
Rusty Blackbird (Euphagus carolinus) Foraging Habitat and Prey Availability in New England: Implications for Conservation of a Declining Boreal Bird Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Methods
2.3. Analysis
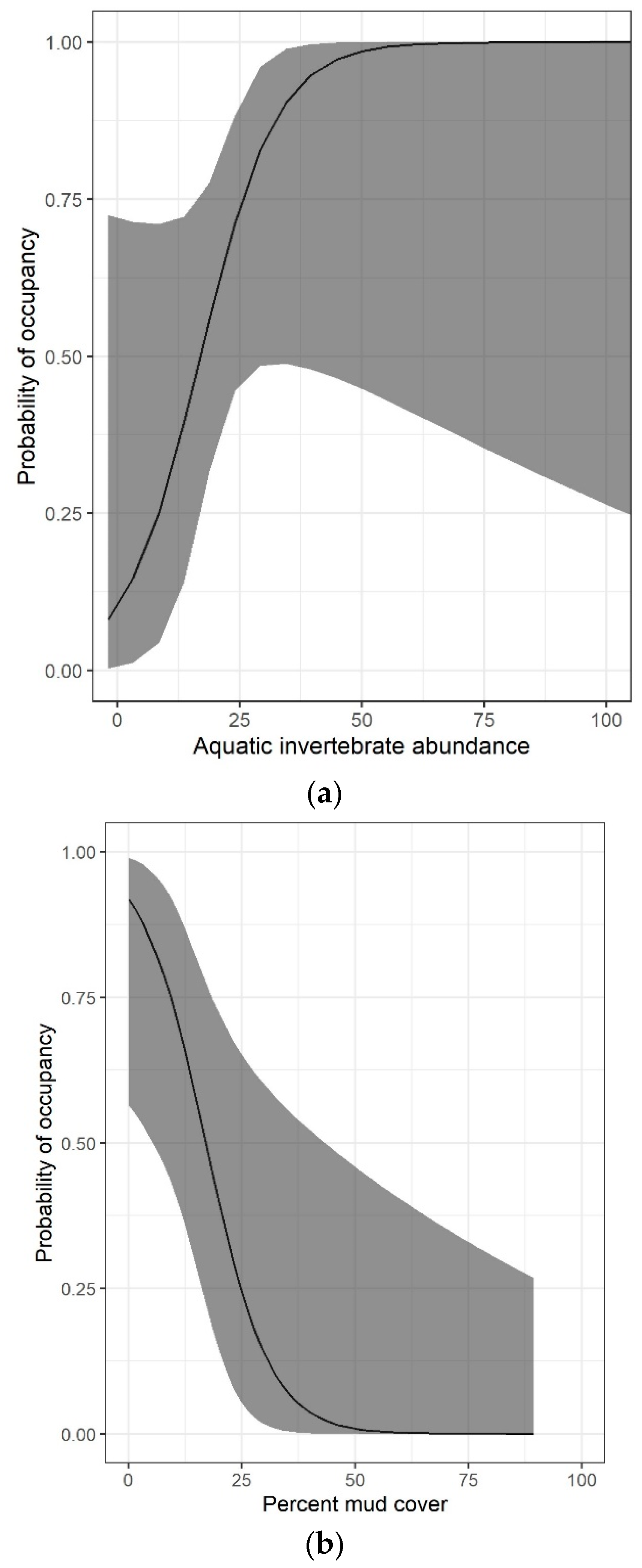
3. Results
4. Discussion
4.1. Wetland Use and Aquatic Macroinvertebrate Prey
4.2. Wetland Characteristics
4.3. Landscape Factors
4.4. Probability of Detection
4.5. Considerations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niven, D.K.; Sauer, J.R.; Butcher, G.S.; Link, W.A. Christmas Bird Count provides insights into population change in land birds hat breed in the boreal forest. Am. Birds 2004, 58, 10–20. [Google Scholar]
- Greenberg, R.; Demarest, D.W.; Matsuoka, S.M.; Mettke-Hofmann, C.; Evers, D.C.; Hamel, P.B.; Luscier, J.D.; Powell, L.L.; Shaw, D.; Avery, M.L.; et al. Understanding Declines in the Rusty Blackbird (Euphagus carolinus). In Studies in Avian Biology, Boreal Birds of North America: A Hemispheric View of Their Conservation Links and Significance; Wells, J.V., Ed.; University of California Press: Berkeley, CA, USA, 2011; pp. 107–126. [Google Scholar]
- Greenberg, R.; Matsuoka, S.M. Rangewide ecology of the declining Rusty Blackbird: Mysteries of a species in decline. Condor 2010, 112, 770–777. [Google Scholar] [CrossRef]
- McClure, C.J.W.; Rolek, B.W.; McDonald, K.; Hill, G.E. Climate change and the decline of a once common bird: Climate change and blackbird decline. Ecol. Evol. 2012, 2, 370–378. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service Migratory Bird Program Focal Species. Available online: https://www.fws.gov/birds/management/managed-species/focal-species.php (accessed on 9 May 2020).
