
Histovariability and Palaeobiological Implications of the Bone Histology of the Dromornithid, Genyornis newtoni


Abstract
:1. Introduction
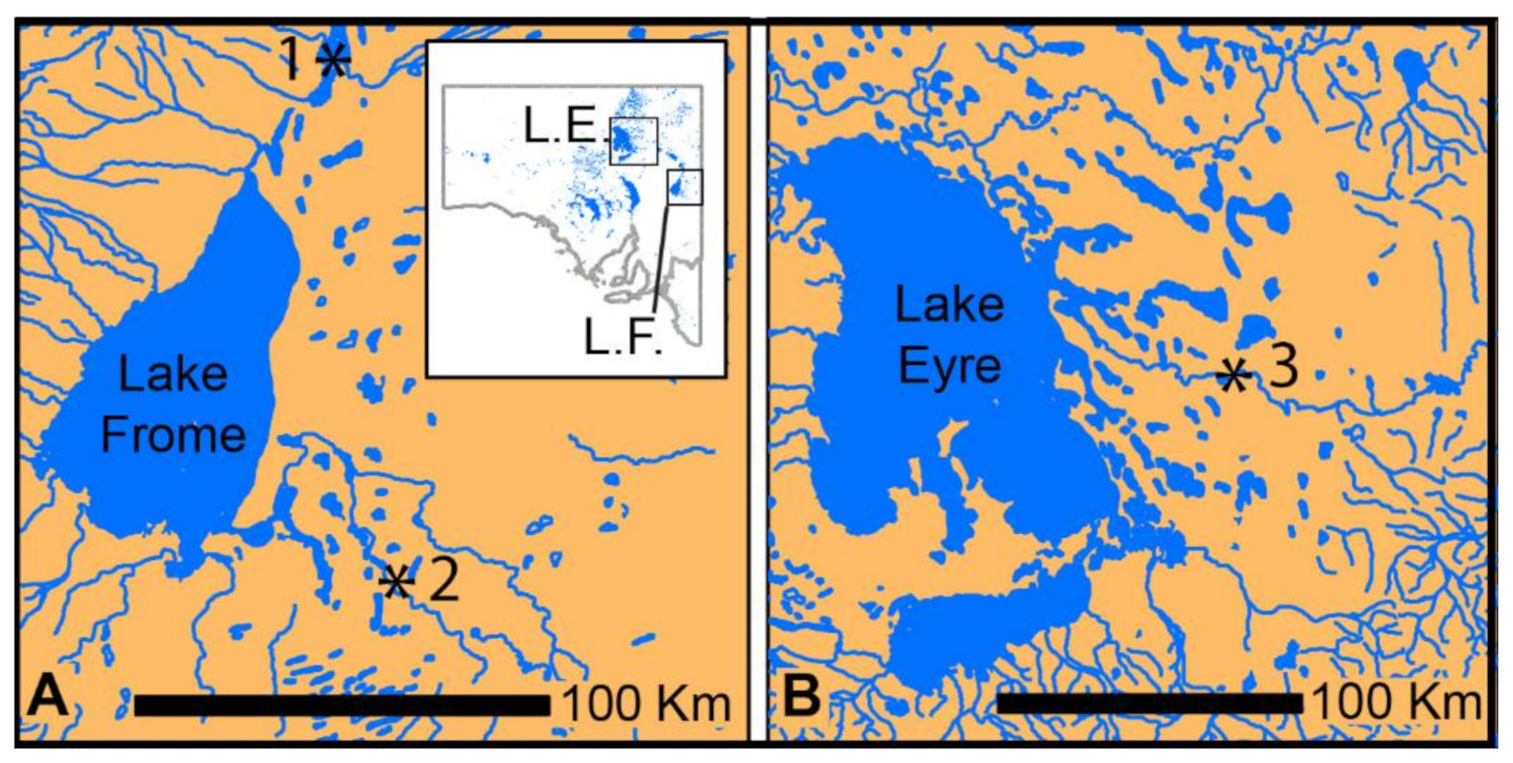
2. Materials
3. Results
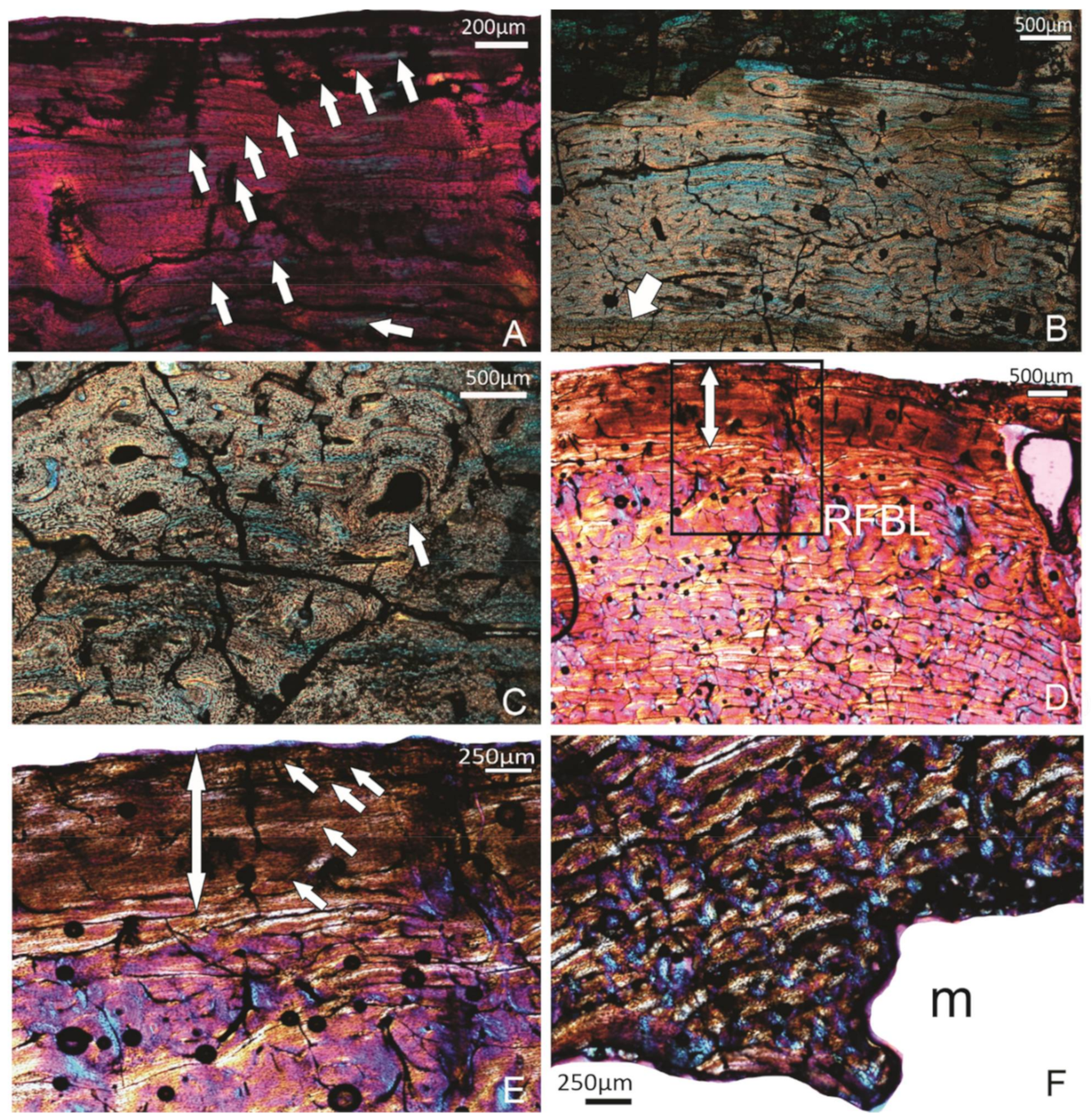
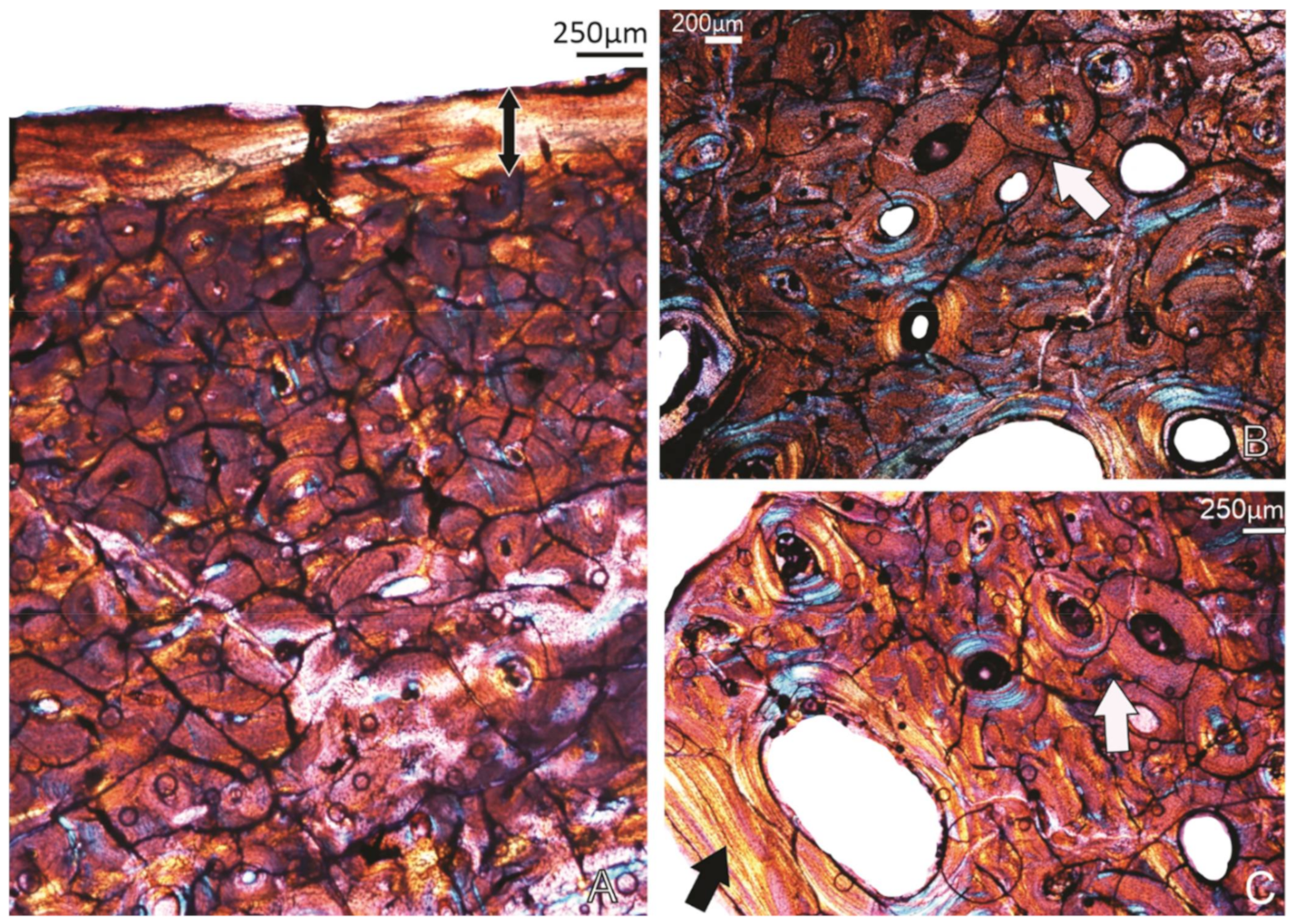
3.1. Tibiotarsi
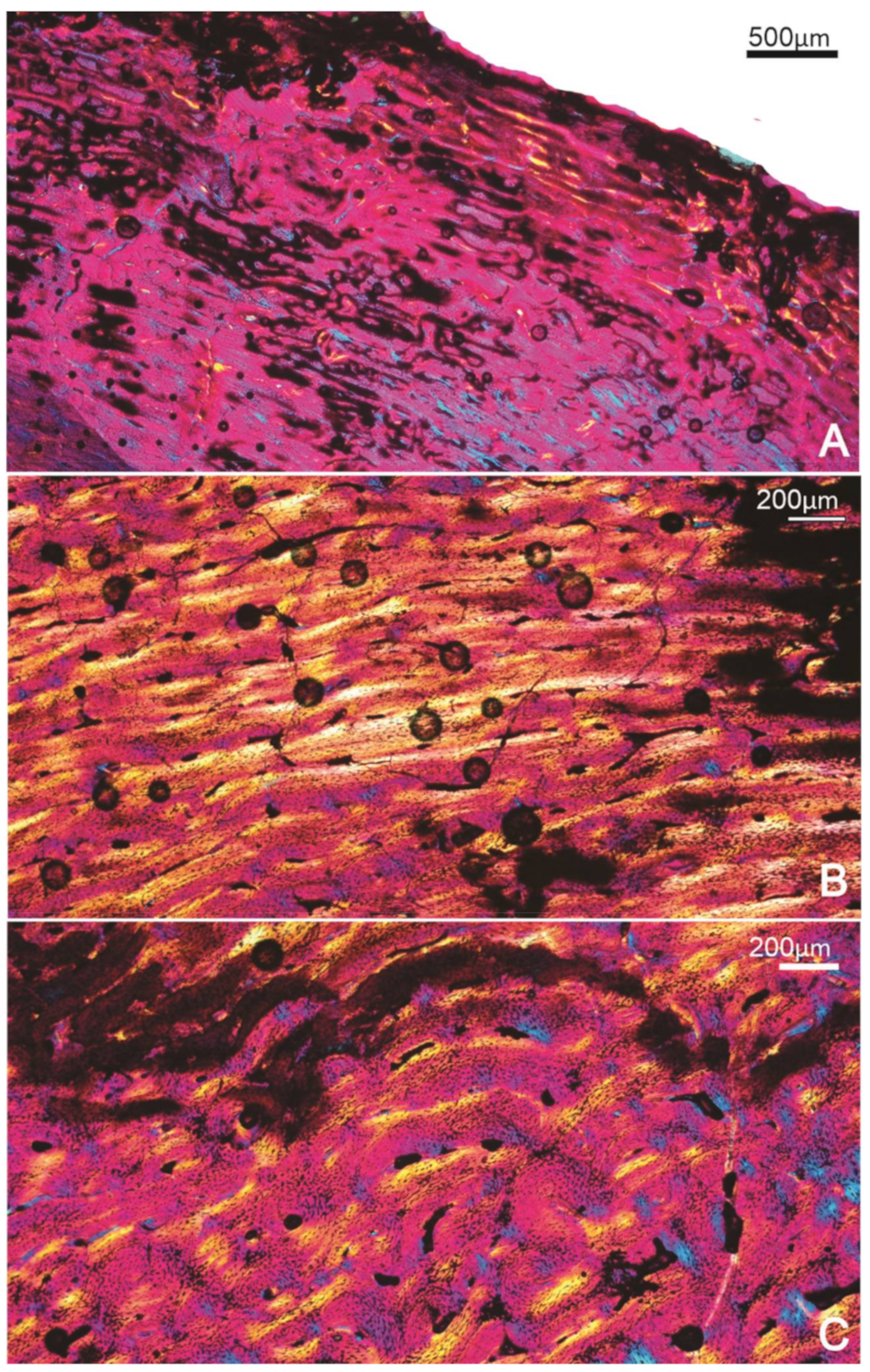
3.1.1. FU2759, Billeroo Creek, F
3.1.2. Specimen SAM P.54333, Cooper Creek (Geny C), ?F
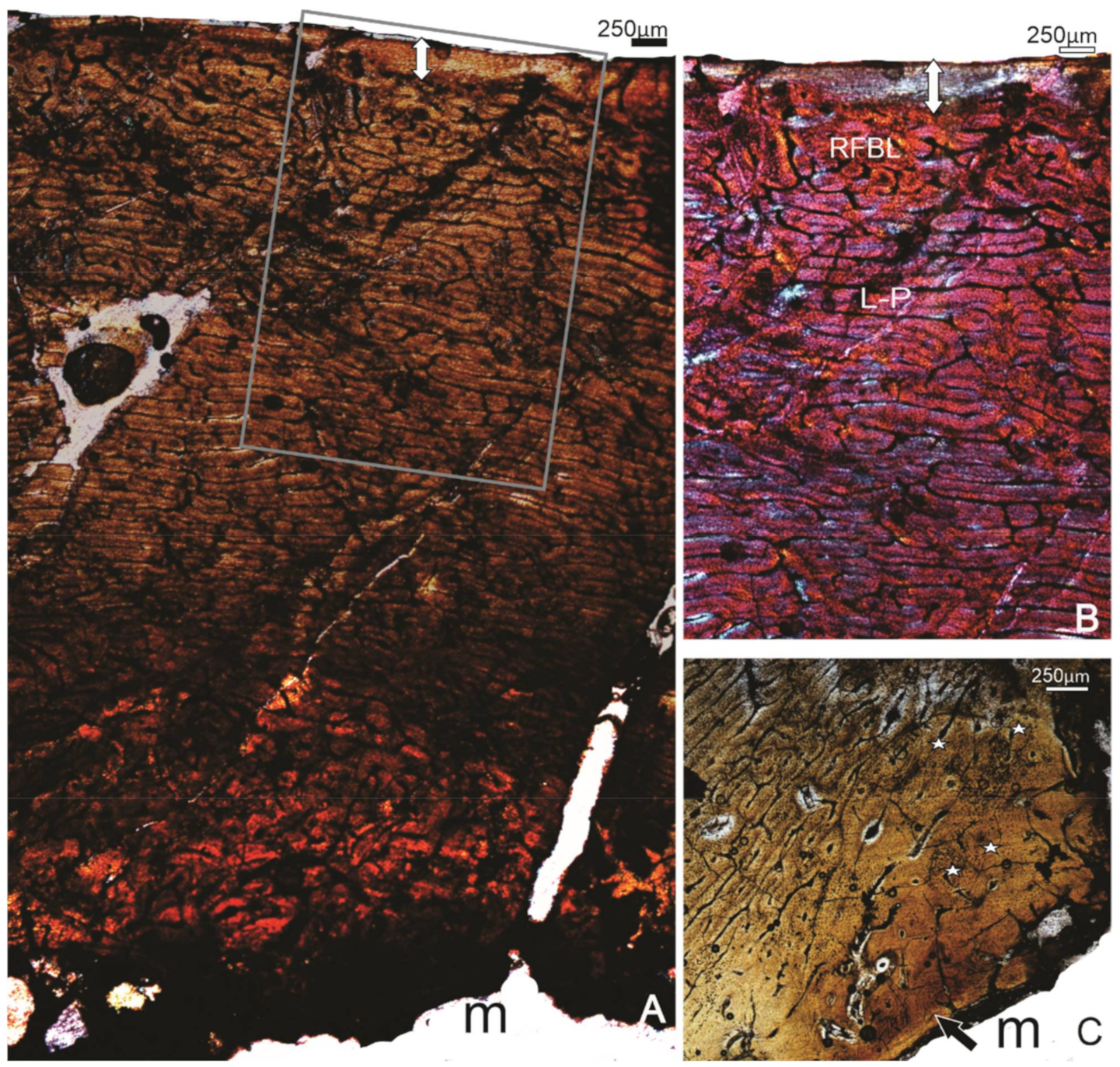
3.1.3. Specimen SAM P.54334, Cooper Creek site 73B, F
3.1.4. Specimen SAM P.53826, Callabonna Geny 1A, F
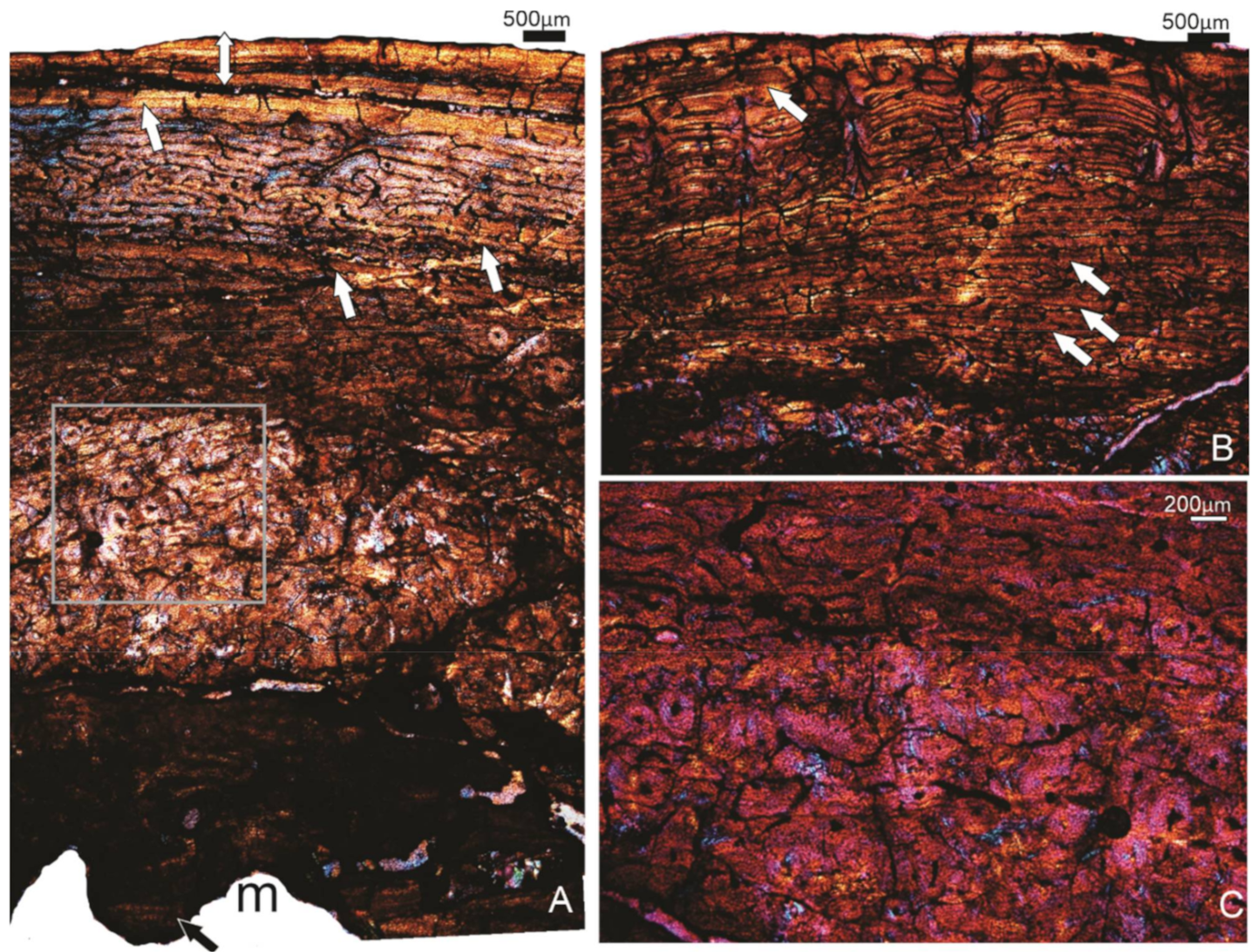
3.1.5. Specimen FU2756 Callabonna CB2018-75, Indiv 2, ?M/F
3.1.6. Specimen FU2755, Callabonna CB2018-75, Indiv 1, F
3.1.7. Specimen SAM P.25017, Cooper Creek, Malkuni WH, F
3.1.8. Specimen SAM P.53833, Callabonna, Geny 10, F?
3.1.9. Summary of the Tibiotarsus Histology
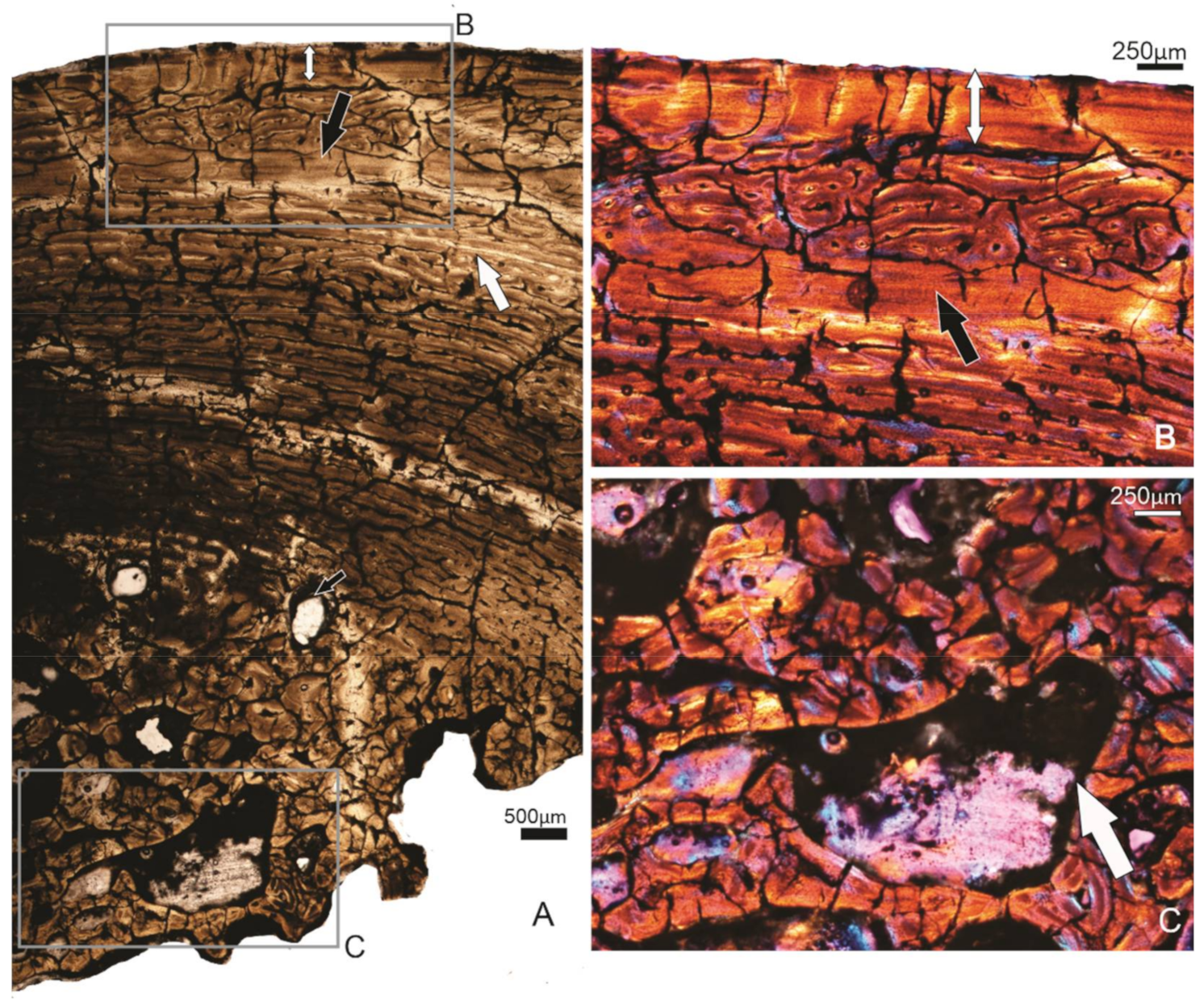
3.2. Femora
3.2.1. Specimen FU2758 Left Femur, Billeroo Creek
3.2.2. Specimen FU2760, Callabonna CB2018-75, Indiv 3, M
3.2.3. Specimen FU2755, Callabonna CB2018-75, Indiv 1, F
3.2.4. Specimen SAM P.53833, Callabonna Geny10, ?F
3.2.5. Summary of the Femoral Histology
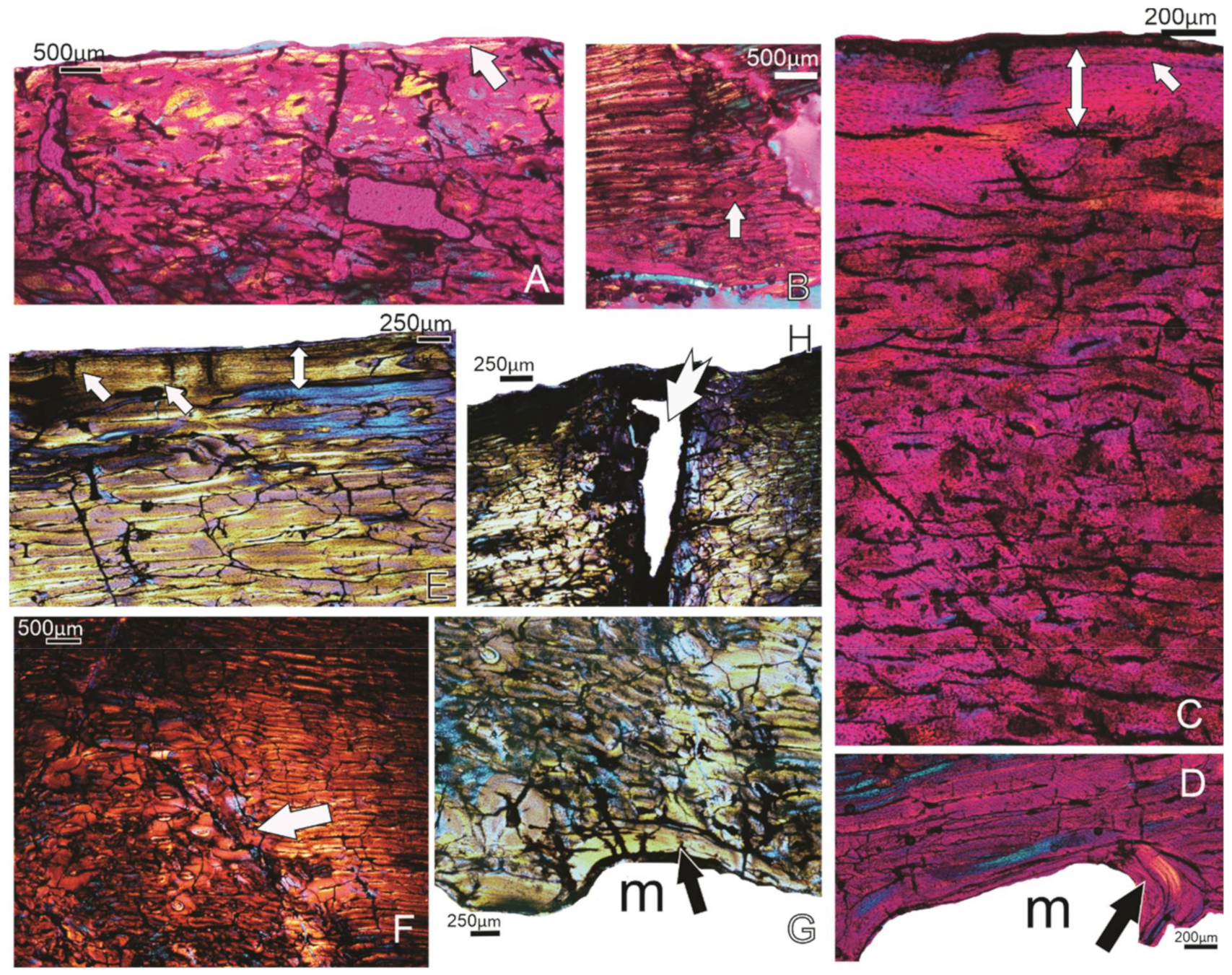
3.3. Tarsometatarsi
3.3.1. Specimen FU2750, Callabonna CB2018-98
3.3.2. Specimen SAM P.53832, Callabonna, Geny 9B, M
3.3.3. Specimen SAM P.53831, Callabonna, Geny 9A, M
3.3.4. Summary of Tarsometatarsi Histology
4. Discussion
4.1. Growth Pattern
4.2. Histological Variations Evident in Different Skeletal Elements
4.2.1. Histological Differences among Bones without OCL
4.2.2. Histological Variations Evident among Bones with OCL
4.2.3. Histological Differences among Specimens Recovered from the Same Site
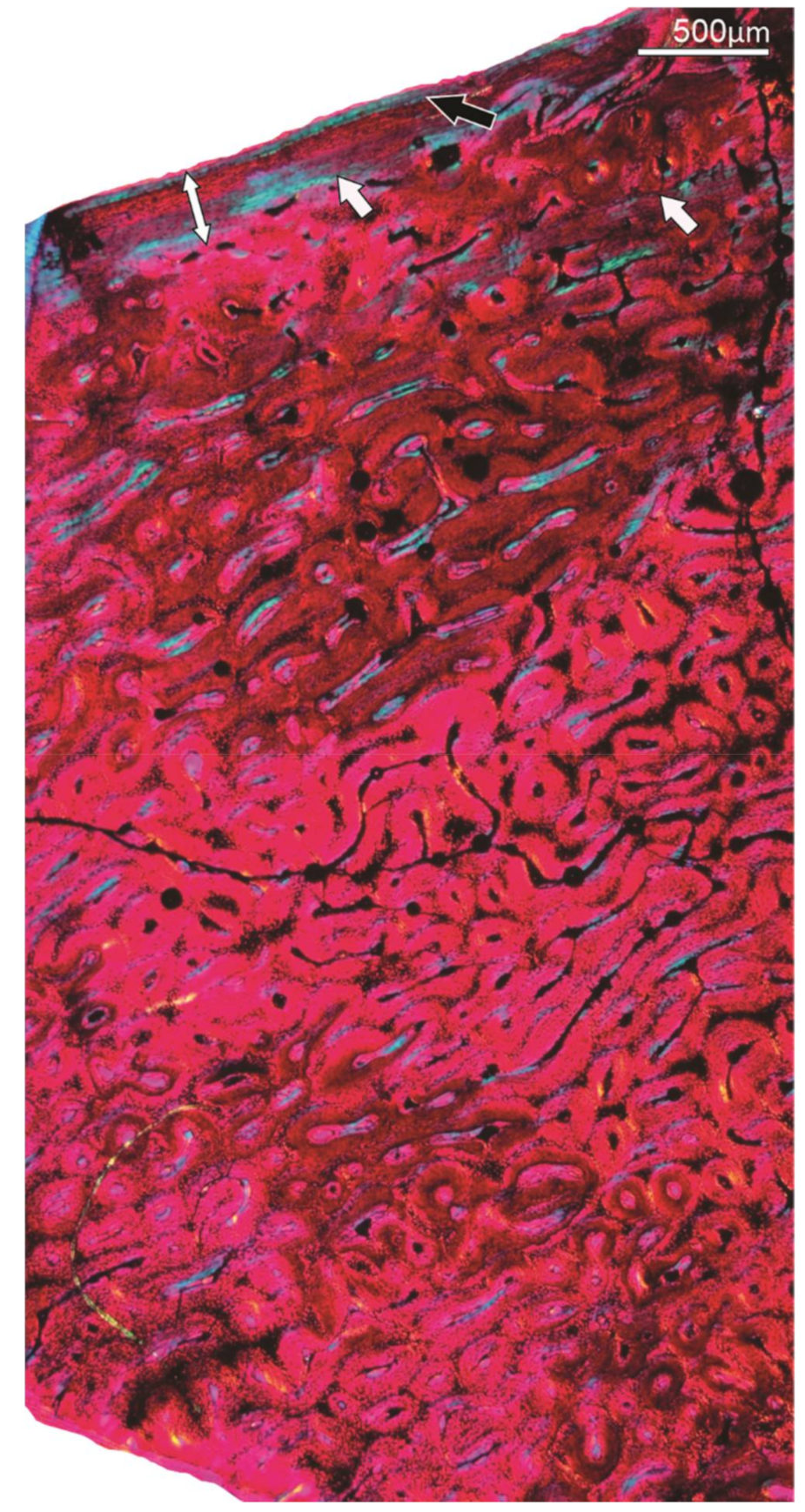
4.3. Secondary Reconstruction
