
Diversity of Testate Amoebae as an Indicator of the Conservation Status of Peatlands in Southwest Europe



Abstract
:1. Introduction
2. Materials and Methods
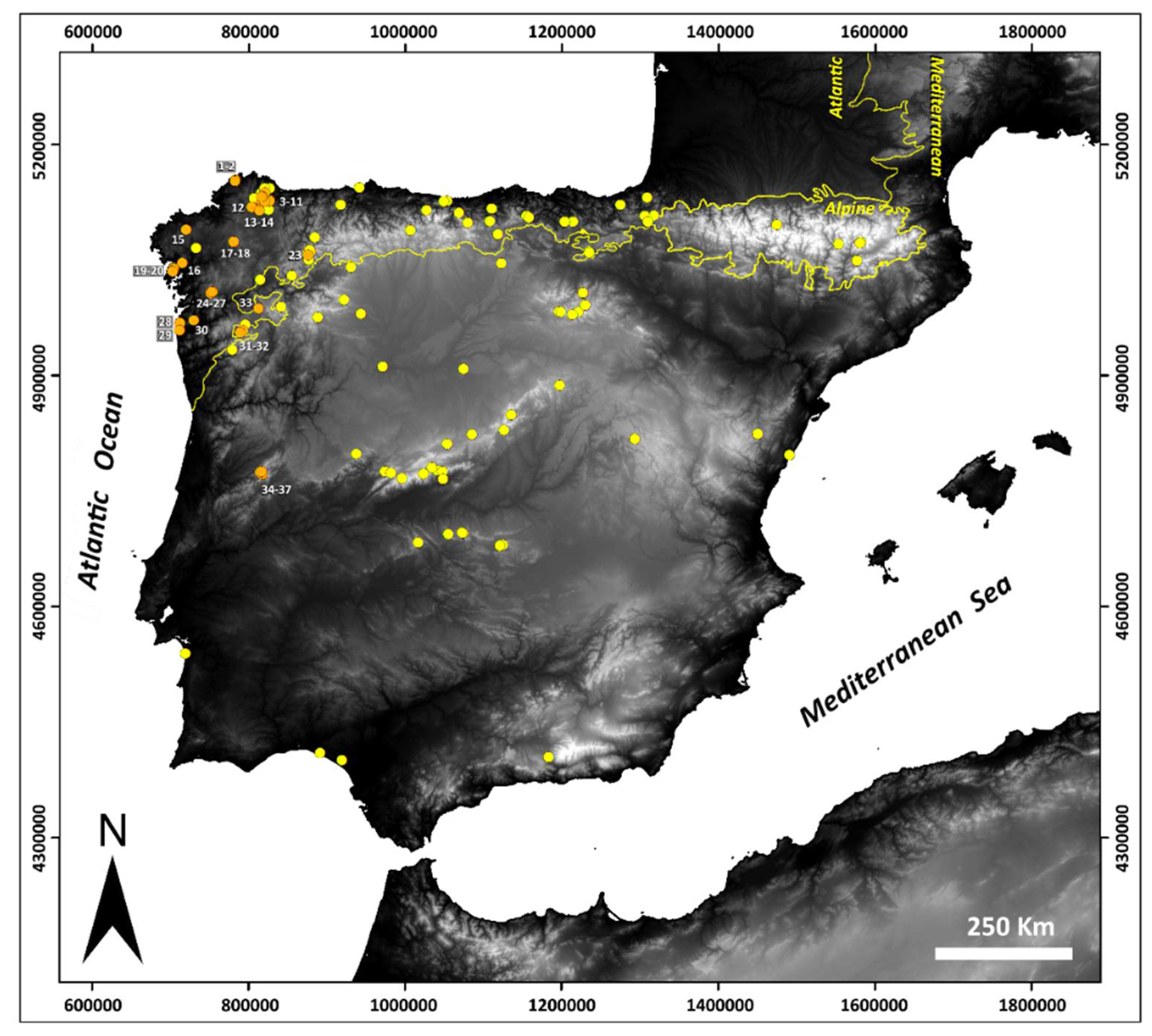
2.1. Study Site
2.2. Sampling and Testate Amoeba Extraction
2.3. Water Chemistry Analysis
2.4. Statistical Analysis
3. Results
3.1. Water Quality
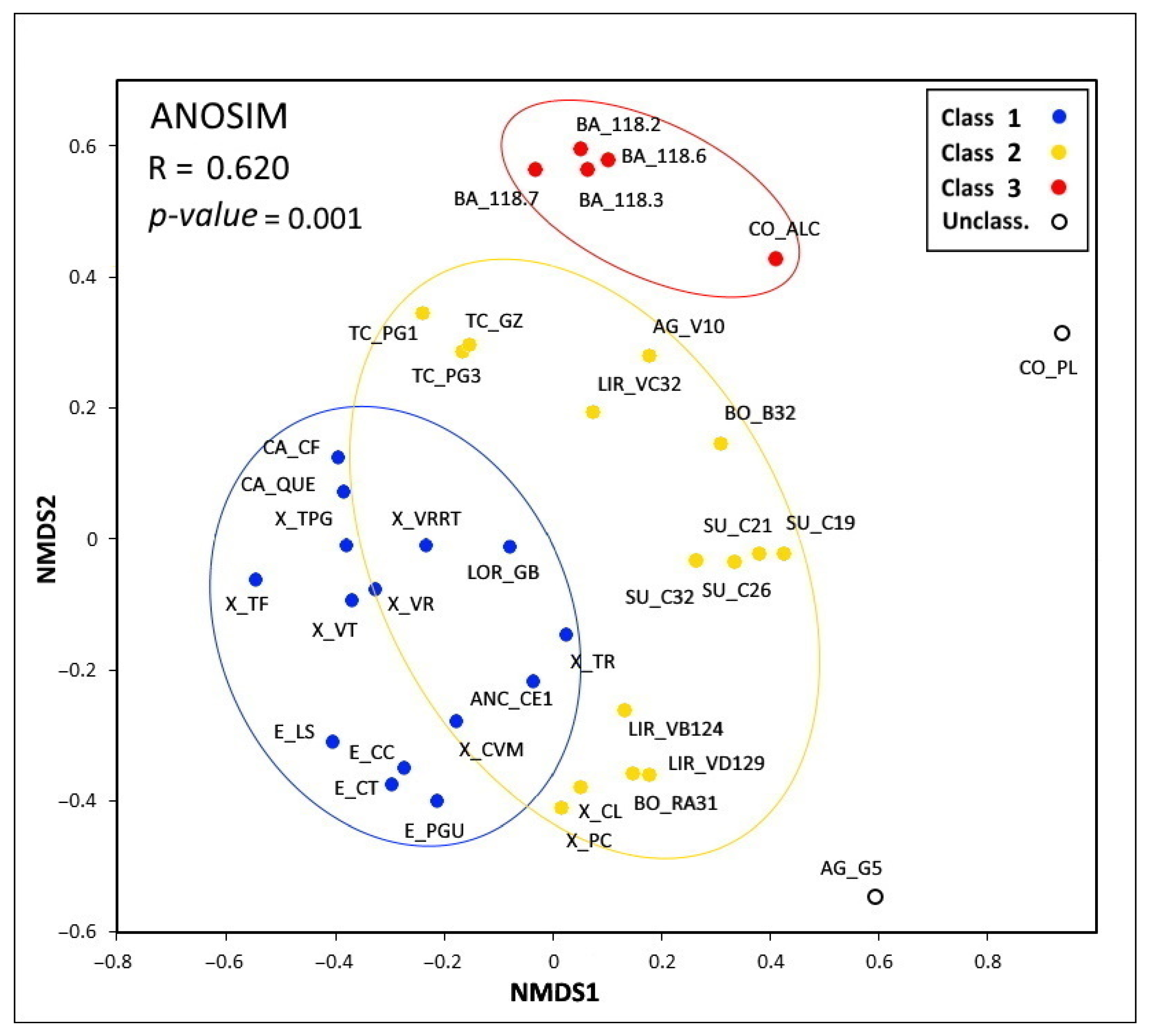
3.2. Testate Amoeba Assemblages
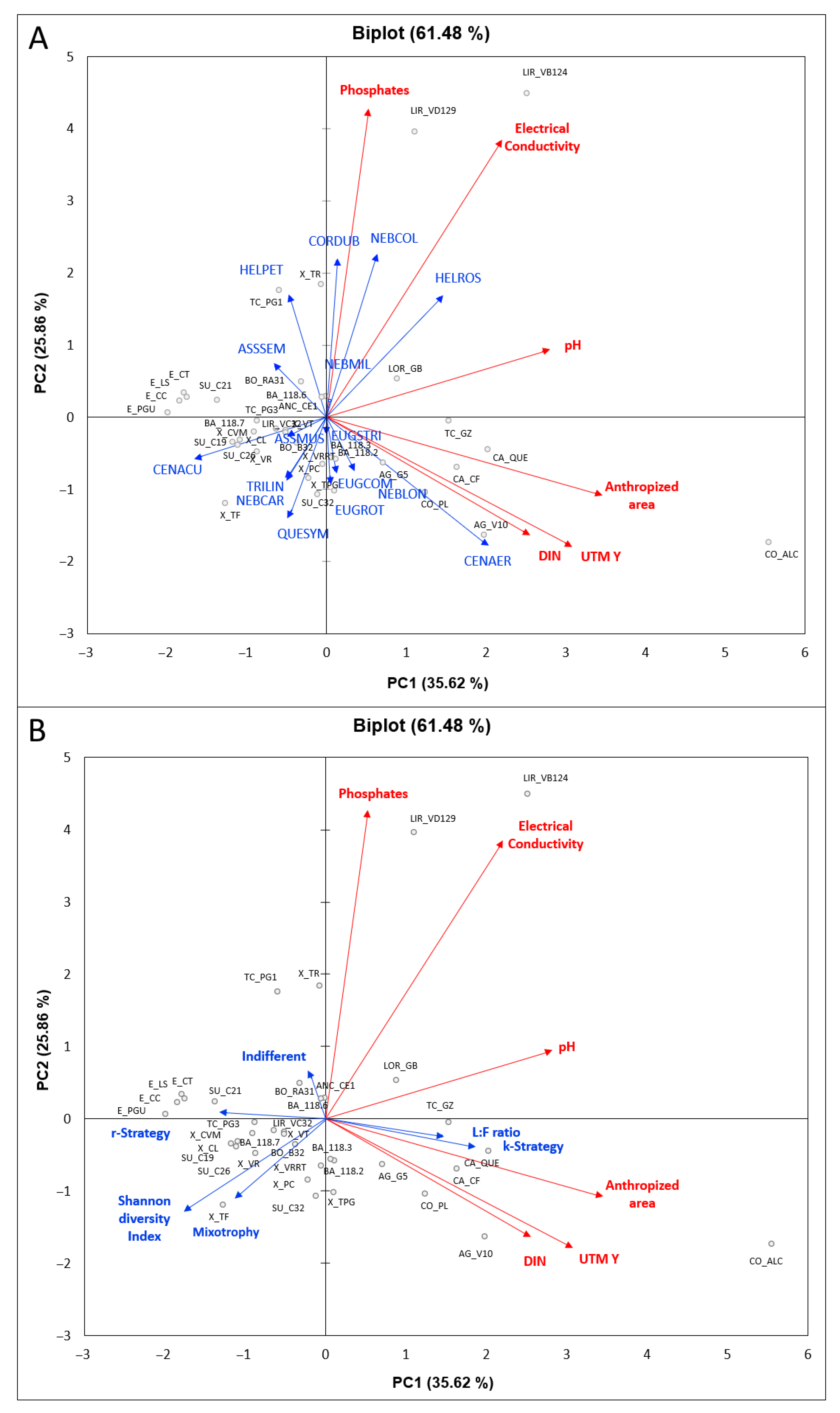
3.3. Effects of Environmental Variables on the Testate Amoeba Assemblages
4. Discussion
4.1. Iberian Peatlands Water Chemistry
4.2. Ecological Drivers of Testate Amoeba Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gorham, E. Northern peatlands: Role in the carbon cycle and probably responses to climate warming. Ecol. Appl. 1991, 1, 182–195. [Google Scholar] [CrossRef]
- Loisel, J.; Yu, Z.; Beilman, D.W.; Camill, P.; Alm, J.; Amesbury, M.J.; Anderson, D.; Andersson, S.; Bochicchio, C.; Barber, K.; et al. A database and synthesis of northern peatland soil properties and Holocene carbon and nitrogen accumulation. Holocene 2014, 24, 1028–1042. [Google Scholar] [CrossRef]
- Leifeld, J.; Menichetti, L. The underappreciated potential of peatlands in global climate change mitigation strategies. Nat. Commun. 2018, 9, 1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, T.R.; Knowles, R. Methane emissions from fen, bog, and swamp peatlands in Quebec. Biogeochemistry 1990, 11, 45–61. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef]
- Shukla, P.R.; Skea, J.; Calvo Buendia, E.; Masson-Delmotte, V.; Pörtner, H.O.; Roberts, D.C.; Zhai, P.; Slade, R.; Connors, S.; van Diemen, R.; et al. An IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems. In IPCC, 2019: Climate Change and Land; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Minayeva, T.Y.; Bragg, O.M.; Sirin, A.A. Towards ecosystem-based restoration of peatland biodiversity. Mires Peat 2017, 1, 1–36. [Google Scholar]
- European Commission. European Commission Interpretation Manual of European Union Habitats, Vers. EUR28; European Commission, DG Environment: Brussel, Belgium, 2013; p. 146. Available online: https://ec.europa.eu/environment/nature/legislation/habitatsdirective/docs/Int_Manual_EU28.pdf (accessed on 15 February 2021).
