3.2. Systematics
Order DECAPODA Latreille, 1802 [
39]
Infraorder STENOPODIDEA Spence Bate, 1888 [
1]
Family SPONGICOLIDAE Schram, 1986 [
40]
Genus
Spongicola de Haan, 1844 [
41]
3.2.1. Spongicola goyi Saito & Komai, 2008
Material examined. Northland. 1 M (PCL: 5.7 mm); Seamount #441, 34.043° S, 174.817° E, 880–792 m; 19 Apr 2002; R/V Kaharoa Stn. KAH0204/47; ‘seamount’ sled; NIWA 3621. Southern Kermadec Ridge. 1 F ov. (PCL: 5.9 mm); Southern Kermadec Ridge, 35.380° S, 178.980° E, 1184.1 m; 7 February 2017; RV Sonne Stn. SO254/33ROV08, Remote Operated Vehicle; NIWA 127111; found inside Corbitella sp.; NIWA 127123.
Diagnosis. Carapace with postrostral submedian and anterolateral spines; branchial region with fine scattered spinules or granules. Merus of third pereiopod unarmed or armed with row of small teeth or denticles on lateral margin; palm distinctly carinate on dorsal margin. Lateral margin of uropodal endopod serrate.
Measurements. CL: 7.6 mm (female), 7.9 mm (male), PCL: 5.9 mm (female), 5.7 mm (male), TL: 26.0 mm (female), 19.4 mm (male).
Distribution. Japan, Indonesia, Vanuatu, New Caledonia, Australia, Madagascar, and now northern New Zealand; 315–1184 m.
Hosts. Previously reported on
Demospongia sp., Hexactinellidae sp.,
Hyalonema sp.,
Euplectella sp. and
Pheronema semiglobosum Lévi & Lévi, 1982 [
43] by Saito & Komai [
12] and
Euhyalonema sp. by Goy [
13]. The female (NIWA 127123) reported here was collected with a sponge host most likely
Pheronema conicum Lévi & Lévi, 1982 [
43] (NIWA 126108, det. Michelle Kelly,
Figure 3). The male (NIWA 3621) included a collection note “with hexactinellid”, of which eight were collected at the same station, including three
Pheronema conicum but also
Chonelasma sp. and
Euryplegma auriculare Schulze, 1886 [
44].
Colour in life. Unknown. Collection note with NIWA 127123 refers to ‘bright blue eggs’.
Remarks. This is the first record for this genus and species in the New Zealand region, although the type series included specimens from New Caledonia and the Norfolk Ridge just north of New Zealand. The two specimens presented here extend the known range slightly southward to seamounts off the Northland Plateau and the southern Kermadec Ridge (
Figure 1). They were both collected from within hexactinellid sponges, possibly both from the amphidiscosid
Pheronema conicum. Genetically, the female (NIWA 127123) aligns with conspecific reference sequences (see below). Morphologically, both specimens match the type description in nearly all aspects. Slight variation for the ovigerous female compared to the original description is an additional large postorbital carapace spine, the first pleonite being more distinctly ridged (
Figure 4A), the posterior margin of the telson being distinctly convex (rather than slightly convex or nearly truncate) (
Figure 4C), the dactylus of the third pereiopod is smooth on the dorsal margin (not with row of teeth or denticles) (
Figure 4D) and the subdivision of the propodi of pereiopods 4 and 5 is indistinct. The specimen bears approximately 100 spherical eggs of a diameter 0.8 mm each. The male (NIWA 3621) matches the paratypes illustrated by Saito & Komai [
12], the third pereiopod is distinctly spinose along the ischium and merus and serrate along the margins of the palm and fingers. A slight difference is apparent in the shape of the eighth thoracic sternite that is anteriorly projected to a distinct, round tooth (instead of subrectangular).
The species resembles
S.
andamanicus Alcock, 1901 [
45] (widely distributed throughout the Indo-West Pacific) but can be chiefly distinguished by the lack of a large tooth on the lateral margin of the third pereiopod merus. The merus is instead furnished with a row of small teeth or denticles only.
DNA sequence data. DNA sequences for the large female (NIWA 127123) aligned well with previously published sequences, with intraspecific levels of divergences for 16S (≤0.8%) compared to four specimens from South China Sea, Papua New Guinea and New Caledonia [
17] (
Figure 2).
Genus
Spongicoloides Hansen, 1908 [
18].
3.2.2. Spongicoloides clarki sp. nov.
Material examined. Holotype: F ov. (PCL: 11.8 mm); Macauley Island, Kermadec Islands, 30.28–30.29° S, 178.20–178.20° W, 1431–1426 m; 29 Oct 2016; RV Tangaroa Stn. TAN1612/71, beam trawl; found inside Regadrella okinoseana; NIWA 118650.
Diagnosis. Carapace with faint hepatic groove; surface with only scattered small postrostral, hepatic and anterolateral spines, cervical groove not lined with distinct spines; anterior margins without antennal spine, few small anterolateral spines. Rostrum reaching distal margin of basal article of antennular peduncle; with nine small dorsal teeth, laterally unarmed. Epistome anteriorly straight, nearly smooth. Second and third pleonite with acute articular knob; fifth and sixth pleonites smooth on dorsal midline, unarmed posteroventral margin. Telson broadly rectangular, about twice as long as wide, small teeth along posterior margin. Antennular peduncle unarmed except for minute distomesial spine on second article, stout stylocerite. Antenna with small ventromedian spines on basicerite; scale with 7–8 small lateral teeth. First maxilliped with single arthrobranch. Second maxilliped with single arthrobranch, well-developed podobranch, lacking epipod. Third maxilliped with two arthrobranchs and rudimentary epipod, setiferous organ well-developed. First pereiopod with setiferous organ on propodus only. Third pereiopod nearly entirely glabrous and unarmed; fixed finger unarmed on distoventral margin; ischium with distodorsal spine and row of small ventral spines; coxa mesially unarmed. Fourth and fifth pereiopods dactyli with ventral unguis bearing a small ventral and dorsal tooth; coxa mesially unarmed; fourth pereiopod with paired arthrobranchs.
Etymology. Named after Malcolm Clark, Principle Scientist at NIWA, for his contribution to deep-sea, specifically seamounts, research, and who led the voyage that collected the holotype (Kermadec-Rangitahua, TAN1612), with KS’s gratitude for his mentoring and guidance.
Body large, robust, surface generally glabrous. Rostrum narrowly triangular in dorsal view, one-third (0.3) as long as PCL, horizontal, slightly over-reaching distal margin of first article of antennular peduncle; dorsal margin with nine small teeth and posterior blunt eminence at level of posterior orbital margin; ventral margin with two small teeth close to rostral tip; ventrolateral ridges unarmed.
Carapace glabrous, slightly inflated. Cervical and branchiostegal grooves distinct. Small spines on anterolateral, anterior branchial surface, smaller spines and granules scattered loosely on postrostral, anterior cardiac and hepatic surfaces. Orbital margin concave, antennal margin unarmed, small branchiostegal spine present, pterygostomian angle round, with series of three small spines on margin.
Thoracic sternites 6–8 anteriorly rounded, minutely serrate but unarmed; sternite 6 posterior margins evenly concave.
Pleonal somites glabrous, surfaces smooth. First pleonite shortest; divided into two sections by distinct transverse carina; ventral margin continuous, straight. Second and third pleonite subequal in length, with feeble transverse carina and acute articular knob; ventral margin rounded. Fourth and fifth pleonites with round posteroventral margin, minute granule on fifth tergite; dorsally smooth and unarmed. Sixth pleonite dorsally smooth; posterolateral corner angular, with very small granule; posterior margin smooth. Pleonal sternites unarmed.
Telson about 2.2 times as long as broad, subquadrangular, posterior margin shallowly convex, with long plumose setae; dorsolateral ridges distinct, with 7–8 spines each, proximalmost largest and placed mesially; lateral margins nearly straight, slightly constricted proximally, each side armed with a subproximal spine and 10–11 small, lateral spines; posterior margin with 11 small spines (4–5 lateral and a pair of median spines).
Eye well developed; cornea semiglobular, about half length of ocular peduncle in dorsal view, distinctly inflated, unpigmented; minute mesial distal spine on both eye stalks, and 1–2 small spines on dorsal surface.
Antennular peduncle reaching to about middle of antennal scale, not reaching spinose lateral margin of antennal scale. Basal article stout, length-width ratio of 2.8 at midlength, without statocyst, about twice as long as second article; lateral margin concave, distally produced to small, rounded lobe, stylocerite short but distinct, acute, barely reaching proximal quarter of basal article; mesial margin almost straight, unarmed other than row of serration where plumose seta are inserted. Second article about 1.5 times longer than distal article, distomesially with small spine. Distal article as long as wide, unarmed. Flagella slender, about twice as long as peduncle.
