
Target Species and Other Residents—An Experiment with Nest Boxes for Red Squirrels in Central Poland


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Data Collection and Laboratory Analyses
2.3. Statistical Analysis
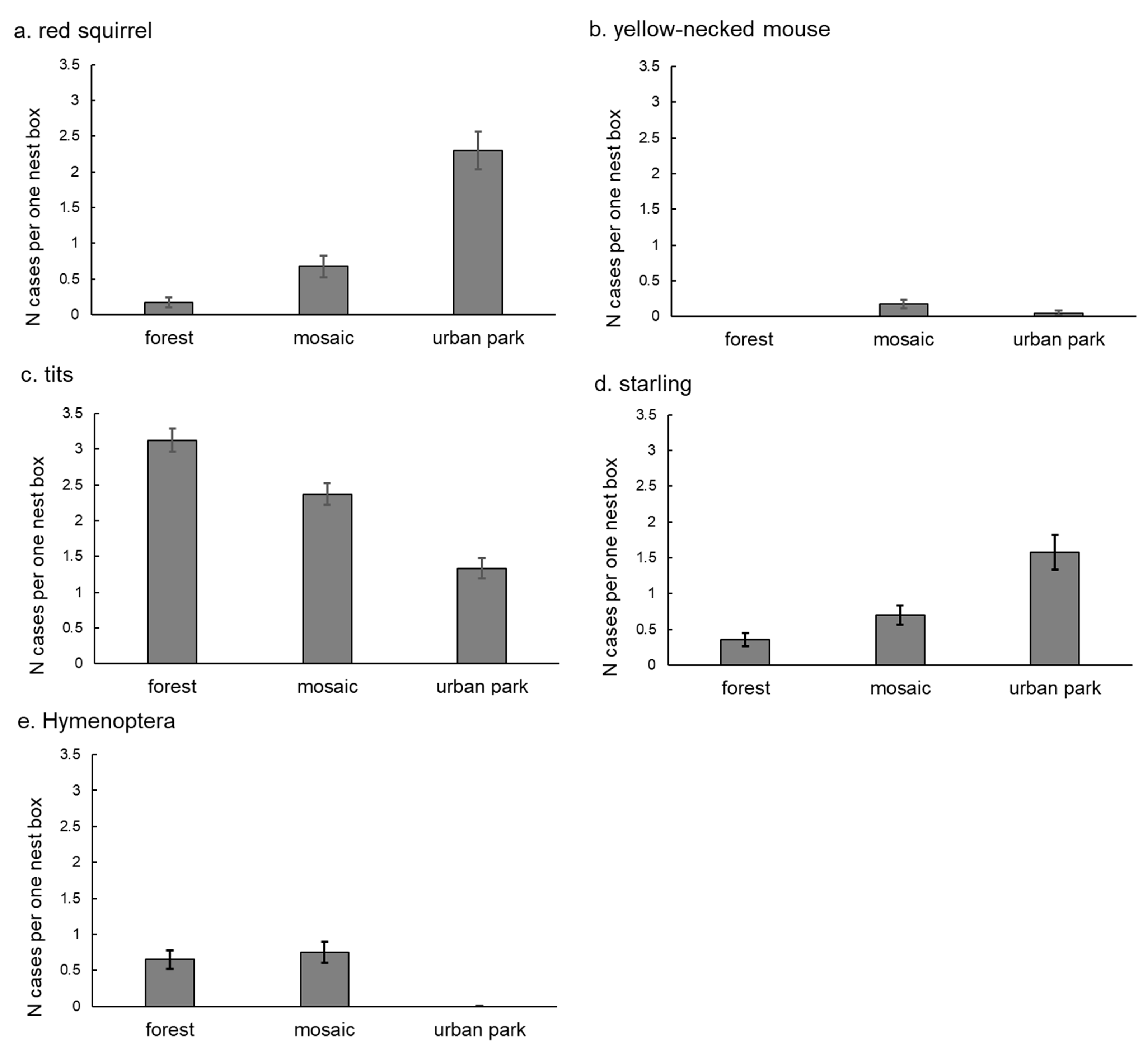
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years/Study Area | Squirrel | Mouse | Tits | Starling | Hymenoptera |
|---|---|---|---|---|---|
| n of records | |||||
| 1. Continous forest | |||||
| 2014 | 1 | 23 | 6 | ||
| 2015 | 29 | 5 | |||
| 2016 | 1 | 24 | 4 | 2 | |
| 2017 | 2 | 13 | 3 | 3 | |
| 2018 | 1 | 12 | 5 | ||
| 2019 | 1 | 16 | 1 | 7 | |
| 2020 | 1 | 8 | 1 | 3 | |
| Total | 7 | 125 | 14 | 26 | |
| 2. Habitat mosaic | |||||
| 2013 | 1 | 14 | 3 | 12 | |
| 2014 | 4 | 2 | 16 | 4 | 5 |
| 2015 | 5 | 2 | 14 | 5 | 1 |
| 2016 | 1 | 2 | 15 | 6 | 2 |
| 2017 | 7 | 11 | 3 | 6 | |
| 2018 | 4 | 10 | 1 | 5 | |
| 2019 | 3 | 9 | 5 | 9 | |
| 2020 | 3 | 1 | 8 | 4 | 2 |
| Total | 27 | 8 | 97 | 31 | 42 |
| 3. Urban park | |||||
| 2013 | 3 | 2 | 10 | ||
| 2014 | 21 | 2 | 13 | ||
| 2015 | 13 | 4 | 10 | ||
| 2016 | 15 | 1 | 10 | ||
| 2017 | 13 | 1 | 10 | ||
| 2018 | 10 | 5 | 4 | ||
| 2019 | 9 | 2 | 1 | 7 | |
| 2020 | 11 | 2 | 9 | ||
| Total | 95 | 2 | 18 | 73 | |
| All study areas | 129 | 10 | 240 | 118 | 68 |
Appendix C
| Order | Family | Family/Species | Total Abundance |
|---|---|---|---|
| Coleoptera | Coccinellidae | Harmonia axyridis (Pallas, 1773) | 1 |