- Joosten, H. Mires in Europe: A preliminary status report. Int. Mire Conserv. Group Memb. Newslett. 1997, 3, 10–13. [Google Scholar]
- Pontevedra-Pombal, X.; Castro, D.; Carballeira, R.; Souto, M.; López-Sáez, J.A.; Pérez-Díaz, S.; Fraga, M.I.; Valcárcel, M.; García-Rodeja, E. Iberian acid peatlands: Types, origin and general trends of development. Mires Peat 2017, 19, 21. [Google Scholar]
- Koenig, I.; Feldmeyer-Christie, E.; Mitchell, E.A.D. Comparative ecology of vascular plant, bryophyte and testate amoeba communities in four Sphagnum peatlands along a altitudinal gradient in Switzerland. Ecol. Indic. 2015, 54, 48–59. [Google Scholar] [CrossRef]
- Marcisz, K.; Colombaroli, D.; Jassey, V.E.J.; Tinner, W.; Kołaczek, P.; Gałka, M.; Karpińska-Kołaczek, M.; Słowiński, M.; Mariusz Lamentowicz, M. A novel testate amoebae trait-based approach to infer environmental disturbance in Sphagnum peatlands. Sci. Rep. 2016, 6, 33907. [Google Scholar] [CrossRef] [Green Version]
- Niedzwiecki, M.; Mieczan, T.; Adamczuk, M. Ecology of testate amoebae (Protists) in a Sphagnum-dominated peat bog and the relationship between species assemblages and environmental parameters. Oceanol. Hydrobiol. Stud. 2016, 45, 344–352. [Google Scholar] [CrossRef]
- Jassey, V.E.J.; Chiapusio, G.; Binet, P.; Buttler, A.; Laggoun-Defarge, F.; Delarue, F.; Bernard, N.; Mitchell, E.A.; Toussaint, M.L.; Francez, A.J.; et al. Above and belowground linkages in Sphagnum peatland: Climate warming affects plant-microbial interactions. Glob. Chang. Biol. 2013, 19, 811–823. [Google Scholar] [CrossRef] [Green Version]
- Jassey, V.E.; Signarbieux, C.; Hättenschwiler, S.; Bragazza, L.; Buttler, A.; Delarue, F.; Fournier, B.; Gilbert, D.; Laggoun-Défarge, F.; Lara, E.; et al. An unexpected role for mixotrophs in the response of peatland carbon cycling to climate warming. Sci. Rep. 2015, 5, 16931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heal, O.W. Observations on the Seasonal and Spatial-Distribution of Testacea (Protozoa, Rhizopoda) in Sphagnum. J. Anim. Ecol. 1964, 33, 395–412. [Google Scholar] [CrossRef]
