
The Effect of Climate and Human Pressures on Functional Diversity and Species Richness Patterns of Amphibians, Reptiles and Mammals in Europe

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Distribution Data
2.2. Trait Data
2.3. Functional Diversity Indices
2.4. Environmental Data
2.5. Statistical Analysis
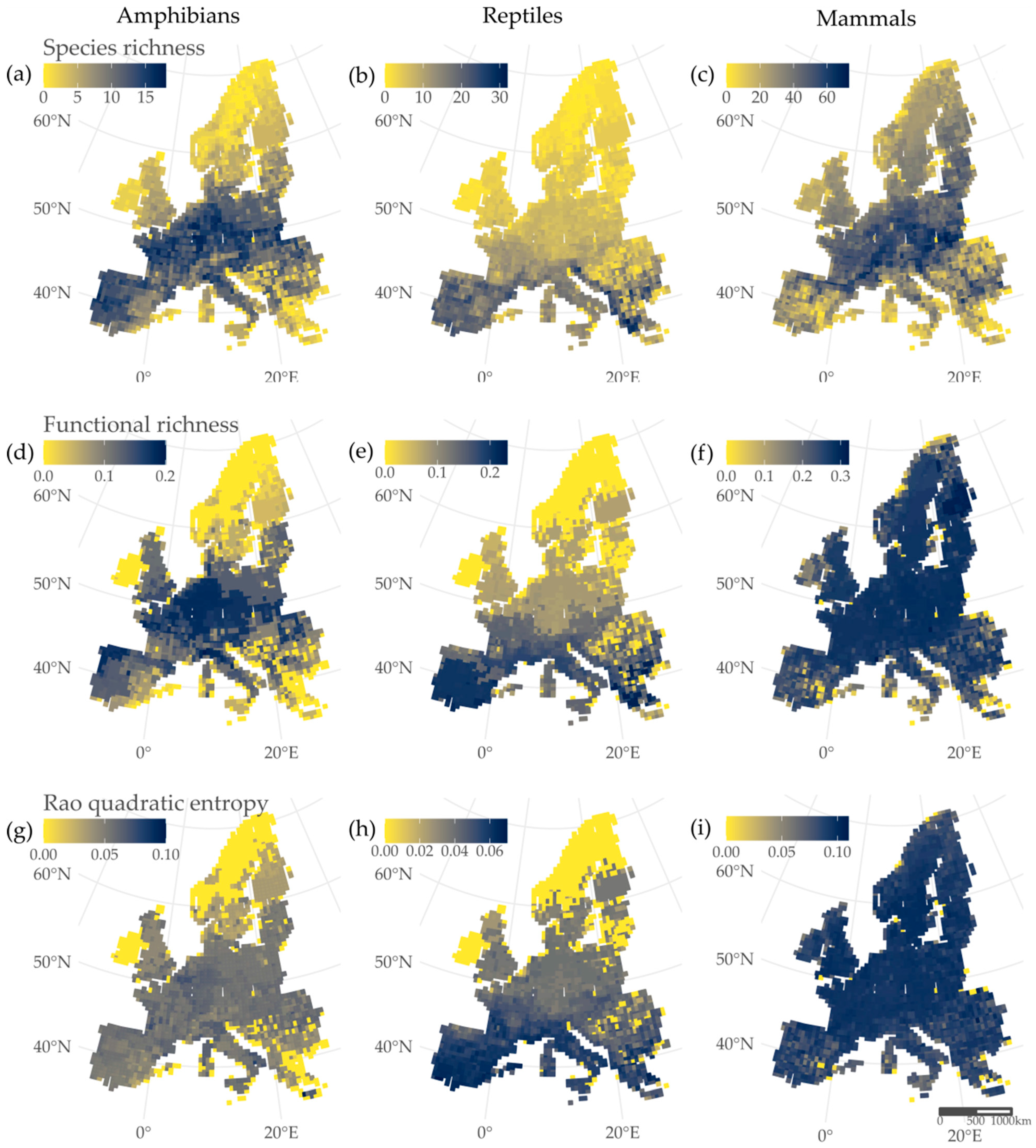
3. Results
4. Discussion
4.1. Climatic Gradients
4.2. Anthropogenic Gradients
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Field, R.; Hawkins, B.A.; Cornell, H.V.; Currie, D.J.; Diniz-Filho, J.A.F.; Guégan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T. Spatial species-richness gradients across scales: A meta-analysis. J. Biogeogr. 2009, 36, 132–147. [Google Scholar] [CrossRef]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional diversity: Back to basics and looking forward. Ecol. Lett. 2006, 9, 741–758. [Google Scholar] [CrossRef] [PubMed]
- Ricklefs, R.E. Disintegration of the ecological community: American Society of Naturalists Sewall Wright award winner address. Am. Nat. 2008, 172, 741–750. [Google Scholar] [CrossRef]
- Münkemüller, T.; Gallien, L.; Pollock, L.J.; Barros, C.; Carboni, M.; Chalmandrier, L.; Mazel, F.; Mokany, K.; Roquet, C.; Smyčka, J. Dos and don’ts when inferring assembly rules from diversity patterns. Glob. Ecol. Biogeogr. 2020, 29, 1212–1229. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Mason, N.W.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Concepción, E.D.; Götzenberger, L.; Nobis, M.P.; de Bello, F.; Obrist, M.K.; Moretti, M. Contrasting trait assembly patterns in plant and bird communities along environmental and human-induced land-use gradients. Ecography 2017, 40, 753–763. [Google Scholar] [CrossRef]
- Mouillot, D.; Bellwood, D.R.; Baraloto, C.; Chave, J.; Galzin, R.; Harmelin-Vivien, M.; Kulbicki, M.; Lavergne, S.; Lavorel, S.; Mouquet, N. Rare species support vulnerable functions in high-diversity ecosystems. PLoS Biol. 2013, 11, e1001569. [Google Scholar] [CrossRef] [Green Version]
- Schleuter, D.; Daufresne, M.; Veslot, J.; Mason, N.W.; Lanoiselée, C.; Brosse, S.; Beauchard, O.; Argillier, C. Geographic isolation and climate govern the functional diversity of native fish communities in European drainage basins. Glob. Ecol. Biogeogr. 2012, 21, 1083–1095. [Google Scholar] [CrossRef]
- Swenson, N.G.; Enquist, B.J.; Pither, J.; Kerkhoff, A.J.; Boyle, B.; Weiser, M.D.; Elser, J.J.; Fagan, W.F.; Forero-Montaña, J.; Fyllas, N. The biogeography and filtering of woody plant functional diversity in North and South America. Glob. Ecol. Biogeogr. 2012, 21, 798–808. [Google Scholar] [CrossRef]
- Bello, F.d.; Lavorel, S.; Lavergne, S.; Albert, C.H.; Boulangeat, I.; Mazel, F.; Thuiller, W. Hierarchical effects of environmental filters on the functional structure of plant communities: A case study in the French Alps. Ecography 2013, 36, 393–402. [Google Scholar] [CrossRef]
