
Unfolding Jellyfish Bloom Dynamics along the Mediterranean Basin by Transnational Citizen Science Initiatives

 ,
, 

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Jellyfish Citizen Science-Based Programs
2.2. Predictor (Environmental) Data
2.3. Data Analysis
3. Results
3.1. Jellyfish Dynamics
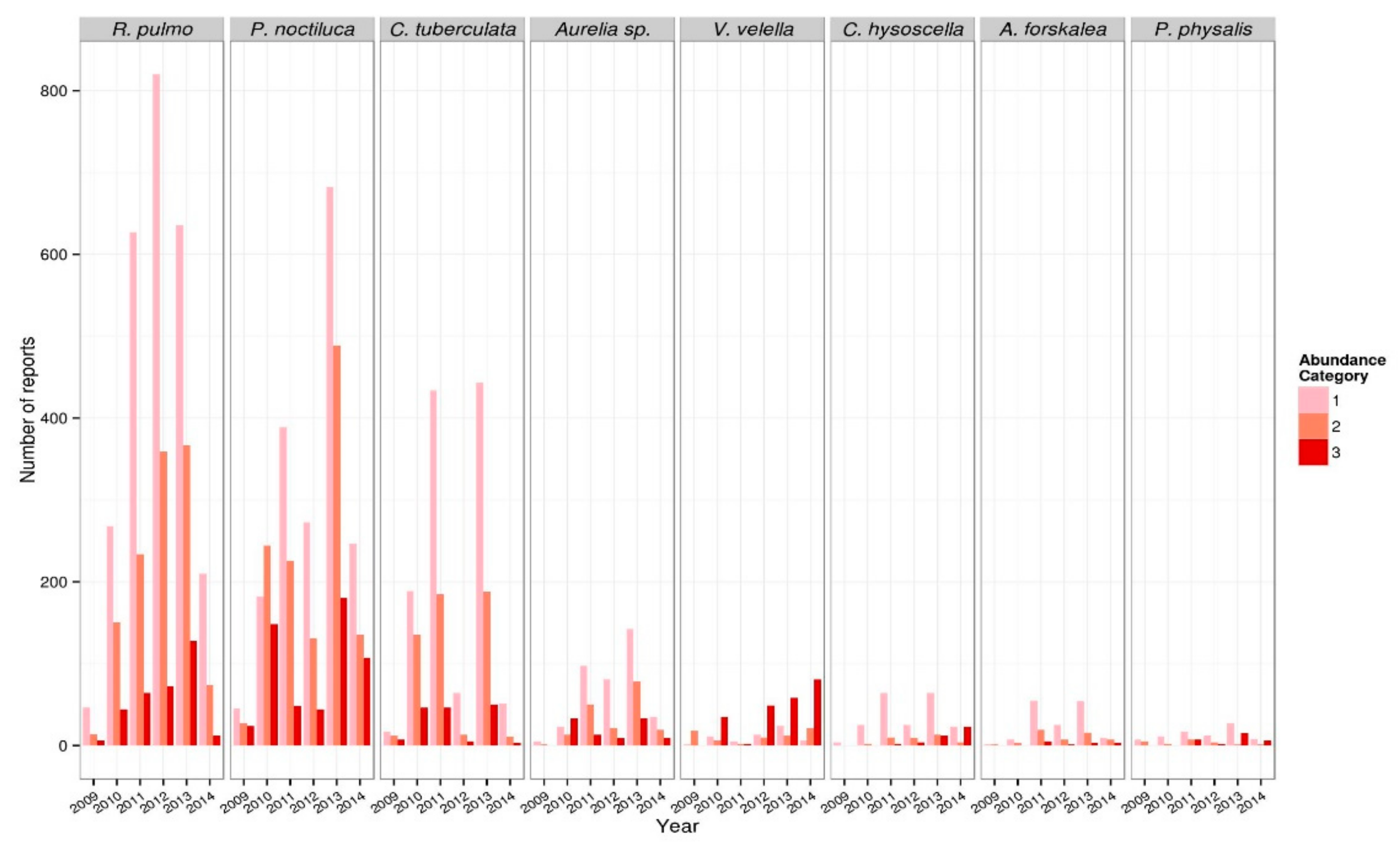
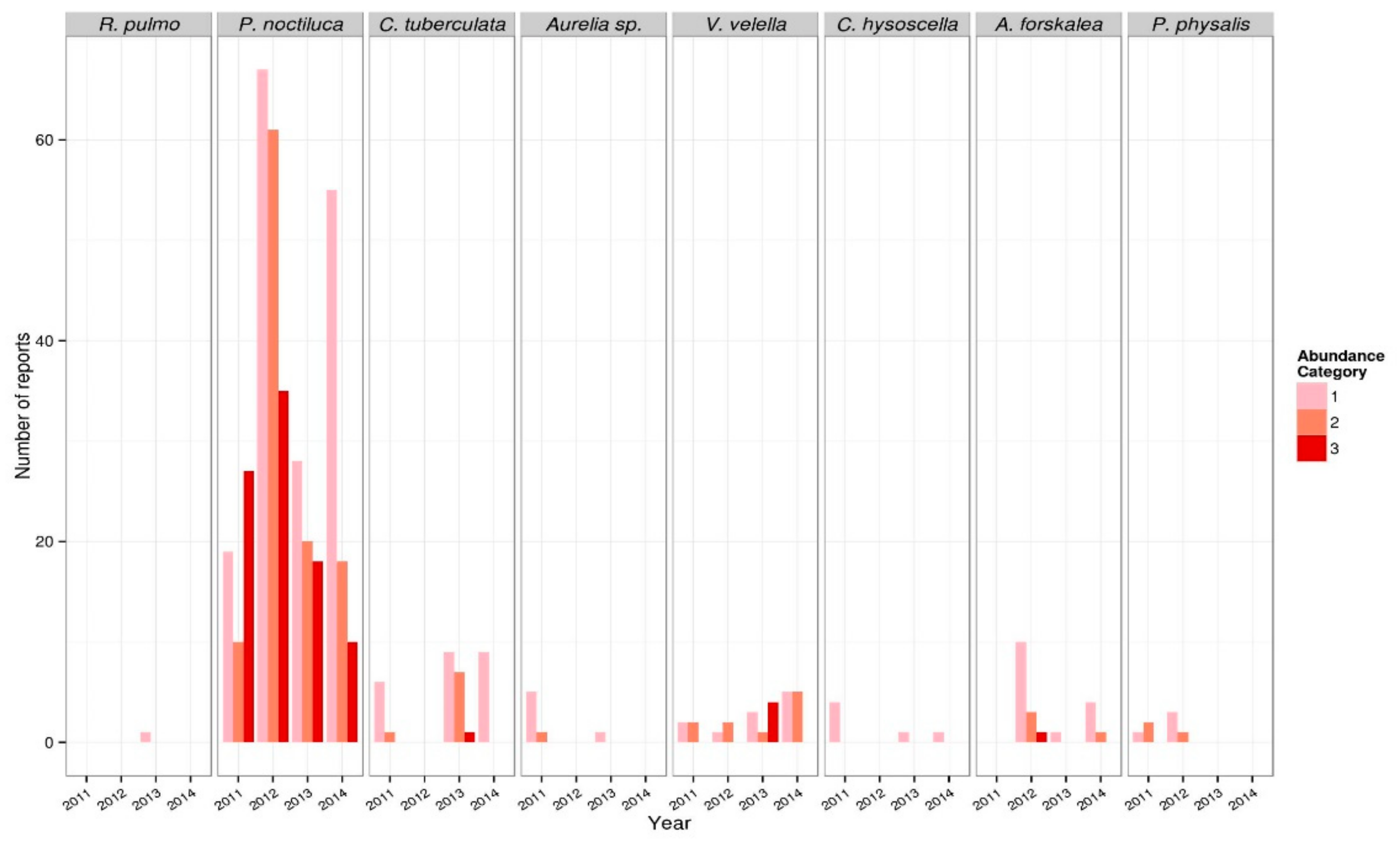
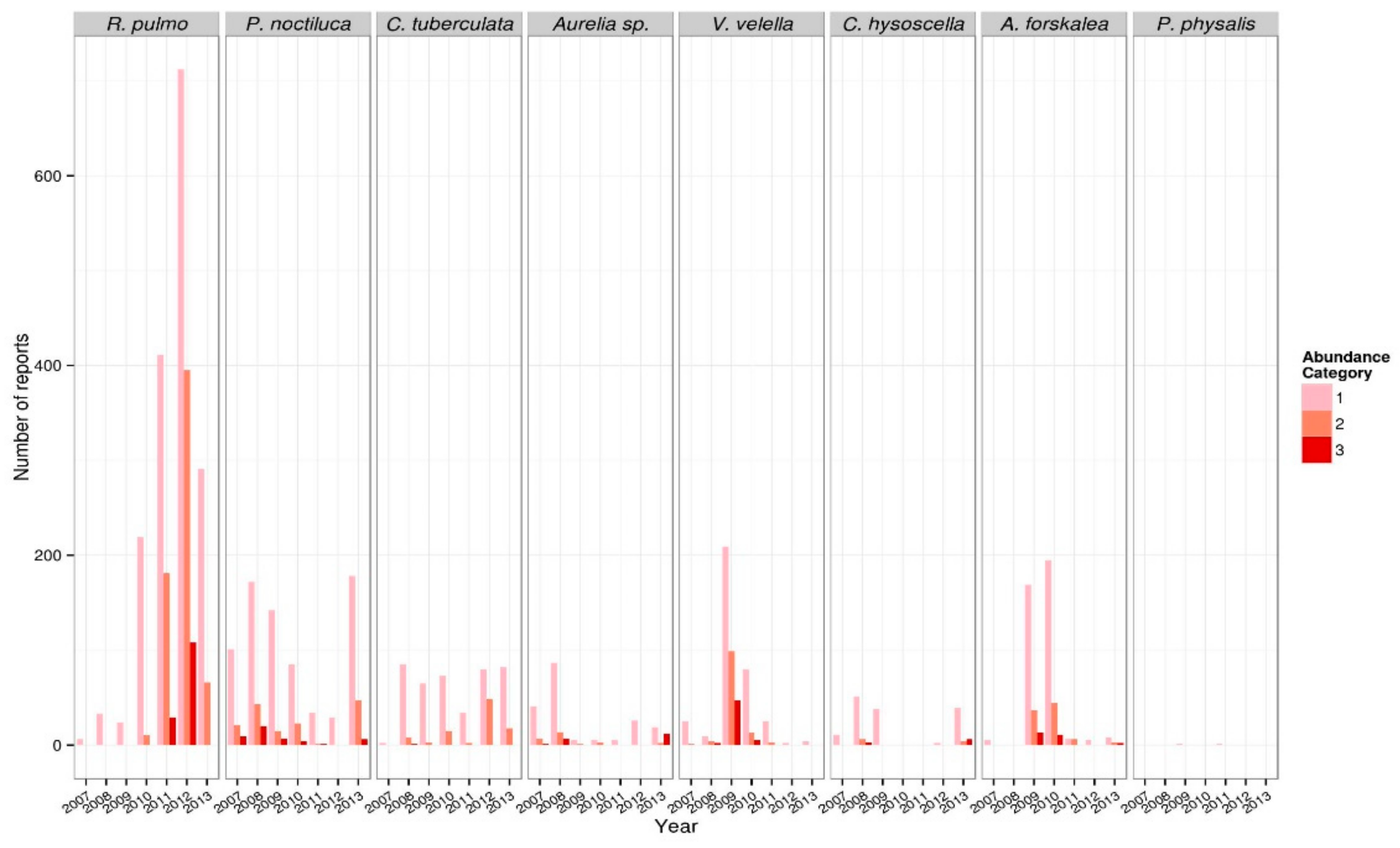
3.1.1. Jellyfish Temporal Dynamics
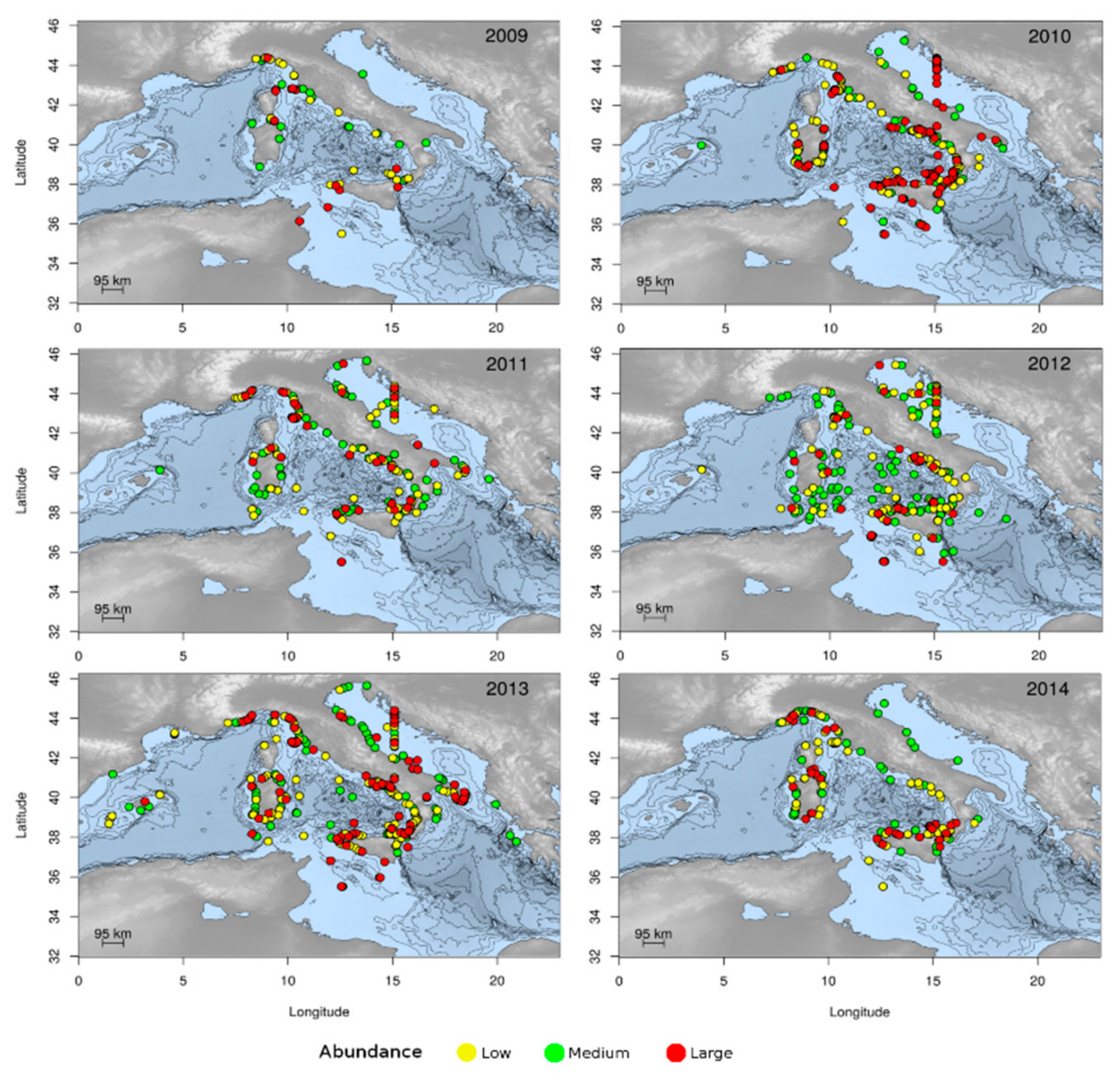
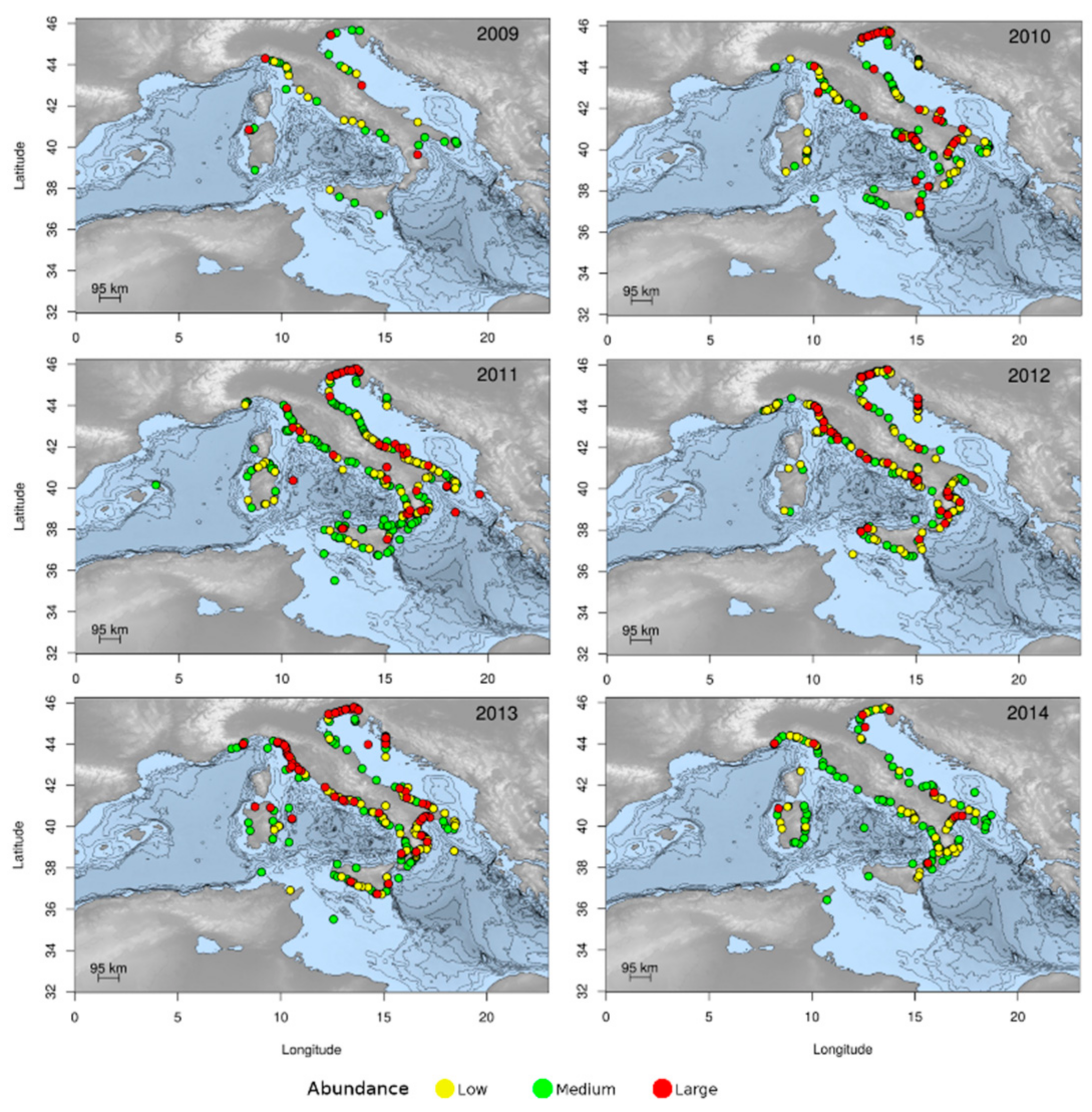
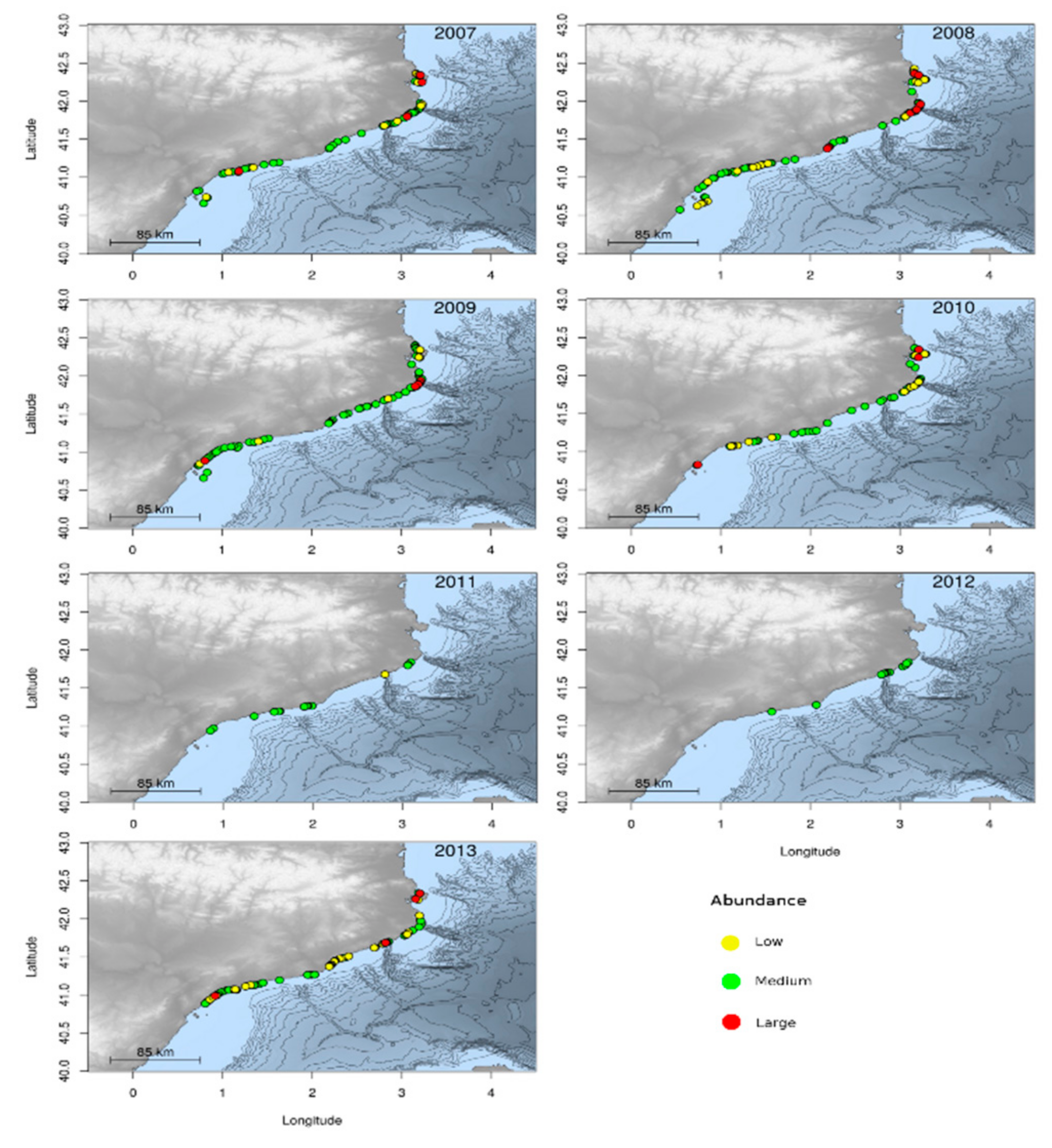
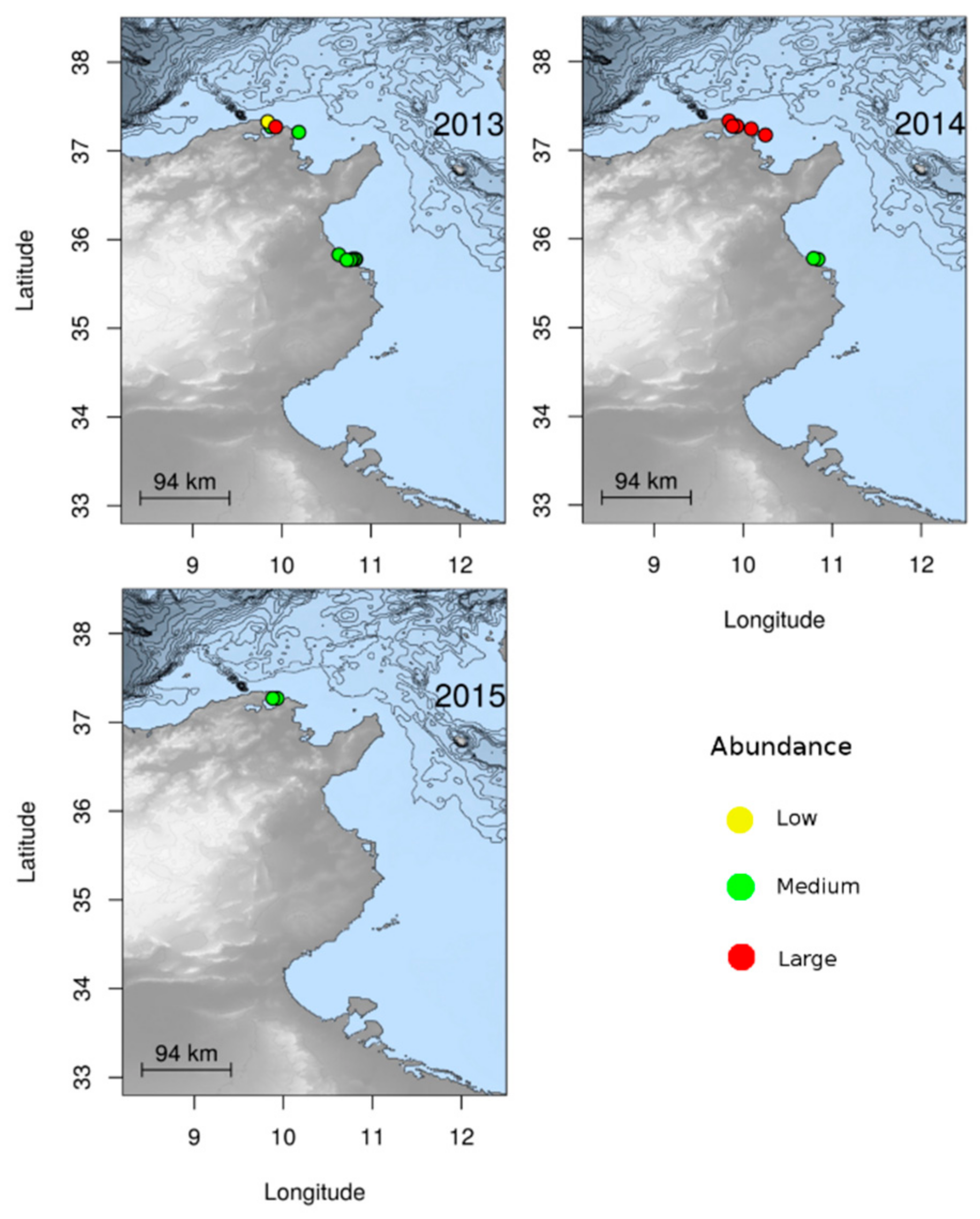
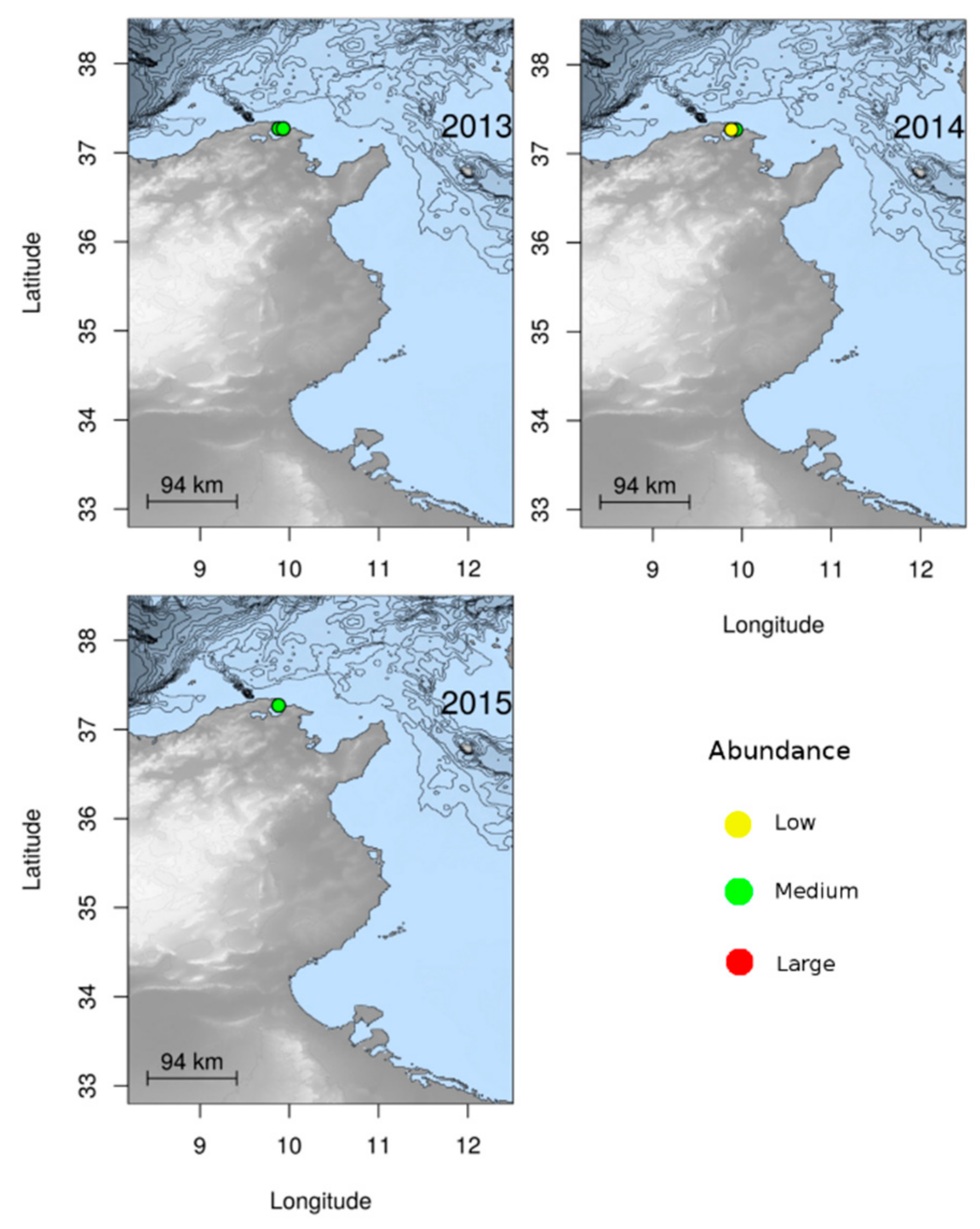
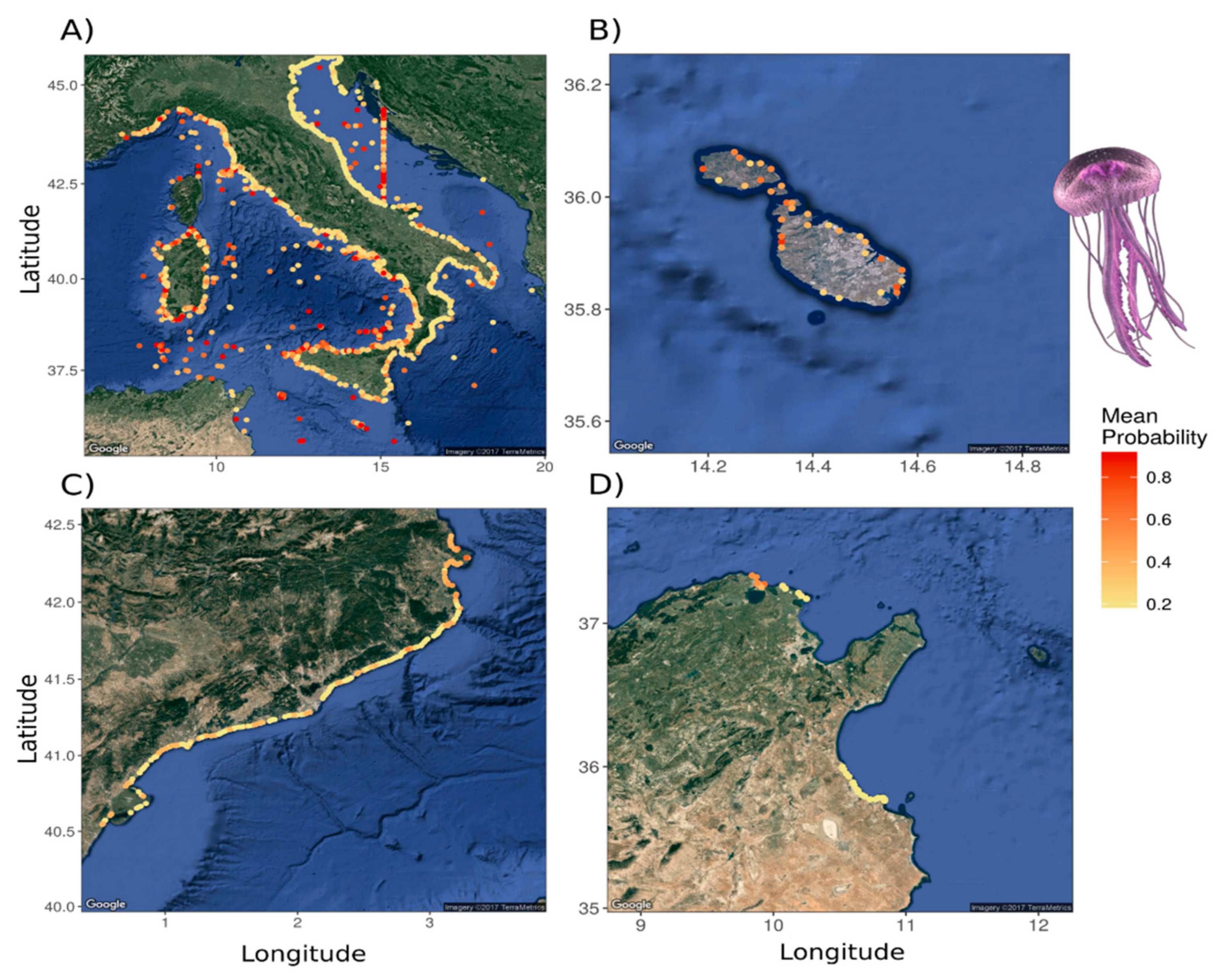
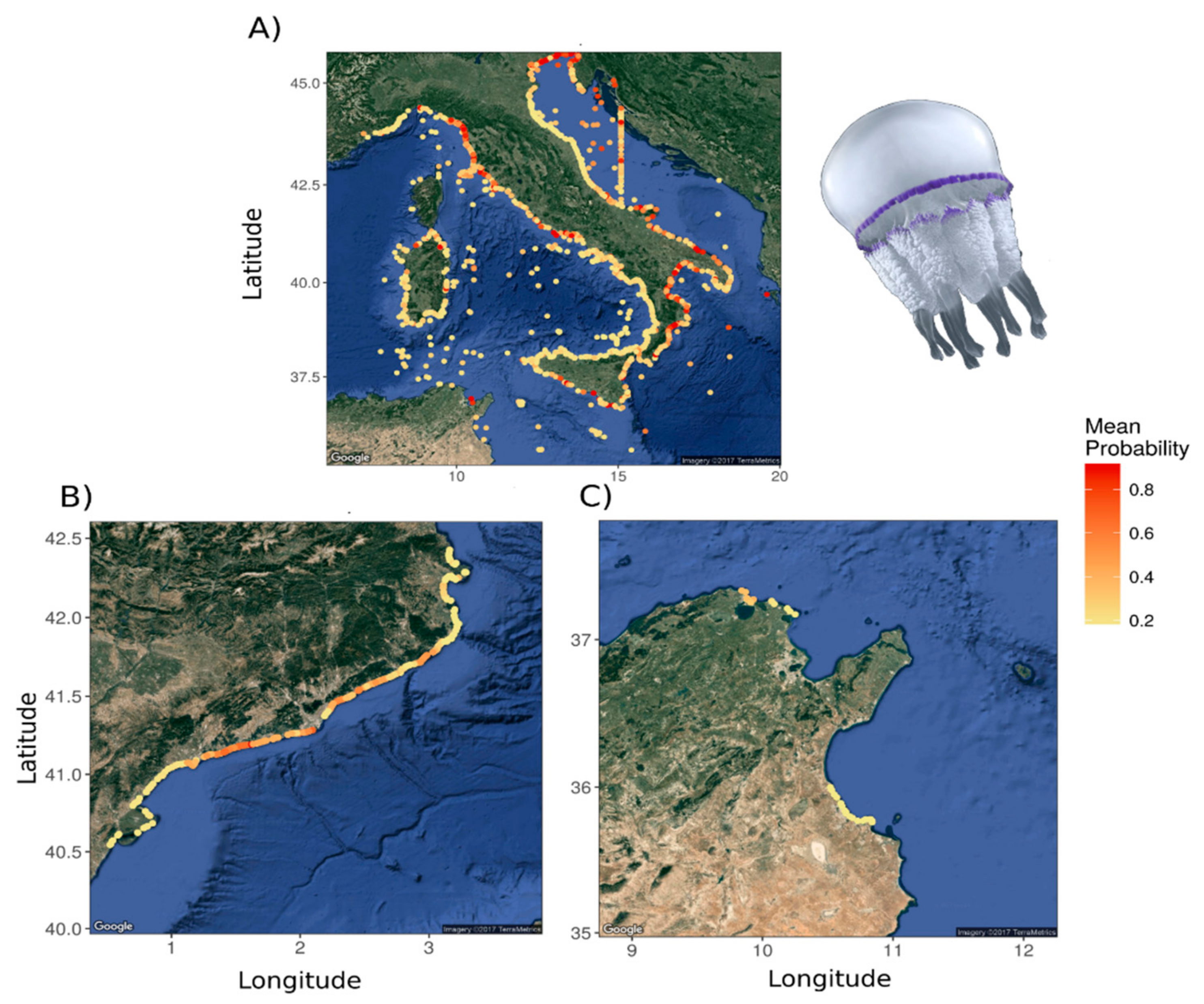
3.1.2. Jellyfish Spatial Dynamics
3.2. Analysis for Jellyfish Potential Spatial Distribution
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hagadorn, J.W.; Dott, R.H.; Damrow, D. Stranded on a Late Cambrian shoreline: Medusae from central Wisconsin. Geology 2002, 30, 147–150. [Google Scholar] [CrossRef]
- Mayoral, E.; Liñán, E.; Vintaned, J.A.G.; Muñiz, F.; Gozalo, R. Stranded jellyfish in the lowermost Cambrian (Corduban) of Spain. Span. J. Palaeontol. 2021, 19, 191–198. [Google Scholar] [CrossRef]
- Goy, J.; Morand, P.; Etienne, M. Long-term fluctuations of Pelagia noctiluca (Cnidaria, Scyphomedusa) in the western Mediterranean Sea. Prediction by climatic variables. Deep Sea Res. Part A Oceanogr. Res. Pap. 1989, 36, 269–279. [Google Scholar] [CrossRef]
- Condon, R.H.; Duarte, C.M.; Pitt, K.; Robinson, K.L.; Lucas, C.H.; Sutherland, K.R.; Mianzan, H.W.; Bogeberg, M.; Purcell, J.E.; Decker, M.B.; et al. Recurrent jellyfish blooms are a consequence of global oscillations. Proc. Natl. Acad. Sci. USA 2013, 110, 1000–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynam, C.; Hay, S.J.; Brierley, A.S. Interannual variability in abundance of North Sea jellyfish and links to the North Atlantic Oscillation. Limnol. Oceanogr. 2004, 49, 637–643. [Google Scholar] [CrossRef]
- Purcell, J.E. Climate effects on formation of jellyfish and ctenophore blooms: A review. J. Mar. Biol. Assoc. UK 2005, 85, 461–476. [Google Scholar] [CrossRef]
- Purcell, J.E. Jellyfish and Ctenophore Blooms Coincide with Human Proliferations and Environmental Perturbations. Annu. Rev. Mar. Sci. 2012, 4, 209–235. [Google Scholar] [CrossRef]
- Brodeur, R.D.; Decker, M.B.; Ciannelli, L.; Purcell, J.E.; Bond, N.A.; Stabeno, P.J.; Acuna, E.; Hunt, G.L. Rise and fall of jellyfish in the eastern Bering Sea in relation to climate regime shifts. Prog. Oceanogr. 2008, 77, 103–111. [Google Scholar] [CrossRef]
- Robinson, K.L.; Graham, W.M. Long-term change in the abundances of northern Gulf of Mexico scyphomedusae Chrysaora sp. and Aurelia spp. with links to climate variability. Limnol. Oceanogr. 2012, 58, 235–253. [Google Scholar] [CrossRef]
- Quiñones, J.; Mianzan, H.; Purca, S.; Robinson, K.L.; Adams, G.; Acha, E.M. Climate-driven population size fluctuations of jellyfish (Chrysaora plocamia) off Peru. Mar. Biol. 2015, 162, 2339–2350. [Google Scholar] [CrossRef]
- Graham, W.M.; Pagès, F.; Hamner, W.M. A physical context for gelatinous zooplankton aggregations: A review. Hydrobiologia 2001, 451, 199–212. [Google Scholar] [CrossRef]
- Purcell, J.E.; Uye, S.-I.; Lo, W.-T. Anthropogenic causes of jellyfish blooms and their direct consequences for humans: A review. Mar. Ecol. Prog. Ser. 2007, 350, 153–174. [Google Scholar] [CrossRef]