| Cryptophagidae | Cryptophagus sp. | 1 | |
| Curculionidae | Phyllobius arborator (Herbst, 1797) | 1 | |
| Dermestidae | Anthrenus museorum (Linnaeus, 1761) | 2 | |
| Anthrenus pimpinellae Fabricius, 1775 | 3 | ||
| Attagenus pellio (Linnaeus, 1758) | 1 | ||
| Ctesias serra (Fabricius, 1792) | 1 | ||
| Dermestes lardarius Linnaeus, 1758 | 2 | ||
| Globicornis corticalis (Eichhoff, 1863) | 1 | ||
| Megatoma undata (Linnaeus, 1758) | 12 | ||
| Latridiidae | Latridius minutus (Linnaeus, 1767) | 1 | |
| Malachiidae | Malachius bipustulatus (Linnaeus, 1758) | 3 | |
| Ptinidae | Ptinus fur (Linnaeus, 1758) | 1 | |
| Ptinus raptor Sturm, 1837 | 1 | ||
| Stegobium paniceum (Linnaeus, 1758) | 1 | ||
| Tenebrionidae | Tenebrio molitor Linnaeus, 1758 | 2 | |
| Dermaptera | Forficulidae | Forficula auricularia Linnaeus, 1758 | 9 |
| Hemiptera | Lygaeidae | Arocatus melanocephalus (Fabricius, 1798) | 1 |
| Lepidoptera | Bucculatricidae | Bucculatrix thoracella (Thunberg, 1794) | 1 |
| Elachistidae | Agonopterix sp. | 1 | |
| Oecophoriadae | Borkhausenia minutella (Linnaeus, 1758) | 1 | |
| Pyralidae | Aphomia sociella (Linnaeus, 1758) | 4 | |
| Tineidae | Tinea pellionella Linnaeus, 1758 | 1 | |
| Neuroptera | Chrysopidae | Chrysopa sp. | 1 |
References
- Atlas of Polish Mammals 2021. Wiewiórka Pospolita Sciurus vulgaris Linnaeus. 1758. Available online: https://www.iop.krakow.pl/Ssaki/gatunek/57 (accessed on 18 May 2021).