- Shiono, T.; Kusumoto, B.; Maeshiro, R.; Fujii, S.J.; Götzenberger, L.; de Bello, F.; Kubota, Y. Climatic drivers of trait assembly in woody plants in Japan. J. Biogeogr. 2015, 42, 1176–1186. [Google Scholar] [CrossRef]
- Mason, N.W.; Irz, P.; Lanoiselée, C.; Mouillot, D.; Argillier, C. Evidence that niche specialization explains species–energy relationships in lake fish communities. J. Anim. Ecol. 2008, 77, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Canavero, A.; Arim, M.; Brazeiro, A. Geographic variations of seasonality and coexistence in communities: The role of diversity and climate. Austral Ecol. 2009, 34, 741–750. [Google Scholar] [CrossRef]
- Safi, K.; Cianciaruso, M.V.; Loyola, R.D.; Brito, D.; Armour-Marshall, K.; Diniz-Filho, J.A.F. Understanding global patterns of mammalian functional and phylogenetic diversity. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2536–2544. [Google Scholar] [CrossRef]
- Brown, J.S.; Kotler, B.P.; Porter, W.P. How foraging allometris and resource dynamics could explain Bergmann’s rule and the body-size diet relationship in mammals. Oikos 2017, 126, 224–230. [Google Scholar] [CrossRef]
- Evans, K.L.; Warren, P.H.; Gaston, K.J. Species-energy relationships at the macroecological scale: A review of the mechanisms. Biol. Rev. 2005, 80, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Huang, Z.Y.; Chi, T.; Chen, B.J.; Zhang, M.; Liu, M. Can local landscape attributes explain species richness patterns at macroecological scales? Glob. Ecol. Biogeogr. 2014, 23, 436–445. [Google Scholar] [CrossRef]
- Sams, M.; Lai, H.; Bonser, S.; Vesk, P.; Kooyman, R.; Metcalfe, D.; Morgan, J.; Mayfield, M. Landscape context explains changes in the functional diversity of regenerating forests better than climate or species richness. Glob. Ecol. Biogeogr. 2017, 26, 1165–1176. [Google Scholar] [CrossRef]
- Rapacciuolo, G.; Marin, J.; Costa, G.C.; Helmus, M.R.; Behm, J.E.; Brooks, T.M.; Hedges, S.B.; Radeloff, V.C.; Young, B.E.; Graham, C.H. The signature of human pressure history on the biogeography of body mass in tetrapods. Glob. Ecol. Biogeogr. 2017, 26, 1022–1034. [Google Scholar] [CrossRef]
- Santini, L.; González-Suárez, M.; Rondinini, C.; Di Marco, M. Shifting baseline in macroecology? Unravelling the influence of human impact on mammalian body mass. Divers. Distrib. 2017, 23, 640–649. [Google Scholar] [CrossRef]
- Concepción, E.D.; Moretti, M.; Altermatt, F.; Nobis, M.P.; Obrist, M.K. Impacts of urbanisation on biodiversity: The role of species mobility, degree of specialisation and spatial scale. Oikos 2015, 124, 1571–1582. [Google Scholar] [CrossRef] [Green Version]
- Sol, D.; Trisos, C.; Múrria, C.; Jeliazkov, A.; González-Lagos, C.; Pigot, A.L.; Ricotta, C.; Swan, C.M.; Tobias, J.A.; Pavoine, S. The worldwide impact of urbanisation on avian functional diversity. Ecol. Lett. 2020, 23, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Flynn, D.F.; Gogol-Prokurat, M.; Nogeire, T.; Molinari, N.; Richers, B.T.; Lin, B.B.; Simpson, N.; Mayfield, M.M.; DeClerck, F. Loss of functional diversity under land use intensification across multiple taxa. Ecol. Lett. 2009, 12, 22–33. [Google Scholar] [CrossRef]
