
Polychaete Diversity Related to Different Mesophotic Bioconstructions along the Southeastern Italian Coast

 ,
,  ,
,
Abstract
:1. Introduction
2. Material and Methods
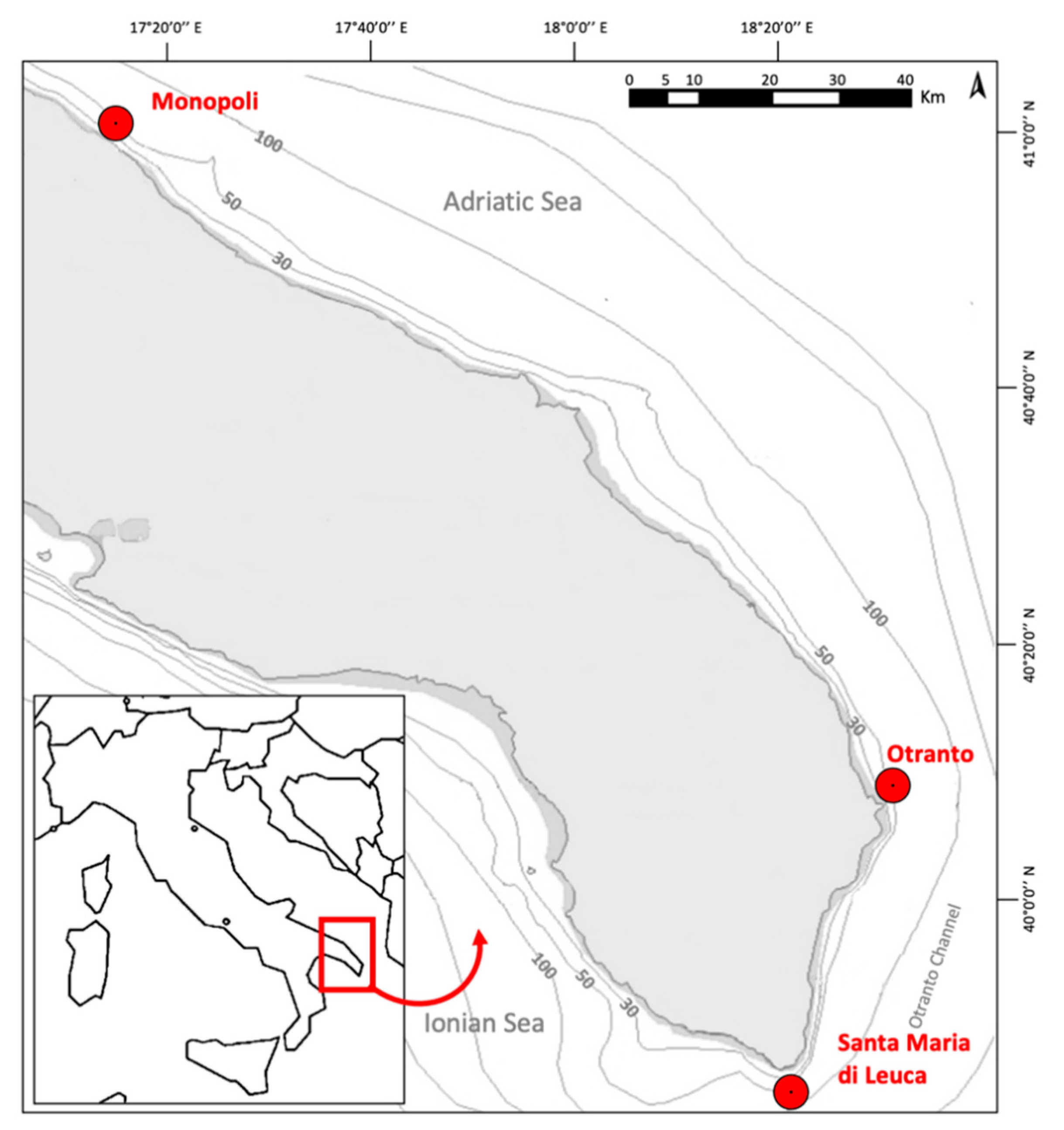
2.1. Study Areas
2.2. Sampling Methods and Taxonomic Analysis
2.3. Data Analysis
3. Results
3.1. Species Composition and Diversity
3.2. Taxonomic Accounts
- Antennae smooth, short and terminating at the same height. Branchiae starting far from the prostomium from 60°–70° chaetigers; peristomial cirri short not reaching the middle of the anterior peristomium ring………………………… Eunice schizobranchiaBranchiae starting within the first chaetigers…………………………………………….… 2
- Acicula yellow…………………………………………………………………………… 3Acicula black………………………………………………………………………………….… 6
- Acicular hooks bidentate………………………………………………………………… 4Acicular hooks tridentate……………………………………………………………………… 5
- Antennae indistinctly annulated or smooth; branchiae pectinate with up to 10–12 filaments, starting at the 3° chaetiger and lacking in the posterior part of the body……………………………………………………………………… Eunice pennataAntennae clearly annulated long, with long cylindrical articles; branchiae pectinate with up to 10–16 filaments; first branchiae at 3°–4° chaetiger and to near posterior end………………………………………………………………………… Eunice harassii
- Antennae deeply annulated, moniliform with short segments. Branchiae pectinate with maximally 12 filaments, starting from chaetiger 4°–7° and present in the posterior segments…………………………………………………………… Eunice antennataAntennae indistinctly articulated. Branchiae pectinate with maximally 12–14 filaments. starting from chaetiger 3 and lacking in the posterior segments………… Eunice vittata
- Antennae smooth or indistinctly articulated ………………………………………… 7Antennae regularly articulated or moniliformis …………………………………………… 8
- Antennae smooth and short, similar in length reaching the 2°–4° chaetiger, branchiae starting at 6°–10°, often 8°–9°, chaetiger, pectinate with numerous up to 15–40 filaments longer than the notopodial cirri, very large species ….………………….……………………………………………………… Eunice roussaeiAntennae indistinctly articulated and long, first branchiae on chaetiger 6°–10° often 7°, pectinate with 3–12 filaments, about as long as the notopodial cirri …………………………………………………………………………… Eunice norvegica
- Antennae articulated with cylindrical articulations; branchiae starting from 3°–4° chaetiger, with 2–3 rarely 5–6 filaments…………………………………. Eunice oerstediAntennae clearly moniliform………………………………………………………………… 9
- First branchiae on chaetiger 7°–10° often 9°, branchiae pectinate with 4–8 filaments longer than the notopodial cirri; antennae articulated with moniliform increasingly distally articulations……………………………………………………… Eunice floridanaFirst branchiae starting before 7° chaetiger ………………………………………………… 10
- Branchiae poor developed, palmate with few 1–2 maximally 3 filaments, starting from 3° chatiger………………………………………………………… Eunice dubitataBranchiae well developed…………………………………………………………………… 11
