
Mesofauna at the Soil-Scree Interface in a Deep Karst Environment


Abstract
:
1. Introduction
2. Materials and Methods
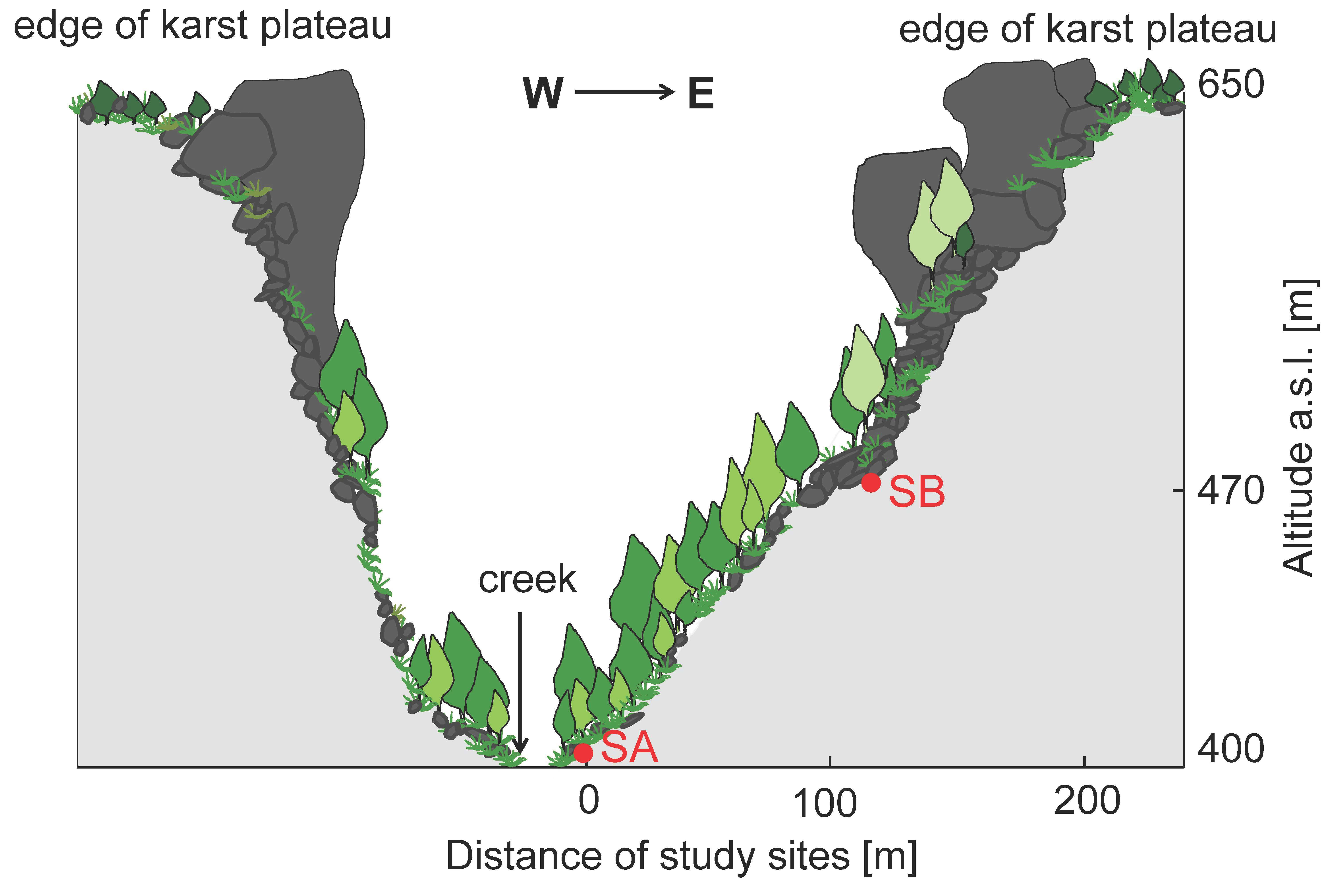
2.1. Study Area and Site Description
2.2. Field Sampling
2.3. Soil Microclimatic and Soil-Chemical Data
2.4. Collembola Diversity, Abundance and Vertical Distribution
2.5. Ecological Groups of Collembola
2.6. Statistical Analyses
3. Results
3.1. Soil Microclimatic and Soil-Chemical Data
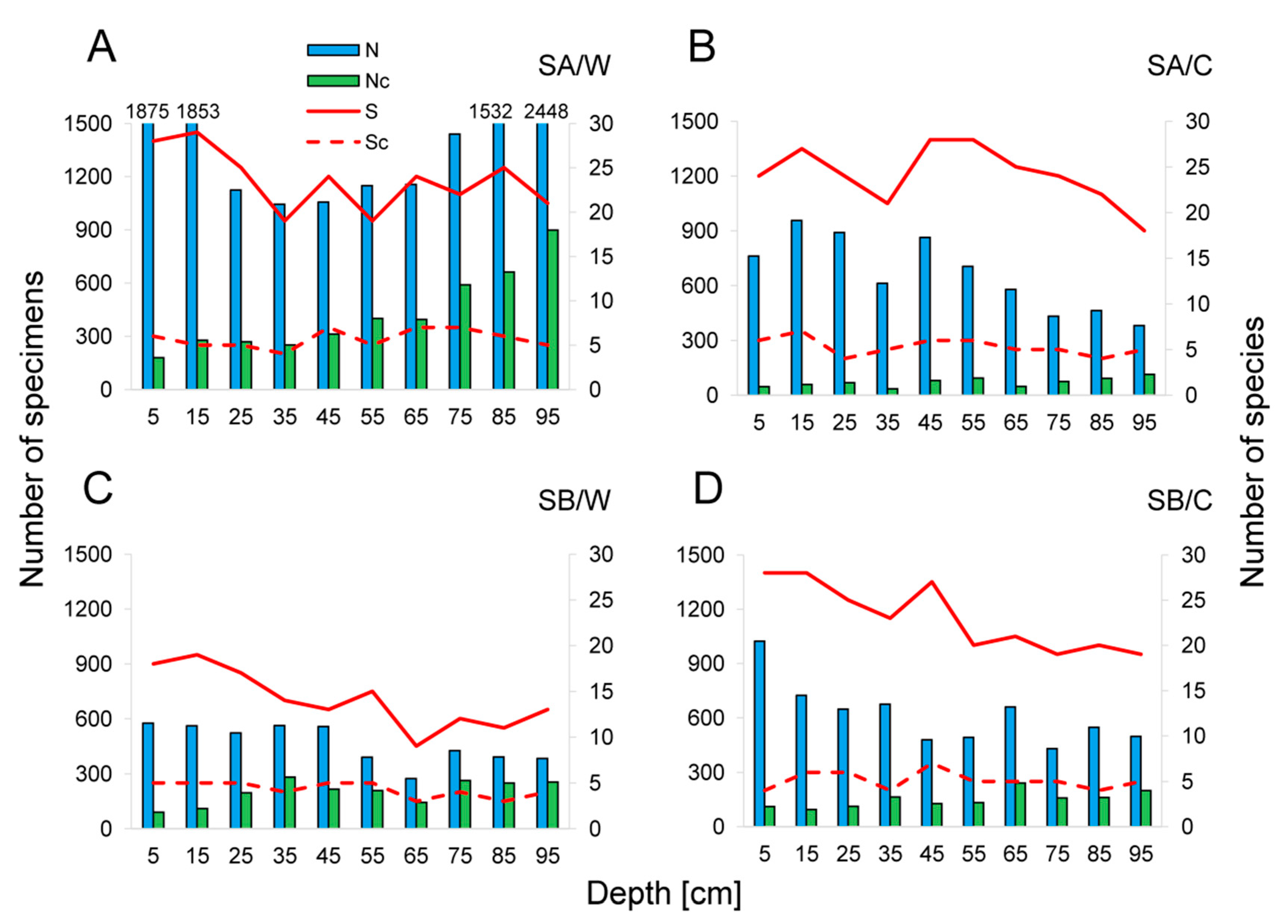
3.2. Collembola Diversity, Abundance and Vertical Distribution
3.3. Vertical Distribution of Ecological Groups
3.4. Community Composition
4. Discussion
4.1. Soil Microclimate and Soil-Chemical Parameters
4.2. Species Diversity, Vertical Distribution and Community Structure of Collembola at Scree Sites
4.3. Ecological Groups of Collembola and Climatic Conditions in the Two Periods
4.4. Scree Habitats of Deep Karst Valleys as Climate Refugia
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| W | C | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Ech | 5 | 15 | 25 | 35 | 45 | 55 | 65 | 75 | 85 | 95 | Nw | 5 | 15 | 25 | 35 | 45 | 55 | 65 | 75 | 85 | 95 | Nc |
| Allacma fusca (Linnaeus, 1758) | tgx | 2 | - | - | - | - | - | - | - | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - |
| Ceratophysella denticulata (Bagnall, 1941) | etp | - | - | 1 | - | - | - | - | 1 | - | 4 | 6 | - | - | - | - | 4 | - | 2 | 1 | - | - | 7 |
| Ceratophysella granulata Stach, 1949 | etp, p a | 37 | 106 | 114 | 134 | 172 | 129 | 176 | 275 | 323 | 531 | 1997 | 6 | 18 | 33 | 16 | 43 | 33 | 20 | 30 | 58 | 68 | 325 |
| Ceratophysella sigillata (Uzel, 1891) | tgx, p a | - | - | - | - | - | - | - | - | - | - | - | 11 | 5 | 8 | 4 | 10 | 3 | 5 | 3 | 5 | 4 | 58 |
| Ceratophysella silvatica (Rusek, 1964) | stp, p a | - | - | - | - | 2 | - | 3 | 2 | 4 | - | 11 | - | - | - | - | - | - | - | - | - | - | - |
| Deharvengiurus denisi (Stach, 1934) | stp, p b | 1 | - | - | - | - | - | - | - | - | - | 1 | - | 2 | - | - | - | - | - | - | - | - | 2 |
| Desoria germanica (Hüther et Winter, 1961) | tgx | - | - | - | - | - | - | - | - | - | - | - | - | 2 | - | - | - | - | - | - | - | - | 2 |
| Desoria hiemalis (Schött, 1839) | stp | 2 | 18 | - | - | 2 | - | - | 1 | - | - | 23 | 10 | 10 | 10 | - | 6 | 2 | - | 1 | - | 1 | 40 |
| Desoria tigrina Nicolet, 1842 | stp | - | - | 4 | - | - | - | - | - | - | - | 4 | 1 | - | - | - | 4 | 1 | - | - | - | - | 6 |
| Deuteraphorura cebennaria (Gisin, 1956) | stp | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 4 | 2 | 1 | 3 | - | - | - | 10 |
| Deutonura conjuncta (Stach, 1926) | tgx | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | 1 |
| Dicyrtoma fusca (Lubbock, 1873) | tgx | 1 | 1 | 1 | 4 | 2 | - | 6 | - | 1 | 1 | 17 | 4 | - | 3 | - | 3 | 5 | 5 | 2 | - | - | 22 |
| Dicyrtomina minuta (Fabricius, 1783) | stp | 5 | 10 | 4 | 8 | 6 | 3 | 13 | 3 | 2 | - | 54 | - | - | - | - | - | - | - | - | - | - | - |
| Entomobrya marginata (Tullberg, 1871) | tgx | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | 1 |
| Entomobrya nivalis (Linnaeus, 1758) | tgx | 10 | - | - | - | - | 1 | - | - | - | 3 | 14 | - | - | - | - | - | - | - | - | - | - | - |
| Entomobryidae juv. | 1 | - | - | - | - | - | - | - | - | - | 1 | 28 | 50 | 47 | 16 | 58 | 10 | 2 | 7 | 3 | 26 | 247 | |
| Folsomia fimetaria (Linné, 1758) | stp | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | 1 | 1 | - | - | - | - | - | 4 |
| Folsomia manolachei Bagnall, 1939 | tgx | 38 | 10 | 15 | 6 | 6 | 15 | 2 | 2 | 2 | 11 | 107 | 18 | 25 | 28 | 11 | 18 | 11 | 9 | 8 | 5 | 6 | 139 |
| Folsomia penicula Bagnall, 1939 | tgx | - | 7 | 1 | 2 | - | 5 | 4 | - | 1 | 2 | 22 | 6 | 8 | 4 | 3 | 1 | 1 | 5 | 1 | 6 | 4 | 39 |
| Folsomia quadrioculata (Tullberg, 1871) | tgx | 24 | 3 | 1 | 3 | 4 | - | - | - | - | - | 35 | 1 | 2 | 1 | 2 | 1 | 1 | - | 2 | 1 | - | 11 |
| Heteraphorura variotuberculata (Stach, 1934) | stp, p b | 1 | - | - | - | 1 | 11 | 10 | 13 | - | 1 | 37 | - | - | - | - | - | - | - | - | - | - | - |