- BirdLife International. The IUCN Red List of Threatened Species, Euphagus carolinus. Available online: https://dx.doi.org/10.2305/IUCN.UK.20182.RLTS.T22724329A131889624.en (accessed on 9 May 2020).
- Edmonds, S.T.; Evers, D.C.; Cristol, D.A.; Mettke-Hofmann, C.; Powell, L.L.; McGann, A.J.; Armiger, J.W.; Lane, O.P.; Tessler, D.F.; Newell, P.; et al. Geographic and seasonal variation in mercury exposure of the declining Rusty Blackbird. Condor 2010, 112, 789–799. [Google Scholar] [CrossRef]
- Barnard, W.H.; Mettke-Hofmann, C.; Matsuoka, S.M. Prevalence of hematozoa infections among breeding and wintering Rusty Blackbirds. Condor 2010, 112, 849–853. [Google Scholar] [CrossRef]
- Powell, L.L.; Hodgman, T.P.; Glanz, W.E. Home ranges of Rusty Blackbirds breeding in wetlands: How much would buffers from timber harvest protect habitat? Condor 2010, 112, 834–840. [Google Scholar] [CrossRef]
- Hamel, P.B.; Steven, D.D.; Leininger, T.; Wilson, R. Historical trends in Rusty Blackbird nonbreeding habitat in forested wetlands. In Proceedings of the Fourth International Partners in Flight Conference: Tundra to Tropics, McAllen, TX, USA, 13–16 February 2009; Rich, T.D., Arizmendi, C., Demarest, D.W., Thompson, C., Eds.; pp. 341–353. [Google Scholar]
- Luepold, S.H.B.; Hodgman, T.P.; McNulty, S.A.; Cohen, J.; Foss, C.R. Habitat selection, nest survival, and nest predators of Rusty Blackbirds in northern New England, USA. Condor 2015, 117, 609–623. [Google Scholar] [CrossRef]
- Rosell, F.; Bozser, O.; Collen, P.; Parker, H. Ecological impact of beavers Castor fiber and Castor canadensis and their ability to modify ecosystems. Mammal Rev. 2005, 35, 248–276. [Google Scholar] [CrossRef]
- Müller-Schwarze, D.; Sun, L. The Beaver: Natural History of a Wetlands Engineer; Cornell University Press: Ithaca, NY, USA, 2003. [Google Scholar]
- Johnston, C.A.; Naiman, R.K. Browse selection by beaver: Effects on riparian forest composition. Can. J. For. Res. 1990, 20, 1036–1043. [Google Scholar] [CrossRef]
- Hood, G.A.; Larson, D.G. Ecological engineering and aquatic connectivity: A new perspective from beaver-modified wetlands. Freshw. Biol. 2015, 60, 198–208. [Google Scholar] [CrossRef]
- Cunningham, J.M.; Calhoun, A.J.K.; Glanz, W.E. Patterns of beaver colonization and wetland change in Acadia National Park. Northeast. Nat. 2006, 13, 583–596. [Google Scholar] [CrossRef]
- Margolis, B.E.; Raesly, R.L.; Shumwaya, D.L. The effects of beaver-created wetlands on the benthic macroinvertebrate assemblages of two Appalachian streams. Wetlands 2001, 21, 554–563. [Google Scholar] [CrossRef]
- McDowell, D.M.; Naiman, R.J. Structure and function of a benthic invertebrate stream community as influenced by beaver (Castor canadensis). Oecologia 1986, 68, 481–489. [Google Scholar] [CrossRef]
- Beecher, W.J. Adaptations for food-getting in the American blackbirds. Auk 1951, 68, 411–440. [Google Scholar] [CrossRef]
- Beal, F.E.L. Food of the Bobolink, Blackbirds, and Grackles; U.S. Department Agriculture Biological Survey Bulletin, 13: Washington, DC, USA, 1900. [Google Scholar]
- Bent, A.C. Life Histories of North American Blackbirds, Orioles, Tanagers, and Their Allies; United States National Museum Bulletin: Washinton, DC, USA, 1958; p. 211. [Google Scholar]
- Martin, A.C.; Zim, H.S.; Nelson, A.L. American Wildlife & Plants: A Guide to Wildlife Food Habits; Dover Publications, Inc.: New York, NY, USA, 1951; ISBN 978-0-486-20793-3. [Google Scholar]
- Meanley, B. Blackbirds and the Southern Rice Crop; U.S. Department of the Interior, Fish and Wildlife Service: Washington, DC, USA, 1971.