- Jassey, V.E.J.; Chiapusio, G.; Mitchell, E.A.D.; Binet, P.; Toussaint, M.L.; Gilbert, D. Fine-sclae horizontal and vertical micro-distribution patterns of testate amoebae along a narrow fen/bog gradient. Microb. Ecol. 2011, 61, 374–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basinska, A.M.; Reczuga, M.K.; Gabka, M.; Strózecki, M.; Łuców, D.; Samson, M.; Urbaniak, M.; Lesny, J.; Chojnicki, B.H.; Gilbert, D.; et al. Experimental warming and precipitation reduction affect the biomass of microbial communities in a Sphagnum peatland. Ecol. Indic. 2020, 112, 106059. [Google Scholar] [CrossRef]
- Tolonen, K.; Warner, B.G.; Vasander, H. Ecology of Testaceans (Protozoa, Rhizopoda) in mires in South-ern Finland. 2. Multivariate-Analysis. Arch. Protistenkd. 1994, 144, 97–112. [Google Scholar] [CrossRef]
- Mitchell, E.A.D.; Borcard, D.; Buttler, A.J.; Grosvernier, P.; Gilbert, D.; Gobat, J.M. Horizontal Distribution Patterns of Testate Amoebae (Protozoa) in a Sphagnum magellanicum Carpet. Microb. Ecol. 2000, 39, 290–300. [Google Scholar] [PubMed]
- Davis, S.R.; Wilkinson, D.M. The conservation management value of testate amoebae as ‘restoration’ indicators: Speculations based on two damaged raised mires in northwest England. Holocene 2004, 14, 135–143. [Google Scholar] [CrossRef]
- Mitchell, E.A.D.; Charman, D.J.; Warner, B.G. Testate amoebae analysis in ecological and paleoecological studies of wetlands: Past, present and future. Biodivers. Conserv. 2008, 17, 2115–2137. [Google Scholar] [CrossRef] [Green Version]
- Mieczan, T. Ecology of testate amoebae (Protists) in Sphagnum peatlands of eastern Poland: Vertical micro-distribution and species assemblages in relation to environmental parameters. Ann. Limnol. Int. J. Limnol. 2009, 45, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Mieczan, T.; Adamczuk, M. Ecology of testate amoebae (Protists) in mosses: Distribution and relation of species assemblages with environmental parameters (King George Island, Antarctica). Polar Biol. 2015, 38, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Lamentowicz, M.; Mitchell, E.A.D. Testate amoebae as ecological and palaeohydrological indicators in peatlands—The Polish experience. In Wetlands: Monitoring, Modelling and Management; Okruszko, T., Maltby, E., Szatylowicz, J., Miroslaw-Swiatek, D., Eds.; Taylor & Francis Group: London, UK, 2007; pp. 85–90. [Google Scholar]
- Mitchell, E.A.D.; Bragazza, L.; Gerdol, R. Testate Amoebae (Protista) Communities in Hylocomium splendens (Hedw.) B.S.G. (Bryophyta): Relationships with Altitude, and Moss Elemental Chemistry. Protist 2004, 155, 423–436. [Google Scholar] [CrossRef] [Green Version]