- Tsianou, M.A.; Kallimanis, A.S. Geographical patterns and environmental drivers of functional diversity and trait space of amphibians of Europe. Ecol. Res. 2020, 35, 123–138. [Google Scholar] [CrossRef]
- Cooke, R.S.; Bates, A.E.; Eigenbrod, F. Global trade-offs of functional redundancy and functional dispersion for birds and mammals. Glob. Ecol. Biogeogr. 2019, 28, 484–495. [Google Scholar] [CrossRef] [Green Version]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.S.; Furukawa, T.; Sasaki, T. Response diversity determines the resilience of ecosystems to environmental change. Biol. Rev. 2013, 88, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Barnagaud, J.Y.; Geniez, P.; Cheylan, M.; Crochet, P.A. Climate overrides the effects of land use on the functional composition and diversity of Mediterranean reptile assemblages. Divers. Distrib. 2021, 27, 50–64. [Google Scholar] [CrossRef]
- Sillero, N.; Campos, J.; Bonardi, A.; Corti, C.; Creemers, R.; Crochet, P.-A.; Isailović, J.C.; Denoël, M.; Ficetola, G.F.; Gonçalves, J. Updated distribution and biogeography of amphibians and reptiles of Europe. Amphib. Reptil. 2014, 35, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Mitchell-Jones, A.J.; Amori, G.; Bogdanowicz, W.; Krystufek, B.; Reijnders, P.; Spitzenberger, F.; Stubbe, M.; Thissen, J.; Vohralik, V.; Zima, J. The Atlas of European Mammals; Academic Press: London, UK, 1999; Volume 3. [Google Scholar]
- Mouchet, M.A.; Villéger, S.; Mason, N.W.; Mouillot, D. Functional diversity measures: An overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 2010, 24, 867–876. [Google Scholar] [CrossRef]
- Tsianou, M.A.; Kallimanis, A.S. Different species traits produce diverse spatial functional diversity patterns of amphibians. Biodivers. Conserv. 2016, 25, 117–132. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B.; Laliberté, M.E. Package ‘FD’. Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology; R Package Version. 2014. Available online: https://cran.r-project.org/web/packages/FD/FD.pdf (accessed on 18 June 2021).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Heymann, Y. Corine Land Cover: Technical Guide; Commission of the European Communities: Luxembourg, 1994. [Google Scholar]
- Klein Goldewijk, K.; Beusen, A.; Van Drecht, G.; De Vos, M. The HYDE 3.1 spatially explicit database of human-induced global land-use change over the past 12,000 years. Glob. Ecol. Biogeogr. 2011, 20, 73–86. [Google Scholar] [CrossRef]