- Avery, M.L. Rusty Blackbird (Euphagus carolinus). In The Birds of North America 2000; Academy of Natural Sciences: Philadelphia, PA, USA; American Ornithologists’ Union: Washington, DC, USA, 1995; p. 200. [Google Scholar] [CrossRef]
- Ellison, W.G. The status and habitat of the Rusty Blackbird in Caledonia and Essex Counties; Vermont Fish and Wildlife Department: Woodstock, VT, USA, 1990. [Google Scholar]
- Cade, T.J. Aerial feeding of the Rusty Blackbird on mosquitoes. Wilson Bull. 1953, 65, 52–53. [Google Scholar]
- Ehrlich, P.; Dobkin, D.S.; Wheye, D. Birder’s Handbook; Simon and Schuster: New York, NY, USA, 1988. [Google Scholar]
- Matsuoka, S.M.; Shaw, D.; Johnson, J.A. Estimating the abundance of nesting Rusty Blackbirds in relation to wetland habitats in Alaska. Condor 2010, 112, 825–833. [Google Scholar] [CrossRef]
- Edmonds, S.T.; O’Driscoll, N.J.; Hillier, N.K.; Atwood, J.L.; Evers, D.C. Factors regulating the bioavailability of methylmercury to breeding rusty blackbirds in northeastern wetlands. Environ. Pollut. 2012, 171, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Orians, G.H. Blackbirds of the Americas; University of Washington Press: Seattle, WA, USA, 1985. [Google Scholar]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Langtimm, C.A. Estimating site occupancy when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- U.S. Climate Data. Available online: https://www.usclimatedata.com/climate/colebrook/new-hampshire/united-states/usnh0044 (accessed on 5 October 2020).
- Pachomski, A.L. Foraging Habitat Characteristics, Prey Availability, and Detectability of Rusty Blackbirds: Implications for Land and Wildlife Management in the Northern Forest. Master’s Thesis, State University of New York College of Environmental Science and Forestry, Syracuse, NY, USA, 2017. [Google Scholar]
- Tavernia, B.G.; Lyons, J.E.; Loges, B.W.; Wilson, A.; Collazo, J.A.; Runge, M.C. An evaluation of rapid methods for monitoring vegetation characteristics of wetland bird habitat. Wetl. Ecol. Manag. 2015, 23, 495–505. [Google Scholar] [CrossRef]
- Peckarsky, B.L.; Fraissinet, P.R.; Penton, M.; Conklin, D.D., Jr. Freshwater Macroinvertebrates of Northeastern North America; Cornell University Press: Ithaca, NY, USA, 1990. [Google Scholar]
- Homer, C.G.; Dewitz, J.A.; Yang, L.; Jin, S.; Danielson, P.; Xian, G.; Coulston, J.; Herold, N.D.; Wickham, J.D.; Megown, K. Completion of the 2011 National Land Cover Database for the conterminous United States representing a decade of land cover change information. Photogramm. Eng. Remote Sens. 2015, 81, 345–354. [Google Scholar]
- Wohner, P.J.; Foss, C.R.; Cooper, R.J. Rusty blackbird habitat selection and survivorship during nesting and post-fledging. Diversity 2020, 12, 221. [Google Scholar] [CrossRef]
- Behnke, L.A.H.; Pejchar, L.; Crampton, L.H. Occupancy and habitat use of the endangered Akikiki and Akekee on Kauai Island, Hawaii. Condor 2016, 118, 148–158. [Google Scholar] [CrossRef]
- Fiske, I.J.; Chandler, R.B. Unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 2011, 43, 1–23. Available online: http://www.jstatsoft.org/v43/i10/ (accessed on 14 August 2016). [CrossRef] [Green Version]
- R Development Core Team. R version 3.2.5: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: http://www.R-project.org (accessed on 14 August 2016).
