1. Introduction
Masdevallia Ruiz. & Pav. in the broad concept comprises about 350 species distributed from southern Mexico to southern Brazil [
1], reaching the highest diversity in the Andes of Ecuador and Colombia, Peru, and Bolivia. It is probably one of the most distinguishable genera of the subtribe Pleurothallidinae. The plants are caespitose and have fleshy, smooth leaves. The often brightly colored, large, and showy flowers commonly have long-tailed sepals and the obscure, small petals.
Genus
Masdevallia has been firstly reported for Guatemala by Ames and Correll [
2], who indicated the presence of seven species. After this publication, two of these species were transferred to the genus
Dracula Luer and two others to the genus
Dryadella Luer, leaving only three species in
Masdevallia:
M. floribunda Lindl.,
M. chontalensis Rchb.f., and
M. tubuliflora Ames. For an unknown reason, Ames did not include
M. tuerckheimii Ames, described by him in 1908. Later, two more species were discovered in Guatemala:
M. adamsii Archila [
3] and
M. corinnea Archila, Jiménez Rodr. & Véliz [
4].
During our studies on orchid diversity carried out in the Estación Experimental de Orquídeas de la Familia Archila in Cobán (Guatemala), we came across plants that in our opinion are the result of the hybridization between two native species of the genus
Masdevallia:
M. floribunda and
M. tuerckheimii. Both species were found on the trunks of numerous trees located across the area of the Station (e.g.,
Coffea arabica L.,
Citrus sinensis (L.) Osbeck, and
Eugenia paniculata Jacq.) and propagated naturally. The flowering period of