Antenna with first article (coxa, bearing antennal gland) mesially glabrous, not carinate, two small mesial spines. Basicerite stout; mesially unarmed, paired spines distolaterally; dorsal and lateral surfaces smooth, one (left) or two (right) central small spines on ventral surface. Antennal scale broad, 2.6 × as long as wide; lateral margin nearly straight, armed with 7–8 small teeth along the distal 0.4 portion, including the most prominent distal tooth; dorsal surface with two distinct longitudinal ridges. Carpocerite reaching to distal end of first article of antennular peduncle, minute dorsal and ventral spine distally. Flagellum at about 1.3 times the CL.
Epistome broadly convex, minutely granulate at anterior margin. Labrum smooth. Paragnaths bilobed, with deep median fissure, distodorsally spatulate.
Mandible robust, fused molar and incisor processes. Molar surface with three teeth; incisor bearing two distal teeth with regular row of small proximal teeth. Palp well-developed, 3-segmented; proximal article shortest, without setae; middle article with few setae on flexor and more, longer setae on extensor margins; distal article suboval, slightly longer than intermediate article (measured along extensor margin), densely setose.
Maxillule with simple palp, with four terminal setae; distal endite broad, round, with numerous simple setae and eight slender spines; proximal endite oval, with simple setae distally.
Maxilla with palp slender, tapering, with plumose setae, nearly reaching end of scaphognathite; distal and proximal endites both deeply bilobed, with numerous plumose setae; scaphognathite well developed, anterior portion longer than posterior portion, about three times longer than broad, with dense fringe of plumose setae along entire margin.
Branchial formula summarized in
Table 2.
First maxilliped with 2-segmented palp, bearing long plumose setae; with distal blunt spine, bearing distal plumose seta; proximal article stout, slightly longer than distal article in length. Distal endite large, subtriangular, densely setose; proximal endite deeply bilobed, with distal setae. Exopod slender, with long plumose setae. Epipod well developed, subequally bilobed. Arthrobranch small.
Second maxilliped with 5-segmented endopod, unarmed; dactylus sub-oval, tapering distally, about twice as long as broad, with dense setae on flexor margin; propodus slightly longer than dactylus, densely setose along flexor margin; carpus triangular, about two-third length of propodus measured at mid-line, with long distodorsal setae; merus nearly straight, about 1.6 × length of propodus, nearly four times longer than broad, with a row of setae along the mesial margin and sparse short setae on surfaces; ischium and basis not fused, each about 0.2 × meral length and long setae along mesial margin. Coxa with blunt mesial process; epipod absent, podobranch and arthrobranch. Exopod absent.
Third maxilliped endopod 5-segmented, slender, unarmed; dactylus narrow, gently tapering, about 4 times longer than broad (at mid-length), setose; propodus slightly less than twice as long as dactylus, with setiferous organ along entire length of flexor margin; carpus slightly longer than propodus; merus longest, about 1.5 × carpal length; ischium broadest, subequal in length to merus; all segments with long setal fringe along flexor margin; basis short. Coxa with rudimentary epipod, arthrobranchs present. Exopod absent.
First pereiopod slender, glabrous and sparsely setose; dactylus about 0.5 × palm length, distally setose; palm subcylindrical, tufts of long setae distally and along ventral margin of fixed finger; carpus 3 × palm length, ventral carpo-propodal setiferous organ pronounced on propodus, absent on carpus; merus about 0.7 × carpal length; ischium half meral length. Basis and coxa short, coxa with sharp mesial spine. Epipod absent.
Second pereiopod similar to first, 1.5 × longer and stronger, sparsely setose; dactylus 0.4 × palm length, distal tip formed into a strong corneous spine, tips of fingers cross when chela closed, cutting edges entire; propodus with a few tufts of setae distally and along fixed finger; carpus about twice palm length; merus 0.8 × carpal length; ischium 0.4 × length of merus. Basis and coxa short; coxa mesially produced to angular process. Epipod absent.
Third pereiopod largest, subequal and similar, about 2 × CL, overreaching the tip of antennal scale by length of chela, very sparsely setose except for a few distal tufts of setae along fingers; dactylus slightly less than 0.5 × palm length, ending in strong, hooked corneous tip, cutting edge with narrow trench along distal half, with broadly rounded tooth at mid-length, otherwise smooth; propodal cutting edge with sharp corneous spine distally, followed by narrow trench along distal half, with shallow trianguloid process at mid-length and distinct notch to accommodate dactylar tooth, proximal quarter straight, with numerous tiny teeth; outer margin of fixed finger unarmed, bearing tufts of long setae; palm sub-cylindrical, 3.1–3.3 times as long as broad, nearly entirely smooth, with few small spines on inner ventrodistal surfaces; carpus about 0.5 × palm length, narrowing proximally, without spines, with two distinct distodorsal lobes; merus 0.9 × length of palm, nearly 5 times longer than broad in lateral view, proximally compressed laterally, distoventral corners blunt, smooth other than ventral ridge with row of minute granules; ischium about three-fourth length of carpus, laterally compressed, with small, sharp distodorsal spine and small ventrodistal spine, preceded by ventral row of granules or small spines, distalmost strongest. Basis and coxa short; coxa mesially densely setose and finely granulate. Epipod absent.
Fourth and fifth pereiopods long and slender, similar, sparsely setose; dactylus about one-fifth the length of propodus, biungulate, both unguis clearly demarcated, both P5 with small accessory tooth on dorsal margin, right P5 with obsolescent accessory tooth on ventral margin proximally (P4 dactyli missing or damaged); propodus not subdivided, about 0.5 × carpal length, P5 propodus as wide as P4 propodus, with single row of 19–21 (P4) and 21 (P5) movable spine along entire flexor margin, both margins with few long, simple setae; carpus longest, not subdivided, with movable spine at distoventral angle; merus about 0.7 (P4)–0.7 (P5) × carpal length, unarmed; ischium about half length of merus, unarmed. Basis and coxa short, coxa mesially with dense setae, unarmed. Epipod absent.
First pleopod uniramous, second to fifth biramous, all lacking appendices, unarmed.
Uropod well developed, about as long as telson. Protopod stout, with sharp posterolateral spine. Exopod broader than endopod; lateral margin slightly convex, with 13 (left) and 16 (right) teeth along distal 0.8 portion of margin, closely spaced in distal quarter, distal margin nearly straight, dorsal surface with two distinct longitudinal ridges. Endopod simple, unarmed, surface with one longitudinal ridge. Exopod and endopod fringed with dense, plumose seta.
Eggs: The ovigerous female incubated around 100 eggs of diameters 1.4 × 2.1 mm.
Measurements. Holotype: CL: 15.2 mm, PCL: 11.8 mm, TL: 42.0 mm.
Distribution. Only known from holotype with certainty, Macauley Island, 1426–1431 m. Specimens that are displaying intraspecific levels of genetic divergence were collected on Yap Seamount, central-western Pacific at 1452.5 m (‘
Spongiocaris panglao’ [
37]) and Caroline Seamount, Micronesia, at a depth of 1205 m (Q. Kou pers. comm.)
Coloration. Unpigmented, most of the integument appears transparent (
Figure 7)
Hosts. Found inside euplectellid glass sponge
Regadrella okinoseana Ijima, 1896 [
46] (Hexactinellida, Lyssacinosida, Euplectellidae) (NIWA 118649, det. Michelle Kelly,
Figure 7). This host species is one of the most common euplectellids in the New Zealand region [
47].