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chinsamy-Turan, A. The Microstructure of Dinosaur bones: Deciphering Biology through Fine Scale Techniques; John Hopkins University Press: Baltimore, MD, USA, 2005. [Google Scholar]
- Chinsamy-Turan, A. The Forerunners of Mammals: Radiation, Histology, Biology; Indiana University Press: Bloomington, IN, USA, 2012. [Google Scholar]
- Erickson, G.M. Assessing dinosaur growth patterns: A microscopic revolution. Trends Ecol. Evol. 2005, 20, 677–684. [Google Scholar] [CrossRef]
- Erickson, G.M. On dinosaur growth. Ann. Rev. Earth Planetary Sci. 2014, 42, 675–697. [Google Scholar] [CrossRef]
- Chinsamy, A.; Angst, D.; Canoville, A.; Göhlich, U.B. Bone histology yields insights into the biology of the extinct elephant birds (Aepyornithidae) from Madagascar. Biol. J. Linn. Soc. 2020, 130, 268–295. [Google Scholar] [CrossRef]
- Chinsamy, A.; Warburton, N.M. Ontogenetic growth and the development of a unique fibrocartilage entheses in Macropus fuliginosus. Zoology 2020, 144, 125860. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.F.; Vickers-Rich, P. Magnificent Mihirungs: The Colossal Flightless Birds of the Australian Dreamtime; Indiana University Press: Bloomington, IN, USA, 2004. [Google Scholar]
- Worthy, T.H.; Degrange, F.J.; Handley, W.D.; Lee, M.S. The evolution of giant flightless birds and novel phylogenetic relationships for extinct fowl (Aves, Galloanseres). Roy. Soc. Open Sci. 2017, 4, 170975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirling, E.C.; Zietz, A.H.C. Fossil remains of Lake Callabonna. Part III. Description of the vertebrae of Genyornis newtoni. Mem. Roy. Soc. S. Aust. 1905, 1, 81–110. [Google Scholar]
- Stirling, E.C.; Zietz, A.H.C. Preliminary notes on Genyornis newtoni: A new genus and species of fossil struthious bird found at Lake Callabonna, South Australia. Trans. Proc. Rep. Roy. Soc. S. Aust. 1896, 20, 171–190. [Google Scholar]