M. floribunda and
M. tuerckheimii ranges from June to February, coinciding with the rainy season. What is more, we were able to document the pollination process in both species done by two still undescribed species of fruit flies belonging to the genus
Zygothrica [
5].
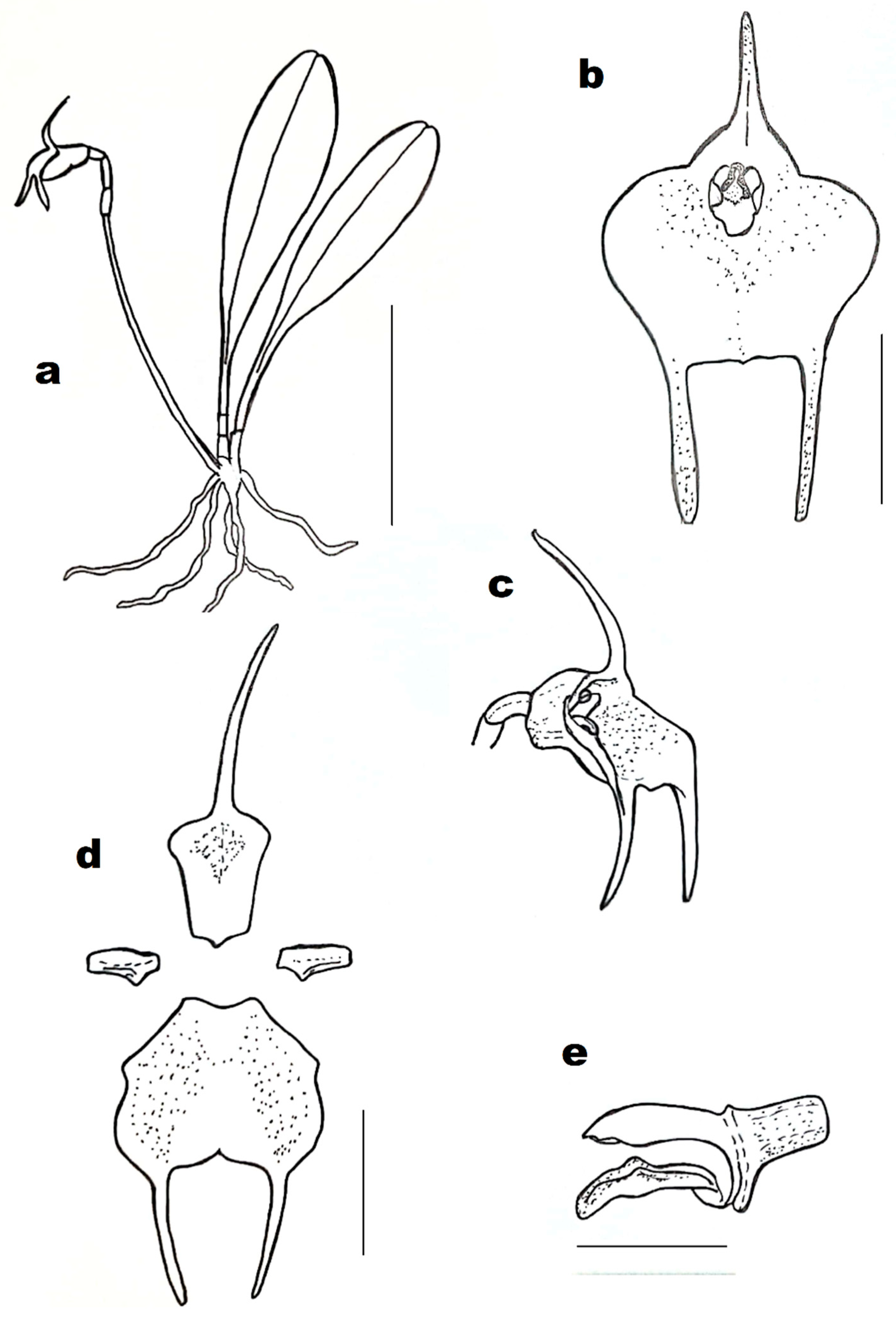
Masdevallia floribunda (
Figure 1b and
Figure 2) is distributed from Mexico to Honduras. It can be found usually on the trees in moist to wet forests, at the elevation of about 900 to 1200 m a.s.l. [
6]. Plants are epiphytic, erect-spreading, and clustered. The ramicaul is covered with short papery sheaths (when young). Leaves are somewhat glossy, broadest above the middle, leathery, with glossy petiole, rounded at the apex, and grooved along the midrib. Inflorescences are single-flowered, arising from the base of the leaf, spreading. Sepals are whitish, finely dotted, and slightly tinged with crimson. Dorsal sepal abruptly tapers into cauda. It is erect to reflexed, joined to lateral sepals. Lateral sepals are fused, yellow at the base, with brownish-yellow to greenish, pointing down to reflexed caudae. Petals are linear-oblong, white, and toothed at the truncated tip. The lip is hinged to the column-foot, reflexed at the tip, white with crimson dots and a yellow tip. The column is greenish with purple edges. The anther produces two obovoid pollinia.
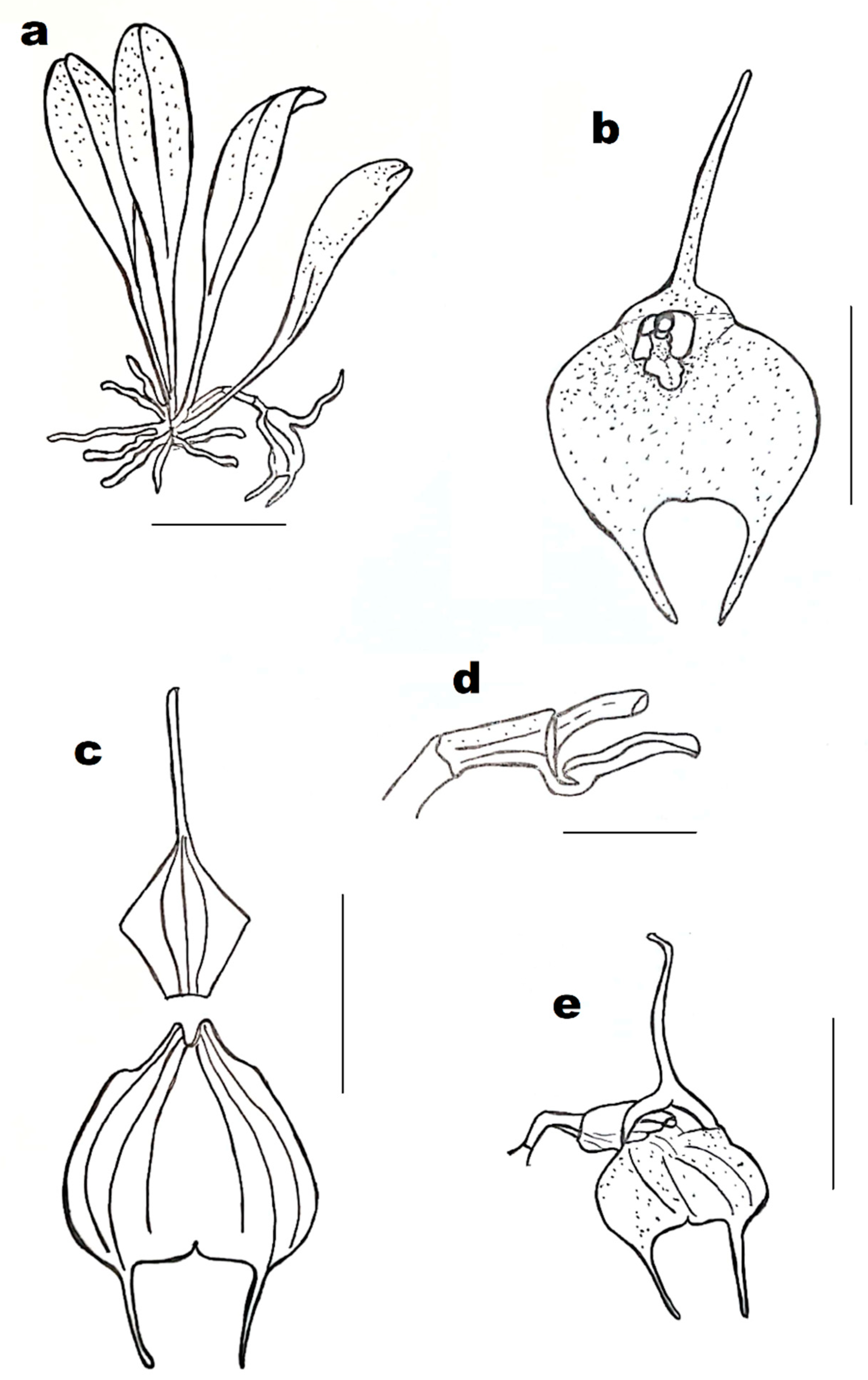
Masdevallia tuerckheimii (
Figure 1d and