- Branchiae starting at 2°–3° chaetiger, pectinate with up to 8–10 filaments; body colour pattern uniform bright orange without spots nor whitish ring on anterior segment; subacicular hooks always in single arrangement……………………. Eunice laurillardiBranchiae starting from 3° chaetiger, pectinate with several up to 10–14, often 7, filaments, body colour pattern red brown with a white collar at the 4° chaetiger; subacicular hooks in double arrangement in medial and posterior segments……… Eunice torquata
4. Discussion
5. Taxonomic Considerations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giangrande, A.; Licciano, M.; Musco, L. Polychaetes as environmental indicators revisited. Mar. Pollut. Bull. 2005, 50, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Musco, L. Ecology and diversity of Mediterranean hard-bottom Syllidae (Annelida): A community level approach. Mar. Ecol. Prog. Ser. 2012, 461, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Giangrande, A.; Gravina, M.F. Brackish-water polychaetes, good descriptors of environmental changes in space and time. Transit. Waters Bull. 2015, 9, 42–55. [Google Scholar] [CrossRef]
- Sardá, R. Polychaete communities related to plant covering in the midlittoral and infralittoral zones of the Balearic Islands (Western Mediterranean). Mar. Ecol. 1991, 12, 341–360. [Google Scholar] [CrossRef]
- Giangrande, A.; Delos, A.L.; Musco, L.; Licciano, M.; Pierri, C. Polychaete assemblages of rocky shore along the South Adriatic coast (Mediterranean Sea). Cah. Biol. Mar. 2004, 45, 85–95. [Google Scholar]
- Mikac, B.; Licciano, M.; Jaklin, A.; Iveša, L.; Giangrande, A.; Musco, L. Diversity and distribution patterns of hard bottom polychaete assemblages in the North Adriatic Sea (Mediterranean). Diversity 2020, 12, 408. [Google Scholar] [CrossRef]
- Ballesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. Oceanogr. Mar. Biol. Annu. Rev. 2006, 44, 123–195. [Google Scholar]
- Bedini, R.; Bonechi, L.; Piazzi, L. Spatial and temporal variability of mobile macro-invertebrate assemblages associated to coralligenous habitat. Medit. Mar. Sci. 2014, 15, 302–312. [Google Scholar] [CrossRef] [Green Version]
- Gatti, G.; Bianchi, C.N.; Morri, C.; Montefalcone, M.; Sartoretto, S. Coralligenous reefs state along anthropized coasts: Application and validation of the COARSE index, based on a rapid visual assessment (RVA) approach. Ecol. Indic. 2015, 52, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Chimienti, G.; Stithou, M.; Dalle Mura, I.; Mastrototaro, F.; D’Onghia, G.; Tursi, A.; Izzi, C.; Fraschetti, S. An Explorative Assessment of the Importance of Mediterranean Coralligenous Habitat to Local Economy: The Case of Recreational Diving. J. Environ. Account. Manag. 2017, 5, 315–325. [Google Scholar] [CrossRef] [Green Version]