- Saltré, F.; Rodríguez-Rey, M.; Brook, B.W.; Johnson, C.N.; Turney, C.S.; Alroy, J.; Cooper, A.; Beeton, N.; Bird, M.I.; Fordham, D.A. Climate change not to blame for late Quaternary megafauna extinctions in Australia. Nat. Commun. 2016, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Grellet-Tinner, G.; Spooner, N.; Handley, W.D.; Worthy, T.H. The Genyornis egg: Response to Miller et al.’s commentary on Grellet-Tinner et al., 2016. Quat. Sci. Rev. 2017, 61, 128–133. [Google Scholar] [CrossRef]
- Handley, W.D.; Chinsamy, A.; Yates, A.M.; Worthy, T.H. Sexual dimorphism in the late Miocene mihirung Dromornis stirtoni (Aves: Dromornithidae) from the Alcoota Local Fauna of central Australia. J. Vert. Paleontol. 2016, 36, e1180298. [Google Scholar] [CrossRef]
- Chinsamy, A.; Chiappe, L.M.; Dodson, P. Growth rings in Mesozoic birds. Nature 1994, 368, 196–197. [Google Scholar] [CrossRef]
- Starck, J.M.; Chinsamy, A. Bone microstructure and developmental plasticity in birds and other dinosaurs. J. Morphol. 2002, 254, 232–246. [Google Scholar] [CrossRef]
- Chinsamy, A. Histological perspectives on growth in the birds Struthio camelus and Sagittarius serpentarius. Acta Palaeornithol. 1995, 181, 317–323. [Google Scholar]
- Chinsamy, A. The Osteohistology of Femoral Growth within a Clade: A Comparison of a Crocodile, Crocodylus niloticus, the Dinosaurs, Massospondylus and Syntarsus and the Birds, Struthio and Sagittarius. Ph.D. Thesis, University of the Witwatersrand, Johannesburg, South Africa, 1990. [Google Scholar]
- Ponton, F.; Elżanowski, A.; Castanet, J.; Chinsamy, A.; Margerie, E.D.; de Ricqlès, A.; Cubo, J. Variation of the outer circumferential layer in the limb bones of birds. Acta Ornithol. 2004, 39, 137–140. [Google Scholar] [CrossRef] [Green Version]