- Richardson, A.; Bakun, A.; Hays, G.; Gibbons, M. The jellyfish joyride: Causes, consequences and management responses to a more gelatinous future. Trends Ecol. Evol. 2009, 24, 312–322. [Google Scholar] [CrossRef]
- Graham, W.M.; Gelcich, S.; Robinson, K.L.; Duarte, C.M.; Brotz, L.; Purcell, J.E.; Madin, L.P.; Mianzan, H.; Sutherland, K.R.; Uye, S.-I.; et al. Linking human well-being and jellyfish: Ecosystem services, impacts, and societal responses. Front. Ecol. Environ. 2014, 12, 515–523. [Google Scholar] [CrossRef] [Green Version]
- Boero, F.; Brotz, L.; Gibbons, M.J.; Piraino, S.; Zampardi, S. Impacts and effects of ocean warming on jellyfish. In Explain Ocean Warm Causes, Scale, Eff Consequences; IUCN: Gland, Switzerland, 2016; pp. 213–237. [Google Scholar]
- Brotz, L.; Cheung, W.W.L.; Kleisner, K.; Pakhomov, E.; Pauly, D. Increasing jellyfish populations: Trends in Large Marine Ecosystems. Hydrobiologia 2012, 690, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Condon, R.; Graham, W.; Duarte, C.; Pitt, K.; Lucas, C.; Haddock, S.; Sutherland, K.; Robinson, K.; Dawson, M.; Decker, M.; et al. Questioning the rise of gelatinous zooplankton in the World’s oceans. BioScience 2012, 62, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Kogovšek, T.; Bogunović, B.; Malej, A. Recurrence of bloom-forming scyphomedusae: Wavelet analysis of a 200-year time series. Hydrobiologia 2010, 645, 81–96. [Google Scholar] [CrossRef]
- Licandro, P.; Conway, D.; Daly Yahia, M.; Fernández de Puelles, M.; Gasparini, S.; Hecq, J.H.; Tranter, P.; Kirby, R. A blooming jellyfish in the northeast Atlantic and Mediterranean. Biol. Lett. 2010, 6, 688–691. [Google Scholar] [CrossRef]
- Marambio, M.; López, L.; Fuentes, V.; Gili, J.M.; Canepa, A. More than 10 years dealing with jellyfish blooms: The jellyfish observation network in the NW Mediterranean. In 6th International Jellyfish Bloom Symposium; IZIKO South African Museum: Cape Town, South Africa, 2019. [Google Scholar]
- Hamner, W.M.; Madin, L.P.; Alldredge, A.L.; Gilmer, R.W.; Hamner, P.P. Underwater observations of gelatinous zooplankton: Sampling problems, feeding biology, and behavior1. Limnol. Oceanogr. 1975, 20, 907–917. [Google Scholar] [CrossRef]
- Purcell, J.E. Extension of methods for jellyfish and ctenophore trophic ecology to large-scale research. In Jellyfish Blooms: Causes, Consequences, and Recent Advances, Proceedings of the Second International Jellyfish Blooms Symposium, Gold Coast, Queensland, Australia, 24–27 June 2007; Pitt, K.A., Purcell, J.E., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 23–50. [Google Scholar]
- Doyle, T.K.; Houghton, J.D.R.; Buckley, S.M.; Hays, G.; Davenport, J. The broad-scale distribution of five jellyfish species across a temperate coastal environment. Hydrobiologia 2007, 579, 29–39. [Google Scholar] [CrossRef]
- Houghton, J.; Doyle, T.; Davenport, J.; Hays, G. Developing a simple, rapid method for identifying and monitoring jellyfish aggregations from the air. Mar. Ecol. Prog. Ser. 2006, 314, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Houghton, J.D.R.; Doyle, T.K.; Davenport, J.; Lilley, M.K.S.; Wilson, R.P.; Hays, G.C. Stranding events provide indirect insights into the seasonality and persistence of jellyfish medusae (Cnidaria: Scyphozoa). Hydrobiologia 2007, 589, 1–13. [Google Scholar] [CrossRef]