- Ingrosso, G.; Abbiati, M.; Badalamenti, F.; Bavestrello, G.; Belmonte, G.; Cannas, R.; Benedetti-Cecchi, L.; Bertolino, M.; Bevilacqua, S.; Bianchi, C.N.; et al. Mediterranean bioconstructions along the Italian coast. Adv. Mar. Biol. 2018, 79, 61–136. [Google Scholar]
- Aguilar, R.; Garcia, S.; Perry, A.L.; Alvarez, H.; Blanco, J.; Chimienti, G.; Montesanto, F.; Mastrototaro, F. Deep-sea habitats and communities in the Aeolian Islands (North Sicily). In Proceedings of the 2nd Mediterranean Symposium on the Conservation of Dark Habitats, Antalya, Turkey, 14–15 January 2019; Langar, H., Ouerghi, A., Eds.; SPA/RAC: Tunis, Tunisia, 2019; pp. 27–33. [Google Scholar]
- Bo, M.; Bavestrello, G.; Canese, S.; Giusti, M.; Angiolillo, M.; Cerrano, C.; Salvati, E.; Greco, S. Coral assemblage off the Calabrian Coast (South Italy) with new observations on living colonies of Antipathes dichotoma. Ital. J. Zool. 2011, 78, 231–242. [Google Scholar] [CrossRef]
- Bo, M.; Bavestrello, G.; Angiolillo, M.; Calcagnile, L.; Canese, S.; Cannas, R.; Cau, A.; D’Elia, M.; Filippo D’Oriano, F.; Follesa, M.C.; et al. Persistence of pristine deep-sea coral gardens in the Mediterranean Sea (SW Sardinia). PLoS ONE 2015, 10, e0119393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chimienti, G.; Bo, M.; Taviani, M.; Mastrototaro, F. Occurrence and biogeography of mediterranean cold-water corals. In Mediterranean Cold-Water Corals: Past, Present and Future; Orejas, C., Jiménez, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 213–243. [Google Scholar] [CrossRef]
- D’Onghia, G.; Capezzuto, F.; Cardone, F.; Carlucci, R.; Carluccio, A.; Chimienti, G.; Corriero, G.; Longo, C.; Maiorano, P.; Mastrototaro, F.; et al. Macro- and megafauna recorded in the submarine Bari Canyon (Southern Adriatic, Mediterranean Sea) using different tools. Mediterr. Mar. Sci. 2015, 16, 180–196. [Google Scholar] [CrossRef]
- Mastrototaro, F.; D’Onghia, G.; Corriero, G.; Matarrese, A.; Maiorano, P.; Panetta, P.; Gherardi, M.; Longo, C.; Rosso, A.; Sciuto, F.; et al. Biodiversity of the white coral bank off cape Santa Maria di Leuca (Mediterranean Sea): An update. Deep-Sea Res. Part II 2010, 57, 412–430. [Google Scholar] [CrossRef]
- Taviani, M.; Angeletti, L.; Cardone, F.; Montagna, P.; Danovaro, R. A unique and threatened deep water coral-bivalve biotope new to the Mediterranean Sea offshore the Naples megalopolis. Sci. Rep. 2019, 9, 3411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerrano, C.; Bastari, A.; Calcinai, B.; Di Camillo, C.; Pica, D.; Puce, S.; Valisano, L.; Torsani, F. Temperate mesophotic ecosystems: Gaps and perspectives of an emerging conservation challenge for the Mediterranean Sea. Eur. Zool. J. 2019, 86, 370–388. [Google Scholar] [CrossRef] [Green Version]
- Bo, M.; Bavestrello, G.; Canese, S.; Giusti, M.; Salvati, E.; Angiolillo, M.; Greco, S. Characteristics of a black coral meadow in the twilight zone of the central Mediterranean Sea. Mar. Ecol. Prog. Ser. 2009, 397, 53–61. [Google Scholar] [CrossRef]