- Powell, L.L.; Hodgman, T.P.; Fiske, I.J.; Glanz, W.E. Habitat occupancy of Rusty Blackbirds (Euphagus carolinus) breeding in northern New England, USA. Condor 2014, 116, 122–133. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package Version 2.0-4. 2016. Available online: http://CRAN.R-project.org/package=AICcmodavg (accessed on 14 August 2016).
- MacKenzie, D.I.; Bailey, L.L. Assessing the fit of site-occupancy models. J. Agric. Biol. Environ. Stat. 2004, 9, 300–318. [Google Scholar] [CrossRef]
- Lebreton, J.D.; Burnham, K.P.; Clobert, J.; Anderson, D.R. Modeling survival and testing biological hypotheses using marked animals: A unified approach with case studies. Ecol. Monogr. 1992, 62, 67–118. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer-Verlag: New York, NY, USA, 2002. [Google Scholar]
- Foss, C.; Wohner, P.J. Rusty Blackbird Nest Locations in Relation to Wetlands. Unpublished.
- Buckley, S.H. Rusty Blackbirds in Northeastern U.S. Industrial Forests: A Multi-Scale Study of Nest Habitat Selection and Nest Survival. Master’s Thesis, State University of New York College of Environmental Science and Forestry, Syracuse, NY, USA, 2013. [Google Scholar]
- Foss, C.; (New Hampshire Audubon, Concord, NH, USA). Personal communication, 2017.
- Sinervo, B. Chapter 6: Optimal Foraging Theory: Constraints and Cognitive Processes. In Behavioral Ecology; University of California: Santa Cruz, CA, USA, 1997. [Google Scholar]
- Loomis, D. Reproductive Success and Foraging Ecology of the Rusty Blackbird on the Copper River Delta, Alaska. Master’s Thesis, Oregon State University, Corvallis, OR, USA, 2013. [Google Scholar]
- Meanley, B. Aging and Sexing Blackbirds, Bobolinks, and Starlings; U.S. Fish Wildlife Service, Patuxent Wildlife Research Center: Laurel, MD, USA, 1967.
- Borchert, S.M. Site-Specific Habitat and Landscape Associations of Rusty Blackbirds Wintering in Louisiana. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2015. [Google Scholar]
- DeLeon, E.E. Ecology of Rusty Blackbirds Wintering in Louisiana: Seasonal Trends, Flock Composition, and Habitat Associations. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2012. [Google Scholar]
- Wright, J.R.; Powell, L.L.; Matthews, S.N.; Tonra, C.M. Rusty blackbirds select areas of greater habitat complexity during stopover. Condor 2020, 122, 1–18. [Google Scholar] [CrossRef]
- Scarl, J.C. Rusty Blackbirds 2012: Building Connections for a Declining Species; Vermont Center for Ecostudies: Norwich, VT, USA, 2013. [Google Scholar]
- Naiman, R.J.; Johnston, C.A.; Kelley, J.C. Alteration of North American streams by beaver. BioScience 1988, 38, 753–762. [Google Scholar] [CrossRef]
- Burcher, C.; Smock, L. Habitat distribution, dietary composition and life history characteristics of odonate nymphs in a blackwater coastal plain stream. Am. Midl. Nat. 2002, 148, 75–89. [Google Scholar] [CrossRef]