- Bourdon, E.; Castanet, J.; de Ricqlès, A.; Scofield, P.; Tennyson, A.; Lamrous, H.; Cubo, J. Bone growth marks reveal protracted growth in New Zealand kiwi (Aves, Apterygidae). Biol. Lett. 2009, 5, 639–642. [Google Scholar] [CrossRef]
- Turvey, S.T.; Green, O.R.; Holdaway, R.N. Cortical growth marks reveal extended juvenile development in New Zealand moa. Nature 2005, 435, 940–943. [Google Scholar] [CrossRef]
- De Ricqlès, A.; Padian, K.; Horner, J.R. The bone histology of basal birds in phylogenetic and ontogenetic perspectives. In New Perspectives on the Origin and Early Evolution of Birds: Proceedings of the International Symposium in Honor of John, H. Ostrom; Gauthier, J., Gall, L.F., Eds.; Allen Press: Lawrence, KS, USA, 2001; pp. 411–426. [Google Scholar]
- Nanson, G.C.; Price, D.M.; Jones, B.G.; Maroulis, J.C.; Coleman, M.; Bowman, H.; Cohen, T.J.; Pietsch, T.J.; Larsen, J.R. Alluvial evidence for major climate and flow regime changes during the middle and late Quaternary in eastern central Australia. Geomorphology 2008, 101, 109–129. [Google Scholar] [CrossRef] [Green Version]
- Chinsamy, A.; Raath, M.A. Preparation of fossil bone for histological examination. Palaeontol. Afr. 1992, 29, 39–44. [Google Scholar]
- Francillon-Vieillot, H.; De Buffrénil, V.; Castanet, J.; Géraudie, J.; Meunier, F.J.; Sire, J.Y.; Zylberberg, L.; De Ricqlès, A. Microstructure and mineralisation of vertebrate skeletal tissues. In Skeletal Biomineralisation: Patterns, Processes and Evolutionary Trends; Carter, J.G., Ed.; Van Nostrand Reinhold: New York, NY, USA, 1990; pp. 471–530. [Google Scholar]
- De Margerie, E.; Robin, J.P.; Verrier, D.; Cubo, J.; Groscolas, R.; Castanet, J. Assessing a relationship between bone microstructure and growth rate: A fluorescent labelling study in the king penguin chick (Aptenodytes patagonicus). J. Exp. Biol. 2004, 207, 869–879. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, J. Ontogeny of surface texture of limb bones in modern aquatic birds and applicability of textural ageing. Anat. Rec. 2018, 301, 1026–1045. [Google Scholar] [CrossRef] [Green Version]