- Shar, S.; Lkhagvasuren, D.; Bertolino, S.; Henttonen, H.; Kryštufek, B.; Meinig, H. Available online: www.iucnredlist.org (accessed on 18 May 2021).
- Stachura, K.; Niedziałkowska, M.; Bartoń, K. Biodiversity of forest mammals. In Essays on Mammals of Białowieża Forest; Jędrzejewska, B., Wójcik, M., Eds.; Mammal Research Institute, Polish Academy of Sciences: Białowieża, Poland, 2004; pp. 13–24. [Google Scholar]
- Wauters, L.A. The ecology of red squirrels in fragmented habitats: A review. In The Conservation of Red Squirrels, Sciurus vulgaris L.; Gurnell, J., Lurz, P., Eds.; People’s Trust for Endangered Species: London, UK, 1997; pp. 5–12. [Google Scholar]
- Babińska-Werka, J.; Żółw, M. Urban Populations of the Red Squirrel (Sciurus vulgaris) in Warsaw. Ann. Zool. Fenn. 2008, 45, 270–276. [Google Scholar] [CrossRef]
- Reher, S.; Dausmann, K.H.; Warnecke, L.; Turner, J.M. Food availability affects habitat use of Eurasian red squirrels (Sciurus vulgaris) in a semi-urban environment. J. Mammal. 2016, 97, 1543–1554. [Google Scholar] [CrossRef]
- Krauze-Gryz, D.; Gryz, J.; Brach, M. Spatial organisation, behaviour, and feeding habits of red squirrels: Differences between an urban park and an urban forest. J. Zool. 2021, 314. [Google Scholar] [CrossRef]
- Krauze-Gryz, D.; Gryz, J.; Klich, D.; Brach, M. Same yet different—individual red squirrels (Sciurus vulgaris) react differently to human presence in an urban park. Hystrix Ital. J. Mammal. 2021, 32. [Google Scholar] [CrossRef]
- Rézouki, C.; Dozières, A.; Coeur, C.; Le Thibault, S.; Pisanu, B.; Chapuis, J.L.; Baudry, E. A viable population of the European red squirrel in an urban park. PLoS ONE 2014, 9, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jokimäki, J.; Selonen, V.; Lehikoinen, A.; Kaisanlahti-Jokimäki, M.-L.L. The role of urban habitats in the abundance of red squirrels (Sciurus vulgaris, L.) in Finland. Urban For. Urban Green. 2017, 27, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Koprowski, J. The response of tree squirrels to fragmentation: A review and synthesis. Anim. Conserv. 2005, 8, 369–376. [Google Scholar] [CrossRef]
- Shuttleworth, C.M.; Schuchert, P. Are nest boxes a useful tool in regional red squirrel conservation programs? Hystrix Ital. J. Mammal. 2014, 25, 91–94. [Google Scholar] [CrossRef]
- Krauze-Gryz, D.; (Warsaw University of Life Sciences, Warsaw, Poland). Personal communication, 2021.
- Walankiewicz, W.; Czeszczewik, C.; Stański, S.; Sahel, M.; Ruczyński, I. Tree cavity resources in spruce-pine managed and protected stands of the Białowieża Forest, Poland. Nat. Areas J. 2014, 34, 423–428. [Google Scholar] [CrossRef]
- Andersson, J.; Gómez, E.D.; Michon, S.; Roberge, J.-M. Tree cavity densities and characteristics in managed and unmanaged Swedish boreal forest. Scand. J. For. Res. 2018, 33, 233–244. [Google Scholar] [CrossRef]
- Le Roux, D.S.; Ikin, K.; Lindenmayer, D.B.; Manning, A.D.; Gibbons, P. The future of large old trees in urban landscapes. PLoS ONE 2014, 9, e99403. [Google Scholar] [CrossRef] [PubMed]
- Lacki, M.J.; Hayes, J.P.; Kurta, A. Bats in Forests: Conservation and Management; Johns Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Cockle, K.L.; Martin, K.; Wesołowski, T. Woodpeckers, decay and the future of cavity-nesting vertebrate communities worldwide. Front. Ecol. Environ. 2011, 9, 377–382. [Google Scholar] [CrossRef] [Green Version]
- van der Hoek, Y.; Gaona, G.V.; Martin, K. The diversity, distribution and conservation status of the tree-cavity-nesting birds of the world. Divers. Distrib. 2017, 23, 1120–1131. [Google Scholar] [CrossRef] [Green Version]
- Siitonen, J. Dead wood in agricultural and urban habitats. In Biodiversity in Dead Wood. Ecology, Biodiversity and Conservation; Stokland, J.N., Siitonen, J., Jonsson, B.G., Eds.; Cambridge University Press: New York, NY, USA, 2012; pp. 380–401. [Google Scholar]