- Marcisz, K.; Jassey, V.E.J.; Kosakyan, A.; Krashevska, V.; Lahr, D.J.G.; Lara, E.; Lamentowicz, M.; Macumber, A.; Mazei, Y.; Mitchell, E.A.D.; et al. Testate amoebae functional traits and their use in paleoecology. Front. Ecol. Evol. 2020, 8, 575966. [Google Scholar] [CrossRef]
- Charman, D.J. Biostratigraphic and palaeoenvironmental applications of testate amoebae. Quat. Sci. Rev. 2001, 20, 1753–1764. [Google Scholar] [CrossRef]
- Turner, T.E.; Swindles, G.T. Ecology of testate amoebae in moorland with a complex fire history: Implications for ecosystem monitoring and sustainable land management. Protist 2012, 163, 844–855. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Mitchell, E.A.D.; Lamentowicz, M.; Payne, R.J.; Lara, E.; Gu, Y.; Huang, X.; Wang, H. Ecology of testate amoebae in peatlands of central China and development of a transfer function for paleohydrological reconstruction. J. Paleolimnol. 2013, 50, 319–330. [Google Scholar] [CrossRef]
- Lamentowicz, M.; Kajukało-Drygalska, K.; Kołaczek, P.; Jassey, V.E.J.; Gabka, M.; Karpinska-Kołaczek, M. Testate amoebae taxonomy and trait diversity are coupled along an openness and wetness gradient in pine-dominated Baltic bogs. Eur. J. Protistol. 2020, 73, 125674. [Google Scholar] [CrossRef]
- Heras-Pérez, P.; Infante-Sánchez, M.; Pontevedra-Pombal, X.; Nóvoa-Muñoz, J.C. Part II, Country chapters: Spain. In Mires and Peatlands of Europe; Joosten, H., Tanneberger, F., Moen, A., Eds.; Schweizerbart Science Publishers: Sttutgart, Germany, 2017; pp. 639–656. [Google Scholar]
- Gutiérrez, F.; Gutiérrez, M.; Martín-Serrano, Á. The Geology and Geomorphology of Spain: A Concise Introduction. In Landscapes and Landforms of Spain; Gutiérrez, F., Gutiérrez, M., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 1–23. [Google Scholar]
- Castro, D.; Souto, M.; Fraga, M.I.; García-Rodeja, E.; Pérez-Díaz, S.; López-Sáez, J.A.; Pontevedra-Pombal, X. High-resolution patterns of palaeoenvironmental changes during the Little Ice Age and the Medieval Climate Anomaly in the northwestern Iberian Peninsula. Geosci. Front. 2020, 11, 1461–1475. [Google Scholar] [CrossRef]
- AEMET; IPM. Iberian Climate Atlas; Agencia Estatal de Meteorología: Madrid, Spain; Instituto Portugués de Meteorologia: Lisboa, Portugal, 2011; p. 81.
