehling JM, Scheid P, Balczun C. Alternative Development Strategies of Clinostomum chabaudi (Digenea) Metacercariae in Frog Hosts (Hyperolius spp.). Diversity. 2021; 13(2):93. https://doi.org/10.3390/d13020093
Chicago/Turabian StyleSinsch, Ulrich, J. Maximilian Dehling, Patrick Scheid, and Carsten Balczun. 2021. "Alternative Development Strategies of Clinostomum chabaudi (Digenea) Metacercariae in Frog Hosts (Hyperolius spp.)" Diversity 13, no. 2: 93. https://doi.org/10.3390/d13020093
APA StyleSinsch, U., Dehling, J. M., Scheid, P., & Balczun, C. (2021). Alternative Development Strategies of Clinostomum chabaudi (Digenea) Metacercariae in Frog Hosts (Hyperolius spp.). Diversity, 13(2), 93. https://doi.org/10.3390/d13020093