- Cerrano, C.; Danovaro, R.; Gambi, C.; Pusceddu, A.; Riva, A.; Schiaparelli, S. Gold coral (Savalia savaglia) and gorgonian forests enhance benthic biodiversity and ecosystem functioning in the mesophotic zone. Biodivers. Conserv. 2010, 19, 153–167. [Google Scholar] [CrossRef] [Green Version]
- Corriero, G.; Pierri, C.; Mercurio, M.; Nonnis Marzano, C.; Tarantini, S.O.; Gravina, M.F.; Lisco, S.; Moretti, M.; De Giosa, F.; Valenzano, E.; et al. A Mediterranean mesophotic coral reef built by non-symbiotic scleractinians. Sci. Rep. 2019, 9, 3601. [Google Scholar] [CrossRef]
- Angeletti, L.; Taviani, M. Offshore Neopycnodonte Oyster Reefs in the Mediterranean Sea. Diversity 2020, 12, 92. [Google Scholar] [CrossRef] [Green Version]
- Cardone, F.; Corriero, G.; Longo, C.; Mercurio, M.; Tarantini, S.O.; Gravina, M.F.; Lisco, S.; Moretti, M.; De Giosa, F.; Giangrande, A.; et al. Massive bioconstructions built by Neopycnodonte cochlear (Mollusca, Bivalvia) in a mesophotic environment in the central Mediterranean Sea. Sci. Rep. 2020, 10, 6337. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.M. Reef-aggregating behaviour by symbiotic eunicid polychaetes from cold-water corals: Do worms assemble reefs? J. Mar. Biol. Assoc. U. K. 2005, 85, 813–819. [Google Scholar] [CrossRef]
- Mueller, C.E.; Lundälv, T.; Middelburg, J.J.; van Oevelen, D. The Symbiosis between Lophelia pertusa and Eunice norvegica stimulates coral calcification and worm assimilation. PLoS ONE 2013, 8, e58660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauvel, P. Polychètes Errantes. Faune Fr. 1923, 5, 1–488. [Google Scholar]
- Day, J.H. A Monograph on the Polychaeta of Southern Africa. Part I. Errantia; Trustees of the British Museum (Natural History): London, UK, 1967; p. 458. [Google Scholar]
- George, J.D.; Hartmann-Schröder, G. Polychaetes: British Amphinomida, Spintherida and Eunicida. Keys and notes for the identification of the species. Synop. Br. Fauna 1985, 32, 1–221. [Google Scholar]
- Fauchald, K. A review of the genus Eunice (Eunicidae: Polychaeta) based upon type material. Smithson. Contrib. Zool. 1992, 523, 1–422. [Google Scholar] [CrossRef] [Green Version]
- Cantone, G. Censimento dei policheti dei mari italiani: Eunicidae Berthold, 1827. Atti Soc. Toscana Sci. Nat. Resid. Pisa Mem. Ser. B 1993, 100, 229–243. [Google Scholar]
- D’Alessandro, M.; Romeo, T.; Castriota, L.; Cosentino, A.; Perzia, P.; Martins, R. New records of Lumbrineridae (Annelida:.Polychaeta) in the Mediterranean biogeographic province, with an updated taxonomic key. Ital. J. Zool. 2016, 83, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Zanol, J.; Bettoso, N. Identity of Eunice roussaei (Eunicidae: Polychaeta: Annelida) from the Adriatic and Mediterranean Seas. J. Mar. Biol. Assoc. U. K. 2006, 86, 1017–1024. [Google Scholar] [CrossRef]