| Heteromurus nitidus (Templeton, 1835) | etp | - | 1 | - | - | - | - | 2 | - | 1 | - | 4 | 11 | 13 | 15 | 4 | 19 | 13 | 15 | 8 | 3 | 7 | 108 |
| Isotomiella minor (Schäffer, 1896) | stp | 4 | 3 | 10 | - | 3 | 5 | 4 | 5 | 12 | - | 46 | 6 | 4 | 4 | 4 | 11 | 6 | 13 | 1 | 2 | - | 51 |
| Lepidocyrtus lignorum (Fabricius, 1775) | stp | 1220 | 1038 | 461 | 494 | 465 | 396 | 396 | 302 | 350 | 538 | 5660 | 431 | 643 | 629 | 484 | 549 | 498 | 377 | 263 | 279 | 154 | 4307 |
| Megalothorax incertus Börner, 1903 | etp | - | - | - | - | - | - | - | - | - | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Megalothorax minimus Willem, 1900 | etp | - | 3 | 7 | 8 | 6 | 15 | 13 | 20 | 13 | 18 | 103 | - | - | - | - | - | - | - | 1 | 5 | 4 | 10 |
| Megalothorax willemi Schneider et d’Haese, 2013 | etp | - | 8 | 4 | 2 | 4 | - | 2 | 7 | 8 | 8 | 43 | - | - | - | - | - | - | - | 3 | 5 | 1 | 9 |
| Morulina verrucosa (Börner, 1903) | tgx, E, p c,d | - | - | - | - | - | - | - | - | - | - | - | 2 | - | - | - | - | - | - | - | - | - | 2 |
| Neanura pseudoparva Rusek, 1963 | tgx | 4 | 5 | 4 | 12 | 10 | 11 | - | - | - | - | 46 | 2 | - | - | - | 1 | - | 1 | - | - | - | 4 |
| Oncopodura crassicornis Shoebotham, 1911 | etp | 3 | 3 | 7 | 5 | 2 | - | - | - | 2 | - | 22 | 11 | 10 | 10 | 4 | 45 | 14 | 41 | 21 | 26 | 11 | 193 |
| Onychiuroides pseudogranulosus (Gisin, 1951) | stp | 14 | 4 | 5 | - | - | 3 | 1 | - | 1 | - | 28 | 1 | 10 | 9 | - | 2 | 5 | 6 | 3 | 5 | 7 | 48 |
| Orchesella bifasciata Nicolet, 1842 | tgx | - | 1 | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Orchesella flavescens (Bourlet, 1843) | tgx | 34 | 12 | - | - | - | - | - | - | 11 | 5 | 62 | 8 | 2 | 1 | 4 | 4 | 2 | 4 | - | 1 | - | 26 |
| Parisotoma notabilis (Schäffer, 1896) | stp | 16 | 13 | 6 | 2 | 3 | 8 | 4 | 3 | 2 | - | 57 | 9 | 5 | 6 | - | 2 | 4 | 14 | 4 | 2 | - | 46 |
| Plutomurus carpaticus Rusek et Weiner 1978 | etp, E, p e | 118 | 122 | 116 | 82 | 96 | 126 | 113 | 213 | 188 | 273 | 1447 | 22 | 30 | 23 | 10 | 18 | 22 | 20 | 33 | 21 | 36 | 235 |
| Pogonognathellus flavescens (Tullberg, 1871) | stp | 200 | 132 | 174 | 92 | 60 | 62 | 82 | 75 | 81 | 177 | 1135 | 150 | 77 | 31 | 22 | 29 | 22 | 21 | 13 | 5 | 24 | 394 |
| Prosisotomodes bipunctatus (Axelson, 1903) | tgx | 2 | 1 | - | - | 3 | - | - | - | - | - | 6 | - | 4 | 3 | 3 | - | 1 | 1 | - | - | - | 12 |
| Protaphorura armata (Tullberg, 1869) | etp, p f | 18 | 36 | 30 | 33 | 34 | 133 | 90 | 82 | 139 | 91 | 686 | 4 | 2 | 4 | 3 | 4 | 34 | 2 | 8 | 8 | 4 | 73 |
| Protaphorura aurantiaca (Ridley, 1880) | stp | - | 1 | - | - | - | - | 3 | 3 | 1 | 6 | 14 | - | - | - | 2 | - | 1 | 2 | - | - | 1 | 6 |
| Protaphorura pannonica (Haybach, 1960) | tgx | - | - | - | - | - | - | 3 | - | - | 2 | 5 | - | - | - | - | - | - | - | - | - | - | - |
| Protaphorura tricampata (Gisin, 1956) | tgx | 2 | - | - | - | 1 | 5 | 3 | 4 | 2 | 9 | 26 | - | 1 | - | - | - | - | 1 | 3 | 3 | - | 8 |
| Pseudachorutes dubius Krausbauer, 1898 | tgx | 3 | - | - | - | - | - | - | - | - | - | 3 | - | - | - | - | - | - | - | - | - | - | - |
| Pseudosinella horaki Rusek, 1985 | stp | 84 | 209 | 62 | 49 | 39 | 76 | 52 | 42 | 66 | 89 | 768 | 11 | 24 | 11 | 9 | 10 | 2 | 4 | 4 | 6 | 7 | 88 |
| Pseudosinella thibaudi Stomp, 1977 | etp | - | 4 | 3 | - | - | - | - | 1 | 1 | - | 9 | 3 | 3 | 2 | 2 | 1 | 1 | - | - | 1 | - | 13 |
| Pygmarrhopalites principalis Stach, 1945 | etp, p g | - | 8 | 1 | - | 5 | 2 | 2 | 4 | 6 | 3 | 31 | - | - | - | - | - | 1 | - | 1 | - | 2 | 4 |
| Pygmarrhopalites pygmaeus (Wankel, 1860) | etp | 15 | 80 | 84 | 105 | 129 | 142 | 170 | 380 | 311 | 671 | 2087 | - | - | 5 | 4 | 11 | 6 | 4 | 10 | 13 | 15 | 68 |
| Sminthurinus aureus (Lubbock, 1862) | tgx | 5 | 4 | 2 | - | - | - | - | - | - | - | 11 | - | 1 | 1 | - | - | - | 1 | - | - | - | 3 |
| Tetrodontophora bielanensis (Waga, 1842) | stp, p h | 5 | 6 | 8 | 2 | 2 | - | 1 | 1 | 3 | - | 28 | 2 | 1 | - | 1 | 2 | - | 1 | - | - | - | 7 |
| Tomoceridae juv. | 1 | 4 | - | - | - | 2 | 2 | 1 | 1 | 4 | 15 | 3 | 1 | - | - | - | 1 | - | - | - | - | 5 | |