Remarks.Spongiocoloides clarki sp. nov. most closely resembles the two other New Zealand congeners
S. novaezelandiae and
S. sonne sp. nov.; the third maxilliped and the first four pereiopods have two arthrobranchs each, the pleonites are dorsally smooth, the telson is sub-rectangular and the fixed finger of the third pereiopod is unarmed on the distoventral margin. The new species is distinct in its branchial formula with the second maxilliped lacking an epipod (usually present in both
S. novaezelandiae and
S. sonne) and a rudimentary epipod on the third maxilliped (present in
S. novaezelandiae and absent in
S. sonne) (
Table 2). Furthermore, the second and third pleonite has a distinctly acute articular knob (round in both other species, although this can be allometric), the labrum is nearly smooth anteriorly (furnished with granules or small spines in both other species), and the coxa of pereiopods 3–5 are mesially smooth (
S. sonne has distinct and
S. novaezelandiae small mesial spines).
Spongiocoloides clarki shares with
S. novaezelandiae the smoother carapace surface with only scattered small spines but it lacks the distinct spines along the cervical groove which are small in
S. novaezelandiae and pronounced in
S. sonne, the shorter rostrum, barely overreaching the first antennular segment (reaching the end of the second antennular segment in
S. sonne). Conversely,
S. clarki shares with
S. sonne the spinose ischium of the third pereiopod (ventrally smooth in
S. novaezelandiae).
The characteristics of the pereiopods 4–5 dactylar spination warrants further investigation, both available intact pereiopods of S. clarki have a minute dorsal accessory tooth on the ventral unguis and lacked a distinct ventral accessory tooth or teeth. The latter are pronounced in both S. novaezelandiae and S. sonne. The collection of further intact specimens might prove this character to be fixed and diagnostic.
DNA sequence data. Closest allies with intraspecific levels of divergences in all cases are a GenBank sequence presented as ‘
Spongiocaris panglao’ by Sun et al. [
37] (MG812382) and a specimen of
Spongicoloides sp. (Qi Kou pers. comm.).
CO1: intra-specific divergences between 1.9% (‘
Spongiocaris panglao’ [
37]) and 2.2%,
Spongicoloides sp. (Q.Kou, unpubl.); intra-generic divergences range from 4.4% (
Spongicoloides novaezelandiae) to 7.1% (
S. corbitellus Kou, Gong & Li, 2018 [
35]).
16S rRNA: intra-specific divergences of 1.0% (‘
Spongiocaris panglao’ [
37] and
Spongicoloides sp. (Q. Kou, unpubl.); intra-generic divergences range from 2.3% (
Spongicoloides weijiaensis) to 15.1% (
S. iheyaensis Saito, Tsuchida & Yamamoto, 2006 [
48]).
3.2.3. Spongicoloides novaezelandiae Baba, 1979
Material examined. Holotype: M (PCL: 14.4 mm); Chatham Rise, 44.73° S, 175.70° E, 1782–1998 m; 16 Jul 1968; RV Kaiyo Maru Stn. KM36, trawl; NMNZ CR.001889. Other material. Macquarie Ridge: 1 F (PCL: ~9.8 mm, specimen damaged); Seamount 7, 53.731–53.733° S, 159.166–159.169° E, 1150–1270 m; 12 Apr 2008; RV Tangaroa Stn. TAN0803/81, ‘seamount’ sled; NIWA 40567. 1 M (PCL: ~6.6 mm, specimen damaged); Seamount 7, 53.705–53.705° S, 159.115–159.106° E; 998–1100 m; 13 Apr 2008, ‘seamount’ sled; RV Tangaroa Stn. TAN0803/84; NIWA 40638.
Diagnosis. Carapace with shallow hepatic groove; postorbital surface smooth; minute hepatic spines; cervical groove with few distinct spines; anterior margins without antennal spine, few small anterolateral spines. Rostrum reaching distal margin of basal segment of antennular peduncle; with 8–11 small dorsal teeth, laterally unarmed. Epistome anteriorly convex, with small teeth. Pleonites smooth on dorsal midline; pleonites 5–6 with 1–2 small spines along posteroventral margin. Telson broadly rectangular, about twice as long as wide; posterior margin with median pair of spines, regular row of spines absent. Antennule unarmed, stout stylocerite. Antennal basicerite with pair of distolateral teeth, ventromedially unarmed; scale with 7–8 lateral teeth. First maxilliped with single arthrobranch developed. Second maxilliped with two arthrobranchs and well-developed podobranch. First pereiopod with setiferous organ on propodus and carpus. Third pereiopod nearly entirely glabrous and unarmed; fixed finger unarmed on distoventral margin; ischium with distodorsal spine, ventrally unarmed; coxa with or without mesial granules. Fourth and fifth pereiopod dactyli with ventral unguis bearing a small ventral and with or without slight indication of dorsal tooth; coxa with small distomesial granules; P4 with paired arthrobranchs.
Description of the holotype male (
Figure 8).
Body large, robust, surface generally glabrous.
Rostrum narrowly triangular in dorsal view, one-third (0.3) as long as PCL, nearly horizontal, slightly overreaching basal segment of antennular peduncle; dorsal margin armed with 11 small, low teeth, posteriormost placed on carapace posterior to level of orbital margin; distoventral margin with two small teeth; no distinct ventrolateral ridges, lateral margins smooth, unarmed.
Carapace glabrous, not inflated. Cervical and branchiostegal grooves distinct; cervical groove lined with five small spines. Group of small spinules placed on lateral portion of gastric region; postrostral, postorbital surfaces nearly smooth, minute dorsal granules. Orbital margin concave, antennal and branchiostegal margin unarmed, anterior pterygostomian angle round, with 1–3 small spines on margin; field of several spinules just behind them.
Thoracic sternites 6–8 anteriorly rounded, unarmed; sternite six posterior margins straight.
Pleonal somites glabrous. First pleonite shortest; divided into two sections by distinct transverse carina; ventral margin continuous, not produced. Second and third pleonite subequal in length, with feeble transverse groove and blunt articular knob; ventral margin of second to fourth segments broadly rounded. Fifth and sixth pleura with acute posteroventral margin; dorsally smooth and unarmed; posterior margins smooth. Pleonal sternites unarmed.
Telson, twice as long as broad, subquadrangular, posterior margin shallowly convex, with long plumose setae; dorsolateral ridges distinct, with 8 spines each, distalmost spine very small, near posterolateral margin, distinctly separated from ordinary row. Lateral margins nearly straight, slightly constricted proximally, each side armed with a subproximal spine and 6–7 small, lateral spines. Posterior margin with two denticles equidistant between lateral groups of 3–4 similar denticles.
Eye well developed; cornea semiglobular, about half length of ocular peduncle in dorsal view, slightly inflated, unpigmented; minute distomesial spine on both eye stalks.
Antennular peduncle overreaching middle of antennal scale, reaching spinose lateral margin of antennal scale. Basal article relatively stout, length-width ratio of about 3.0 at mid-length, without statocyst, about twice as long as second article, distally unarmed but with short distodorsal carina; lateral margin concave, stylocerite small and blunt but distinct; mesial margin almost straight, unarmed other than row of serration where plumose seta are inserted. Second article about 1.5 times longer than distal article, unarmed. Distal article as long as wide, unarmed. Flagella slender, about twice as long as peduncle.
Antenna with first article (coxa, bearing antennal gland) mesially glabrous, not inflated or carinate, one small mesial spine and a row of small setiferous denticles. Basicerite stout; mesially unarmed, one (left) or two (right) outer terminal spines; surfaces smooth, unarmed. Antennal scale broad, around twice as long as broad; lateral margin almost straight, not setiferous, with 6–7 spines in distal half, distally paired on both sides; dorsal surface with two distinct longitudinal ridges; inner margin convex, inner and distal margins with long setae. Carpocerite reaching to distal end of first article of antennular peduncle, unarmed. Flagellum at least as long as PCL.
Epistome narrowly convex, with short transverse row of granules at anterior margin. Labrum smooth. Paragnaths bilobed, with deep median fissure, distodorsally spatulate.
Mandible robust, fused molar and incisor processes. Molar surface with two teeth; incisor bearing distal tooth followed by regular row of small teeth along midlength. Palp well-developed, 3-segmented; proximal article shortest, without setae; middle article with few setae on flexor and more, longer setae on extensor margins; distal article suboval, longer than intermediate article (measured along extensor margin), densely setose.
Maxillule with simple palp, with four terminal setae; distal endite broad, round, with numerous simple setae and eight slender spines; proximal endite oval, with four simple setae distally.
Maxilla with palp slender, tapering, with plumose setae, falling well short of end of scaphognathite; distal and proximal endites both deeply bilobed, with numerous plumose setae; scaphognathite well developed, anterior portion longer than posterior portion, about 3.5 times longer than broad, with dense fringe of plumose setae along entire margin.
Branchial formula summarized in
Table 2.
First maxilliped with 2-segmented palp, bearing long plumose setae; distal article as long as wide, rounded, with plumose seta; proximal article stout, slightly longer than distal article in length. Distal endite large, subtriangular, densely setose; proximal endite deeply bilobed, with distal setae. Exopod slender, with long plumose setae. Epipod well developed, subequally bilobed. Arthrobranch small.