- Mitchell, E.A.D.; Gilbert, D. Vertical micro-distribution and response to nitrogen deposition of testate amoebae in Sphagnum. J. Eukaryot. Microb. 2004, 51, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Booth, R.K.; Lamentowicz, M.; Charman, D. Preparation and analysis of testate amoebae in peatland paleoenvironmental studies. Mires Peat 2010, 7, 1–7. [Google Scholar]
- APHA-AWWA-WEF. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Bonnet, L. Le peuplement thécamoebien édaphique de la Côte-d’Ivoire. Sols Reg. Lamto Protistol. 1976, 12, 539–554. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; p. 117. [Google Scholar]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- XLSTAT. Data Analysis and Statistical Solution for Microsoft Excel Addisonf (XLSTAT Version 2021.2.2); XLSTAT: Paris, France, 2021; Available online: https://www.xlstat.com/ (accessed on 1 March 2021).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; Stevens, H.; et al. Community Ecology Package: Ordination, Diversity and Dissimilarities. Version 2013, 2, 295. Available online: https://cran.r-project.org; https://github.com/vegandevs/vegan (accessed on 10 January 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 10 January 2021).
- Muñoz, J.; Aldasoro, J.J.; Negro, A.; de Hoyos, C.; Vega, J.C. Flora and water chemistry in a relictic mirre complex: The Sierra Segundera mire area (Zamora, NW Spain). Hydrobiologia 2003, 495, 1–16. [Google Scholar] [CrossRef]
- Holden, J.; Chapman, P.J.; Labadz, J.C. Artificial drainage of peatlands: Hydrological and hydrochemical process and wetland restoration. Prog. Phys. Geogr. 2004, 28, 95–123. [Google Scholar] [CrossRef] [Green Version]
- Luscombe, D.J.; Anderson, K.; Grand-Clement, E.; Gatis, N.; Ashe, J.; Benaud, P.; Smith, D.; Brazier, R.E. How does drainage alter the hydrology of shallow degraded peatlands across multiple spatial scales? J. Hydrol. 2016, 541, 1329–1339. [Google Scholar] [CrossRef] [Green Version]
- Menberu, M.W.; Tahvanainen, T.; Marttila, H.; Irannezhad, M.; Bonkanen, A.K.; Penttinen, J.; Klove, B. Water-table-dependent hydrological changes following peatland forestry drainage and restoration: Analysis of restoration success. Water Resour. Res. 2016, 52, 3742–3760. [Google Scholar] [CrossRef] [Green Version]
- Lundin, L.; Nilsson, T.; Jordan, S.; Lode, E.; Strömgren, M. Impacts of rewetting on peat, hydrology and water chemical composition over 15 years in two finished peat extraction areas in Sweden. Wetl. Ecol. Manag. 2017, 25, 405–419. [Google Scholar] [CrossRef] [Green Version]
- Bourbonniere, R.A. Review of Water Chemistry Research in Natural and Disturbed Peatlands. Can. Water. Resour. J. 2009, 34, 393–414. [Google Scholar] [CrossRef] [Green Version]
- Bragazza, L.; Parisod, J.; Buttler, A.; Bardgett, R.D. Biogeochemical plant-soil microbe feedback in response to climate warming in peatlands. Nat. Clim. Chang. 2013, 3, 273–277. [Google Scholar] [CrossRef]
- Lamentowicz, M.; Bragazza, L.; Buttler, A.; Jassey, V.E.J.; Mitchell, E.A.D. Seasonal patterns of testate amoeba diversity, community structure and species environment relationships in four Sphagnum-dominated peatlands along a 1300 m altitudinal gradient in Switzerland. Soil Biol. Biochem. 2013, 67, 1–11. [Google Scholar] [CrossRef]
- Rydin, H.; Jeglum, J.K. The Biology of Peatlands; Oxford University Press: Oxford, UK, 2006; pp. 1–296. [Google Scholar]
- Alonso, M. Las lagunas de la España peninsular. Limnetica 1998, 15, 1–176. [Google Scholar]
- Varela-Silvalde, O.; Castro, D.; Pontevedra-Pombal, X.; Antelo, J. Chemical composition and dynamics of the physico-chemical properties of peatland sub-catchments from the Xistral Mountains (NW Spain). In Challenges and Opportunities in Soil Scienc; Andavira Editora S.L.: Santiago de Compostela, Spain, 2014; pp. 185–189. [Google Scholar]
- Pérez-Haase, A.; Ninot, J.M. Hydrological heterogeneity rather than water chemistry explains the high plant diversity and uniqueness of a Pyrenean mixed mire. Folia Geobot. 2017, 52, 143–160. [Google Scholar] [CrossRef] [Green Version]
- Colomer, J.; Pérez-Haase, A.; Carrillo, E.; Ventura, M.; Ninot, J.M. Fine-scale vegetation mosaics in Pyrenean mires are driven by complex hydrological regimes and threatened by extreme weather events. Ecohydrology 2018, 12, e2070. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, X.; Wang, Z.; Xu, Z.; He, C.; Sheng, L.; Liu, H.; Wang, Z. Shift in nitrogen transformation in peatland soil by nitrogen inputs. Sci. Total Environ. 2021, 764, 142924. [Google Scholar] [CrossRef] [PubMed]