- Castanet, J.; Vieillot, H.F.; Meunier, F.J.; De Ricqlès, A. Bone and individual aging. In Bone, Vol. 7, Bone Growth; Hall, B.K., Ed.; CRC Press: Boca Raton, FL, USA, 1993; pp. 245–283. [Google Scholar]
- Angst, D.; Chinsamy, A.; Steel, L.; Hume, J.P. Bone histology sheds new light on the ecology of the dodo (Raphus cucullatus, Aves, Columbiformes). Sci. Rep. 2017, 7, 7993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanet, J.; Grandin, A.; Arbourachid, A.; de Ricqles, A. Expression of growth dynamic in the structure of the periosteal bone in the mallard, Anas platyrhynchos. C. R. Acad. Sci. Paris Ser. 3 Sci. Vie 1996, 319, 301–308. [Google Scholar]
- Hansford, J.P.; Turvey, S.T. Unexpected diversity within the extinct elephant birds (Aves: Aepyornithidae) and a new identity for the world’s largest bird. R. Soc. Open Sci. 2018, 5, 181295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinsamy, A.; Buffetaut, E.; Angst, D.; Canoville, A. Insight into the growth dynamics and systematic affinities of the Late Cretaceous Gargantuavis from bone microstructure. Naturwissenschaften 2014, 101, 447–452. [Google Scholar] [CrossRef]
- Hume, J.P.; Steel, L. Fight club: A unique weapon in the wing of the solitaire, Pezophaps solitaria (Aves: Columbidae), an extinct flightless bird from Rodrigues, Mascarene Islands. Biol. J. Linn. Soc. 2013, 110, 32–44. [Google Scholar] [CrossRef] [Green Version]
- Turvey, S.T.; Holdaway, R.N. Postnatal ontogeny, population structure, and extinction of the giant moa Dinornis. J. Morphol. 2005, 265, 70–86. [Google Scholar] [CrossRef]
- Chinsamy, A.; Marugán-Lobón, J.; Serrano, F.J.; Chiappe, L. Osteohistology and life history of the basal pygostylian, Confuciusornis sanctus. Anat. Rec. 2019, 303, 949–996. [Google Scholar] [CrossRef]