Second maxilliped with 5-segmented endopod, unarmed; dactylus sub-oval, tapering distally, about 1.5 times as long as broad, with dense setae on flexor margin; propodus slightly longer than dactylus, densely setose along flexor margin; carpus triangular, about two-third length of propodus measured at mid-line, with long distodorsal setae; merus nearly straight, about twice as long as propodus, three times longer than broad, with row of setae along mesial margin and sparse short setae on surfaces; ischium and basis not fused, each about 0.2 × meral length and long setae along mesial margin. Coxa with blunt mesial process; small epipod, podobranch and pair of arthrobranchs present. Exopod absent.
Third maxilliped endopod 5-segmented, slender, unarmed; dactylus narrow, gently tapering, about 4 times longer than broad (at mid-length), setose; propodus slightly less than twice as long as dactylus, with setiferous organ along entire length of flexor margin; carpus subequal in length to propodus; merus longest, about twice carpal length; ischium broadest, subequal in length to merus; all articles with long setal fringe along flexor margin; basis short. Coxa with small epipod, mesial margin bluntly triangular; arthrobranchs present. Exopod absent.
First pereiopod slender, glabrous and sparsely setose; fingers unarmed, half as long as palm, distally setose; palm subcylindrical, tufts of long setae distally and along ventral margin of fixed finger; carpus 2.3 times as long as palm, ventral carpo-propodal setiferous organ pronounced on propodus and carpus; merus about 0.7 × carpal length; ischium half meral length. Basis and coxa short, coxa with small mesial spine. Epipod absent.
Second pereiopod similar to first, 1.5 × longer and stronger, sparsely setose; dactylus 0.4 × palm length, distal tip formed into a strong corneous spine, tips of fingers cross when chela closed, cutting edges entire; propodus with a few tufts of setae distally and along fixed finger; carpus about 1.5 × palm length; merus 0.8 × carpal length; ischium 0.4 × length of merus. Basis and coxa short. Epipod absent.
Third pereiopod largest, subequal and similar, about 1.7 × CL, very sparsely setose except for a few distal tufts of setae along fingers; dactylus slightly less than 0.5 × palm length, ending in strong, hooked corneous tip, cutting edge with narrow trench along distal half, with broadly rounded tooth at proximal third, otherwise smooth; propodal cutting edge with sharp corneous spine distally, followed by narrow trench along distal half, mid-length barely produce, distinct proximal notch to accommodate dactylar tooth; outer margin of fixed finger unarmed, bearing tufts of long setae; palm sub-cylindrical, moderately compressed, 3.4 times as long as broad, smooth, with few minute spines on inner ventrodistal surfaces; carpus about 0.4 × palm length, narrowing proximally, without spines, with knoblike process at anterior end of inner surface; merus 0.8–0.9 × length of palm, around four times longer than broad in lateral view, proximally compressed laterally, distoventral corners blunt, smooth; ischium about three-fourth length of carpus, laterally compressed, with small, sharp distodorsal spine only, ventral margin smooth. Basis and coxa short; coxa mesially densely setose, unarmed. Epipod absent.
Fourth and fifth pereiopods long and slender, similar, sparsely setose; dactylus about one-fifth the length of propodus, biungulate; both unguis clearly demarcated, with small accessory tooth on ventral margin, distinct shoulder dorsally; propodus not subdivided, slightly less than 0.5 × carpal length, fifth pereiopod propodus as wide as fourth pereiopod propodus, with single row of 10–12 movable spine along entire flexor margin, both margins with few long, simple setae; carpus longest, not subdivided, with movable spine at distoventral angle; merus about 0.9 × carpal length, unarmed; ischium about half length of merus, unarmed. Basis and coxa short, coxa mesially with dense setae, with small granules. Epipod absent.
First pleopod uniramous, second to fifth biramous, all lacking appendices, unarmed.
Uropod well developed, about as long as telson. Protopod stout, with sharp posterolateral spine. Exopod broader than endopod; lateral margin slightly convex, with 8–10 teeth along distal 0.8 portion of margin, distal margin convex, dorsal surface with 2 distinct longitudinal ridges. Endopod simple, unarmed, surface with 1 longitudinal ridge. Exopod and endopod fringed with dense, plumose seta.
Measurements. Holotype: CL: 19.5 mm, PCL: 14.4 mm, TL: 55 mm. NIWA 40567*: CL: 12.7 mm, PCL: 9.8 mm, TL: 34.2 mm. NIWA 40638*: CL: 9.3 mm, PCL: 6.6 mm, TL: 29.3 mm. *Measurements for both NIWA specimens are approximate as the specimens are damaged.
Distribution. New Zealand (type locality), southern Chatham Rise, 990–1110 m. New records presented here extend the range southwards to Macquarie Ridge, 998–1270 m. Based on the confirmed records at hand, this appears to be the only species in this genus with a southern temperate to sub-Antarctic distribution (
Figure 1). Records recently presented by Goy [
13] of
S. novaezelandiae from New Caledonia are referable to
S. sonne sp. nov. and
S. weijiaensis based on DNA sequence similarities (
Figure 2 and see comments below). Further specimens from Tasmania, Madagascar, Indian Ocean and Fiji mentioned by Goy [
13] require more detailed examination.
Coloration. Body and appendages transparent, cornea yellow [
20].
Hosts. Unknown.
Remarks. Following the examination of the male holotype of
S.
novaezelandiae, the following characters were not previously presented and can be added as follows: the epistome is anteriorly rounded, bearing a row of small granules along the anterior margin; the labrum is anteriorly rounded and smooth (
Figure 8D); both ocular peduncles bear a small dorsomesial granule (
Figure 8A); the basal antennal articles are smooth and not bearing any ventromesial processes (
Figure 8F); the coxa of pereiopods 4–5 bear small distomesial granules only (
Figure 8E). The proposed apomorphy for
S.
novaezelandiae is the first maxilliped distal segment broad and rounded (not tapering) [
20], this is consistent on both sides of the holotype, however, the additional specimens presented below do not share this character (
Figure 10E). Baba [
20] illustrates the entire dorsal and the left anterior carapace but fails to include the small but distinct series of spines that line the cervical groove. They are less distinct on the left than the right, illustrated here (
Figure 8A).
Two single specimens collected at two stations on Macquarie Ridge at >53° S provide the most southerly records for the genus to date. Unfortunately, both specimens are badly damaged, both lack third maxilliped and pereiopod 3, and only one pereiopod 4 or 5 of NIWA 40567 is preserved (
Figure 9,
Figure 10 and
Figure 11). Genetically, these specimens clearly align with the holotype of
S.
novaezelandiae (see below). Morphologically, they share the following characters that may prove to be diagnostic: e.g., the nearly smooth postrostral carapace surface; the telson is sub-rectangular, length-width ratio is ≤ 2.0, posterior margin with few or irregular row of spines; anterior margin of epistome bearing at least two granules; maxillipeds 2 and 3 bear a small epipod each. Notably, the distal segment of the first maxilliped palp bears a distinct spine (
Figure 10E).
Spongicoloides novaezelandiae was described from a single male collected on the New Zealand Chatham Rise (
Figure 1) and was recently reported again from a wide geographic range and with considerable morphological variation [
13]. Following the examination of New Zealand material presented here in combination with DNA sequencing, we propose that only the additional New Zealand material reported, NIWA 40567 and 40638 from Macquarie Ridge, belong to
S.
novaezelandiae and that all remaining specimens are referable to the following species:
- (1).
Spongicoloides weijiaensis Xu, Zhou & Wang, 2017 [
36]
Three of four specimens reported from New Caledonia (BIOCAL specimens listed in Goy [
13] as MNHN-Na-11996 [IU-2013-19630], Na-11997 [IU-2013-19488] and Na-11998 [IU-2013-19627]) were successfully sequenced and are referred to
S. weijiaensis (
Figure 2), see comments under DNA sequence section below). According to figures provided (Goy [
13]:
Figure 10), these all share dorsal carapace spines on parts of the postrostral, hepatic and cardiac regions, but specimens will need to be examined in more detail. In the meantime, we propose the following characteristics to separate
S.
novaezelandiae and
S. weijiaensis to include:
- (a).
The postorbital carapace region is nearly entirely smooth in S. novaezelandiae (with at least some scattered spines in S. weijiaensis).
- (b).
The epistome bears anterior teeth or row of small granules (smooth in S. weijiaensis).
- (c).
The antennal basicerite with nearly entirely smooth ventral surface in S. novaezelandiae (small ventral ridge armed with 1–3 spines present in S. weijiaensis).
- (d).
P3 ischium is smooth on ventral margin in S. novaezelandiae (irregular, with at least some proximal granules in S. weijiaensis).