- Boero, F. Review of Jellyfish Blooms in the Mediterranean and Black Sea; GFCM Studies and Reviews; FAO: Rome, Italy, 2013; p. 92. [Google Scholar]
- Bosch-Belmar, M.; Azzurro, E.; Pulis, K.; Milisenda, G.; Fuentes, V.; Yahia, O.K.-D.; Micallef, A.; Deidun, A.; Piraino, S. Jellyfish blooms perception in Mediterranean finfish aquaculture. Mar. Policy 2017, 76, 1–7. [Google Scholar] [CrossRef]
- Bernard, P.; Berline, L.; Gorsky, G. Long term (1981–2008) monitoring of the jellyfish Pelagia noctiluca (Cnidaria, Scyphozoa) on Mediterranean Coasts (Principality of Monaco and French Riviera). J. Oceanogr. Res. Data 2011, 4, 1–10. [Google Scholar]
- Baumann, S.; Schernewski, G. Occurrence and public perception of jellyfish along the German Baltic coastline. J. Coast. Conserv. 2012, 16, 555–566. [Google Scholar] [CrossRef]
- Canepa, A.; Fuentes, V.; Sabatés, A.; Piraino, S.; Boero, F.; Gili, J.-M. Pelagia noctiluca in the Mediterranean Sea. In Jellyfish Blooms; Springer: Dordretch, The Netherlands, 2013; pp. 237–266. [Google Scholar]
- Purcell, J.E.; Milisenda, G.; Rizzo, A.; Carrion, S.A.; Zampardi, S.; Airoldi, S.; Zagami, G.; Guglielmo, L.; Boero, F.; Doyle, T.K.; et al. Digestion and predation rates of zooplankton by the pleustonic hydrozoan Velella velella and widespread blooms in 2013 and 2014. J. Plankton Res. 2015, 37, 1056–1067. [Google Scholar] [CrossRef] [Green Version]
- Edelist, D.; Guy-Haim, T.; Kuplik, Z.; Zuckerman, N.; Nemoy, P.; Angel, D.L. Phenological shift in swarming patterns of Rhopilema nomadica in the Eastern Mediterranean Sea. J. Plankton Res. 2020, 42, 211–219. [Google Scholar] [CrossRef]
- Aceves-Bueno, E.; Adeleye, A.S.; Bradley, D.; Tyler Brandt, W.; Callery, P.; Feraud, M.; Garner, K.L.; Gentry, R.; Huang, Y.; McCullough, I.; et al. Citizen Science as an Approach for Overcoming Insufficient Monitoring and Inadequate Stakeholder Buy-in in Adaptive Management: Criteria and Evidence. Ecosystems 2015, 18, 493–506. [Google Scholar] [CrossRef]
- Dickinson, J.L.; Zuckerberg, B.; Bonter, D.N. Citizen Science as an Ecological Research Tool: Challenges and Benefits. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 149–172. [Google Scholar] [CrossRef] [Green Version]
- Miller-Rushing, A.; Primack, R.B.; Bonney, R. The history of public participation in ecological research. Front. Ecol. Environ. 2012, 10, 285–290. [Google Scholar] [CrossRef]
- Chandler, M.; See, L.; Copas, K.; Bonde, A.M.; López, B.C.; Danielsen, F.; Legind, J.K.; Masinde, S.; Miller-Rushing, A.J.; Newman, G.; et al. Contribution of citizen science towards international biodiversity monitoring. Biol. Conserv. 2017, 213, 280–294. [Google Scholar] [CrossRef] [Green Version]
- Delaney, D.G.; Sperling, C.D.; Adams, C.S.; Leung, B. Marine invasive species: Validation of citizen science and implications for national monitoring networks. Biol. Invasions 2007, 10, 117–128. [Google Scholar] [CrossRef]
- Theobald, E.; Ettinger, A.; Burgess, H.; DeBey, L.; Schmidt, N.; Froehlich, H.; Wagner, C.; HilleRisLambers, J.; Tewksbury, J.; Harsch, M.; et al. Global change and local solutions: Tapping the unrealized potential of citizen science for biodiversity research. Biol. Conserv. 2015, 181, 236–244. [Google Scholar] [CrossRef] [Green Version]
- Purcell, J.E.; Arai, M.N. Interactions of pelagic cnidarians and ctenophores with fish: A review. Hydrobiologia 2001, 451, 27–44. [Google Scholar] [CrossRef]