- Müller, J.; Jarzabek-Müller, A.; Bussler, H.; Gossner, M.M. Hollow beech trees identified as keystone structures for saproxylic beetles by analyses of functional and phylogenetic diversity. Anim. Conserv. 2013, 17, 154–162. [Google Scholar] [CrossRef]
- Micó, E. Saproxylic Insects in Tree Hollows. Diversity, Ecology and Conservation. In Saproxylic Insects; Zoological Monographs; Ulyshen, M., Ed.; Springer: Cham, Switzerland, 2018; Volume 1, pp. 693–727. [Google Scholar]
- Lambrechts, M.; Adriaensen, F.; Ardia, D.R.; Artemyev, A.V.; Atienzar, F.; Banbura, J.; Barba, E.; Bouvier, J.-C.; Camprodon, J.; Cooper, C.B.; et al. The design of artificial nestboxes for the study of secondary hole-nesting birds: A review of methodological inconsistencies and potential biases. Acta Ornithol. 2010, 45, 1–26. [Google Scholar] [CrossRef] [Green Version]
- General Directorate of State Forests. unpublished data. 2021.
- Broughton, R.K.; Hebda, G.; Maziarz, M.; Smith, K.W.; Smith, L.; Hinsley, S.A. Nest-site competition between bumblebees (Bombidae), social wasps (Vespidae) and cavity-nesting birds in Britain and the Western Palearctic. Bird Study 2015, 62, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Hicks, E.A. Check-List and Bibliography on the Occurrence of Insects in Birds’ Nests; Iowa State College Press: Ames, IA, USA, 1959. [Google Scholar]
- Petersen, G. Tineiden als Bestandteil der Nidicolenfauna. Beiträge Zur Entomol. 1963, 13, 411–427. [Google Scholar]
- Robinson, G.S. Moth and bird interactions: Guano, feathers, and detritophagous caterpillars (Lepidoptera, Tineidae). In Insect and Bird Interactions; van Emden, H.F., Rothschild, M., Eds.; Intercept: Andover, UK, 2004; pp. 271–285. [Google Scholar]
- Rushton, S.P.; Lurz, P.W.W.; Gurnell, J.; Nettleton, P.; Bruemmer, C.; Shirley, M.D.F.; Sainsbury, A.W. Disease threats posed by alien species: The role of a poxvirus in the decline of the native red squirrel in Britain. Epidemiol. Infect. 2006, 134, 521–533. [Google Scholar] [CrossRef]
- Gurnell, J.; Wauters, L.A.; Lurz, P.W.W.; Tosi, G. Alien species and interspecific competition: Effects of introduced eastern grey squirrels on red squirrel population dynamics. J. Anim. Ecol. 2004, 73, 26–35. [Google Scholar] [CrossRef]
- Romeo, C.; McInnes, C.J.; Dale, T.D.; Shuttleworth, C.; Bertolino, S.; Wauters, L.A.; Ferrari, N. Disease, invasions and conservation: No evidence of squirrelpox virus in grey squirrels introduced to Italy. Anim. Conserv. 2019, 22, 14–23. [Google Scholar] [CrossRef]
- Bertolino, S.; di Montezemolo, N.C.; Preatoni, D.G.; Wauters, L.A.; Martinoli, A. A grey future for Europe: Sciurus carolinensis is replacing native red squirrels in Italy. Biol. Invasions 2014, 16, 53–62. [Google Scholar] [CrossRef]
- de Raad, A.L.; Balafa, F.; Heitkonig, I.; Lurz, P.W.W. Mitigating the impact of forest management for conservation of an endangered forest mammal species: Drey surveys and nest boxes for red squirrels (Sciurus vulgaris). Hystrix Ital. J. Mammal. 2021, 32. [Google Scholar] [CrossRef]
- Shuttleworth, C.M. The use of nest boxes by the Red Squirrel Sciurus vulgaris in a coniferous habitat. Mammal Rev. 1999, 29, 63–68. [Google Scholar] [CrossRef]
- Shuttleworth, C.M. Interactions between the red squirrel (Sciurus vulgaris), great tit (Parus major) and jackdaw (Corvus monedula) whilst using nest boxes. J. Zool. 2001, 255, 269–272. [Google Scholar] [CrossRef]
- Turkia, T.; Korpimäki, E.; Villers, A.; Selonen, V. Predation risk landscape modifies flying and red squirrel nest site occupancy independently of habitat amount. PLoS ONE 2018, 13, e0194624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krauze-Gryz, D.; (Warsaw University of Life Sciences, Warsaw, Poland); Gryz, J.; (Forest Research Institute, Sękocin Stary, Poland). Personal communications, 2007.