- Hood, G.A.; Larson, D.G. Beaver-created habitat heterogeneity influences aquatic invertebrate assemblages in boreal Canada. Wetlands 2014, 34, 19–29. [Google Scholar] [CrossRef]
- Bush, B.M.; Stenert, C.; Maltchik, L.; Batzer, D.P. Beaver-created successional gradients increase β-diversity of invertebrates by turnover in stream-wetland complexes. Freshw. Biol. 2019, 64, 1265–1274. [Google Scholar] [CrossRef]
- Glennon, M. Analysis of Boreal Bird Trends in the Adirondack Park, 2007–2016: Report to the New York State Department of Environmental Conservation; Wildlife Conservation Society: Saranac Lake, NY, USA, 2017. [Google Scholar]
- Corcoran, R.M.; Lovvorn, J.R.; Heglund, P.J. Long-term change in limnology and invertebrates in Alaskan boreal wetlands. Hydrobiologia 2009, 620, 77–89. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Hood, G.A.; Bayley, S.E. Beaver (Castor canadensis) mitigate the effects of climate on the area of open water in boreal wetlands in western Canada. Biol. Conserv. 2008, 141, 556–557. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Covariate | Description | Method |
|---|---|---|
| Beaver | Binary measure of observed beaver activity | Field |
| Day | Ordinal survey date | Field |
| Elevation | Site elevation in meters | GIS |
| Water.depth | Average depth (centimeters) of open water near pond edge | Field |
| Invert.abundance | Average number of invertebrate individuals observed in three samples | Field |
| Invert.richness | Total number of invertebrate families observed in three samples | Field |
| Min | Survey start time converted to minute of day | Field |
| Mud | Visual estimate of percent exposed mud within a wetland | Field |
| Open.water | Visual estimate of percent open water within a wetland | Field |
| Pct.conifer | Percent conifer cover within a 500-m buffer of wetland using NLCD 2011 | GIS |
| Puddles | Binary measure of puddles observed 0, 1, 2, or 3 times out of three surveys | Field |
| Precip | Binary measure of presence of precipitation during survey | Field |
| Size | Wetland size measured in meters squared | GIS |
| Temp | Measure of temperature (Celsius) at the start of the survey | Field |
| Visit | Survey period 1, 2, or 3 | Field |
| Wind | Wind speed (km per hour) measured using an anemometer held at 4.5 m height at the start of the survey | Field |
| Yng.conifer | Binary measure of presence of dense regenerating spruce and/or fir trees < 1.5 m tall | Field |
| Model | K a | AIC b | ΔAIC c | wi d | −2 Log-Likelihood |
|---|---|---|---|---|---|
| p(visit) Ψ(.) | 4 | 214.38 | 0.00 | 0.513 | 206.4 |
| p(size) Ψ(.) | 3 | 218.18 | 3.80 | 0.077 | 212.2 |
| p(precip) Ψ(.) | 3 | 219.19 | 4.81 | 0.046 | 213.2 |
| p(temp) Ψ(.) | 3 | 219.26 | 4.88 | 0.045 | 213.2 |