- Wood, S.; Wood, M.S. Package ‘mgcv’. R Package Version 2015, 1, 29. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Buckley, L.B.; Hurlbert, A.H.; Jetz, W. Broad-scale ecological implications of ectothermy and endothermy in changing environments. Glob. Ecol. Biogeogr. 2012, 21, 873–885. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Global analysis of thermal tolerance and latitude in ectotherms. Proc. R. Soc. B Biol. Sci. 2011, 278, 1823–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunday, J.M.; Pecl, G.T.; Frusher, S.; Hobday, A.J.; Hill, N.; Holbrook, N.J.; Edgar, G.J.; Stuart-Smith, R.; Barrett, N.; Wernberg, T. Species traits and climate velocity explain geographic range shifts in an ocean-warming hotspot. Ecol. Lett. 2015, 18, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Tsianou, M.A.; Koutsias, N.; Mazaris, A.D.; Kallimanis, A.S. Climate and landscape explain richness patterns depending on the type of species’ distribution data. Acta Oecol. 2016, 74, 19–27. [Google Scholar] [CrossRef]
- Rodríguez, M.Á.; Belmontes, J.A.; Hawkins, B.A. Energy, water and large-scale patterns of reptile and amphibian species richness in Europe. Acta Oecol. 2005, 28, 65–70. [Google Scholar] [CrossRef]
- Aragón, P.; Lobo, J.M.; Olalla-Tárraga, M.Á.; Rodríguez, M.Á. The contribution of contemporary climate to ectothermic and endothermic vertebrate distributions in a glacial refuge. Glob. Ecol. Biogeogr. 2010, 19, 40–49. [Google Scholar] [CrossRef]
- Buckley, L.B.; Jetz, W. Insularity and the determinants of lizard population density. Ecol. Lett. 2007, 10, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Fløjgaard, C.; Normand, S.; Skov, F.; Svenning, J.C. Deconstructing the mammal species richness pattern in Europe–towards an understanding of the relative importance of climate, biogeographic history, habitat heterogeneity and humans. Glob. Ecol. Biogeogr. 2011, 20, 218–230. [Google Scholar] [CrossRef]
- Oliveira, B.F.; Machac, A.; Costa, G.C.; Brooks, T.M.; Davidson, A.D.; Rondinini, C.; Graham, C.H. Species and functional diversity accumulate differently in mammals. Glob. Ecol. Biogeogr. 2016, 25, 1119–1130. [Google Scholar] [CrossRef]
- Ochoa-Ochoa, L.M.; Mejía-Domínguez, N.R.; Velasco, J.A.; Marske, K.A.; Rahbek, C. Amphibian functional diversity is related to high annual precipitation and low precipitation seasonality in the New World. Glob. Ecol. Biogeogr. 2019, 28, 1219–1229. [Google Scholar] [CrossRef]
- Carrara, R.; Vázquez, D.P. The species–energy theory: A role for energy variability. Ecography 2010, 33, 942–948. [Google Scholar] [CrossRef]
- Cordier, J.M.; Aguilar, R.; Lescano, J.N.; Leynaud, G.C.; Bonino, A.; Miloch, D.; Loyola, R.; Nori, J. A global assessment of amphibian and reptile responses to land-use changes. Biol. Conserv. 2021, 253, 108863. [Google Scholar] [CrossRef]
- Araújo, M.B. The coincidence of people and biodiversity in Europe. Glob. Ecol. Biogeogr. 2003, 12, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Luck, G.W. A review of the relationships between human population density and biodiversity. Biol. Rev. 2007, 82, 607–645. [Google Scholar] [CrossRef] [PubMed]
- Guerra, C.; Aráoz, E. Amphibian diversity increases in an heterogeneous agricultural landscape. Acta Oecol. 2015, 69, 78–86. [Google Scholar] [CrossRef]