- Gryz, J.; (Forest Research Institute, Sękocin Stary, Poland). Personal communication, 2021.
- Gryz, J.; Chojnacka-Ożga, L.; Krauze-Gryz, D. Long-term stability of tawny owl (Strix aluco) population despite varying environmental conditions–a case study from central Poland. Pol. J. Ecol. 2019, 67, 75–83. [Google Scholar] [CrossRef]
- Jaworski, T.; Gryz, J.; Buszko, J. Monopis fenestratella (Heyden, 1863) (Lepidoptera, Tineidae)–new records from Poland with notes on species biology. Fragm. Faun. 2011, 54, 149–151. [Google Scholar] [CrossRef] [Green Version]
- Jaworski, T.; Gryz, J.; Krauze-Gryz, D. Skrzynki lęgowe puszczyków (Strix aluco L.) jako środowisko występowania niektórych gatunków motyli (Lepidoptera). Wiadomości Entomol. 2012, 31, 17–22. [Google Scholar]
- Goszczyński, J.; Gryz, J.; Krauze, D. Fluctuations of a common buzzard Buteo buteo population in Central Poland. Acta Ornithol. 2005, 40, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Gryz, J.; Krauze-Gryz, D. Pigeon and poultry breeders, friends or enemies of northern goshawk Accipiter gentilis? A long term study of the population in central Poland. Animals 2019, 9, 141. [Google Scholar] [CrossRef] [Green Version]
- Gryz, J.; Krauze-Gryz, D. Common buzzard Buteo buteo population in a changing environment, central Poland as a case study. Diversity 2019, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Kruczek, Z. Analiza frekwencji w polskich atrakcjach turystycznych. Turyzm 2015, 25, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Krauze-Gryz, D.; Gryz, J. A review of the diet of the red squirrel (Sciurus vulgaris) in different types of habitats. In Red Squirrels: Ecology, Conservation & Management in Europe; Shuttleworth, C.M., Lurz, P.W., Hayward, M., Eds.; European Squirrel Initiative: Stoneleigh Park, Warwickshire CV8 2L, England, UK, 2015; pp. 39–50. [Google Scholar]
- Kostrzewa, A.; Krauze-Gryz, D. The choice and handling of supplemental food by red squirrels in an urban park. Behav. Process. 2020, 178, 104153. [Google Scholar] [CrossRef] [PubMed]
- ter Braak, C.J.F.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5). 2002. Available online: https://edepot.wur.nl/405659 (accessed on 15 May 2021).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 15 May 2021).