- Eurola, S.; Holappa, K. The Finnish mire type system. Aquil. Ser. Bot. 1985, 21, 101–110. [Google Scholar]
- Malmer, N. Vegetational gradients in relation to environmental conditions in northwestern European mires. Can. J. Bot. 1986, 64, 375–383. [Google Scholar] [CrossRef]
- Gracia-Royo, M.P. Tecamebas muscícolas del Valle de Ribas (Gerona). Publ. Inst. Biol. Apl. 1964, 37, 67–71. [Google Scholar]
- Gracia-Royo, M.P. Tecamebas esfagnícolas de Panticosa (Pirineos Aragoneses). Instituto de Estudios Pirenaicos CSIC Jaca. Actas V Congr. Int. Estud. Pirenaicos 1966, 79, 7–12. [Google Scholar]
- Gracia-Royo, M.P. Tecamebas esfagnícolas de la Península Ibérica: Contribución al estudio de las tecamebas (Protozoa, Thecameboidea). Publ. Inst. Biol. Apl. 1972, 52, 5–42. [Google Scholar]
- Margalef, R. Datos para la hidrobiología de la Sierra de Guadarrama. Publ. Inst. Biol. Apl. 1949, 6, 5–21. [Google Scholar]
- Margalef, R. Comunidades bióticas de las aguas dulces del noroeste de España. Publ. Inst. Biol. Apl. 1955, 21, 137–171. [Google Scholar]
- Margalef, R. Algas de agua dulce del noroeste de España. Publ. Instit. Biol. Apli. 1956, 22, 43–152. [Google Scholar]
- Pontevedra-Pombal, X.; Carballeira, R.; Crujeiras, U.; García-Rodeja, E. Characterization of testate amoebae in edaphic microhabitats of a histosol of the Serra do Xistral (Galicia). In Soil, a Sustainable Resource for the Bioeconomy; Besga, G., Gartzia, N., Arias, A., Zabaleta, A., Antigüedad, I., Virto, I., Mataiz-Solera, J., Alexandre, C., Eds.; Instituto Vasco de Investigación y Desarrollo Agrario: Donostia, Spain, 2018; pp. 630–633. [Google Scholar]
- Mitchell, E.A.D.; Buttler, J.; Warner, B.G.; Gobat, J.M. Ecology of testate amoebae (Protozoa: Rhizopoda) in Sphagnum peatlands in the Jura mountains, Switzerland and France. Ecoscience 1999, 6, 565–576. [Google Scholar] [CrossRef]
- Lamentowicz, M.; Mitchell, E.A.D. The ecology of testate amoebae (Protists) in Sphagnum in north-western Poland in relation to peatland ecology. Microb. Ecol. 2005, 50, 48–63. [Google Scholar] [CrossRef]
- Kosakyan, A.; Gomaa, F.; Mitchell, E.A.D.; Heger, T.J.; Lara, E. Using DNA-barcoding for sorting out protist species complex: A case study of the Nebela tincta-collaris-bohemica group (Amoebozoa; Arcellinida, Hyalospheniidae). Eur. J. Protistol. 2013, 49, 222–237. [Google Scholar] [CrossRef] [PubMed]
- Warner, B.; Asada, T.; Quinn, N. Seasonal influences on the ecology of testate amoebae (Protozoa) in a small Sphagnum peatland in Southern Ontario, Canada. Microb. Ecol. 2007, 54, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, E.A.D.; Buttler, A.; Grosvernier, P.; Rydin, H.; Al-binsson, C.; Greenup, A.L.; Heijmans, M.M.P.D.; Hoos-beek, M.R.; Saarinen, T. Relationships among testate amoebae (Protozoa), vegetation and water chemistry in five Sphagnum-dominated peatlands in Europe. New Phytol. 2000, 145, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, E.A.D. Response of testate amoebae (Protozoa) to N and P fertilization in an Arctic wet sedge tundra. Arct. Antarct. Alp. Res. 2004, 36, 77–82. [Google Scholar] [CrossRef]
- Heger, T.J.; Straub, F.; Mitchell, E.A.D. Impact of farming practices on soil diatoms and testate amoebae: A pilot study in the DOK-trial at Therwil, Switzerland. Eur. J. Soil Biol. 2012, 49, 31–36. [Google Scholar] [CrossRef]
- Carballeira, A.; Devesa, C.; Retuerto, R.; Santillán, E.; Ucieda, F. Bioclimatología de Galicia; Fundación Pedro Barrié de la Maza Conde de FENOSA: La Coruña, Spain, 1983; p. 250. [Google Scholar]
- Krashevska, V.; Bonkowski, M.; Maraun, M.; Scheu, S. Testate amoebae (protista) of an elevational gradient in the tropical mountain rain forest of Ecuador. Pedobiologia 2007, 51, 319–331. [Google Scholar] [CrossRef]
- Krashevska, V.; Bonkowski, M.; Maraun, M.; Ruess, L.; Kandeler, L.; Scheu, S. Microorganisms as driving factors for the community structure of testate amoebae along an altitudinal transect in tropical mountain rain forests. Soil Biol. Biochem. 2008, 40, 2427–2433. [Google Scholar] [CrossRef]
- Bonnet, L. Les thécamobiens, indicateurs pédologiques, et la notion de climax. Bull. Soc. Hist. Nat. Toulouse 1961, 96, 80–86. [Google Scholar]
- Bonnet, L. Le peuplement thécamobiens des sols. Rev. Écol. Biol. Sol 1964, 1, 123–408. [Google Scholar]
- Bonnet, L. Le signalement écologique des Thécamoebiens du sol. Bull. Soc. Hist. Nat. Toulouse 1984, 120, 103–110. [Google Scholar]