- De Ricqlès, A.; Bourdon, E.; Legendre, L.J.; Cubo, J. Preliminary assessment of bone histology in the extinct elephant bird Aepyornis (Aves, Palaeognathae) from Madagascar. Comptes Rendus Palevol. 2016, 15, 197–208. [Google Scholar] [CrossRef]
- Martin, R.B.; Burr, D.B. Structure, Function, and Adaptation of Compact Bone; Raven Pr: New York, NY, USA, 1989. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catalogue No. | Locality | Field ID | Element | Histology Sample | Comment | Sex |
|---|---|---|---|---|---|---|
| SAM P.25017 | Cooper Creek | Distal left tibiotarsus | y | distal width, 85 mm; minimum shaft circumference, 140 mm; sampled section of shaft 1/3 length from distal end. | Female? | |
| SAM P.53826 | Callabonna | Geny 1A | Distal right tibiotarsus | y | distal width, 88 mm, shaft inflated by salt degradation; sampled caudal facies. | Female? |
| SAM P.53826 | Callabonna | Geny 1A | Right tarsometatarsus | max proximal width, 105 mm; min shaft width, 45 mm; distal width, 99 mm | ||
| SAM P.53831 | Callabonna | Geny 9A | Right tarsometatarsus | y | Length trochlea III-cotyla lateralis, 358 mm; min shaft diameter, 50 mm; distal width, 110 mm; sampled medioplantar facies. | Male |
| SAM P.53832 | Callabonna | Geny 9B | Left tarsometatarsus | y | min shaft diameter, 44 mm; distal width, 95 mm; sampled medioplantar facies. | Female |
| SAM P.53833 | Callabonna | Geny 10 | Right femur | y | midshaft diameter, 76 mm; max distal width, 154 mm, sampled mid-caudal facies | |
| SAM P.53833 | Callabonna | Geny 10 | Left tibiotarsus | y | distal width, 86 mm; total length, 610 mm; minimum shaft circumference, 144 mm; sampled mid-medial facies | Female? |
| SAM P.53833 | Callabonna | Geny 10 | Right tarsometatarsus | length, 347 mm; TL2, 355 mm, distal width medial-lateral, 102; min shaft width, 39 mm; max distal width, 92 mm | ||