| p(time) Ψ(.) | 3 | 219.34 | 4.96 | 0.043 | 213.3 |
| p(day) Ψ(.) | 3 | 219.39 | 5.01 | 0.042 | 213.4 |
| p(wind) Ψ(.) | 3 | 219.50 | 5.12 | 0.040 | 213.5 |
| p(day+precip) Ψ(.) | 4 | 220.87 | 6.49 | 0.020 | 212.9 |
| p(time+precip) Ψ(.) | 4 | 220.96 | 6.58 | 0.019 | 213.0 |
| p(day+time) Ψ(.) | 4 | 220.99 | 6.61 | 0.019 | 213.0 |
| p(temp+wind) Ψ(.) | 4 | 221.05 | 6.67 | 0.018 | 213.1 |
| p(time+temp) Ψ(.) | 4 | 221.07 | 6.69 | 0.018 | 213.1 |
| p(day+temp) Ψ(.) | 4 | 221.08 | 6.70 | 0.018 | 213.1 |
| p(time+wind) Ψ(.) | 4 | 221.19 | 6.81 | 0.017 | 213.2 |
| p(day+wind) Ψ(.) | 4 | 221.25 | 6.87 | 0.016 | 213.3 |
| p(time2) Ψ(.) | 4 | 221.32 | 6.94 | 0.016 | 213.3 |
| p(day2) Ψ(.) | 4 | 221.36 | 6.98 | 0.015 | 213.4 |
| p(temp*wind) Ψ(.) | 5 | 222.43 | 8.05 | 0.009 | 212.4 |
| p(time+precip+wind) Ψ(.) | 5 | 222.90 | 8.52 | 0.007 | 212.9 |
| Model | K a | QAIC b | ΔQAIC c | Qwi d | −2 Log-Likelihood |
|---|---|---|---|---|---|
| p(visit) Ψ(invert.abundance+mud) | 7 | 123.44 | 0.00 | 0.600 | 188.2 |
| p(visit) Ψ(mud) | 6 | 128.06 | 4.63 | 0.060 | 199.6 |
| p(visit) Ψ(invert.abundance) | 6 | 128.31 | 4.87 | 0.053 | 200.0 |
| p(visit) Ψ(invert.abundance+yng.conifer) | 7 | 129.87 | 6.43 | 0.024 | 199.3 |
| p(visit) Ψ(.) | 5 | 129.99 | 6.55 | 0.023 | 206.4 |
| p(visit) Ψ(mud+yng.conifer) | 7 | 130.05 | 6.61 | 0.022 | 199.6 |
| p(visit) Ψ(beaver+invert.abundance) | 7 | 130.12 | 6.69 | 0.021 | 199.7 |
| p(.) Ψ(.) | 3 | 130.21 | 6.77 | 0.020 | 213.6 |
| p(visit) Ψ(invert.abundance+pct.conifer) | 7 | 130.35 | 6.92 | 0.019 | 200.1 |
| p(visit) Ψ(invert.richness) | 6 | 130.73 | 7.29 | 0.016 | 204.2 |
| p(visit) Ψ(open.water) | 6 | 130.94 | 7.51 | 0.014 | 204.6 |
| p(visit) Ψ(pct.conifer) | 6 | 131.07 | 7.63 | 0.013 | 204.8 |
| p(visit) Ψ(yng.sof+beaver+invert.abundance) | 8 | 131.58 | 8.14 | 0.010 | 198.8 |
| p(visit) Ψ(yng.conifer) | 6 | 131.91 | 8.47 | 0.009 | 206.2 |
| p(visit) Ψ(puddles3x) | 6 | 131.92 | 8.49 | 0.009 | 206.3 |
| p(visit) Ψ(elevation) | 6 | 131.93 | 8.49 | 0.009 | 206.3 |
| p(visit) Ψ(size) | 6 | 131.97 | 8.53 | 0.008 | 206.3 |
| p(visit) Ψ(water.depth) | 6 | 131.97 | 8.53 | 0.008 | 206.4 |
| p(visit) Ψ(beaver) | 6 | 131.99 | 8.55 | 0.008 | 206.4 |
| p(visit) Ψ(invert.richness+yng.conifer) | 7 | 132.68 | 9.24 | 0.006 | 204.1 |
| p(visit) Ψ(beaver+invert.richness) | 7 | 132.73 | 9.29 | 0.006 | 204.2 |
| p(visit) Ψ(open.water+pct.conifer) | 7 | 132.91 | 9.48 | 0.005 | 204.5 |
| p(visit) Ψ(pct.conifer+puddles.3x) | 7 | 132.97 | 9.53 | 0.005 | 204.6 |
| p(visit) Ψ(pct.conifer+beaver) | 7 | 133.05 | 9.61 | 0.005 | 204.8 |
| p(visit) Ψ(yng.conifer) | 7 | 133.84 | 10.41 | 0.003 | 206.1 |
| p(visit) Ψ(size+yng.conifer) | 7 | 133.86 | 10.43 | 0.003 | 206.1 |
| p(visit) Ψ(water.depth+yng.conifer) | 7 | 133.89 | 10.46 | 0.003 | 206.2 |
| p(visit) Ψ(yng.conifer+beaver) | 7 | 133.90 | 10.46 | 0.003 | 206.2 |