- Aitken, K.E.H.; Martin, K. Experimental test of nest-site limitation in mature mixed forests of central British Columbia, Canada. J. Wildl. Manag. 2012, 76, 557–565. [Google Scholar] [CrossRef]
- Wauters, L.; Dhondt, A. Nest-use by red squirrels (Sciurus vulgaris Linnaeus, 1758). Mammalia 1990, 54, 377–390. [Google Scholar] [CrossRef]
- Gryz, J.; Krauze-Gryz, D.; Lesiński, G. Mammals in the vicinity of Rogów (central Poland). Fragm. Faun. 2011, 54, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Croose, E.; Birks, J.D.S.; Martin, J. Den boxes as a tool for pine marten Martes martes conservation and population monitoring in a commercial forest in Scotland. Conserv. Evid. 2016, 13, 57–61. [Google Scholar]
- Krauze-Gryz, D.; Gryz, J.; Goszczyński, J.; Chylarecki, P.; Żmihorski, M. The Good, the Bad and the Ugly: Space use and intraguild interactions among three opportunistic predators–cat Felis catus, dog Canis familiaris and fox Vulpes vulpes –under human pressure. Can. J. Zool. 2012, 90, 1402–1413. [Google Scholar] [CrossRef] [Green Version]
- Krauze-Gryz, D.; Żmihorski, M.; Gryz, J. Annual variation in prey composition of domestic cats in rural and urban environment. Urban Ecosyst. 2017, 20, 945–952. [Google Scholar] [CrossRef] [Green Version]
- Krauze-Gryz, D.; Gryz, J.; Żmihorski, M. Cats kill millions of vertebrates in Polish farmland annually. Glob. Ecol. Conserv. 2019, 17, e00516. [Google Scholar] [CrossRef]
- Krauze-Gryz, D.; Mazur, K.; Gryz, J. Density of red squirrels and their use of non-native tree species in the Rogów arboretum. For. Res. Pap. 2016, 77, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Burger, G. Response of gray squirrels to nest boxes at Remington Farms, Maryland. J. Wildl. Manag. 1969, 33, 796–801. [Google Scholar] [CrossRef]
- Bosch, S.; Lurz, P. The Eurasian Red Squirrel; Westarp Wissenschaften: Hohenwarsleben, Germany, 2012. [Google Scholar]
- Marsh, A.C.W.; Morris, P.A. The use of dormouse Muscardinus avellanarius nest boxes by two species of Apodemus in Britain. Acta Theriol. 2000, 45, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Bosch, S.; Lurz, P.W.W. The process of drey construction in red squirrels-nestbox observations based on a hidden camera. Hystrix Ital. J. Mammal. 2013, 24, 199–202. [Google Scholar] [CrossRef]
- Berthier, K.; Leippert, F.; Fumagalli, L.; Arlettaz, R. Massive nest-box supplementation boosts fecundity, survival and even immigration without altering mating and reproductive behaviour in a rapidly recovered bird population. PLoS ONE 2012, 7, e36028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesiński, G. Location of bird nests in vertical metal pipes in suburban built-up area of Warsaw. Acta Ornithol. 2000, 35, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Chylarecki, P.; Chodkiewicz, T.; Neubauer, G.; Sikora, A.; Meissner, W.; Woźniak, B.; Wylegała, P.; Ławicki, Ł.; Marchowski, D.; Betleja, J.; et al. Trendy Liczebności ptaków w Polsce; GIOŚ: Warsaw, Poland, 2018. [Google Scholar]
- Mennechez, G.; Clergeau, P. Effect of urbanisation on habitat generalists: Starlings not so flexible? Acta Oecologica 2006, 30, 182–191. [Google Scholar] [CrossRef]
- Mazgajski, T. Nest site choice in relation to the presence of old nests and cavity depth in the starling Sturnus vulgaris. Ethol. Ecol. Evol. 2003, 15, 273–281. [Google Scholar] [CrossRef]
- Mazgajski, T. Effect of old nest material in nestboxes on ectoparasites abundance and reproductive output in the European Starling (Sturnus vulgaris L.). Pol. J. Ecol. 2007, 55, 377–385. [Google Scholar]
- Mazgajski, T. Competition for nest sites between the starling Sturnus vulgaris and other cavity nesters–study in forest park. Acta Ornithol. 2000, 35, 103–107. [Google Scholar]