- Zanol, J.; Fauchald, K.; Paiva, P.C. A phylogenetic analysis of the genus Eunice (Eunicidae, Polychaete, Annelida). Zool. J. Linn. Soc. 2007, 150, 413–434. [Google Scholar] [CrossRef] [Green Version]
- Salazar-Vallejo, S.I.; Carrera-Parra, L.F.; de León-González, J.A. Giant Eunicid Polychaetes (Annelida) in shallow tropical and temperate seas. Rev. Biol. Trop. 2011, 59, 1463–1474. [Google Scholar] [CrossRef] [Green Version]
- Zanol, J.; Halanych, K.M.; Fauchald, K. Reconciling taxonomy and phylogeny in the bristleworm family Eunicidae (Polychaete, Annelida). Zool. Scr. 2014, 43, 79–100. [Google Scholar] [CrossRef]
- Arias, A.; Fernández-Álvarez, F.Á.; Martins, R.; Anadón, N. Rediscovery and redescription of Leodice laurillardi (Quatrefages, 1866) comb. nov. (Annelida: Eunicidae)-A rare European polychaete or just an overlooked species? Zootaxa 2015, 3964, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Zanol, J.; Carrera-Parra, L.F.; Steiner, T.M.; Amaral, A.C.Z.; Wiklund, H.; Ravara, A.; Budaeva, N. The Current State of Eunicida (Annelida) Systematics and Biodiversity. Diversity 2021, 13, 74. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef] [Green Version]
- Koleff, P.; Gaston, K.J.; Lennon, J.J. Measuring beta diversity for presence-absence data. J. Anim. Ecol. 2003, 72, 367–382. [Google Scholar] [CrossRef] [Green Version]
- Castelli, A.; Bianchi, C.N.; Cantone, G.; Çinar, M.E.; Gambi, M.C.; Giangrande, A.; Iraci Sareri, D.; Lanera, P.; Licciano, M.; Musco, L.; et al. Annelida Polychaeta. Biol. Mar. Mediterr. 2008, 15, 323–373. [Google Scholar]
- Casellato, S.; Stefanon, A. Coralligenous habitat in the northern Adriatic Sea: An overview. Mar. Ecol. 2008, 29, 321–341. [Google Scholar] [CrossRef]
- Giampaoletti, J.; Cardone, F.; Corriero, G.; Gravina, M.F.; Nicoletti, L. Sharing and Distinction in Biodiversity and Ecological Role of Bryozoans in Mediterranean Mesophotic Bioconstructions. Front. Mar. Sci. 2020, 7, 581292. [Google Scholar] [CrossRef]
- Damiani, V.; Bianchi, C.N.; Ferretti, O.; Bedulli, D.; Morri, C.; Viel, M.; Zurlini, G. Risultati di una ricerca ecologica sul sistema marino pugliese. Thalass. Salentina 1988, 18, 153–169. [Google Scholar]
- Russo, A.; Artegiani, A. Adriatic Sea hydrography. Sci. Mar. 1996, 60, 33–43. [Google Scholar]
- Poulain, P.M. Adriatic Sea surface circulation as derived from drifter data between 1990 and 1999. J. Mar. Syst. 2001, 29, 3–32. [Google Scholar] [CrossRef]
- Zavatarelli, M.; Pinardi, N.; Kourafalou, V.H.; Maggiore, A. Diagnostic and prognostic model studies of the Adriatic Sea general circulation: Seasonal variability. J. Geophys. Res. Ocean. 2002, 107, 2-1–2-20. [Google Scholar] [CrossRef] [Green Version]
- Zavatarelli, M.; Pinardi, N. The Adriatic Sea modelling system: A nested approach. Ann. Geophys. 2003, 21, 345–364. [Google Scholar] [CrossRef]