| Tomocerus minor (Lubbock, 1862) | stp, p e,i | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | 3 | 1 | - | - | - | - | 5 |
| Tomocerus vulgaris (Tullberg, 1871) | stp | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | 1 | 1 | - | 3 |
| Willowsia nigromaculata (Lubbock, 1873) | tgx | 5 | - | - | 1 | - | - | - | - | - | - | 6 | - | 2 | 1 | - | 2 | 2 | - | - | - | - | 7 |
| Ntotal | 1875 | 1853 | 1125 | 1044 | 1057 | 1150 | 1157 | 1440 | 1532 | 2448 | 14,681 | 762 | 957 | 891 | 613 | 864 | 705 | 579 | 432 | 464 | 382 | 6649 | |
| Stotal | 28 | 29 | 25 | 19 | 24 | 19 | 24 | 22 | 25 | 21 | 41 | 24 | 27 | 24 | 21 | 28 | 28 | 25 | 24 | 22 | 18 | 41 | |
Appendix B
| W | C | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Ech | 5 | 15 | 25 | 35 | 45 | 55 | 65 | 75 | 85 | 95 | Nw | 5 | 15 | 25 | 35 | 45 | 55 | 65 | 75 | 85 | 95 | Nc |
| Allacma fusca (Linnaeus, 1758) | tgx | 1 | 1 | - | - | - | - | - | - | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - |
| Ceratophysella denticulata (Bagnall, 1941) | etp | - | - | - | - | - | - | - | - | - | - | - | 5 | - | - | - | - | - | - | - | - | - | 5 |
| Desoria hiemalis (Schött, 1839) | stp | - | - | - | - | - | - | - | - | - | - | - | 5 | - | - | - | - | - | - | - | - | - | 5 |
| Desoria tigrina Nicolet, 1842 | stp | - | 2 | 1 | - | - | - | - | - | - | - | 3 | 2 | 4 | 1 | - | - | - | - | - | - | - | 7 |
| Deuteraphorura cebennaria (Gisin, 1956) | stp | - | - | - | - | - | - | - | - | - | - | - | 10 | 2 | - | 1 | 1 | - | - | - | - | - | 14 |
| Deutonura albella (Stach, 1920) | tgx, p h | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | 1 |
| Dicyrtoma fusca (Lubbock, 1873) | tgx | 16 | 33 | 13 | 13 | 5 | 1 | - | 1 | - | - | 82 | 42 | 40 | 24 | 20 | 10 | 5 | 7 | 3 | 1 | - | 152 |
| Dicyrtomina minuta (Fabricius, 1783) | stp | 7 | 4 | - | - | - | - | - | - | - | - | 11 | 115 | 65 | 24 | 3 | 1 | - | 2 | 2 | 1 | 3 | 216 |
| Entomobrya nivalis (Linnaeus, 1758) | tgx | - | - | - | - | - | 2 | - | - | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - |
| Entomobryidae juv. | - | - | - | 1 | - | - | - | 1 | - | - | 2 | 67 | 59 | 47 | 22 | 19 | 1 | 4 | 5 | 5 | 3 | 232 | |
| Folsomia manolachei Bagnall, 1939 | tgx | 13 | 2 | - | - | 1 | - | - | - | - | - | 16 | 14 | 8 | 4 | 5 | 11 | 5 | 6 | - | - | - | 53 |
| Folsomia penicula Bagnall, 1939 | tgx | - | - | - | - | - | - | - | - | - | 1 | 1 | 4 | 3 | 4 | 1 | 5 | - | - | - | - | - | 17 |
| Folsomia quadrioculata (Tullberg, 1871) | tgx | - | - | - | - | - | - | - | - | - | - | - | 1 | 1 | - | - | 2 | - | - | 1 | - | - | 5 |
| Heteraphorura variotuberculata (Stach, 1934) | stp, p b | - | - | - | - | - | - | - | - | - | - | - | 1 | - | 4 | - | 2 | 5 | 4 | 1 | - | 3 | 20 |
| Heteromurus nitidus (Templeton, 1835) | etp | - | - | - | - | - | - | - | - | - | - | - | 16 | 15 | 9 | 16 | 10 | 9 | 17 | 4 | 17 | 8 | 121 |
| Isotomiella minor (Schäffer, 1896) | stp | 5 | - | 3 | 3 | - | 1 | - | - | - | - | 12 | 8 | 6 | 6 | 6 | 6 | 6 | 11 | 6 | 5 | 8 | 68 |
| Kalaphorura carpenteri (Stach, 1919) | etp, p b | 21 | 10 | 2 | 4 | 2 | 1 | 3 | 1 | - | 2 | 46 | 8 | 12 | 1 | 8 | 7 | 7 | 17 | 12 | 18 | 33 | 123 |
| Kalaphorura paradoxa (Schäffer, 1900) | etp, p b | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | 1 |
| Lepidocyrtus lignorum (Fabricius, 1775) | stp | 92 | 87 | 39 | 35 | 39 | 38 | 27 | 29 | 13 | 24 | 423 | 370 | 210 | 180 | 182 | 158 | 145 | 169 | 97 | 121 | 108 | 1740 |
| Lepidocyrtus serbicus Denis, 1936 | tgx | 3 | 2 | 3 | - | 2 | - | - | - | - | - | 10 | - | - | - | - | - | - | - | - | - | - | - |
| Megalothorax minimus Willem, 1900 | etp | - | - | - | - | - | - | - | - | - | 1 | 1 | 1 | 5 | 1 | 11 | - | 30 | 14 | 17 | 18 | 3 | 100 |
| Megalothorax willemi Schneider et d’Haese, 2013 | etp | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | 6 | 4 | - | 4 | 2 | 17 |
| Neanura pseudoparva Rusek, 1963 | tgx | - | - | - | - | - | - | - | - | - | - | - | 1 | 4 | - | - | - | - | - | - | - | - | 5 |