- Pawlikowski, T.; Osmański, M. Atrakcyjność środowisk miejskich dla os społecznych (Hymenoptera: Vespinae) na obszarze Torunia. Wiadomości Entomol. 1998, 17, 95–104. [Google Scholar]
- Pawlikowski, K.; Pawlikowski, T. Nesting interactions of the social wasp Dolichovespula saxonica [F.] (Hymenoptera: Vespinae) in wooden nest boxes for birds in the forest reserve “Las Piwnicki” in the Chełmno Land (Northern Poland). Ecol. Quest. 2010, 13, 67–72. [Google Scholar]
- Harriott, N. Mosquito control and pollinator health. Pestic. You 2016, 36, 9–17. [Google Scholar]
- Seress, G.; Sándor, K.; Evans, K.L.; Liker, A. Food availability limits avian reproduction in the city: An experimental study on great tits Parus major. J. Anim. Ecol. 2020, 89, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Mroczkowski, M. Dermestidae. Skórnikowate. (Insecta: Coleoptera). Fauna Pol. Warszawa 1975, 4, 1–163. [Google Scholar]
- Buszko, J. Mole (Tineidae, Lepidoptera) zasiedlające huby i gniazda ptaków w rezerwacie Las Piwnicki. Acta Univ. Nicolai Copernic 1996, 96, 49–55. [Google Scholar]
- Buszko, J.; Pałka, K. Nowe dla fauny Polski gatunki Tineidae i Tortricidae (Lepidoptera). Wiadomości Entomol. 1992, 11, 105–111. [Google Scholar]
- Buszko, J.; Śliwinski, Z. Nowe dla fauny Polski i rzadko spotykane gatunki motyli (Lepidoptera). Wiadomości Entomol. 1979, 49, 653–662. [Google Scholar]
- Gaedike, R. Tineidae II (Myrmecozelinae, Perissomasticinae, Tineinae, Hieroxestinae, Teichobiinae and Stathmopolitinae). Microlepidoptera of Europe; Brill: Leiden, The Netherlands, 2019; Volume 9. [Google Scholar]
- Tokár, Z.; Lvovsky, A.; Huemer, P. Die Oecophoridae s. l. (Lepidoptera) Mitteleuropas. Bestimmung-Verbreitung-Habitat–Bionomie; Slamka: Bratislava, Slovakia, 2005. [Google Scholar]
- Lees, A.D. The physiology and biochemistry of diapause. Annu. Rev. Entomol. 1956, 1, 1–16. [Google Scholar] [CrossRef]
- Buszko, J.; Pacuk, B. Uwagi o zimowaniu imago Gracillariidae (Lepidoptera) w budkach lęgowych w Polsce. Wiadomości Entomol. 2009, 28, 255. [Google Scholar]
- Nadolski, J. Factors restricting the abundance of wasp colonies of the European hornet Vespa crabro and the Saxon wasp Dolichovespula saxonica (Hymenoptera: Vespidae) in an urban area in Poland. Entomol. Fenn. 2013, 24, 204–215. [Google Scholar] [CrossRef]
- Bourne, A.; Fountain, M.T.; Wijnen, H.; Shaw, B. Potential of the European earwig (Forficula auricularia) as a biocontrol agent of the soft and stone fruit pest Drosophila suzukii. Pest Manag. Sci. 2019, 75, 3340–3345. [Google Scholar] [CrossRef] [PubMed]
- Mänd, R.; Tilgar, V.; Lõhmus, A.; Leivits, A. Providing nest boxes for hole-nesting birds–Does habitat matter? Biodivers. Conserv. 2005, 14, 1823–1840. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gryz, J.; Jaworski, T.; Krauze-Gryz, D. Target Species and Other Residents—An Experiment with Nest Boxes for Red Squirrels in Central Poland. Diversity 2021, 13, 277. https://doi.org/10.3390/d13060277
Gryz J, Jaworski T, Krauze-Gryz D. Target Species and Other Residents—An Experiment with Nest Boxes for Red Squirrels in Central Poland. Diversity. 2021; 13(6):277. https://doi.org/10.3390/d13060277
Chicago/Turabian StyleGryz, Jakub, Tomasz Jaworski, and Dagny Krauze-Gryz. 2021. "Target Species and Other Residents—An Experiment with Nest Boxes for Red Squirrels in Central Poland" Diversity 13, no. 6: 277. https://doi.org/10.3390/d13060277
APA StyleGryz, J., Jaworski, T., & Krauze-Gryz, D. (2021). Target Species and Other Residents—An Experiment with Nest Boxes for Red Squirrels in Central Poland. Diversity, 13(6), 277. https://doi.org/10.3390/d13060277