- Lynam, C.; Gibbons, M.; Axelsen, B.E.; Sparks, C.; Coetzee, J.; Heywood, B.; Brierley, A. Jellyfish overtake fish in a heavily fished ecosystem. Curr. Biol. 2006, 16. [Google Scholar] [CrossRef] [Green Version]
- Utne-Palm, A.C.; Salvanes, A.G.V.; Currie, B.; Kaartvedt, S.; Nilsson, G.E.; Braithwaite, V.A.; Stecyk, J.A.W.; Hundt, M.; Van Der Bank, M.; Flynn, B.; et al. Trophic Structure and Community Stability in an Overfished Ecosystem. Science 2010, 329, 333–336. [Google Scholar] [CrossRef] [Green Version]
- Brodeur, R.D.; Link, J.; Smith, B.E.; Ford, M.D.; Kobayashi, D.R.; Jones, T.T. Ecological and Economic Consequences of Ignoring Jellyfish: A Plea for Increased Monitoring of Ecosystems. Fisheries 2016, 41, 630–637. [Google Scholar] [CrossRef]
- Bosch-Belmar, M.; Milisenda, G.; Girons, A.; Taurisano, V.; Accoroni, S.; Totti, C.; Piraino, S.; Fuentes, V. Consequences of Stinging Plankton Blooms on Finfish Mariculture in the Mediterranean Sea. Front. Mar. Sci. 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Bosch-Belmar, M.; Milisenda, G.; Basso, L.; Doyle, T.K.; Leone, A.; Piraino, S. Jellyfish Impacts on Marine Aquaculture and Fisheries. Rev. Fish. Sci. Aquac. 2021, 29, 242–259. [Google Scholar] [CrossRef]
- Graham, W.M.; Martin, D.L.; Felder, D.L.; Asper, V.L.; Perry, H.M. Ecological and economic implications of a tropical jellyfish invader in the Gulf of Mexico. Biol. Invasions 2003, 5, 53–69. [Google Scholar] [CrossRef]
- Kawahara, M.; Uye, S.-I.; Ohtsu, K.; Iizumi, H. Unusual population explosion of the giant jellyfish Nemopilema nomurai (Scyphozoa: Rhizostomeae) in East Asian waters. Mar. Ecol. Prog. Ser. 2006, 307, 161–173. [Google Scholar] [CrossRef]
- Gershwin, L.-A.; Denardi, D.; Fenner, P.; Winkel, K. Marine stingers: Review of an under-recognised global coastal management issue. Coast. Manag. J. 2009, 38, 22–41. [Google Scholar] [CrossRef]
- Palmieri, M.; Barausse, A.; Luisetti, T.; Turner, R. Jellyfish blooms in the Northern Adriatic Sea: Fishermen’s perceptions and economic impacts on fisheries. Fish. Res. 2014, 155, 51–58. [Google Scholar] [CrossRef]
- Nunes, P.A.L.D.; Loureiro, M.L.; Piñol, L.; Sastre, S.; Voltaire, L.; Canepa, A. Analyzing Beach Recreationists’ Preferences for the Reduction of Jellyfish Blooms: Economic Results from a Stated-Choice Experiment in Catalonia, Spain. PLoS ONE 2015, 10, e0126681. [Google Scholar] [CrossRef] [Green Version]
- Kingsford, M.J.; Becken, S.; Bordehore, C.; Fuentes, V.L.; Pitt, K.A.; Yangihara, A.A. Empowering Stakeholders to Manage Stinging Jellyfish: A Perspective. Coast. Manag. 2017, 46, 1–18. [Google Scholar] [CrossRef]
- Decker, M.; Brown, C.W.; Hood, R.; Purcell, J.; Gross, T.; Matanoski, J.; Bannon, R.; Setzler-Hamilton, E. Predicting the distribution of the scyphomedusa Chrysaora quinquecirrha in Chesapeake Bay. Mar. Ecol. Prog. Ser. 2007, 329, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Lucas, C.H.; Gelcich, S.; Uye, S.-I. Living with Jellyfish: Management and Adaptation Strategies. In Jellyfish Blooms; Springer: Dordretch, The Netherlands, 2014; pp. 129–150. [Google Scholar]
- Uye, S. The Giant Jellyfish Nemopilema nomurai in East Asian Marginal Seas. In Jellyfish Blooms; Pitt, K.A., Lucas, C.H., Eds.; Springer: Dordretch, The Netherlands, 2014; pp. 185–205. [Google Scholar]