- Giangrande, A.; Gambi, M.C.; Gravina, M.F. Paradigm shifts in community ecology: Open versus closed units, challenges and limits of connectivity studies. Mar. Ecol. 2017, 38, e12480. [Google Scholar] [CrossRef]
- Donnarumma, L.; Appolloni, L.; Chianese, E.; Bruno, R.; Baldrighi, E.; Guglielmo, R.; Russo, G.; Zeppilli, D.; Sandulli, R. Environmental and benthic community patterns of the shallow hydrothermal area of Secca delle Fumose (Baia, Naples, Italy). Front. Mar. Sci. 2019, 6, 685. [Google Scholar] [CrossRef] [Green Version]
- Longo, C.; Cardone, F.; Pierri, C.; Mercurio, M.; Mucciolo, S.; Nonnis Marzano, C.; Corriero, G. Sponges associated with coralligenous formations along the Apulian coasts. Mar. Biodivers. 2018, 48, 2151–2163. [Google Scholar] [CrossRef]
- Cardone, F.; Corriero, G.; Fianchini, A.; Gravina, M.F.; Nonnis Marzano, C. Biodiversity of transitional waters: Species composition and comparative analysis of hard bottom communities from the south-eastern Italian coast. J. Mar. Biol. Assoc. 2014, 94, 25–34. [Google Scholar] [CrossRef]
- Gravina, M.F.; Cabiddu, S.; Como, S.; Floris, A.; Padedda, B.M.; Pusceddu, A.; Magni, P. Disentangling heterogeneity and commonalities in nanotidal Mediterranean lagoons through environmental features and macrozoobenthic assemblages. Estuar. Coast. Shelf Sci. 2020, 237, 106688. [Google Scholar] [CrossRef]
- Cerrano, C.; Bavestrello, G.; Bianchi, C.N.; Calcinai, B.; Cattaneo-Vietti, R.; Morri, C.; Sarà, M. The Role of Sponge Bioerosion in Mediterranean Coralligenous Accretion. In Mediterranean Ecosystems; Faranda, F.M., Guglielmo, L., Spezie, G., Eds.; Springer: Milano, Italy, 2001; pp. 235–240. [Google Scholar] [CrossRef]
- Sanfilippo, R.; Mòllica, E. Serpula cavernicola Fassari & Mòllica, 1991 (Annelida Polychaeta); diagnostic features of the tube and new Mediterranean records. Mar. Life 2000, 10, 27–32. [Google Scholar]
- Calcinai, B.; Arillo, A.; Cerrano, C.; Bavastrello, G. Taxonomy-related differences in the excavating micro-patterns of boring sponges. J. Mar. Biol. Assoc. U. K. 2003, 83, 37–39. [Google Scholar] [CrossRef]
- Fauvel, P. Annelides Polychetes de L’Australie meridionale. Arch. Zool. Exp. Gen. (Paris) 1917, 56, 159–277, plates 4–8. [Google Scholar]
- Tursi, A.; Mastrototaro, F.; Matarrese, A.; Maiorano, P.; D’Onghia, G. Biodiversity of the white coral reefs in the Ionian Sea (Central Mediterranean). Chem. Ecol. 2004, 20, S107–S116. [Google Scholar] [CrossRef]
- Vertino, A.; Savini, A.; Rosso, A.; Di Geronimo, I. Benthic habitat characterization and distribution from two representative sites of the deep-water SML Coral Province (Mediterranean). Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 380–396. [Google Scholar] [CrossRef]
- Taviani, M.; Angeletti, L.; Canese, S.; Cannas, R.; Cardone, F.; Cau, A.; Cau, A.B.; Follesa, M.C.; Marchese, F.; Montagna, P.; et al. The “Sardinian cold-water coral province” in the context of the Mediterranean coral ecosystems. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 145, 61–78. [Google Scholar] [CrossRef]