- Bonnet, L. Le signalement écologique des Thécamoebiens du sol (deuxième partie). Bull. Soc. Hist. Nat. Toulouse 1985, 121, 7–12. [Google Scholar]
- Bonnet, L. Signalement des préférences écologiques des Thécamoebiens. Bull. Lab. Biol. Quant. 1986, 2, 22–25. [Google Scholar]
- Bonnet, L. Ecologie de quelques Euglyphidae (Thécamoebiens, Filosea) des milieux édaphiques et para édaphiques (Troisième partie: Genre Euglypha). Bull. Soc. Hist. Nat. Toulouse 1992, 128, 11–20. [Google Scholar]
- Booth, R.K.; Zygmunt, J.R. Biogeography and comparative ecology of testate amoebae inhabiting Sphagnum-dominated peatlands in the Great Lakes and Rocky Mountain regions of North America. Divers. Distrib. 2005, 11, 577–590. [Google Scholar] [CrossRef]
- Booth, R.K. Testate amoebae as proxies for mean annual water-table depth in Sphagnum-dominated peatlands of North America. J. Quat. Sci. 2008, 23, 43–57. [Google Scholar] [CrossRef]
- Booth, R.K.; Sullivan, M.E.; Sousa, V.A. Ecology of testate amoebae in a North Carolina pocosin and their potential use as environmental and paleoenvironmental indicators. Ecoscience 2008, 15, 277–289. [Google Scholar] [CrossRef]
- Sullivan, M.E.; Booth, R.K. The potential influence of short-term environmental variability on the composition of testate amoeba communities in Sphagnum peatlands. Microb. Ecol. 2011, 62, 80–93. [Google Scholar] [CrossRef]
- Beyens, L.; Ledeganck, P.; Graae, B.J.; Nijs, I. Are soil biota buffered against climatic extremes? An experimental test on testate amoebae in arctic tundra (Qeqertarsuaq, West Greenland). Polar Biol. 2009, 32, 453–462. [Google Scholar] [CrossRef]
- Bonnet, L. Les thécamoebiens des Bouillouses. Bull. Soc. D’histoire Nat. Toulouse 1958, 93, 529–543. [Google Scholar]
- Schönborn, W. Untersuchungen über die Zoochlorellen-Symbiose der Hochmoor-Testaceen. Limnologica 1965, 3, 173–176. [Google Scholar]
- Gómez-Sánchez, M.S. Introducción al estudio de los Tecamebidos, (Protozoa) en Asturias. Bol. Cienc. Nat. Inst. Estud. Astur 1980, 25, 83–90. [Google Scholar]
- Bankov, N.; Todorov, M.; Ganeva, A. Checklist of Sphagnum-dwelling testate amoebae in Bulgaria. Biodivers. Data J. 2019, 6, e25295. [Google Scholar] [CrossRef] [Green Version]
- Török, J. Study on moss-dwelling testate amoebae. Opusc. Zool. 1993, 26, 95–104. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peatlands | Code | Locality (Province) | Country | Geographic Area | EU Habitat Directive | Coordinates (Datum ETRS89) | Altitude | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Huse | UTM X | UTM Y | (m a.s.l.) | |||||||
| Cruz do Frade (1) | CA_CF | Cariño (A Coruña, Galicia) | Spain | Serra da Capelada (CA) | Meso-eutrophic | 7230 | 29 T | 583605 | 4837869 | 530 |
| Queizal (2) | CA_QUE | Cariño (A Coruña, Galicia) | Spain | Serra da Capelada (CA) | Meso-eutrophic | 7230 | 29 T | 584005 | 4837885 | 490 |
| Rego do Tremoal (3) | X_VRRT | Abadín (Lugo, Galicia) | Spain | Serra do Xistral (X) | Acidic fens | 7140 | 29 T | 626494 | 4812597 | 570 |
| Tremoal de Penas Gordas (4) | X_TPG | Abadín (Lugo, Galicia) | Spain | Serra do Xistral (X) | Acidic fens | 7140 | 29 T | 626534 | 4811601 | 625 |
| Veiga do Rial (5) | X_VR | Abadín (Lugo, Galicia) | Spain | Serra do Xistral (X) | Acidic fens | 7140 | 29 T | 616311 | 4810992 | 750 |
| Tremoal da Revolta (6) | X_TR | Abadín (Lugo, Galicia) | Spain | Serra do Xistral (X) | Acidic fens | 7140 | 29 T | 619398 | 4812159 | 700 |
| Chao de Veiga Mol (7) | X_CVM | O Valadouro (Lugo, Galicia) | Spain | Serra do Xistral (X) | Raised bog | 7110 | 29 T | 620745 | 4822070 | 695 |
| Tremoal do Fiuco (8) | X_TF | Abadín (Lugo, Galicia) | Spain | Serra do Xistral (X) | Raised bog | 7110 | 29 T | 618996 | 4811886 | 700 |
| Veiga do Tremoal (9) | X_VT | Abadín (Lugo, Galicia) | Spain | Serra do Xistral (X) | Acidic fens | 7140 | 29 T | 620106 | 4811318 | 700 |
| Chao de Lamoso (10) | X_CL | O Valadouro (Lugo, Galicia) | Spain | Serra do Xistral (X) | Blanket bog | 7130 | 29 T | 617074 | 4817170 | 1020 |
| Pena da Cadela (11) | X_PC | Muras (Lugo, Galicia) | Spain | Serra do Xistral (X) | Blanket bog | 7130 | 29 T | 616449 | 4816677 | 1014 |
| Rego de Pena Grande (12) | TC_PG1 | Xermade (Lugo, Galicia) | Spain | Terra Chá (TC) | Acidic fens | 7140 | 29 T | 605556 | 4692824 | 480 |
| Rego de Pena Grande (13) | TC_PG3 | Xermade (Lugo, Galicia) | Spain | Terra Chá (TC) | Acidic fens | 7140 | 29 T | 605785 | 4692891 | 485 |
| Lagoa de Fabás (14) | TC_GZ | Vilalba (Lugo, Galicia) | Spain | Terra Chá (TC) | Acidic fens | 7140-pond | 29 T | 613605 | 4799346 | 435 |