- Pontin, D.R.; Watts, M.J.; Worner, S.P. Using Multi-Layer Perceptrons to predict the presence of jellyfish of the genus Physalia at New Zealand beaches. In Proceedings of the International Joint Conference on Neural Networks, IJCNN 2008, Part of the IEEE World Congress on Computational Intelligence, WCCI 2008, Hong Kong, China, 1–6 June 2008; pp. 1170–1175. [Google Scholar]
- Record, N.; Tupper, B.; Pershing, A. The jelly report: Forecasting jellyfish using email and social media. Anthr. Coasts 2018, 1, 34–43. [Google Scholar]
- Canepa, A.; Purcell, J.E.; Córdova, P.; Fernández, M.; Palma, S. Massive strandings of pleustonic Portuguese Man-of-War (Physalia physalis) related to ENSO events along the southeastern Pacific Ocean. Lat. Am. J. Aquat. Res. 2020, 48, 806–817. [Google Scholar] [CrossRef]
- Benedetti-Cecchi, L.; Canepa, A.; Fuentes, V.; Tamburello, L.; Purcell, J.E.; Piraino, S.; Roberts, J.; Boero, F.; Halpin, P. Deterministic Factors Overwhelm Stochastic Environmental Fluctuations as Drivers of Jellyfish Outbreaks. PLoS ONE 2015, 10, e0141060. [Google Scholar] [CrossRef] [PubMed]
- Pante, E.; Simon-Bouhet, B. marmap: A Package for Importing, Plotting and Analyzing Bathymetric and Topographic Data in R. PLoS ONE 2013, 8, e73051. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems: Data exploration. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Austin, M. Species distribution models and ecological theory: A critical assessment and some possible new approaches. Ecol. Model. 2007, 200, 1–19. [Google Scholar] [CrossRef]
- Austin, M.P.; Van Niel, K.P. Improving species distribution models for climate change studies: Variable selection and scale. J. Biogeogr. 2010, 38, 1–8. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F. Biomod2: Ensemble Platform for Species Distribution Modeling; R Package Version 3.4.6. 2020. Available online: https://CRAN.R-project.org/package=biomod2 (accessed on 1 April 2021).
- Wood, S. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman & Hall: London, UK, 2017; p. 496. [Google Scholar]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.; Huettmann, F.; Leathwick, J.; Leh-mann, A.; et al. Novel methods improve prediction of species’ distributions from occurence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- McCulloch, W.S.; Pitts, W. A logical calculus of the ideas immanent in nervous activity. Bull. Math. Biol. 1943, 5, 115–133. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S. A Review of Machine Learning Based Species’ Distribution Modelling. In Proceedings of the 2017 International Conference on Industrial Informatics—Computing Technology, Intelligent Technology, Industrial Information Integration, Wuhan, China, 2–3 December 2017. [Google Scholar]
- Scorrano, S.; Aglieri, G.; Boero, F.; Dawson, M.N.; Piraino, S. Unmasking Aurelia species in the Mediterranean Sea: An integrative morphometric and molecular approach. Zool. J. Linn. Soc. 2017, 180, 243–267. [Google Scholar]
- Gueroun, S.K.; Molinero, J.C.; Piraino, S.; Yahia, M.N.D. Population dynamics and predatory impact of the alien jellyfish Aurelia solida (Cnidaria, Scyphozoa) in the Bizerte Lagoon (southwestern Mediterranean Sea). Mediterr. Mar. Sci. 2020, 21, 22–35. [Google Scholar] [CrossRef]
- Devictor, V.; Whittaker, R.J.; Beltrame, C. Beyond scarcity: Citizen science programmes as useful tools for conservation biogeography. Divers. Distrib. 2010, 16, 354–362. [Google Scholar] [CrossRef]