The presence and number of arthrobranchs on the maxilliped 1 and 2 are usually considered diagnostic and the holotype of S. novaezelandiae has two arthrobranchs on maxilliped 2 which would differ from the single arthrobranch in S. weijaensis. However, this character is unusually variable in S. novaezelandiae with only one arthrobranch on maxilliped 2 of NIWA 40567 and one rudimentary arthrobranch on NIWA 40638.
The distribution of S. weijiaensis now extends from the northwestern Pacific to New Caledonia, north of the New Zealand region.
- (2).
Spongicoloides sonne sp. nov.
The figured CALSUB specimen (MNHN-NA 11999 [IU-2013-19487]) (Goy [
13]:
Figure 9A and
Figure 10A) shows a distinct antennal spine on the anterolateral margin of the carapace (rounded in
S.
novaezelandiae) and a distinctly spinose ischium of the third pereiopod (smooth in
S.
novaezelandiae). Additionally, the rostrum reaches at least to the distal end of the second antennular article (reaching the end of the first article in
S.
novaezelandiae). These characteristics match those of
S.
sonne sp. nov. described and discussed below.
The material from Tasmania, Madagascar, Indian Ocean and Fiji referred to by Goy [
13] will need to be examined in more detail in light of the review presented here.
Spongicoloides novaezelandiae most closely resembles
S. clarki sp. nov. and
S. sonne sp. nov., both reported from the northern New Zealand region (
Figure 1). Morphological differences are discussed under those species below.
DNA sequence data. The holotype of
S. novaezelandiae (NMNZ CR.001889) could be sequenced for both genes and aligned with the two specimens from Macquarie Ridge (NIWA 40567, 40638). The specimen reported as
S.
novaezelandiae by Chen et al. ([
17] MNHN-IU-2014-6347) from the Solomon Islands belongs to a different species and aligns more closely with an undescribed species of
Spongicoloides from the northwest Pacific (Zhao et al., in press.). Three of the four specimens presented as
S. novaezelandiae by Goy [
13] were successfully sequenced and align with the holotype sequence of
S. weijiaensis (SRSIO-16050001,
Figure 2).
CO1: intra-specific divergences between 0.6–1.7%; intra-generic divergences range from 4.4% (Spongicoloides clarki sp. nov.) to 8.5% (S. corbitellus).
16S rRNA: intra-specific divergences were 0.0%; intra-generic divergences range from 2.6% (Spongicoloides clarki sp. nov.) to 16.0% (S. iheyaensis).
3.2.4. Spongicoloides sonne sp. nov.
In (
Figure 1,
Figure 2,
Figure 12,
Figure 13,
Figure 14,
Figure 15,
Figure 16 and
Figure 17).
Spongicoloides novaezelandiae.— Goy 2015 [
13]: 310,
Figure 9,
Figure 10 and
Figure 11 (in part, New Caledonia, specimen MNHN-NA 11999 [IU-2013-19487]).
Material examined. Holotype: F ov. (PCL: 15.0 mm); Southern Kermadec Ridge, 35.380° S, 178.980° E, 1184.1 m; 7 February 2017; RV Sonne Stn. SO254/33ROV08, Remote Operated Vehicle; NIWA 127111; found inside Corbitella sp. Allotype: M (PCL: 11.4 mm); Southern Kermadec Ridge, 35.380° S, 178.980° E, 1184.1 m; 7 February 2017; RV Sonne Stn. SO254/33ROV08, Remote Operated Vehicle; NIWA 127110; found inside Corbitella sp. Other material. 1 F (PCL: 11.0 mm); West Norfolk Ridge, 34.28° S, 168.41° E, 1246–1249 m; 2 June 2003; TAN0308/142, Orange Roughy trawl; NMNZ CR.019650. 1 M (PCL: 12.0 mm); Lillie Seamount, Kermadec Ridge, 35.857–35.857° S, 178.448–178.443° E, 1237–1460 m; 19 March 2011; RV Tangaroa Stn. TAN1104/124, ‘seamount’ sled; NIWA 72929.
Diagnosis. Carapace with distinct hepatic groove; scattered small spines on postrostral, cardiac, hepatic and branchial surfaces; cervical groove lined with distinct spines; antennal spine and anterolateral spines small but distinct. Rostrum at least reaching distal margin of basal article of antennular peduncle; with 8–10 dorsal teeth. Epistome anteriorly produced, with small anterior teeth. Second and third pleonite with blunt articular knob. Fourth to sixth pleonites smooth on dorsal midline; one or more small spine along each posteroventral margin. Telson broadly rectangular, about twice as long as wide; regular row of teeth along posterior margin. Ocular peduncle with 2–3 granules on dorsal surface. Antennule basal article unarmed on mesial margin; stout stylocerite. Antenna basal article with distinct ventral spines; basicerite with scattered spines on ventral surface; scale with 4–9 lateral teeth along distal half of margin. First maxilliped distal article with sharp distal spine; single arthrobranch developed. Second maxilliped with single arthrobranch and well-developed podobranch; epipod present. Third maxilliped with well-developed setiferous organ; lacking epipod; with paired arthrobranchs. First pereiopod with setiferous organ on propodus only. Second pereiopod similar to first, 1.5 × longer and stronger. Third pereiopod nearly entirely glabrous and smooth; fixed finger unarmed on distoventral margin; palm with few to many minute granules scattered along distoventral portion; ischium with distinct distodorsal spine, with row of ventral spines; coxa mesially granulate. Fourth and fifth pereiopods dactyli with ventral unguis bearing a number of small ventral teeth. First four pereiopods with paired arthrobranchs.
Etymology. Named after the German Research Vessel Sonne that collected the first specimens assigned to this new species during the 2015 PoribacNewZ voyage SO254. Used as a substantive in apposition.
Body large, robust, surface generally glabrous.
Rostrum narrowly triangular in dorsal view, one-third (0.33) as long as PCL, horizontal, barely reaching to distal margin of second article of antennular peduncle; dorsal margin with seven small teeth and posterior blunt eminence at level of posterior orbital margin; ventral margin with one small tooth close to rostral tip; ventrolateral ridges with 1–2 (right-left) small teeth on distal half.
Carapace glabrous, slightly inflated. Cervical and branchiostegal grooves distinct. Gastric, hepatic, and anterior portions of cardiac and branchial regions armed with numerous scattered small spines. Orbital margin concave, inferior orbital angle slightly produced and furnished with 1–2 small antennal spines. Small branchiostegal spine present. Pterygostomian angle round, with series of five small spines on margin.
Thoracic sternites 6–8 anteriorly rounded, minutely serrate but unarmed; sternite six posterior margins evenly concave.
Pleonal somites glabrous, surfaces smooth. First pleonite shortest; divided into two sections by distinct transverse carina; ventral margin continuous, straight. Second and third pleonite subequal in length, with feeble transverse carina and small articular knob; ventral margin rounded, left side with slight indication of a granule. Fourth and fifth pleonites each with small sharp or blunt posteroventral tooth on broad, round margin; dorsally smooth and unarmed. Sixth pleonite dorsally smooth; posterolateral process straight, angular, with small, sharp spine at corner; posterior margin smooth. Pleonal sternites unarmed.
Telson about 1.9 times as long as broad, subquadrangular, posterior margin shallowly convex, with long plumose setae; dorsolateral ridges distinct, with nine spines each, proximalmost largest and placed mesially; lateral margins relatively straight, slightly constricted proximally, each side armed with a subproximal spine and nine lateral spines; posterior margin with 10 small pines (4–5 lateral and one median spine).
Eye well developed; cornea semiglobular, about half length of ocular peduncle in dorsal view, distinctly inflated, unpigmented; minute mesial proximal spine on both eye stalks, and small distal spine on left eye stalk only.
Antennular peduncle reaching to about middle of antennal scale, does not reach spinose lateral margin of antennal scale. Basal article stout, length-width ratio of 2.7 at midlength, without statocyst, about three times as long as second article; lateral margin concave, distally produced to small, rounded lobe, stylocerite distinct, acute, reaching 1/3 length of basal article; mesial margin almost straight, unarmed other than row of serration where plumose seta are inserted. Second article about 1.5 times longer than distal article, distomesially with two small spines. Distal article as long as wide, unarmed. Flagella slender, about twice as long as peduncle.