| Lagoa de Alcaián (15) | CO_ALC | Coristanco (A Coruña, Galicia) | Spain | Chairas Occidentais (Western Plains) (CO) | Acidic fens | 7140-6410 Molinea meadows-Atlantic wet heath | 29 T | 520943 | 4775406 | 400 |
| Poza do Lameiro (16) | CO_PL | Lousame (A Coruña, Galicia) | Spain | Chairas Occidentais (Western Plains) (CO) | Acidic fens | 7140 | 29 T | 515709 | 4733290 | 450 |
| Rego Grande (17) | BO_B32 | Toques (A Coruña, Galicia) | Spain | Montes do Bocelo (BO) | Acidic fens | 7140-grasslands | 29 T | 581205 | 4759180 | 600 |
| Río Ameneiros (18) | BO_RA31 | Toques (A Coruña, Galicia) | Spain | Montes do Bocelo (BO) | Acidic fens | 7140 | 29 T | 581388 | 4760452 | 705 |
| Barranqueira da Devesa da Graña (19) | BA_118.2 | Boiro (A Coruña, Galicia) | Spain | Serra do Barbanza (BA) | Acidic fens | 7140 | 29 T | 505495 | 4727159 | 540 |
| Rego da Puntiña (20) | BA_118.3 | Boiro (A Coruña, Galicia) | Spain | Serra do Barbanza (BA) | Acidic fens | 7140 | 29 T | 505318 | 4726200 | 555 |
| Chan das Brañas (21) | BA_118.6 | Boiro (A Coruña, Galicia) | Spain | Serra do Barbanza (BA) | Acidic fens | 7140 | 29 T | 502641 | 4723368 | 650 |
| Rego das Brañas (22) | BA_118.7 | Boiro (A Coruña, Galicia) | Spain | Serra do Barbanza (BA) | Acidic fens | 7140 | 29 T | 503665 | 4722536 | 525 |
| Campa dos Extremeños (23) | ANC_CE1 | Cervantes (Lugo, Galicia) | Spain | Serra dos Ancares (ANC) | Acidic fens | 7140 | 29 T | 675506 | 4743659 | 1615 |
| As Pontellas 1 (24) | SU_C19 | A Lama (Pontevedra, Galicia) | Spain | Serra do Suído (SU) | Acidic fens | 7140-7110 | 29 T | 554323 | 4696467 | 855 |
| As Pontellas 2 (25) | SU_C21 | A Lama (Pontevedra, Galicia) | Spain | Serra do Suído (SU) | Acidic fens | 7140 | 29 T | 554364 | 4696930 | 860 |
| Costa de Uxía (26) | SU_C26 | A Lama (Pontevedra, Galicia) | Spain | Serra do Suído (SU) | Acidic fens | 7140 | 29 T | 551521 | 4695120 | 820 |
| As Landras (27) | SU_C32 | A Lama (Pontevedra, Galicia) | Spain | Serra do Suído (SU) | Acidic fens | 7140 | 29 T | 553636 | 4696430 | 870 |
| Brañas da Groba (28) | AG_G5 | Oia (Pontevedra, Galicia) | Spain | Serra da Groba (AG) | Acid peaty soil | Para-peaty wetland | 29 T | 513096 | 4657227 | 525 |
| A Portela (29) | AG_V10 | Oia (Pontevedra, Galicia) | Spain | Serra de Valga (VA) | Acidic fens | 7140 | 29 T | 512525 | 4648061 | 350 |
| Gándaras de Budiño (30) | LOR_GB | Porriño (Pontevedra, Galicia) | Spain | Louro River Valley (LOR) | Acidic fens | 7140 | 29 T | 530591 | 4660889 | 30 |
| Veiga de Golpellás (31) | LIR_VB124 | Calvos de Randín-Muíños (Ourense, Galicia) | Spain | Limia River Valley (LIR) | Acidic fens | 7140 | 29 T | 593321 | 4647552 | 865 |
| Val de Deus (32) | LIR_VD129 | Muíños (Ourense, Galicia) | Spain | Limia River Valley (LIR) | Acidic fens | 7140 | 29 T | 589484 | 4645252 | 875 |
| Veiga do Calvelo do Río (33) | LIR_VC32 | Baños de Molgas (Ourense, Galicia) | Spain | Limia River Valley (LIR) | Acidic fens | 7140 | 29 T | 612374 | 4675533 | 520 |
| Covao da Clareza (34) | E_CC | Seia (Beira Alta) | Portugal | Serra da Estrela (E) | Acidic fens | 7140 | 29 T | 617938 | 4466145 | 1820 |
| Cabeço do Talao (35) | E_CT | Seia (Beira Alta) | Portugal | Serra da Estrela (E) | Acidic fens | 7140 | 29 T | 616955 | 4466623 | 1850 |
| Pedras Guieiras (36) | E_PGU | Seia (Beira Alta) | Portugal | Serra da Estrela (E) | Acidic fens | 7140 | 29 T | 614406 | 4468721 | 1590 |
| Lagoa Seca (37) | E_LS | Seia (Beira Alta) | Portugal | Serra da Estrela (E) | Acidic fens | 7140 | 29 T | 616302 | 4469814 | 1625 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carballeira, R.; Pontevedra-Pombal, X. Diversity of Testate Amoebae as an Indicator of the Conservation Status of Peatlands in Southwest Europe. Diversity 2021, 13, 269. https://doi.org/10.3390/d13060269
Carballeira R, Pontevedra-Pombal X. Diversity of Testate Amoebae as an Indicator of the Conservation Status of Peatlands in Southwest Europe. Diversity. 2021; 13(6):269. https://doi.org/10.3390/d13060269
Chicago/Turabian StyleCarballeira, Rafael, and Xabier Pontevedra-Pombal. 2021. "Diversity of Testate Amoebae as an Indicator of the Conservation Status of Peatlands in Southwest Europe" Diversity 13, no. 6: 269. https://doi.org/10.3390/d13060269
APA StyleCarballeira, R., & Pontevedra-Pombal, X. (2021). Diversity of Testate Amoebae as an Indicator of the Conservation Status of Peatlands in Southwest Europe. Diversity, 13(6), 269. https://doi.org/10.3390/d13060269