| SAM P.54333 | Cooper Creek | Geny C | Distal left tibiotarsus | y | distal width, 91 mm; sampled anterolateral facies | Female? |
| SAM P.54334 | Cooper Creek | Cooper Creek 73-B | Distal left tibiotarsus | y | distal width, 92 mm; minimum shaft circumference, ±140 mm; sampled section midshaft | Female? |
| FU2750 | Callabonna | CB2018-98 | part left tarsometatarsus | y | min shaft diameter, 38 mm; sampled medial facies | Female? |
| FU2755 | Callabonna | CB2018-75 Ind 1 | Right femur | y | shaft width, 87 mm; small indiv; sampled mid-caudal facies | |
| FU2755 | Callabonna | CB2018-75 Ind 1 | Right tibiotarsus | y | distal width, 88 mm; minimum shaft circumference, 144 mm; small indiv; 2 samples distomedial facies. | Female? |
| FU2755 | Callabonna | CB2018-75 Ind 1 | Left tarsometatarsus | distal width, 90 mm; min shaft width, 40 mm; max proximal width, 102 mm; small indiv. | ||
| FU2756 | Callabonna | CB2018-75 Ind 2 | Crushed left and right femora | |||
| FU2756 | Callabonna | CB2018-75 Ind 2 | Right tibiotarsus | y | distal width, 96 mm; minimum shaft circumference 146 mm; big indiv; sampled midshaft medial facies | boundary- male/female? |
| FU2756 | Callabonna | CB2018-75 Ind 2 | Left tarsometatarsus | min shaft diameter, 46 mm; max proximal width, 109 mm; max distal width, ±105 mm | ||
| FU2760 | Callabonna | CB2018-75 Ind 3 | Right femur | y | midshaft width, ~66 mm; sampled mid-caudal facies | Male |
| FU2758 | Billeroo Creek | NA | Left femur | y | midshaft width, 70 mm; surface texture is porous; crista trochanteris is not fully formed; sampled mid-caudal facies. | young indiv? |
| FU2759 | Billeroo Creek | NA | Distal left tibiotarsus | y | Sampled medial facies | Small-female? |
| Specimen | Element | OCL | Lags in OCL | Annuli | ICL | Dense Hav Bone | Early Bone | Resorpt. Perimedull. | Ontog. State |
|---|---|---|---|---|---|---|---|---|---|