Antenna with first article (coxa, bearing antennal gland) mesially carinate, bearing three distinct spines. Basicerite stout; mesially bearing minute (left) or distinct (right) spine, paired spines distolaterally; dorsal and lateral surfaces smooth, minute spine at midlength mesially, scattered spines on ventral surface. Antennal scale broad, 2.6 × as long as wide; lateral margin nearly straight, armed with seven (left) or eight (right) teeth along the distal 0.4 portion, including the distal tooth; dorsal surface with two distinct longitudinal ridges. Carpocerite reaching to distal end of second article of antennular peduncle, unarmed. Flagellum at least 1.7 times the CL.
Epistome narrow, subquadrate, with two small spines at anterolateral angles. Labrum smooth. Paragnaths bilobed, with deep median fissure, distodorsally spatulate.
Mandible robust, fused molar and incisor processes. Molar surface with three teeth; incisor bearing two distal teeth with five small proximal teeth. Palp well-developed, 3-segmented; proximal article shortest, without setae; middle article with few setae on flexor and more, longer setae on extensor margins; distal article suboval, slightly longer than intermediate article (measured along extensor margin), densely setose.
Maxillule with simple palp, with few terminal setae; distal endite broad, round, with numerous simple setae and seven slender spines; proximal endite oval, with simple setae distally.
Maxilla with palp stout, tapering, with plumose setae, falling short of end of scaphognathite; distal and proximal endites both deeply bilobed, with numerous plumose setae; scaphognathite well developed, anterior portion longer than posterior portion, about three times longer than broad, with dense fringe of plumose setae along entire margin.
Branchial formula summarized in
Table 2.
First maxilliped with 2-segmented palp, bearing long plumose setae; distal segment broad, with distal blunt spine; proximal article stout, about 1.6 × distal article in length. Distal endite large, subtriangular, densely setose; proximal endite deeply bilobed, with distal setae. Exopod slender, with long plumose setae. Epipod well developed, subequally bilobed. Arthrobranch present.
Second maxilliped with 5-segmented endopod; dactylus sub-oval, tapering distally, about twice as long as broad, with dense setae on flexor margin; propodus subequal in length to dactylus, densely setose along flexor margin; carpus triangular, about three-forth length of propodus measured at mid-line, with long distodorsal setae; merus nearly straight, about 1.8 × length of propodus, nearly 4 times longer than broad, with row of setae along mesial margin and sparse short setae on surfaces; ischium and basis not fused, each about 0.2 × meral length and long setae along mesial margin. Coxa with mesial spine; oval epipod, podobranch and arthrobranch. Exopod absent.
Third maxilliped endopod 5-segmented, slender; dactylus narrow, about 4.5 times longer than broad (at mid-length), setose; propodus about twice as long as dactylus, with setiferous organ along distal two-thirds of flexor margin; carpus subequal in length to propodus; merus longest, about 1.5 × carpal length, minute distolateral spine on left, absent on right; ischium broadest, 0.8 × meral length, small distolateral spine on both sides; all segments with long setal fringe along flexor margin; basis short. Coxa without epipod, arthrobranchs present. Exopod absent.
First pereiopod slender, just overreaching antennal scale when extended, glabrous and sparsely setose; dactylus about 0.5 × palm length; palm subcylindrical, tufts of long setae dorsodistally and along ventral margin of fixed finger; carpus 3 × palm length, ventral carpo-propodal setiferous organ more pronounced on propodus, very weak on carpus; merus about 0.8 × carpal length; ischium one-third meral length. Basis and coxa short, coxa with sharp mesial spine. Epipod absent.
Second pereiopod similar to first, 1.5 × longer and stronger, sparsely setose; dactylus 0.4 × palm length, distal tip formed into a strong corneous spine, tips of fingers cross when chela closed, cutting edges entire; propodus with a few tufts of setae distally and along fixed finger; carpus about 2.2 × palm length; merus 0.8 × carpal length; ischium one-third length of merus. Basis and coxa short; coxa with mesial spine. Epipod absent.
Third pereiopod largest, subequal and similar, about 2 × CL, overreaching the tip of antennal scale by length of chela, very sparsely setose except for a few distal tufts of setae along fingers; dactylus about 0.5 × palm length, ending in strong, hooked tip, cutting edge with narrow trench along distal half, with strong, hook-shaped tooth at mid-length, otherwise smooth; propodal cutting edge with sharp corneous spine distally, followed by narrow trench along distal half, rounded tooth at mid-length and molariform process with numerous tiny teeth at proximal third; outer margin of fixed finger unarmed, bearing tufts of long setae; palm sub-cylindrical, 3.3 times as long as broad, dorsal margin smooth, with few scattered small granules on surface, along ventral margin and more dense across inside palm surface; carpus about 0.5 × palm length, narrowing proximally, without spines; merus subequal in length to palm, five times longer than broad in lateral view, proximally compressed laterally, with small distoventral tooth, dorsally with few distal granules and ventral ridge with row of small teeth and granules, otherwise smooth; ischium about three-fourth length of carpus, laterally compressed, with sharp distodorsal spine and ventral row of spines, distalmost strongest. Basis and coxa short; coxa mesially granulate. Epipod absent.
Fourth and fifth pereiopods long and slender, similar, sparsely setose; dactylus about one-fourth the length of propodus, biungulate, both unguis clearly demarcated, with small, irregularly shaped accessory tooth on ventral margin proximally; propodus not subdivided, about 0.4 × carpal length, pereiopod 5 propodus 1.1 × wider than pereiopod 4 propodus, with single row of 21–22 (P4) and 24–25 (P5) movable spines along entire flexor margin, both margins with few long, simple and very few plumose setae; carpus longest, not subdivided, with movable spine at distoventral angle; merus about 0.8 × carpal length, unarmed; ischium less than half length of merus, unarmed. Basis and coxa short, coxa with blunt, serrated distomesial tooth. Epipod absent.
First pleopod uniramous, second to fifth biramous, all lacking appendices, unarmed.
Uropod well developed, about as long as telson. Protopod stout, with sharp posterolateral spine. Exopod broader than endopod; lateral margin slightly convex, with 17 (left) and 19 (right) teeth along distal three-fourth of margin, distal margin shallowly convex, dorsal surface with two distinct longitudinal ridges. Endopod simple, unarmed, surface with 1 longitudinal ridge. Exopod and endopod fringed with dense, plumose seta.
Eggs: The ovigerous female incubates around 200 eggs of diameters 1.8 × 2.2 mm.
Body smaller than female holotype.
Rostrum proportionally longer, 0.4 × PCL, dorsal margin with eight small teeth in addition to proximal shallow rounded eminence at level of posterior orbital margin; ventral margin with two teeth; laterally unarmed.
The carapace less inflated, distribution of spines similar, but spines generally longer with exception of antennal spine smaller, otherwise similar to holotype.
Sixth thoracic sternites with narrowly triangular process, anteriorly furnished with sharp spine each. Seventh and eight sternites broadly rounded, unarmed.
Pleonal somites similar to holotype; first pleonite with pronounced anteroventral spine; ventral margins of tergites 3–6 more angular and with sharper distal spine each; dorsally smooth. Fifth pleomere with small median spine distally, directed posteriorly.
Telson similar, dorsal ridges with 6–7 spines, laterally with 7–8 spines; distal margin with 9 small spines (median spine with four lateral pairs).
Eye similar, eye stalk with 2–3 dorsal spines.
Antennule similar, second article with single, small distomesial granule only.
Antenna with first article (coxa) bearing strong mesial tooth (not carinate). Basicerite similar. Antennal scale with 4–5 lateral spines.
Epistome with two pairs of small spines and smooth median margin.
Mouthparts as for female holotype. Left third maxilliped merus with sharp distolateral spine, merus unarmed (right third maxilliped missing).
Pereiopod 1, 2, 4 and 5 as for female holotype. Pereiopod 3 slightly longer, 2.5 × CL; the palm is slightly more massive at 2.7–2.8 times longer than wide (left–right); ischium with double distodorsal spine, otherwise as holotype.
Uropodal exopod with 14–15 teeth along lateral margins.
Measurements. CL: 16.3–18.5 mm (M), 16.6–[20.0] mm (F), PCL: 12.0–11.4 mm (M), 11–[
15] mm (F), TL: 38.5–46.0 mm (M), 40.2–[53.2] mm (F). Holotype measurements are given in square brackets.
Distribution. Confirmed in New Zealand, 1184–1460 m. Sequence only, pending morphological examination: Vanuatu, 1210–1250 m (MNHN-IU 2013-19622).
Coloration. Collection notes are retained as “white” (NIWA 127110) and “pink” (NIWA 127111).