- McKinley, D.C.; Miller-Rushing, A.J.; Ballard, H.L.; Bonney, R.; Brown, H.; Cook-Patton, S.C.; Evans, D.M.; French, R.A.; Parrish, J.K.; Phillips, T.B.; et al. Citizen science can improve conservation science, natural resource management, and environmental protection. Biol. Conserv. 2017, 208, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Turrini, T.; Dörler, D.; Richter, A.; Heigl, F.; Bonn, A. The threefold potential of environmental citizen science-Generating knowledge, creating learning opportunities and enabling civic participation. Biol. Conserv. 2018, 225, 176–186. [Google Scholar] [CrossRef]
- Bonney, R.; Cooper, C.B.; Dickinson, J.; Kelling, S.; Phillips, T.; Rosenberg, K.V.; Shirk, J. Citizen Science: A Developing Tool for Expanding Science Knowledge and Scientific Literacy. BioScience 2009, 59, 977–984. [Google Scholar] [CrossRef]
- Cigliano, J.A.; Meyer, R.; Ballard, H.L.; Freitag, A.; Phillips, T.B.; Wasser, A. Making marine and coastal citizen science matter. Ocean Coast. Manag. 2015, 115, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Bonter, D.N.; Cooper, C.B. Data validation in citizen science: A case study from Project FeederWatch. Front. Ecol. Environ. 2012, 10, 305–307. [Google Scholar] [CrossRef]
- Crall, A.; Greg, N.; Stohlgren, T.; Holfelder, K.; Graham, J.; Waller, D. Assessing citizen science data quality: An invasive species case study. Conserv. Lett. 2011, 4, 433–442. [Google Scholar] [CrossRef]
- Freitag, A.; Meyer, R.; Whiteman, L. Strategies Employed by Citizen Science Programs to Increase the Credibility of Their Data. Citiz. Sci. Theory Pract. 2016, 1, 2. [Google Scholar] [CrossRef]
- Angel, D.L.; Edelist, D.; Freeman, S. Local perspectives on regional challenges: Jellyfish proliferation and fish stock management along the Israeli Mediterranean coast. Reg. Environ. Chang. 2014, 16, 315–323. [Google Scholar] [CrossRef]
- Boero, F.; Bouillon, J.; Gravili, C.; Miglietta, M.; Parsons, T.; Piraino, S. Gelatinous plankton: Irregularities rule the world (some-times). Mar. Ecol. Prog. Ser. 2008, 356, 299–310. [Google Scholar] [CrossRef]
- Brotz, L.; Pauly, D. Jellyfish populations in the Mediterranean Sea. Acta Adriat. 2012, 53, 211–230. [Google Scholar]
- Mariottini, G.; Giacco, E.; Pane, L. The Mauve Stinger Pelagia noctiluca (Forsskål, 1775). Distribution, Ecology, Toxicity and Epidemiology of Stings. A Review. Mar. Drugs 2008, 6, 496–513. [Google Scholar] [CrossRef]
- Rusell, F.S. The Medusae of the British Isles II. Pelagic Svyphozoa with a Supplement to the First Volume on Hydromedusae; Cambridge University Press: Cambridge, UK, 1970; p. 284. [Google Scholar]
- UNEP. UNEP: Report on the Workshop on Jellyfish Blooms in the Mediterranean. In Proceedings of the Rapport de Journees D’etude sur les Proliferations Anormales de Meduses en Mediterranee, Athens, Greece, 31 October–4 November 1984; p. 221. [Google Scholar]
- UNEP. Jellyfish blooms in the Mediterranean. In Proceedings of the II Workshop on jellyfish in the Mediterranean Sea, Athens, Greece, 2–5 September 1991. [Google Scholar]
- Mariottini, G.L.; Pane, L. Mediterranean Jellyfish Venoms: A Review on Scyphomedusae. Mar. Drugs 2010, 8, 1122–1152. [Google Scholar] [CrossRef] [Green Version]
- Gili, J.M.; Pagès, F. Les Proliferacions de Meduses; Bolletí de la Societat d’Història Natural de les Balears: Palma, Illes Balears, Spain, 2005; pp. 9–22. Available online: https://raco.cat/index.php/BolletiSHNBalears/article/view/170002 (accessed on 1 June 2021).