Figure 3) has been reported in Mexico (Chiapas) and Guatemala, in the wet lower mountain rain forest, at the altitude of about 350–800 m a.s.l. This species is locally abundant along creeks and in gallery forest, on shaded tree trunks. Plants of
M. tuerckheimii are epiphytic, caespitose herbs. Their ramicauls are terete, sulcate, very short, and covered by two membranaceous sheaths. Leaves’ blades are convex, arcuate, oblanceolate-spatulate with the apex obtuse-subacute and the base attenuate into a sulcate petiole. Inflorescences are short, pendulous, and facultatively bifloral—when the first flower falls off, the second one develops, but in the case when the first one has been pollinated, the plant does not produce the second flower. Sepals form a basal tube. The dorsal sepal is usually yellowish, slightly tinged with purple at the opening of the tube. Lateral sepals are usually pinkish or creamy-white with many clustered dots and/or short, transverse, purple stripes and yellow caudae. Petals are white, same as the lip, which additionally is spotted with purple. The dorsal sepal is partially united to the laterals, rhombic, abruptly caudate. Lateral sepals are connate, forming an obovate to transversely elliptic-oblong blade, abruptly bicaudate. Petals are oblong, truncate, erose-dentate at the apex, with a marginal keel in the lower margin. The lip is shortly clawed, oblong, with two plates separated by an axial groove, truncate-obtuse, deflexed, thickened, and erose-dentate at the apex. The column is slender, subclavate, slightly arcuate, cream-white with a solid purple line on the ventral margins and the clinandrium. Pollinia are two, ovoid-subclavate, compressed, with a small viscidium.