Hosts. The pair of type specimens collected by ROV from the Southern Kermadec Ridge was taken with a sponge host of
Corbitella sp. nov. (det. Henry Reiswig,
Figure 15). This genus of glass sponges was recently reported as a host for the first time by Kou et al. [
35] and this is the second record of a
Spongicoloides species with
Corbitella. An unidentified species of
Farrea glass sponge is the only sponge collected at the same station with NIWA 72929 and is the most likely host at present.
Variation and remarks. Two further specimens are reported in addition to the pair of type specimens, displaying a limited amount of variation: NMNZ CR.019650 appears more spinose both considering the surface of the carapace and rostrum, the antennal peduncle and scale and the uropodal endopod (
Figure 16 and
Figure 17). The epistome of the holotype is distinctly bispinose anteriorly, but all other specimens have more, indistinct spines distally. The palp of the first maxilliped bears a long and slender distal spine in this female specimen, which is shorter and blunter in all others, including the female holotype. Saito and colleagues [
48] indicated this might be sexually dimorphic when describing
S. iheyaensis. The armature of the third maxilliped ischium and merus varies slightly across specimens but all have a small or distinct distal spine on at least one side or article (NIWA 72929 had a small spine on both merus and ischium of the one remaining third maxilliped). Some (sexually dimorphic and/or allometric) variation is apparent in the proportions of the third pereiopod: it is slightly shorter for the two females (1.7–2.0 × CL) and longer (2.5 × CL) for the male allotype (pereiopod 3 are missing in the male NIWA 72929). Typical sexual dimorphism is evident with the palm more slender in the large holotype female (length-width ratio of 3.3) and proportionally broader (2.7–2.9) for the male allotype and female NMNZ CR.019650 that are both approximately the same size.
Spongicoloides sonne sp. nov. closely resembles the group of species with two arthrobranchs on third maxilliped and pereiopods 1–4, dorsally unarmed uropodal endopod, dorsally smooth pleonites and smooth distoventral margin of pereiopod 3 fixed finger. This includes
S. clarki sp. nov.,
S. novaezelandiae,
S. hawaiiensis Baba, 1983 [
49],
S. weijiaensis and
S. corbitellus.
Spongicoloides sonne sp. nov. can be distinguished from these species by the following fixed characters:
- -
The postrostral, cardiac and at least the anterior branchial carapace surface is covered with small spines; S. hawaiiensis has a nearly smooth carapace and the remaining species only have a few scattered small spinules.
- -
Third maxilliped is lacking an epipod; which all others have present (rudimentary in S. clarki).
- -
Third maxilliped endopod ischium and/or merus with small distolateral spine (typically varies from left to right but present on at least one of the articles); the endopod is unarmed in all other species.
- -
Coxa of third maxilliped and pereiopods 1–2 with distinct mesial spine or process; rounded in all other species.
- -
Rostrum overreaches the basal antennular article in S. sonne, S. novaezelandiae, S. weijiaensis and S. clarki but it falls short of the distal end of the article in S. corbitellus (the rostrum in S. hawaiiensis remains unknown).
- -
Pereiopod 3 ischium with distinct distodorsal spine and row of sharp spines along ventral margin, the distodorsal spine is small in S. novaezelandiae, S. weijiaensis, S. clarki and S. corbitellus and absent in S. hawaiiensis; the ventral margin smooth or irregular but not furnished with sharp spines in all other species.
A notable characteristic that has in the past been rarely considered is the ventromesial ornamentation of the first antennal segment (coxa, bearing the antennal gland) which is distinctly ridged and carinate (as in the holotype), always furnished with at least one distinct spine. In S. novaezelandiae the first antennal article is simple, with only a small spine. This character might hold additional phylogenetic value and could be considered further across the remaining species.
At least one of the specimens presented by Goy [
13] as
S. novaezelandiae from New Caledonia most likely belong to this new species based on the clearly spinose carapace surface and the row of distinct spines illustrated on the third pereiopod ischium of MNHN Na-11999 (Goy [
13]:
Figure 9; MNHN-IU-2013-19487). Most of the remaining material presented by Goy [
13] are referable to
S. weijiaensis (see comments under
S. novaezelandiae above).
DNA sequence data. A clade formed by 3–4 sequences were monophyletic for all genes considered (
Figure 2). This included a sequence from a specimen from Vanuatu (1210–1250 m) held at the MNHN (IU-2013-19622) which is considered conspecific and pending more detailed examination.
CO1: intra-specific divergences between 0–1.2%; intra-generic divergences range from 4.2% (Spongicoloides clarki sp. nov.) to 7.3% (S. weijiaensis).
16S rRNA: intra-specific divergences between 0–0.3%; intra-generic divergences range from 3.3% (Spongicoloides weijiaensis) to 16.7% (S. iheyaensis).
Genus
Spongiocaris Bruce & Baba, 1973 [
21].
3.2.5. Spongiocaris antipodes sp. nov.
In (
Figure 1,
Figure 2,
Figure 18,
Figure 19,
Figure 20,
Figure 21 and
Figure 22).
Material examined. Holotype: F ov. (PCL: 7.7 mm); West Norfolk Ridge, 34.342° S, 168.387° E, 382–390 m; 2 June 2003; NORFANZ Stn. TAN0308/139; NMNZ CR.025704.
Allotype: M (PCL: 5.2 mm); West Norfolk Ridge, 34.285° S, 168.358° E, 785–800 m; 2 June 2003; NORFANZ Stn. TAN0308/141; NMNZ CR.019492.
Paratype: 1 M (poor condition, PCL: ~6.2 mm); station details as for holotype; NMNZ CR.019493. 1 F (PCL: 8.0 mm); Seamount No. 986, off Hawkes Bay shelf, 39.991° S, 178.215° E, 792 m; 9 February 2017; method: manipulator arm; R/V Sonne, ROV
KIEL 6000, cruise: SO254, dive: 36 ROV 10; NIWA 127133.
Other material: Norfolk Ridge. 1 F ov. (PCL: 12.9 mm), 1 M (PCL: 8.7 mm), Wanganella bank (International Waters); 33.5–33.4° S, 167.7–167.6° E, 677–546 m; 11 November 2013; SOP Stn. TRIP3933/23; bottom longline; NIWA 88622. 2 M (PCL: 6.4, 7.3 mm); southern Norfolk Ridge, NW of Three Kings Islands, 33.390–33.391° S, 170.210–170.196° E, 469–490 m; 1 June 2003; beam trawl; NORFANZ Stn. TAN0308/136; NMNZ CR.019494 (with Regadrella sp.). 1 F ov. (PCL: 8.0), 2 M (PCL: 7.5, 5.2 mm); southern Norfolk Ridge, NW of Three Kings Islands, 33.390–33.391° S, 170.210–170.196° E, 469–490 m; 1 June 2003; beam trawl; NORFANZ Stn. TAN0308/136; NMNZ CR.019491. 3 F (PCL: 7.8, 7.9, 8.7 mm), 1 M (PCL: 6.3 mm); southern Norfolk Ridge, Reinga Ridge, 33.390–33.396° S, 170.190–170.203° E; 469–526 m; 31 May 2003; NORFANZ Stn. TAN0308/126; NMNZ CR.019259. Three Kings Ridge. 1 F (PCL: 6.0 mm), 2 M (PCL: 7.2, 4.6 mm); Seamount 148, 31.980° S, 174.265° E, 700 m; date unknown; Stn. Z9026; NIWA 135621. Kermadec Ridge. 1 M (PCL: 7.5 mm); Raoul Island, 6.4 km NNE of Herald Islet, 29.20° S, 177.82° W, 1189–1225 m; 5 April 1973; NMNZ Stn. 73312: NMNZ CR.016806; “inside glass sponge”. Hikurangi Margin. 1 F ov. (damaged, TL ~ 33 mm), East Coast, E of Cape Kidnappers, 40.03° S, 178.06° E, 935 m; 28 Aug 1986; bottom trawl; RV James Cook; Stn. J10/40/86; NMNZ CR.005952 (collection note: “from Euplectella”). 1 F ov. (poorly preserved), 1 M (PCL: 7.0 mm); SE of Cape Kidnappers, 40.02° S, 178.08° E, 840 m; 28 August 1986; bottom trawl; RV James Cook; Stn. J10/39/86; NMNZ CR.004792.