- Vučetić, T. Some causes of the blooms and unusual distribution of the jellyfish Pelagia noctiluca in the Mediterranean (Adriatic). In Proceedings of the UNEP: Report on the Workshop on Jellyfish Blooms in the Mediterranean, Athens, Greece, 31 October–4 November 1984; pp. 167–176. [Google Scholar]
- Rottini-Sandrini, L. Preliminary report on the occurrence of Pelagia noctiluca (Semaeostomeae, Pelagiidae) in Northern Adriatic. Rapp. Comm. Int. Mer Médit. 1981, 27, 2. [Google Scholar]
- Maretić, Z. The bloom of jellyfish Pelagia noctiluca along the coasts of Pula and Istria 1977–1983, with special reference to epidemiology, clinics and treatment. In Proceedings of the UNEP: Report on the Workshop on Jellyfish Blooms in the Mediterranean, Athens, Greece, 31 October–4 November 1984; pp. 83–88. [Google Scholar]
- Malej, A. Some data on the occurrence and biology of the scyphomedusa Pelagia noctiluca in the Gulf of Trieste, and the impact of jellyfish swarming on human activities. In Proceedings of the UNEP: Report on the Workshop on Jellyfish Blooms in the Mediterranean, Athens, Greece, 31 October–4 November 1984. [Google Scholar]
- Rosa, S.; Pansera, M.; Granata, A.; Guglielmo, L. Interannual variability, growth, reproduction and feeding of Pelagia noctiluca (Cnidaria: Scyphozoa) in the Straits of Messina (Central Mediterranean Sea): Linkages with temperature and diet. J. Mar. Syst. 2013, 111–112, 97–107. [Google Scholar] [CrossRef]
- Leoni, V.; Bonnet, D.; Ramírez-Romero, E.; Molinero, J.C. Biogeography and phenology of the jellyfish Rhizostoma pulmo (Cnidaria: Scyphozoa) in southern European seas. Glob. Ecol. Biogeogr. 2021, 30, 622–639. [Google Scholar] [CrossRef]
- Mariottini, G.L.; Giacco, E.; Pane, L. Distribution de Pelagia noctiluca (Forskal) en Méditerranée dans l’été 1983. In Proceedings of the UNEP: Report on the Workshop on Jellyfish Blooms in the Mediterranean, Athens, Greece, 31 October–4 November 1984. [Google Scholar]
- Stravisi, F. Meteoclimatic influence on the blooms of Pelagia noctiluca in the northern Adriatic. In Proceedings of the UNEP: Report on the Workshop on Jellyfish Blooms in the Mediterranean, Athens, Greece, 31 October–4 November 1984. [Google Scholar]
- Zavodnik, D. Spatial aggregations of the swarming jellyfish Pelagia noctiluca (Scyphozoa). Mar. Biol. 1987, 94, 265–269. [Google Scholar] [CrossRef]
- Lilley, M.; Houghton, J.; Hays, G. Distribution, extent of inter-annual variability and diet of the bloom-forming jellyfish Rhizostoma in European waters. J. Mar. Biol. Assoc. UK 2008, 89, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Axiak, V.; Galea, C.; Schembri, P. Coastal aggregations of the jellyfish Pelagia noctiluca (Scyphozoa) in Maltese coastal waters during 1980–1986. Mediterr. Action Plan Tech. Rep. Ser. 1991, 47, 32–40. [Google Scholar]
- Gatt, M.P.; Deidun, A.; Galea, A.; Gauci, A. Is Citizen Science a Valid Tool to Monitor the Occurrence of Jellyfish? The Spot the Jellyfish Case Study from the Maltese Islands. J. Coast. Res. 2018, 85, 316–320. [Google Scholar] [CrossRef]
- Benovic, A. The aspect of jellyfish distribution in the Adriatic Sea. UNEP: Jellyfish blooms in the Mediterranean. In Proceedings of the II workshop on jellyfish in the Mediterranean Sea MAP Tech Rep Ser, No 47, Athens, Greece, 2–5 September 1991. [Google Scholar]
- Zavodnik, D. On the food and feeding in the northern Adriatic of Pelagia noctiluca (Scyphozoa). UNEP: Jellyfish blooms in the Mediterranean. In Proceedings of the II workshop on jellyfish in the Mediterranean Sea MAP Tech Rep Ser, No 47, Athens, Greece, 2–5 September 1991; pp. 212–216. [Google Scholar]
- Fuentes, V.; Straehler-Pohl, I.; Atienza, D.; Franco, I.; Tilves, U.; Gentile, M.; Acevedo, M.; Olariaga, A.; Gili, J.-M. Life cycle of the jellyfish Rhizostoma pulmo (Scyphozoa: Rhizostomeae) and its distribution, seasonality and inter-annual variability along the Catalan coast and the Mar Menor (Spain, NW Mediterranean). Mar. Biol. 2011, 158, 2247–2266. [Google Scholar] [CrossRef]
- Daly Yahia, M. Distribution and ecology of Medusae and Scyphomedusae (Cnidaria) in Tunis Gulf (SW Mediterranean). Oceanol. Acta 2003, 26, 645–655. [Google Scholar] [CrossRef] [Green Version]
- Addad, S.; Yahia, M.; Belghith, H.; Zamamouch, S.; Goy, J.; Yahia-KÈfi, O.D. Analyse taxinomique, écologique et caractérisation biochimique des scyphoméduses de la baie de Bizerte et de Tunis (Juillet-D 2004). Bull. Soc. Zool. Franc. 2008, 133, 33–41. [Google Scholar]
- Touzri, C.; Kefi-Daly Yahia, O.; Hamdi, H.; Goy, J.; Daly Yahia, M. Spatio-temporal distribution of Medusae (Cnidaria) in the Bay of Bizerte (South Western Mediterranean Sea). Cah. Biol. Mar. 2010, 51, 167–176. [Google Scholar]
- Touzri, C.; Hamdi, H.; Goy, J.; Daly Yahia, M. Diversity and distribution of gelatinous zooplankton in the South-western Mediterranean Sea. Mar. Ecol. 2012, 33. [Google Scholar] [CrossRef]
- Touzri, C.; Daly Yahia, M.; Goy, J. Gelatinous plankton of Sousse bay: Systematics and ecology. Bull. Soc. Zool. Franc. 2004, 129, 379–392. [Google Scholar]
- Molinero, J.C.; Ibanez, F.; Nival, P.; Buecher, E.; Souissi, S. North Atlantic climate and northwestern Mediterranean plankton variability. Limnol. Oceanogr. 2005, 50, 1213–1220. [Google Scholar] [CrossRef]
- Leone, A.; Lecci, R.M.; Durante, M.; Meli, F.; Piraino, S. The Bright Side of Gelatinous Blooms: Nutraceutical Value and Antioxidant Properties of Three Mediterranean Jellyfish (Scyphozoa). Mar. Drugs 2015, 13, 4654–4681. [Google Scholar] [CrossRef] [Green Version]
- De Domenico, S.; De Rinaldis, G.; Paulmery, M.; Piraino, S.; Leone, A. Barrel Jellyfish (Rhizostoma pulmo) as Source of Antioxidant Peptides. Mar. Drugs 2019, 17, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Donno, A.; Idolo, A.; Bagordo, F.; Grassi, T.; Leomanni, A.; Serio, F.; Guido, M.; Canitano, M.; Zampardi, S.; Boero, F.; et al. Impact of Stinging Jellyfish Proliferations along South Italian Coasts: Human Health Hazards, Treatment and Social Costs. Int. J. Environ. Res. Public Health 2014, 11, 2488–2503. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Year | NDS | PPR | Intensity Index | NBS | NPR | TR |