- Corriero, G.; Gherardi, M.; Giangrande, A.; Longo, C.; Mercurio, M.; Musco, L.; Nonnis Marzano, C. Inventory and distribution of hard bottom fauna from the marine protected area of Porto Cesareo (Ionian Sea): Porifera and Polychaeta. It. J. Zool. 2004, 71, 237–245. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Site | Monopoli | Otranto | Santa Maria di Leuca | |
|---|---|---|---|---|
| Depth | 43 m | 52 m | 60 m | |
| Main Structuring species | Scleractinians | N. cochlear | N. cochlear | |
| Family | Species | |||
| Sabellidae Latreille, 1825 | Hypsicomus stichophthalmos (Grube, 1863) | x | ||
| Serpulidae Rafinesque, 1815 | Hydroides pseudouncinata (Zibrowius, 1968) | x | x | |
| Serpula vermicularis Linnaeus, 1767 | x | x | x | |
| Serpula cavernicola Fassari & Mollica, 1991 | x | x | x | |
| Serpula concharum Langerhans, 1880 | x | x | x | |
| Serpula lobiancoi Rioja, 1917 | x | x | ||
| Serpula israelitica Amoureux, 1977 | x | |||
| Spiraserpula massiliensis (Zibrowius, 1968) | x | x | x | |
| Vermiliopsis infundibulum (Philippi, 1844) | x | x | x | |
| Vermiliopsis striaticeps (Grube, 1862) | x | x | x | |
| Vermiliopsis monodiscus Zibrowius, 1968 | x | x | ||
| Vermiliopsis labiata (O. G. Costa, 1861) | x | x | x | |
| Bathyvermilia eliasoni (Zibrowius, 1970) | x | |||
| Metavermilia multicristata (Philippi, 1844) | x | x | x | |
| Semivermilia agglutinata (Marenzeller, 1893) | x | x | ||
| Semivermilia crenata (O. G. Costa, 1861) | x | x | x | |
| Semivermilia cribrata (O. G. Costa, 1861) | x | x | ||
| Semivermilia pomatostegoides (Zibrowius, 1969) | x | x | ||
| Filogranula gracilis Langerhans, 1884 | x | x | ||
| Filogranula calyculata (O. G. Costa, 1861) | x | x | ||
| Filogranula annulata (O. G. Costa, 1861) | x | x | ||
| Janita fimbriata (Delle Chiaje, 1822) | x | x | ||
| Spirobranchus lima (Grube, 1862) | x | |||
| Spirobranchus polytrema (Philippi, 1844) | x | |||
| Spirobranchus triqueter (Linnaeus, 1758) | x | x | x | |
| Placostegus tridentatus (Fabricius, 1779) | x | x | ||
| Verminia cristallina (Philippi, 1844) | x | x | x | |
| Josephella marenzelleri Caullery & Mesnil, 1896 | x | x | ||
| Filograna implexa Berkeley, 1835 | x | x | x | |
| Protula tubularia (Montagu, 1803) | x | x | ||
| Spirorbis cuneatus Gee, 1964 | x | |||
| Spirorbis marioni Caullery & Mesnil, 1897 | x | x | ||
| Protolaeospira striata (Quiévreux, 1963) | x | x | ||
| Janua heterostropha (Montagu, 1803) | x | x | ||
| Neodexiospira pseudocorrugata (Bush, 1905) | x | x | ||
| Pileolaria militaris Claparède, 1870 | x | x | ||
| Pileolaria heteropoma (Zibrowius, 1968) | x | |||
| Vinearia koehleri (Caullery & Mesnil, 1897) | x | |||
| Nidificaria clavus (Harris, 1968) | x | |||
| Euphrosinidae Williams, 1852 | Euphrosine foliosa Audouin & H Milne Edwards, 1833 | x | x | x |