| Neelus koseli Kováč et Papáč, 2010 | tgb, E | - | - | 12 | 14 | 16 | 13 | 25 | 15 | 16 | 17 | 128 | - | 4 | 4 | 13 | 4 | 26 | 34 | 10 | 49 | 26 | 170 |
| Oncopodura crassicornis Shoebotham, 1911 | etp | - | - | - | - | - | 1 | 3 | 2 | - | - | 6 | 13 | 15 | 20 | 27 | 12 | 5 | 14 | - | 12 | 11 | 129 |
| Oncopodura reyersdorfensis Stach, 1936 | etp | - | - | - | - | - | - | - | - | - | - | - | 1 | - | 1 | 1 | 1 | - | - | - | - | - | 4 |
| Onychiuroides pseudogranulosus (Gisin, 1951) | stp | 7 | 9 | - | - | - | - | - | - | - | - | 16 | 8 | 11 | - | 3 | - | - | - | - | - | - | 22 |
| Orchesella bifasciata Nicolet, 1842 | tgx | - | 1 | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Orchesella flavescens (Bourlet, 1843) | tgx | 27 | 19 | 3 | - | - | - | - | - | - | - | 49 | 8 | - | 1 | - | - | - | - | - | - | - | 9 |
| Parisotoma notabilis (Schäffer, 1896) | stp | - | - | - | 2 | - | - | - | - | - | - | 2 | 5 | 5 | 4 | 2 | 2 | 1 | 5 | 4 | 3 | 3 | 34 |
| Plutomurus carpaticus Rusek et Weiner 1978 | etp, E, p e | 53 | 58 | 55 | 68 | 47 | 75 | 65 | 191 | 152 | 181 | 945 | 87 | 75 | 95 | 136 | 97 | 107 | 191 | 132 | 126 | 140 | 1186 |
| Pogonognathellus flavescens (Tullberg, 1871) | stp | 288 | 193 | 207 | 158 | 118 | 61 | 30 | 32 | 8 | 10 | 1105 | 171 | 119 | 92 | 41 | 29 | 6 | - | - | 6 | 7 | 471 |
| Prosisotomodes bipunctatus (Axelson, 1903) | tgx | 2 | - | - | - | - | - | - | - | - | - | 2 | 5 | 3 | 1 | - | 1 | - | - | - | - | - | 10 |
| Protaphorura armata (Tullberg, 1869) | etp, p f | 4 | 16 | 7 | 16 | 1 | 5 | - | 3 | 12 | 3 | 67 | - | 2 | 3 | 2 | 8 | 7 | 10 | 7 | 15 | 19 | 73 |
| Protaphorura aurantiaca (Ridley, 1880) | stp | - | 3 | - | - | - | - | - | - | - | - | 3 | - | - | - | - | - | 3 | 1 | 1 | 2 | 2 | 9 |
| Protaphorura campata (Gisin, 1952) | tgx | - | - | - | - | - | - | - | - | 6 | - | 6 | - | - | - | - | - | - | - | - | - | - | - |
| Protaphorura tricampata (Gisin, 1956) | tgx | - | - | 2 | 4 | - | 1 | - | 2 | 4 | 4 | 17 | - | - | - | 2 | 2 | 2 | 2 | 1 | 5 | 2 | 16 |
| Pseudosinella horaki Rusek, 1985 | stp | 21 | 30 | 19 | 33 | 132 | 42 | 23 | 56 | 69 | 49 | 474 | 25 | 26 | 11 | 27 | 11 | 24 | 25 | 22 | 24 | 15 | 210 |
| Pseudosinella thibaudi Stomp, 1977 | etp | - | - | - | - | - | - | - | - | - | - | - | 2 | - | 1 | 2 | 2 | - | - | - | - | - | 7 |
| Pygmarrhopalites principalis Stach, 1945 | etp, p g | 6 | 80 | 130 | 194 | 165 | 126 | 76 | 69 | 86 | 69 | 1001 | - | 1 | 8 | 18 | 12 | 7 | 19 | 7 | 2 | 4 | 78 |
| Pygmarrhopalites pygmaeus (Wankel, 1860) | etp | 4 | 3 | 23 | 17 | 27 | 23 | 22 | 24 | 25 | 22 | 190 | 13 | 20 | 100 | 125 | 59 | 85 | 96 | 97 | 113 | 98 | 806 |
| Sminthurinus aureus (Lubbock, 1862) | tgx | - | - | - | 1 | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Tetrodontophora bielanensis (Waga, 1842) | stp, p h | 7 | 5 | 3 | - | 2 | 1 | - | - | - | - | 18 | 15 | 4 | 2 | - | 1 | - | - | - | - | - | 22 |
| Tomoceridae juv. | - | 3 | - | - | - | - | - | - | - | - | 3 | - | - | - | 1 | - | - | 5 | - | - | - | 6 | |
| Tomocerina minuta (Tullberg, 1877) | stp | - | - | 1 | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| Tomocerus vulgaris (Tullberg, 1871) | stp | - | - | - | - | - | - | - | - | 1 | 2 | 3 | - | 3 | - | - | 4 | - | 3 | 1 | 1 | - | 12 |
| Ntotal | 577 | 561 | 523 | 563 | 557 | 391 | 274 | 426 | 392 | 385 | 4649 | 1023 | 724 | 648 | 675 | 478 | 492 | 660 | 430 | 548 | 498 | 6176 | |
| Stotal | 18 | 19 | 17 | 14 | 13 | 15 | 9 | 12 | 11 | 13 | 32 | 28 | 28 | 25 | 23 | 27 | 20 | 21 | 19 | 20 | 19 | 37 | |
References
- Juberthie, C.; Delay, B.; Bouillon, M. Extension du milieu souterrain en zone non calcaire:description d’un nouveau milieu et de son peuplement par les Coléoptéres troglobies. Mem. Biospeol. 1980, 7, 19–52. [Google Scholar]
- Juberthie, C.; Delay, B.; Bouillon, M. Extension du milieu souterrain superficiel en zone calcaire. Mem. Biospeol. 1981, 8, 77–93. [Google Scholar]