| p(visit) Ψ(water.depth+size) | 7 | 133.95 | 10.52 | 0.003 | 206.3 |
| p(visit) Ψ(beaver+size) | 7 | 133.96 | 10.53 | 0.003 | 206.3 |
| p(visit) Ψ(pct.conifer+beaver+puddles.3x) | 8 | 134.97 | 11.53 | 0.002 | 204.6 |
| RUBL Used a | RUBL Undetected b | Poisson Estimates | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Order | Max Count | N | Mean | Max Count | N | Mean | Estimate for Difference c | 95% Lower Bound | Z | p-Value d |
| Amphipoda | 112 | 35 | 3.20 | 28 | 25 | 1.12 | 2.08 | 1.47 | 5.64 | <0.001 |
| Aranae e | 4 | 35 | 0.11 | 3 | 25 | 0.12 | −0.01 | −0.15 | −0.06 | 0.525 |
| Coleoptera | 49 | 35 | 1.40 | 16 | 25 | 0.64 | 0.76 | 0.34 | 2.97 | 0.002 |
| Collembola e | 143 | 35 | 4.09 | 0 | 25 | 0.00 | 4.09 | 3.52 | 11.96 | <0.001 |
| Diptera | 789 | 35 | 22.54 | 284 | 25 | 11.36 | 11.18 | 9.46 | 10.67 | <0.001 |
| Ephemeroptera | 74 | 35 | 2.11 | 165 | 25 | 6.60 | −4.49 | −5.42 | −7.88 | 1.000 |
| Hemiptera e | 8 | 35 | 0.23 | 1 | 25 | 0.04 | 0.19 | 0.04 | 2.09 | 0.018 |
| Hirudinea e, f | 1 | 35 | 0.03 | 6 | 25 | 0.24 | −0.21 | −0.38 | −2.07 | 0.981 |
| Lepidoptera e | 2 | 35 | 0.06 | 0 | 25 | 0.00 | 0.06 | −0.01 | 1.41 | 0.079 |
| Megaloptera e | 5 | 35 | 0.14 | 14 | 25 | 0.56 | −0.41 | −0.68 | −2.56 | 0.995 |
| Odonata | 110 | 35 | 3.14 | 38 | 25 | 1.52 | 1.62 | 0.98 | 4.18 | <0.001 |
| Oligochaeta e, f | 5 | 35 | 0.14 | 0 | 25 | 0.00 | 0.14 | 0.04 | 2.24 | 0.013 |
| Plecoptera e | 7 | 35 | 0.20 | 0 | 25 | 0.00 | 0.20 | 0.08 | 2.65 | 0.004 |
| Trichoptera | 74 | 35 | 2.11 | 37 | 25 | 1.48 | 0.63 | 0.07 | 1.83 | 0.033 |
| Veneroida e | 8 | 35 | 0.23 | 29 | 25 | 1.16 | −0.93 | −1.31 | −4.05 | 1.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pachomski, A.; McNulty, S.; Foss, C.; Cohen, J.; Farrell, S. Rusty Blackbird (Euphagus carolinus) Foraging Habitat and Prey Availability in New England: Implications for Conservation of a Declining Boreal Bird Species. Diversity 2021, 13, 99. https://doi.org/10.3390/d13020099
Pachomski A, McNulty S, Foss C, Cohen J, Farrell S. Rusty Blackbird (Euphagus carolinus) Foraging Habitat and Prey Availability in New England: Implications for Conservation of a Declining Boreal Bird Species. Diversity. 2021; 13(2):99. https://doi.org/10.3390/d13020099
Chicago/Turabian StylePachomski, Amanda, Stacy McNulty, Carol Foss, Jonathan Cohen, and Shannon Farrell. 2021. "Rusty Blackbird (Euphagus carolinus) Foraging Habitat and Prey Availability in New England: Implications for Conservation of a Declining Boreal Bird Species" Diversity 13, no. 2: 99. https://doi.org/10.3390/d13020099
APA StylePachomski, A., McNulty, S., Foss, C., Cohen, J., & Farrell, S. (2021). Rusty Blackbird (Euphagus carolinus) Foraging Habitat and Prey Availability in New England: Implications for Conservation of a Declining Boreal Bird Species. Diversity, 13(2), 99. https://doi.org/10.3390/d13020099