| Eunicidae Berthold, 1827 | Eunice dubitata Fauchald, 1974 | x | x | |
| Eunice schizobranchia Claparède, 1870 | x | |||
| Eunice pennata (Müller, 1776) | x | |||
| Eunice floridana (Pourtalès, 1867) | x | |||
| Eunice torquata (Quatrefages, 1866) | x | x | ||
| Palola siciliensis (Grube, 1840) | x | x | ||
| Palola valida (Gravier, 1900) | x | |||
| Paucibranchia fallax (Marion & Bobretzky, 1875) | x | x | ||
| Lysidice collaris Grube, 1870 | x | x | x | |
| Lysidice ninetta Audouin & H Milne Edwards, 1833 | x | x | ||
| Lumbrineridae Schmarda, 1861 | Lumbrineris coccinea (Renier, 1804) | x | x | |
| Scoletoma laurentiana (Grube, 1863) | x | |||
| Oenonidae Kinberg, 1865 | Arabella geniculata (Claparède, 1868) | x | x | |
| Arabella iricolor (Montagu, 1804) | x | |||
| Drilonereis filum (Claparède, 1868) | x | x | ||
| Glyceridae Grube, 1850 | Glycera tesselata Grube, 1863 | x | ||
| Glycera unicornis Lamarck, 1818 | x | |||
| Goniadidae Kinberg, 1866 | Glycinde nordmanni (Malmgren, 1866) | x | ||
| Goniada emerita Audouin & H Milne Edwards, 1833 | x | |||
| Goniada maculata Örsted, 1843 | x | x | ||
| Nereididae Blainville, 1818 | Ceratonereis costae (Grube, 1840) | x | x | |
| Polynoidae Kinberg, 1856 | Harmothoe antilopes McIntosh, 1876 | x | ||
| Harmothoe pagenstecheri Michaelsen, 1896 | x | x | ||
| Lepidasthenia elegans (Grube, 1840) | x | |||
| Syllidae Grube, 1850 | Haplosyllis spongicola (Grube, 1855) | x | ||
| Syllis alternata Moore, 1908 | x | |||
| Syllis gracilis Grube, 1840 | x | x | ||
| Syllis ferrani Alós & San Martín, 1987 | x | |||
| Syllis variegata Grube, 1860 | x | x | ||
| Sphaerosyllis hystrix Claparède, 1863 | x |
| Sites | α | SI | β |
|---|---|---|---|
| MON (α)/MON-OTR (SI, β) | 40 | 0.59 | 0.41 |
| OTR (α)/MON-SML (SI, β) | 46 | 0.43 | 0.55 |
| SML (α)/OTR-SML (SI, β) | 42 | 0.64 | 0.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gravina, M.F.; Pierri, C.; Mercurio, M.; Nonnis Marzano, C.; Giangrande, A. Polychaete Diversity Related to Different Mesophotic Bioconstructions along the Southeastern Italian Coast. Diversity 2021, 13, 239. https://doi.org/10.3390/d13060239
Gravina MF, Pierri C, Mercurio M, Nonnis Marzano C, Giangrande A. Polychaete Diversity Related to Different Mesophotic Bioconstructions along the Southeastern Italian Coast. Diversity. 2021; 13(6):239. https://doi.org/10.3390/d13060239
Chicago/Turabian StyleGravina, Maria Flavia, Cataldo Pierri, Maria Mercurio, Carlotta Nonnis Marzano, and Adriana Giangrande. 2021. "Polychaete Diversity Related to Different Mesophotic Bioconstructions along the Southeastern Italian Coast" Diversity 13, no. 6: 239. https://doi.org/10.3390/d13060239
APA StyleGravina, M. F., Pierri, C., Mercurio, M., Nonnis Marzano, C., & Giangrande, A. (2021). Polychaete Diversity Related to Different Mesophotic Bioconstructions along the Southeastern Italian Coast. Diversity, 13(6), 239. https://doi.org/10.3390/d13060239