- Culver, D.C.; Pipan, T. Shallow Subterranean Habitats: Ecology, Evolution, and Conservation; Oxford University Press: Oxford, MS, USA, 2014. [Google Scholar]
- Mammola, S.; Giachino, P.M.; Piano, E.; Jones, A.; Barberis, M.; Badino, G.; Isaia, M. Ecology and sampling techniques of an understudied subterranean habitat: The milieu Souterrain Superficiel (MSS). Sci. Nat. 2016, 103, 88. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. Superficial subterranean habitats–gateway to the subterranean realm. Cave Karst Sci. 2008, 35, 5–12. [Google Scholar]
- Růžička, V.; Zacharda, M. Arthropods of stony debris in the Krkonoše Mountains, Czech Republic. Arct. Alp. Res. 1994, 26, 332–338. [Google Scholar] [CrossRef]
- Delaloye, R.; Lambiel, C. Evidence of winter ascending air circulation throughout talus slopes and rock glaciers situated in the lower belt of alpine discontinuous permafrost (Swiss Alps). Nor. Geogr. Tidsskr. 2005, 59, 194–203. [Google Scholar] [CrossRef]
- Gutiérrez, M. Chapter 7. The periglacial environment. Dev. Earth Surf. Process. 2005, 8, 145–170. [Google Scholar]
- Ledesma, E.; Jiménez-Valverde, A.; Baquero, E.; Jordana, R.; de Castro, A.; Ortuño, V.M. Arthropod biodiversity patterns point to the Mesovoid Shallow Substratum (MSS) as a climate refugium. Zoology 2020, 141, 125771. [Google Scholar] [CrossRef]
- Bárány-Kevei, I. Microclimate of karstic dolines. Acta Climatol. 1999, 32, 19–27. [Google Scholar]
- Růžička, V. The freezing scree slopes and their arachnofauna. In Lebensraum Blockhalde—Zur Ökologie Periglazialer Blockhalden im Ausseralpinen Mitteleuropa. Decheniana-Beihefte; Möseler, B.M., Molenda, R., Eds.; Naturhistorischer Verein der Rheinlande und Westfalens: Bonn, Germany, 1999; Volume 37, pp. 141–147. [Google Scholar]
- Gude, M.; Dietrich, S.; Mausbacher, R.; Hauck, C.; Molenda, R.; Růžička, V.; Zacharda, M. Probable occurrence of sporadic permafrost in non-alpine scree slopes in central Europe. In Proceedings of the 8th International Conference on Permafrost, Zürich, Switzerland, 21–25 July 2003; Phillips, M., Springman, S.M., Arenson, L.U., Eds.; Balkema: Zürich, Switzerland; pp. 331–336.
- Zacharda, M.; Gude, M.; Kraus, S.; Hauck, C.; Molenda, R.; Růžička, V. The relict mite Rhagidia gelida (Acari, Rhagidiidae) as a biological cryoindicator of periglacial microclimate in European highland screes. Arct. Antarct Alp. Res. 2005, 37, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Zacharda, M.; Gude, M.; Růžička, V. Thermal regime of three low elevation scree slope in central Europe. Permafr. Periglac. Process. 2007, 18, 301–308. [Google Scholar] [CrossRef]
- Růžička, V.; Zacharda, M.; Němcová, L.; Šmilauer, P.; Nekola, J.C. Periglacial microclimate in low-altitude scree slopes supports relict biodiversity. J. Nat. Hist. 2012, 46, 2145–2157. [Google Scholar] [CrossRef]
- Růžička, V.; Zacharda, M.; Šmilauer, P.; Kučera, T. Can paleorefugia of cold-adapted species in talus slopes resist global warming? Boreal. Environ. Res. 2015, 20, 403–412. [Google Scholar]
- Šupinský, J.; Kaňuk, J.; Hochmuth, Z.; Gallay, M. Detecting dynamics of cave floor ice with selective cloud-to-cloud approach. Cryosphere 2019, 13, 2835–2851. [Google Scholar] [CrossRef] [Green Version]
- Růžička, V.; Hajer, J.; Zacharda, M. Arachnid population patterns in underground cavities of a stony debris field (Araneae, Opiliones, Pseudoscorpionidea, Acari: Prostigmata, Rhagidiidae). Pedobiologia 1995, 39, 42–51. [Google Scholar]
- Raška, P.; Kirchner, K.; Raška, M. Winter microclimatic regime of low-altitude scree slopes and its relation to topography: Case study from the Ceske Stredohori Mts. (N Czech Republic). Geogr. Fis. Din. Quat. 2011, 34, 235–246. [Google Scholar] [CrossRef]
- Delaloye, R.; Reynard, E.; Lambiel, C.; Marescot, L.; Monnet, R. Thermal anomaly in a cold scree slope (Creux du Van, Switzerland). In Proceedings of the 8th International Conference on Permafrost, Zürich, Switzerland, 21–25 July 2003; Phillips, M., Springman, S.M., Arenson, L.U., Eds.; Balkema: Zürich, Switzerland; pp. 175–180.