In the past,
M. tuerckheimii has been considered as subspecies [
1] or variety [
7] of
M. floribunda. However,
M. tuerckheimii can be easily distinguished by the larger, darker, and more showily colored flowers (yellow caudae and densely purple-spotted sepals), less floriferous habit, and short, pendulous inflorescences. The appearance of the flowers of
M. tuerckheimii is very different from that of
M. floribunda. It is clear that both species are ecologically parapatric;
M. floribunda is a species of cloud forest, while
M. tuerckheimii grows in tropical rain forests. However, there are ecotone zones between these two ecosystems where both species and some intermediate individuals can be found growing together and hereby, we describe them as a new natural hybrid.
3. Results
3.1. Taxonomic Treatment
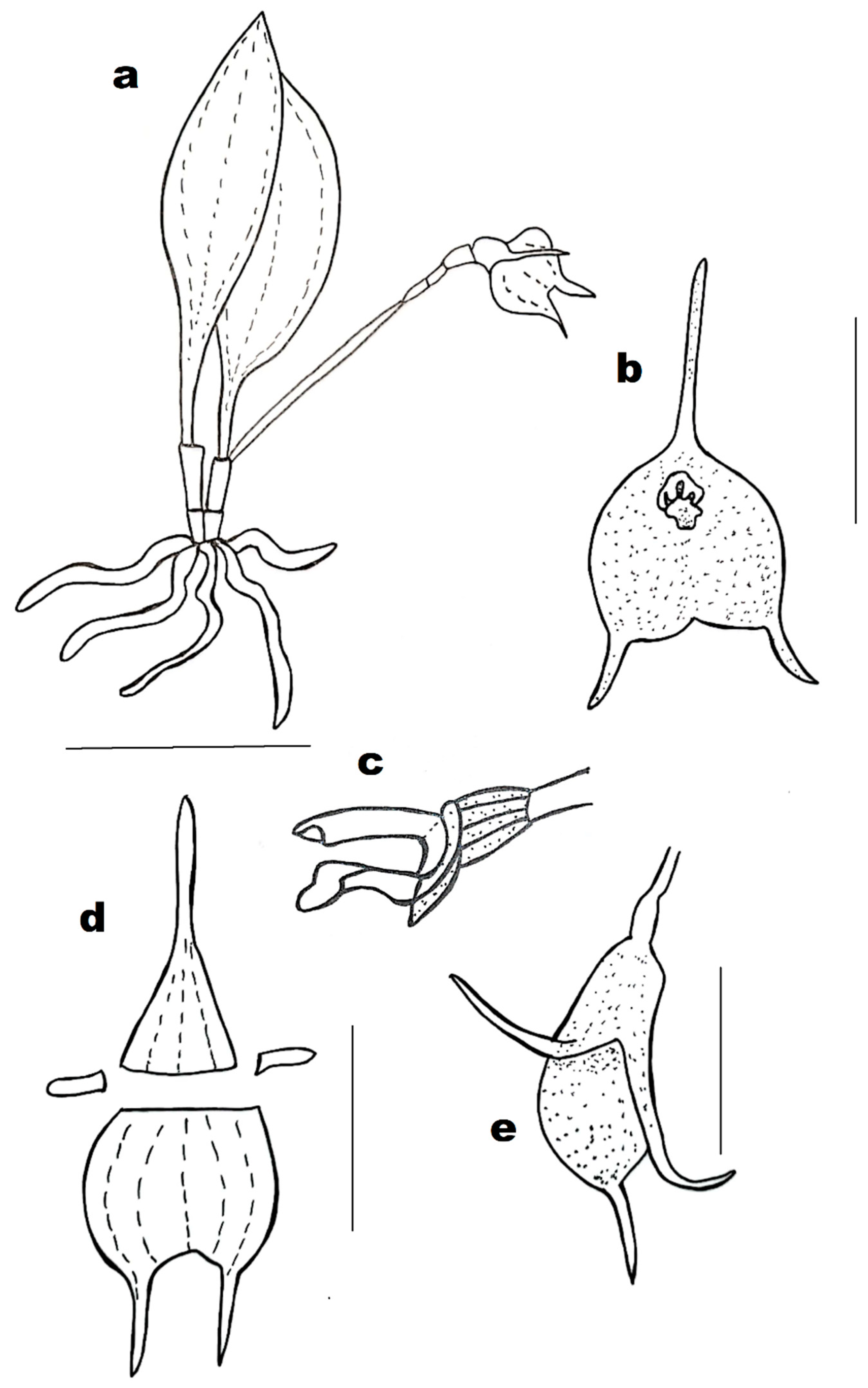
Masdevallia × urbanae Archila, Lipińska & Szlach., hybr. nov. (
Figure 1c and
Figure 4).