- Brüning, L.Z.; Krieger, M.; Meneses-Pelayo, E.; Eisenhauer, N.; Pinilla, M.P.R.; Reu, B.; Ernst, R. Land-use heterogeneity by small-scale agriculture promotes amphibian diversity in montane agroforestry systems of northeast Colombia. Agric. Ecosyst. Environ. 2018, 264, 15–23. [Google Scholar] [CrossRef]
- Mendoza, M.; Araujo, M.B. Climate shapes mammal community trophic structures and humans simplify them. Nat. Commun. 2019, 10, 5197. [Google Scholar] [CrossRef]
- De Solan, T.; Renner, I.; Cheylan, M.; Geniez, P.; Barnagaud, J.Y. Opportunistic records reveal Mediterranean reptiles’ scale-dependent responses to anthropogenic land use. Ecography 2019, 42, 608–620. [Google Scholar] [CrossRef] [Green Version]
- Ferger, S.W.; Peters, M.K.; Appelhans, T.; Detsch, F.; Hemp, A.; Nauss, T.; Otte, I.; Böhning-Gaese, K.; Schleuning, M. Synergistic effects of climate and land use on avian beta-diversity. Divers. Distrib. 2017, 23, 1246–1255. [Google Scholar] [CrossRef] [Green Version]
- Maiorano, L.; Amori, G.; Capula, M.; Falcucci, A.; Masi, M.; Montemaggiori, A.; Pottier, J.; Psomas, A.; Rondinini, C.; Russo, D. Threats from climate change to terrestrial vertebrate hotspots in Europe. PLoS ONE 2013, 8, e74989. [Google Scholar]



{kind=link}
{kind=link}
| Diversity Aspect | Taxonomic Group | Climatic Model | Land–Human Model | Overall Model |
|---|---|---|---|---|
| Species richness | ||||
| Amphibians | 0.41 | 0.35 | 0.44 | |
| Reptiles | 0.60 | 0.34 | 0.65 | |
| Mammals | 0.30 | 0.19 | 0.35 | |
| Functional richness | ||||
| Amphibians | 0.42 | 0.37 | 0.47 | |
| Reptiles | 0.59 | 0.33 | 0.61 | |
| Mammals | 0.25 | 0.10 | 0.30 | |
| Rao’s quadratic entropy | ||||
| Amphibians | 0.48 | 0.43 | 0.50 | |
| Reptiles | 0.61 | 0.38 | 0.61 | |
| Mammals | 0.23 | 0.10 | 0.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsianou, M.A.; Lazarina, M.; Michailidou, D.-E.; Andrikou-Charitidou, A.; Sgardelis, S.P.; Kallimanis, A.S. The Effect of Climate and Human Pressures on Functional Diversity and Species Richness Patterns of Amphibians, Reptiles and Mammals in Europe. Diversity 2021, 13, 275. https://doi.org/10.3390/d13060275
Tsianou MA, Lazarina M, Michailidou D-E, Andrikou-Charitidou A, Sgardelis SP, Kallimanis AS. The Effect of Climate and Human Pressures on Functional Diversity and Species Richness Patterns of Amphibians, Reptiles and Mammals in Europe. Diversity. 2021; 13(6):275. https://doi.org/10.3390/d13060275
Chicago/Turabian StyleTsianou, Mariana A., Maria Lazarina, Danai-Eleni Michailidou, Aristi Andrikou-Charitidou, Stefanos P. Sgardelis, and Athanasios S. Kallimanis. 2021. "The Effect of Climate and Human Pressures on Functional Diversity and Species Richness Patterns of Amphibians, Reptiles and Mammals in Europe" Diversity 13, no. 6: 275. https://doi.org/10.3390/d13060275
APA StyleTsianou, M. A., Lazarina, M., Michailidou, D. -E., Andrikou-Charitidou, A., Sgardelis, S. P., & Kallimanis, A. S. (2021). The Effect of Climate and Human Pressures on Functional Diversity and Species Richness Patterns of Amphibians, Reptiles and Mammals in Europe. Diversity, 13(6), 275. https://doi.org/10.3390/d13060275