| SAM P.54334 | Tibiotarsus | Y | 4–5 | 3–4 | Y | Y | Y | Y | adult |
| SAM P.54333 | Tibiotarsus | Y | N | 1? | Y | N | Y | Y | young adult |
| SAM P.53833 | Femur | Y | 2 | Y | Y | Y | N | Y | adult |
| SAM P.53833 | Tibiotarsus | Y | 3 | 2? | N | Y | N | Y | adult |
| SAM P.53832 | Tarsometatarsus | Y | 2? | N | Y | Y | N | Y | adult |
| SAM P.53831 | Tarsometatarsus | Y | 2? | N | ? | Y | N | Y | adult |
| SAM P.53826 | Tibiotarsus | Y | 3–4 | 2? | ? | ? | ? | ? | adult |
| SAM P.25017 | Tibiotarsus | Y | 5 | 3 | Y | N | Y | Y | adult |
| FU2760 | Femur | Y | 1 | ? | Y | N | N | Y | adult |
| FU2759 | Tibiotarsus | N | N | N | N | N | Y | ? | immature |
| FU2758 | Femur | Y | N | N | Y | N | Y | Y | young adult |
| FU2756 | Tibiotarsus | Y | ~10 | 1 | N | N | Y | Y | mature |
| FU2750 | Tarsometatarsus | Y | 1 | 2 | ? | N | Y | Y | young adult |
| FU2755 | Tibiotarsus | ? | ? | ? | ? | N | Y? | ? | ? |
| FU2755 | Femur | Y | 6–7 | ? | ? | N | Y? | ? | mature |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chinsamy, A.; Worthy, T.H. Histovariability and Palaeobiological Implications of the Bone Histology of the Dromornithid, Genyornis newtoni. Diversity 2021, 13, 219. https://doi.org/10.3390/d13050219
Chinsamy A, Worthy TH. Histovariability and Palaeobiological Implications of the Bone Histology of the Dromornithid, Genyornis newtoni. Diversity. 2021; 13(5):219. https://doi.org/10.3390/d13050219
Chicago/Turabian StyleChinsamy, Anusuya, and Trevor H. Worthy. 2021. "Histovariability and Palaeobiological Implications of the Bone Histology of the Dromornithid, Genyornis newtoni" Diversity 13, no. 5: 219. https://doi.org/10.3390/d13050219
APA StyleChinsamy, A., & Worthy, T. H. (2021). Histovariability and Palaeobiological Implications of the Bone Histology of the Dromornithid, Genyornis newtoni. Diversity, 13(5), 219. https://doi.org/10.3390/d13050219