- Zacharda, M.; Kučera, T. Diversity of predatory Rhagidiid mites (Acari: Rhagidiidae) inhabiting montane stony debris in the Otztal Alps, North Tyrol. Arct. Antarct. Alp. Res. 2006, 38, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Meynier, S.; Brun, J.J. Humus forms pathways in low-elevation cold scree slopes: Tangel or Mor? Appl. Soil. Ecol. 2018, 123, 572–580. [Google Scholar] [CrossRef]
- Christian, E. Composition and origin of underground arthropod fauna in an extrazonal permafrost soil of Central Europe. Biol. Fertil. Soils 1987, 3, 27–30. [Google Scholar] [CrossRef]
- Nitzu, E.; Dorobăţ, M.L.; Popa, I.; Giurginca, A.; Baba, Ş. The influence of geological substrate on the faunal structure of the superficial subterranean habitats. Carpathian J. Earth Environ. Sci. 2018, 13, 383–393. [Google Scholar] [CrossRef]
- Nitzu, E.; Nae, A.; Giurginca, A.; Popa, I. Invertebrate communities from the mesovoid shallow substratum of the carpatho-euxinic area: Eco-faunistic and zoogeographic analysis. Trav. Inst. Spéol. Émile Racovitza 2010, 49, 41–79. [Google Scholar]
- Nitzu, E.; Popa, I.; Giurginca, A. Invertebrate fauna (Coleoptera, Collembola, Diplopoda, Isopoda) collected in the karst areas of the Aninei-Locvei Mountains. Trav. Inst. Spéol. Émile Racovitza 2011, 50, 15–35. [Google Scholar]
- Nitzu, E.; Nae, A.; Băncilă, R.; Popa, I.; Giurginca, A.; Plăiaşu, R. Scree habitats: Ecological function, species conservation and spatial temporal variation in the arthropod community. Syst. Biodivers. 2014, 12, 65–75. [Google Scholar] [CrossRef]
- Pipan, T.; López, H.; Oromí, P.; Polak, S.; Culver, D.C. Temperature variation and the presence of troglobionts in terrestrial shallow subterranean habitats. J. Nat. Hist. 2011, 45, 253–273. [Google Scholar] [CrossRef]
- Rendoš, M.; Mock, A.; Jászay, T. Spatial and temporal dynamics of invertebrates dwelling karstic mesovoid shallow substratum of Sivec National Nature Reserve (Slovakia), with emphasis on Coleoptera. Biologia 2012, 67, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Rendoš, M.; Raschmanová, N.; Kováč, Ľ.; Miklisová, D.; Mock, A.; Ľuptáčik, P. Organic carbon content and temperature as substantial factors affecting diversity and vertical distribution of Collembola on forested scree slopes. Eur. J. Soil Biol. 2016, 75, 180–187. [Google Scholar] [CrossRef]
- Rendoš, M.; Miklisová, D.; Kováč, Ľ.; Mock, A. Dynamics of Collembola (Hexapoda) in a forested limestone scree slope, Western Carpathians, Slovakia. J. Cave Karst Stud. 2020, 82, 18–29. [Google Scholar] [CrossRef]
- Raschmanová, N.; Miklisová, D.; Kováč, L. Dynamics of soil Collembola communities (Hexapoda: Collembola) along the mesoclimatic gradient in a deep karst valley. Biologia 2016, 71, 184–193. [Google Scholar] [CrossRef]
- Marcin, M.; Raschmanová, N.; Miklisová, D.; Kováč, L. Microclimate and habitat heterogeneity as important drivers of soil Collembola in a karst collapse doline in the temperate zone. Invertebr. Biol. 2021, e12315. [Google Scholar] [CrossRef]
- Gers, C. Diversity of energy fluxes and interactions between arthropod communities: Fromsoil to cave. Acta Oecol. 1998, 19, 205–213. [Google Scholar] [CrossRef]
- Jureková, N.; Raschmanová, N.; Kováč, Ľ.; Miklisová, D.; Červená, M.; Christophoryová, J. Type of fixative solution in pitfall traps as a decisive factor affecting community parameters of Collembola (Hexapoda) inhabiting superficial subterranean habitats. Sci. Nat. 2019, 106, 21. [Google Scholar] [CrossRef]
- Norris, C.; Hobson, P.; Ibisch, P.L. Microhabitat and vegetation function as indicators of forest thermodynamic efficiency. J. Appl. Ecol. 2012, 49, 562–570. [Google Scholar] [CrossRef]
- Ashcroft, M.B.; Gollan, J.R. Moisture, thermal inertia, and the spatial distributions of near-surface soil and air temperatures: Understanding factors that promote microrefugia. Agric. For. Meteorol. 2013, 176, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Keppel, G.; Van Niel, K.P.; Wardell-Johnson, G.W.; Yates, C.J.; Byrne, M.; Mucina, L.; Schut, A.G.T.; Hopper, S.D.; Franklin, S.E. Refugia: Identifying and understanding safe havens for biodiversity under climate change. Glob. Ecol. Biogeogr. 2012, 21, 393–404. [Google Scholar] [CrossRef]
- Raschmanová, N.; Miklisová, D.; Kováč, Ľ.; Šustr, V. Community composition and cold tolerance of soil Collembola in a collapse karst doline with strong microclimate inversion. Biologia 2015, 70, 802–811. [Google Scholar] [CrossRef]
- Bátori, Z.; Vojtkó, A.; Maák, I.E.; Lőrinczi, G.; Farkas, T.; Kántor, N.; Tanács, E.; Kiss, P.J.; Juhász, O.; Módra, G.; et al. Karst dolines provide diverse microhabitats for different functional groups in multiple phyla. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Raschmanová, N.; Kováč, Ľ.; Miklisová, D. The effect of mesoclimate on Collembola diversity in the Zádiel Valley, Slovak karst (Slovakia). Eur. J. Soil Biol. 2008, 44, 463–472. [Google Scholar] [CrossRef]
- Raschmanová, N.; Miklisová, D.; Kováč, Ľ. A unique small-scale microclimatic gradient in a temperate karst harbours exceptionally high diversity of soil Collembola. Int. J. Speleol. 2018, 47, 247–262. [Google Scholar] [CrossRef]
- Mammola, S.; Piano, E.; Cardoso, P.; Vernon, P.; Domínguez-Villar, D.; Culver, D.C.; Pipan, T.; Isaia, M. Climate change going deep: The effects of global climatic alterations on cave ecosystems. Anthr. Rev. 2019, 6, 98–116. [Google Scholar] [CrossRef]