|---|---|---|---|---|---|---|---|
| Italy | 2009 | 117 | 100 | 0.51 | 197 | 303 | 303 |
| 2010 | 201 | 100 | 0.12 | 811 | 1818 | 1818 | |
| 2011 | 191 | 100 | 0.14 | 720 | 3738 | 3738 | |
| 2012 | 217 | 100 | 0.04 | 2387 | 2671 | 2671 | |
| 2013 | 214 | 100 | 0.09 | 1130 | 5422 | 5422 | |
| 2014 | 227 | 100 | 0.18 | 542 | 1779 | 1779 | |
| Malta | 2011 | 72 | 100 | 3.13 | 32 | 124 | 124 |
| 2012 | 128 | 100 | 2.94 | 34 | 242 | 242 | |
| 2013 | 84 | 100 | 3.57 | 28 | 122 | 122 | |
| 2014 | 75 | 100 | 3.03 | 33 | 134 | 134 | |
| Spain | 2007 | 101 | 1.3 | 0.01 | 243 | 204 | 15,762 |
| 2008 | 101 | 2.9 | 0.01 | 243 | 456 | 15,729 | |
| 2009 | 107 | 4.9 | 0.02 | 242 | 773 | 15,797 | |
| 2010 | 108 | 4.2 | 0.02 | 243 | 709 | 16,907 | |
| 2011 | 123 | 30.2 | 0.38 | 79 | 1509 | 4995 | |
| 2012 | 103 | 43.5 | 0.43 | 101 | 2982 | 6863 | |
| 2013 | 134 | 5.1 | 0.03 | 197 | 1135 | 22,320 | |
| Tunisia | 2013 | 50 | 16.5 | 0.50 | 33 | 66 | 400 |
| 2014 | 46 | 25.5 | 0.80 | 32 | 61 | 239 | |
| 2015 | 1 | -- * | -- * | 2 | 2 | 2 | |
| TOTAL | -- | 2400 | 7329 | 115,367 | 7329 | 24,250 | 115,367 |
| Country | Environmental Variable | GAM | ANN | RF | All Models (Average) |
|---|---|---|---|---|---|
| Italy | Sea surface temperature (°C) | 0.08 | 0.09 | 0.35 | 0.17 |
| Salinity | 0.62 | 0.24 | 0.45 | 0.44 | |
| Current direction (East, m s−1) | 0.08 | 0.00 | 0.08 | 0.05 | |
| Current direction (North, m s−1) | 0.06 | 0.00 | 0.10 | 0.05 | |
| Current speed (m s−1) | 0.08 | 0.05 | 0.12 | 0.08 | |
| Slope index | 0.29 | 0.93 | 0.66 | 0.62 | |
| Malta | Sea surface temperature °C) | 0.13 | 0.93 | 0.23 | 0.43 |
| Salinity | 0.09 | 0.01 | 0.15 | 0.08 | |
| Phosphate (mmol m−3) | 0.50 | 0.00 | 0.02 | 0.17 | |
| Nitrate (mmol m−3) | 0.07 | 0.00 | 0.11 | 0.06 | |
| Current direction (East, m s−1) | 0.02 | 0.01 | 0.11 | 0.05 | |
| Current direction (North, m s−1) | 0.09 | 0.00 | 0.08 | 0.06 | |
| Slope index | 0.02 | 0.36 | 0.09 | 0.15 | |
| Spain | Sea surface temperature (°C) | 0.39 | 0.78 | 0.63 | 0.60 |
| Salinity | 0.17 | 0.07 | 0.60 | 0.28 | |
| Chlorophyll a (mg m−3) | 0.04 | 0.07 | 0.34 | 0.15 | |
| Wind direction (East, m s−1) | 0.14 | 0.51 | 0.41 | 0.35 | |
| Wind direction (North, m s−1) | 0.19 | 0.62 | 0.59 | 0.47 | |
| Current direction (East, m s−1) | 0.36 | 0.01 | 0.35 | 0.24 | |
| Current direction (North, m s−1) | 0.26 | 0.01 | 0.40 | 0.22 | |
| Current speed (m s−1) | 0.51 | 0.00 | 0.36 | 0.29 | |
| Slope index | 0.18 | 0.55 | 0.47 | 0.40 | |
| Tunisia | Sea surface temperature (°C) | 0.54 | 0.28 | 0.42 | 0.41 |
| Chlorophyll a (mg m−3) | 0.23 | 0.00 | 0.26 | 0.16 | |
| Phosphate (mmol m−3) | 0.25 | 0.00 | 0.02 | 0.09 | |
| Nitrate (mmol m−3) | 0.48 | 0.15 | 0.13 | 0.25 | |
| Wind direction (East, m s−1) | 0.25 | 0.43 | 0.05 | 0.24 | |
| Wind direction (North, m s−1) | 0.25 | 0.26 | 0.11 | 0.21 | |
| Wind speed (m s−1) | 0.23 | 0.20 | 0.06 | 0.17 | |
| Current direction (East, m s−1) | 0.10 | 0.00 | 0.02 | 0.04 | |
| Current direction (North, m s−1) | 0.27 | 0.00 | 0.04 | 0.10 | |
| Current speed (m s−1) | 0.07 | 0.00 | 0.03 | 0.03 | |
| Slope Index | 0.97 | 0.93 | 0.45 | 0.78 |
| Country | Environmental Variable | GAM | ANN | RF | All Models (Average) |
|---|---|---|---|---|---|
| Italy | Sea surface temperature (°C) | 0.03 | 0.10 | 0.22 | 0.12 |
| Salinity | 0.29 | 0.38 | 0.41 | 0.40 | |
| Current direction (East, m s−1) | 0.12 | 0.00 | 0.07 | 0.06 | |
| Current direction (North, m s−1) | 0.10 | 0.01 | 0.10 | 0.07 | |
| Current speed (m s−1) | 0.12 | 0.05 | 0.12 | 0.09 | |
| Slope index | 0.42 | 0.8 | 0.64 | 0.62 | |
| Spain | Sea surface temperature (°C) | 0.40 | 0.65 | 0.32 | 0.46 |
| Salinity | 0.36 | 0.03 | 0.45 | 0.28 | |
| Chlorophyll a (mg m−3) | 0.01 | 0.07 | 0.20 | 0.09 | |
| Wind direction (East, m s−1) | 0.02 | 0.08 | 0.40 | 0.17 | |
| Wind direction (North, m s−1) | 0.07 | 0.25 | 0.35 | 0.22 | |
| Current direction (East, m s−1) | 0.06 | 0.03 | 0.29 | 0.13 | |
| Current direction (North, m s−1) | 0.05 | 0.02 | 0.19 | 0.08 | |
| Current speed (m s−1) | 0.02 | 0.01 | 0.31 | 0.11 | |
| Slope index | 0.29 | 0.68 | 0.39 | 0.45 | |
| Tunisia | Sea surface temperature (°C) | 0.85 | 0.33 | 0.24 | 0.48 |
| Chlorophyll a (mg m−3) | 0.70 | 0.00 | 0.16 | 0.29 | |
| Phosphate (mmol m−3) | 0.80 | 0.00 | 0.00 | 0.27 | |
| Nitrate (mmol m−3) | 0.80 | 0.25 | 0.17 | 0.40 | |
| Wind direction (East, m s−1) | 0.84 | 0.40 | 0.06 | 0.43 | |
| Wind direction (North, m s−1) | 0.84 | 0.48 | 0.11 | 0.48 | |
| Wind speed (m s−1) | 0.83 | 0.48 | 0.55 | 0.62 | |
| Current direction (East, m s−1) | 0.49 | 0.00 | 0.16 | 0.22 | |
| Current direction (North, m s−1) | 0.52 | 0.00 | 0.12 | 0.21 | |
| Current speed (m s−1) | 0.24 | 0.00 | 0.12 | 0.12 | |
| Slope Index | 0.91 | 0.84 | 0.37 | 0.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marambio, M.; Canepa, A.; Lòpez, L.; Gauci, A.A.; Gueroun, S.K.M.; Zampardi, S.; Boero, F.; Yahia, O.K.-D.; Yahia, M.N.D.; Fuentes, V.; et al. Unfolding Jellyfish Bloom Dynamics along the Mediterranean Basin by Transnational Citizen Science Initiatives. Diversity 2021, 13, 274. https://doi.org/10.3390/d13060274
Marambio M, Canepa A, Lòpez L, Gauci AA, Gueroun SKM, Zampardi S, Boero F, Yahia OK-D, Yahia MND, Fuentes V, et al. Unfolding Jellyfish Bloom Dynamics along the Mediterranean Basin by Transnational Citizen Science Initiatives. Diversity. 2021; 13(6):274. https://doi.org/10.3390/d13060274
Chicago/Turabian StyleMarambio, Macarena, Antonio Canepa, Laura Lòpez, Aldo Adam Gauci, Sonia K. M. Gueroun, Serena Zampardi, Ferdinando Boero, Ons Kéfi-Daly Yahia, Mohamed Nejib Daly Yahia, Verónica Fuentes, and et al. 2021. "Unfolding Jellyfish Bloom Dynamics along the Mediterranean Basin by Transnational Citizen Science Initiatives" Diversity 13, no. 6: 274. https://doi.org/10.3390/d13060274
APA StyleMarambio, M., Canepa, A., Lòpez, L., Gauci, A. A., Gueroun, S. K. M., Zampardi, S., Boero, F., Yahia, O. K. -D., Yahia, M. N. D., Fuentes, V., Piraino, S., & Deidun, A. (2021). Unfolding Jellyfish Bloom Dynamics along the Mediterranean Basin by Transnational Citizen Science Initiatives. Diversity, 13(6), 274. https://doi.org/10.3390/d13060274