TYPE: FA s.n.; deposited in herbario BIGU
TYPE LOCALITY: Guatemala, Alta Verapaz, Municipio de Cobán, ciudad de Cobán, en jardines de la Estación Experimental de Orquídeas de la Familia Archila, 1300 m asl, Noviembre 2017.
HABITAT AND DISTRIBUTION: Cloud forest, at type locality
ETYMOLOGY: The name refers to the place where the first plants of the new nothotaxon have been found—the urban area of the city of Cobán.
DIAGNOSIS: Masdevallia × urbanae can be easily distinguished from parental species by the shape of synsepal with shorter and thicker sepaline tails directed outward.
3.2. Description
Plants small, caespitose. Leaves petiolate with basal bract sheaths 6 to 8.5 cm long, and 1.6 cm wide, minutely tridentate at the apex. Inflorescence single-flowered (although this is relative in this group and depends on nutritional and humidity conditions, if the first flower is not fertilized, a second flower is produced). Axil 2.2 cm long, petiole 0.8 cm long, ovary 0.4 cm long and 0.15 cm wide. Flowers variable in color but usually white with purple spots or purple with spots. Sepals united in a tube, separated apically. Tube 0.5 cm long and 0.45 cm wide. Dorsal sepal in its free part triangular, 0.3 cm long and 0.35 cm wide, apically aciculate-attenuated, 0.9 cm long. Lateral sepals united, forming a broadly ovate synsepal, 1.5 cm long and 1.4 cm wide. Petals oblong with the internal part ornamented and the apiculate-truncated apex 0.25 cm long and 0.1 cm wide. Lip oblong with subcarinate surface, 0.3 cm long and 0.1 cm wide.
4. Discussion
Both M. floribunda and M. tuerckheimii are thought to be extremely variable species in terms of coloration. As it is stated in type protologue of M. floribunda, it probably has been collected in Mexico (possibly by H. Galeotti in Veracruz in 1840) and later cultivated in Sevenoaks, England by J. Rogers. With time, other specimens with different color patterns or regions of the collection were described by the different scientists. These were for example Masdevallia galeottiana A. Rich. & Galeotti with purple flowers and M. lindeniana A. Rich. & Galeotti with pale yellow flowers, both found in Mexico and both with time reduced to synonymy under M. floribunda. M. tuerckheimii, described from Guatemala, is sometimes wrongly considered by some taxonomists to be just a synonym or color variation of M. floribunda. It tends to be more brightly colored, white-spotted purple or cream speckled red, but still quite variable. The ecological niches of both species are rather separated, as in general, M. floribunda is considered to be a species of the cloud forest and M. tuerckheimii grows in tropical rain forests.
Ecological evidence, such as the coexistence of both orchid species in a relatively small area, the same habitat and flowering time, as well as common pollinators, strongly support our theory that hybridization was the source of plants with intermediate features. The Estación Experimental de Orquídeas de la Familia Archila is located in a cloud forest, at the altitude of 1320 m asl, from where the plants of M. tuerckheimii are taken to the stacion’s facilities. This apparently is a reason for a greater intensity of pollination of both orchid species and the hybridization process stimulated effectively by the Zygothrica fruit flies.
During our study, a population of ca. 50 individuals of Masdevallia × urbanae have been carefully examined. Field observations suggested that seeds produced by most of them are vivid and during the past five years M. × urbanae successfully populated trees in the area of Station, forming dense, regularly expanding clumps. We were not able to observe whether cross-pollination between the hybrid and parental species occurs, thus we leave it as a possible hypothesis until further investigation, also experimental, will be performed.









{kind=link}
{kind=link}
{kind=link}
{kind=link}