- Raschmanová, N.; Miklisová, D.; Kováč, Ľ. Soil Collembola communities along a steep microclimatic gradient in the collapse doline of the Silická ľadnica Cave, Slovak Karst (Slovakia). Biologia 2013, 68, 470–478. [Google Scholar] [CrossRef]
- Mammola, S.; Leroy, B. Applying species distribution models to caves and other subterranean habitats. Ecography 2018, 41, 1194–1208. [Google Scholar] [CrossRef] [Green Version]
- Ledesma, E.; Jiménez-Valverde, A.; de Castro, A.; Aguado-Aranda, P.; Ortuño, V.M. The study of hidden habitats sheds light on poorly known taxa: Spiders of the Mesovoid Shallow Substratum. ZooKeys 2019, 841, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Rozložník, M.; Szöllös, F.; Uhrin, M.; Karasová, E. Slovenský kras–Slovak Karst Biosphere Reserve. In Biosphere Reserves on the Crossroads of Central Europe; Jeník, J., Price, F., Eds.; Czech National Committee for UNESCO’s MaB Programme: Empora, Prague, Czech Republic, 1994; pp. 113–128. [Google Scholar]
- Hofierka, J.; Gallay, M.; Bandura, P.; Šašak, J. Identification of karst sinkholes in a forested karst landscape using airborne lasser scanning data and water flow analysis. Geomorphology 2018, 308, 265–277. [Google Scholar] [CrossRef]
- Ľuptáčik, P.; Tajovský, K.; Pižl, V. Symphyla, Pauropoda, Protura and Diplura Dynamics Along a Climate Inversion in the Zádiel Gorge (Slovak Karst). Soil Zoology in Central Europe. In Proceedings of the 5th Central European Workshop on Soil Zoology, České Budejovice, Czech Republic, 27–30 April 1999; pp. 197–201. [Google Scholar]
- Mock, A.; Šašková, T.; Raschmanová, N.; Jászay, T.; Ľuptáčik, P.; Rendoš, M.; Tajovský, K.; Jászayová, A. An introductory study of subterranean communities of invertebrates in forested talus habitats in southern Slovakia. Acta Soc. Zool. Bohem. 2015, 79, 243–256. [Google Scholar]
- Schlick-Steiner, B.C.; Steiner, F.M. Eine neue Subterranfalle und Fänge aus Kärnten. Carinthia II 2000, 190/110, 475–482. [Google Scholar]
- Rusek, J. Eine Präparationstechnik für Springschwänze und ähnliche Gliederfüsser. Mikrokosmos 1975, 12, 376–381. [Google Scholar]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark. Part I: Poduromorpha. Fauna Entomologica Scandinavica; Brill: Leiden, The Netherlands, 1998; Volume 35, pp. 1–183. [Google Scholar]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark. Part II: Entomobryomorpha and Symphypleona. Fauna Entomologica Scandinavica; Brill: Leiden, The Netherlands, 2007; Volume 42, pp. 1–263. [Google Scholar]
- Pomorski, R.J. Onychiurinae of Poland (Collembola: Onychiuridae): Genus; Polish Taxonomical Society: Wrocław, Poland, 1998; pp. 1–201. [Google Scholar]
- Bretfeld, G. Synopses on Palaearctic Collembola: Symphypleona. Abh. Ber. Nat. Görlitz 1999, 71, 1–318. [Google Scholar]
- Potapov, M. Synopses on Palaearctic Collembola: Isotomidae. Abh. Ber. Nat. Görlitz 2001, 73, 1–603. [Google Scholar]
- Thibaud, J.M.; Schulz, H.J.; da Gama Assalino, M.M. Synopses on Palaearctic Collembola: Hypogastruridae. Abh. Ber. Nat. Görlitz 2004, 75, 1–287. [Google Scholar]
- StatSoft Inc. STATISTICA (Data Analysis Software System), Version 12. 2013. Available online: https://www.statsoft.com/ (accessed on 7 December 2020).
- Kobza, J.; Hrivňáková, K.; Makovníková, J.; Barančíková, G.; Bezák, P.; Bezáková, Z.; Dodok, R.; Grečo, V.; Chlpík, J.; Lištjak, M.; et al. Jednotné Pracovné Postupy Rozborov Pôd; Výskumný ústav pôdoznalectva a ochrany pôdy: Bratislava, Slovakia, 2011; pp. 1–136. [Google Scholar]
- Tischler, W. Synökologie der Landtiere; Gustav Fischer, Verlag: Stuttgart, Germany, 1955; pp. 1–414. [Google Scholar]
- Sket, B. Can we agree on an ecological classification of subterranean animals? J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Kováč, L.; Parimuchová, A.; Miklisová, D. Distributional patterns of cave Collembola (Hexapoda) in association with habitat conditions, geography and subterranean refugia in the Western Carpathians. Biol. J. Linn. Soc. 2016, 119, 571–592. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data; Version 6.07; MjM Software: Gleneden Beach, OR, USA, 2011. [Google Scholar]
- Shanks, R.E.; Norris, F.H. Microclimatic variation in a small valley in eastern Tennessee. Ecology 1950, 31, 532–539. [Google Scholar] [CrossRef]
- Pipan, T.; Culver, D.C. Organic carbon in shallow subterranean habitats. Acta Carsologica 2013, 42, 91–300. [Google Scholar] [CrossRef] [Green Version]
- Swift, M.J.; Heal, O.W.; Anderson, J.M. Decomposition in Terrestrial Ecosystems; University of California Press: Berkeley, CA, USA, 1979. [Google Scholar]
- Laška, V.; Kopecký, O.; Růžička, V.; Mikula, J.; Véle, A.; Šarapatka, B.; Tuf, I.H. Vertical distribution of spiders in soil. J. Arachnol. 2011, 39, 393–398. [Google Scholar] [CrossRef]
- Nitzu, E.; Nae, A.; Popa, I. Eco-faunistic study on the invertebrate fauna (Araneae, Collembola and Coleoptera) from the Vârghis Gorge Natural Reserve (Eastern Carpathians, Romania), with special note on the micro-refugial role of the subterranean habitats. Trav. Inst. Spéol. Émile Racovitza 2006, 45, 31–50. [Google Scholar]
- Dorobăţ, M.L.; Nitzu, E.; Popa, I.; Giurginca, A.; Nae, A.; Baba, Ş.; Dobrescu, C.M. A systematic conspectus of the invertebrate species identified in the scree and lithosol areas from the North-Western sector of the Leaota Mountains (Southern Carpathians), Romania. Stud. Cercet. Ştiinţifice Biol. 2019, 28, 60–65. [Google Scholar]
- Rendoš, M.; Červená, M.; Haľková, B.; Jakšová, P.; Jureková, N.; Rudy, J.; Jászay, T.; Raschmanová, N.; Mock, A.; Kováč, Ľ.; et al. Forested scree slopes—A noteworthy subterranean habitat of the Western Carpathians. Acta Carsologica Slovaca 2019, 57, 229–245. [Google Scholar]
- Kemencei, Z.; Farkas, R.; Páll-Gergely, B.; Vilisics, F.; Nagy, A.; Hornung, E.; Sólymos, P. Microhabitat associations of land snails in forested dolinas: Implications for coarse filter conservation. Community Ecol. 2014, 15, 180–186. [Google Scholar] [CrossRef]
- Růžička, V. Central European habitats inhabited by spiders with disjunctive distributions. Pol. J. Ecol. 2011, 59, 367–380. [Google Scholar]
- Zettel, J.; Zettel, U. Development, phenology and surface activity of Ceratophysella sigillata (Uzel) (Collembola, Hypogastruridae). Acta Zool. Fenn. 1994, 195, 150–153. [Google Scholar]
- Zettel, J.; Zettel, U. Seasonal and reproductional polymorphism in Ceratophysella sigillata (Uzel) (Collembola, Hypogastruridae). Acta Zool. Fenn. 1994, 195, 154–156. [Google Scholar]
- Block, W.; Zettel, J. Activity and dormancy in relation to body water and cold tolerance in a winter-active springtail (Collembola). Eur. J. Entomol. 2003, 100, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Zacharda, M. Glacial relict Rhagidiidae (Acari: Prostigmata) from superficial underground enclosures in the Krkonoše Mountains, Czechoslovakia. J. Nat. Hist. 1993, 27, 47–61. [Google Scholar] [CrossRef]
- Růžička, V.; Zacharda, M. Variation and diversity of spider assemblages along a thermal gradient in scree slopes and adjacent cliffs. Pol. J. Ecol. 2010, 8, 361–369. [Google Scholar]
- Urbanovičová, V.; Kováč, Ľ.; Miklisová, D. Epigeic arthropod communities of spruce forest stands in the High Tatra Mts.(Slovakia) with special reference to Collembola–first year after windthrow. Acta Soc. Zool. Bohem. 2010, 74, 141–152. [Google Scholar]
- Weiner, W.M. Collembola of the Pieniny National Park in Poland. Acta Zool. Cracov. 1981, 25, 417–500. [Google Scholar]
- Kováč, Ľ.; Papáč, V. Revision of the genus Neelus Folsom, 1896 (Collembola, Neelida) with the description of two new troglobiotic species from Europe. Zootaxa 2010, 2663, 36–52. [Google Scholar] [CrossRef] [Green Version]
- Nosek, J. The investigation of the Apterygotan fauna of the Low Tatras. Acta Univ. Carol. Biol. 1967, 5–6, 349–528. [Google Scholar]
- Urbanovičová, V.; Miklisová, D.; Kováč, Ľ. Forest disturbance enhanced the activity of epedaphic collembola in windthrown stands of the High Tatra mountains. J. Mt. Sci. 2014, 11, 449–463. [Google Scholar] [CrossRef]
- Smolis, A.; Skarzyński, D. Springtails (Collembola) of the “Przelom Jasiolki” reserve in the Beskid Niski Mountains (Polish Carpathians). Fragm. Faunist. 2003, 46, 121–129. [Google Scholar] [CrossRef]





| Site | SA | SB | SA | SB | |
|---|---|---|---|---|---|
| TW [°C] | W [%] | ||||
| 5 cm | 8.9 ± 1.9 a | 16.5 ± 3.1 b | 5 cm | 75.6 ± 2.9 | 71.1 ± 13.0 |
| 35 cm | 6.2 ± 2.2 a | 16.7 ± 2.3 b | 35 cm | 53.8 ± 4.3 | 68.4 ± 3.4 |
| 65 cm | 5.8 ± 2.2 a | 16.7 ± 2.1 b | 65 cm | 47.1 ± 2.8 | 65.4 ± 8.5 |
| 95 cm | 5.4 ± 2.1 a | 16.6 ± 2.1 b | 95 cm | 54.2 ± 10.3 | 21.1 ± 3.1 |
| TC [°C] | pHH2O | ||||
| 5 cm | 1.1 ± 4.3 a | 6.4 ± 3.7 b | 5 cm | 6.9 | 7.2 |
| 35 cm | 0.7 ± 3.8 a | 8.5 ± 3.2 b | 35 cm | 7.7 | 7.3 |
| 65 cm | 0.7 ± 3.7 a | 8.7 ± 3.1 b | 65 cm | 7.7 | 7.7 |
| 95 cm | 0.6 ± 3.6 a | 8.7 ± 3.1 b | 95 cm | 7.6 | 7.0 |
| Corg [%] | |||||
| TW min [°C] | −0.1 | 9.1 | 5 cm | 52.5 | 47.0 |
| TW max [°C] | 12.0 | 23.0 | 35 cm | 31.3 | 46.0 |
| TC min [°C] | −8.4 | −0.5 | 65 cm | 28.6 | 24.3 |
| TC max [°C] | 9.9 | 16.0 | 95 cm | 29.4 | 13.8 |
| Parameter | Pearson Coefficient | |
|---|---|---|
| S_Collembola | TC | −0.56 |
| TC min | −0.79 | |
| Corg | 0.77 | |
| N_Collembola | TW | −0.74 |
| TC | −0.81 | |
| TW min | −0.77 | |
| TW max | −0.71 | |
| TC min | −0.61 | |
| TC max | −0.77 | |
| S_trogloxenes | TC | −0.51 |
| TW min | −0.52 | |
| Corg | 0.61 | |
| N_trogloxenes | pHH2O | −0.65 |
| Corg | 0.79 | |
| S_subtroglophiles | TW | 0.82 |
| TC | 0.77 | |
| TW min | 0.75 | |
| TW max | 0.86 | |
| TC min | 0.71 | |
| TC max | 0.81 | |
| N_subtroglophiles | TW | −0.56 |
| TC | −0.72 | |
| TC max | −0.63 | |
| TW min | −0.63 | |
| TW max | −0.51 | |
| TC min | −0.86 | |
| Corg | 0.69 | |
| Lepidocyrtus lignorum | TW | −0.69 |
| TC | −0.81 | |
| TC max | −0.74 | |
| TW min | −0.71 | |
| TW max | −0.67 | |
| TC min | −0.91 | |
| Corg | 0.51 | |
| Plutomurus carpaticus | TC min | 0.78 |
| Corg | −0.58 | |
| Pogonognathellus flavescens | Corg | 0.86 |
| Pygmarrhopalites pygmaeus | TW | −0.50 |
| TW max | −0.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jureková, N.; Raschmanová, N.; Miklisová, D.; Kováč, Ľ. Mesofauna at the Soil-Scree Interface in a Deep Karst Environment. Diversity 2021, 13, 242. https://doi.org/10.3390/d13060242
Jureková N, Raschmanová N, Miklisová D, Kováč Ľ. Mesofauna at the Soil-Scree Interface in a Deep Karst Environment. Diversity. 2021; 13(6):242. https://doi.org/10.3390/d13060242
Chicago/Turabian StyleJureková, Nikola, Natália Raschmanová, Dana Miklisová, and Ľubomír Kováč. 2021. "Mesofauna at the Soil-Scree Interface in a Deep Karst Environment" Diversity 13, no. 6: 242. https://doi.org/10.3390/d13060242
APA StyleJureková, N., Raschmanová, N., Miklisová, D., & Kováč, Ľ. (2021). Mesofauna at the Soil-Scree Interface in a Deep Karst Environment. Diversity, 13(6), 242. https://doi.org/10.3390/d13060242