Diagnosis. Small commensal spongicolid shrimp so far known to be associated with euplectellid glass sponges; body slightly depressed. Carapace with distinct cervical groove, each side furnished with 6–10 small spines; small branchiostegal spine and antennal spine present or absent; 2–3 pairs of postrostral spines; typically two pairs of supraorbital spines; pterygostomian angle furnished with a few small spines; a few anterolateral spines always present; hepatic spines typically present. Rostrum length reaching to distinctly overreaching last antennular peduncle article, entire dorsal margin with spines with 5–11 dorsal, 1–3 ventral, 0–3 lateral spines; cornea unpigmented. Antennal scale subrectangular, with 4–9 spines along the lateral margin. Epistome nearly always unarmed, may have single anterior spine; endopod of maxillule unarmed. Third maxilliped setiferous organ covering nearly entire length of propodal ventral margin; epipod always absent. First pereiopod with well-developed setiferous organ. Third pereiopod robust; carpi unarmed except for low blunt distal spines; meri smooth or with dorsal row of low spines; palm with a few scattered spines along distal portion of outer surface, at least three times longer than wide; fixed finger with dense fringe of setal brushes along two-thirds of ventral margin, unarmed. Fourth and fifth pereiopods with carpi distinctly segmented by 1–3 sutures; coxa may be armed with short distomesial spinose ridge (males) or granular (females); all pereiopods lack epipods. Posterior margin of fifth pleonal tergite ending in a few spines, posterior margins of pleura irregular; sixth tergite with 3–4 posterior dorsal spines, surface smooth, posterior margin of pleura with 1–3 spines. Telson with 6–8 spines along each lateral margin. Uropodal endopodite with two terminal dorsal hairs.
Etymology. Named antipodes, an archaic vernacular for Australia and New Zealand, as this species is so far restricted to the southwestern Pacific region. The term is used as a noun in apposition.
Body robust, surface generally glabrous.
Rostrum narrowly triangular in dorsal view, one-third (0.3) as long as PCL, horizontal, nearly reaching to distal end of antennular peduncle; dorsal margin with 7 small teeth or granules and small posterior blunt eminence at level of posterior orbital margin; ventral margin with three small teeth in distal one-third; ventrolateral ridges with 1–2 (right-left) small teeth on distal half.
Carapace glabrous, not inflated. Cervical and branchiostegal grooves distinct, seven (right)–eight (left) distinct spines line cervical groove. Postrostral region with two small submedian spines on either side; two pairs of postorbital spines; hepatic region smooth; four pairs of anterolateral spines; two small spines on anterior branchial region; otherwise carapace surface is smooth. Orbital margin concave, inferior orbital angle slightly produced and furnished with one small antennal spine (left) or granule (right); branchiostegal spine small to minute; pterygostomian angle round, with 1–2 spines on margin.
Thoracic sternites 6–8 anteriorly rounded, minutely serrate but unarmed; sternite 6 posterior margins evenly concave.
Pleonal somites glabrous, surfaces smooth. First pleonite shortest; divided into two sections by distinct transverse carina; ventral margin continuous, straight. Second to fourth pleonites subequal in length, with feeble transverse carina and blunt articular knob; ventral margin rounded. Fifth pleonite with two small posteroventral granules on broad, round margin; dorsal surface smooth, unarmed; posterior margin with four small spines. Sixth pleonite dorsally smooth; posterolateral process straight, angular, with small spine at corner; posterior margin with three small spines. Pleonal sternites unarmed.
Telson about twice as long as broad, subquadrangular, posterior margin distinctly convex, with long plumose setae; dorsolateral ridges distinct, with six spines each, proximalmost largest and placed mesially; lateral margins relatively straight, slightly constricted proximally, each side armed with a subproximal spine and six lateral spines; posterior margin with three small spines (one median spine and one lateral pair).
Eye well developed; cornea semiglobular, about half length of ocular peduncle in dorsal view, moderately inflated, unpigmented; unarmed.
Antennular peduncle nearly reaching to middle of antennal scale, does not reach spinose lateral margin of antennal scale. Basal article stout, length-width ratio of 2.0 at midlength, without statocyst, about three times as long as second article; lateral margin concave, distally produced to small, rounded lobe, stylocerite distinct, acute, reaching one-third length of basal article; mesial margin almost straight, unarmed other than row of serration where plumose seta are inserted. Second article slightly longer than distal article, unarmed. Distal article as long as wide, unarmed. Flagella slender, about twice as long as peduncle.
Antenna with first article (coxa, bearing antennal gland) with 1–2 mesial spines. Basicerite stout; 1 mesial spine proximal to base of carpocerite; 2–3 small spines on ventral surface; dorsal and lateral surfaces smooth. Margins with paired spines distolaterally; small spine below ventral angle; small spine below mesial angle (
Figure 18H inset). Antennal scale broad, 2.5 × as long as wide; lateral margin slightly concave, armed with 4 (left) or 6 (right) small teeth along the distal 0.4–0.5 portion, including the distal tooth; dorsal surface with two distinct longitudinal ridges. Carpocerite not reaching to distal end of second article of antennular peduncle; first article with small mesial spine; second article with minute distoventral spine. Flagellum at least as long as CL.
Epistome narrow, inflated, subtriangular, unarmed. Labrum smooth, with median ridge. Paragnaths bilobed, with deep median fissure, distodorsally spatulate.
Mandible robust, fused molar and incisor processes. Molar surface with three blunt teeth; incisor with irregular teeth. Palp well-developed, 3-segmented; proximal article shortest, without setae; middle article with few distal setae; distal article suboval, slightly longer than intermediate article (measured along extensor margin), densely setose.
Maxillule with simple palp, with few terminal setae; distal endite broad, round, with numerous simple setae and eight slender spines; proximal endite oval, with simple setae distally.
Maxilla with palp stout, tapering, with plumose setae, falling short of end of scaphognathite; distal and proximal endites both deeply bilobed, with numerous plumose setae; scaphognathite well developed, anterior portion longer than posterior portion, about 3 times longer than broad, with dense fringe of plumose setae along entire margin.
Branchial formula summarized in
Table 2.
First maxilliped with 2-segmented palp, bearing long plumose setae; distal article longer than broad, with distal spine; proximal article stout, about 1.5 × distal article in length. Distal endite large, subtriangular, densely setose; proximal endite deeply bilobed, with distal setae. Exopod slender, with long plumose setae. Epipod well developed, subequally bilobed. Arthrobranch well developed.
Second maxilliped with 5-segmented endopod; dactylus sub-oval, tapering distally, with dense setae on flexor margin; propodus twice as long as dactylus, densely setose along flexor margin; carpus triangular, about two-third length of propodus measured at mid-line, with long distodorsal setae; merus nearly straight, about 1.8 × length of propodus, 4 times longer than broad, with row of setae along mesial margin and sparse short setae on surfaces; ischium and basis not fused, ischium slightly longer, about one-fourth meral length, long setae along mesial margin. Coxa mesially produced to blunt process; exopod well developed; oval epipod present, podobranch and arthrobranch well developed.
Third maxilliped endopod 5-segmented, slender, unarmed; dactylus narrow, distally tapering, setose; propodus about twice as long as dactylus, with setiferous organ along entire flexor margin; carpus subequal in length to propodus; merus 1.5 times longer than carpus, about as long as ischium; all segments with long setal fringe along flexor margin; basis short. Coxa mesially angular, not produced to spine, without epipod, arthrobranchs well developed. Exopod absent.
First pereiopod slender, glabrous and sparsely setose, slightly overreaching antennal scale when extended. Dactylus about 0.4 × palm length; palm subcylindrical, tufts of long setae dorsodistally and along ventral margin of fixed finger; carpus about 3 times palm length, ventral carpo-propodal setiferous organ well developed on both articles; merus about 0.7 × carpal length, about 3 times longer than ischium. Basis and coxa short, unarmed. Epipod absent.
Second pereiopod similar to first, 1.5 × longer and stronger, sparsely setose; dactylus 0.4 × palm length, distal tip formed into a strong corneous spine, tips of fingers cross when chela closed, cutting edges entire; propodus with a few tufts of setae distally and along fixed finger; carpus about 1.8 × palm length; merus 0.8 × carpal length; ischium 0.4 × merus. Basis and coxa short; coxa unarmed. Epipod absent.
Third pereiopod largest, subequal and similar, about 2 × CL, sparsely setose except for a few distal tufts of setae along fingers; dactylus about 0.5 × palm length, ending in strong, hooked tip, cutting edge with narrow trench along distal half, with blunt, trianguloid tooth at distal quarter, preceding fossa not pronounced; propodal cutting edge with sharp corneous spine distally, followed by narrow trench along distal half, distinct fossa to accommodate dactylar tooth at distal third, followed proximally by flat molariform process with numerous tiny teeth; outer margin of fixed finger unarmed, distally bearing tufts of long setae; surface with small proximal spine; palm sub-cylindrical, 3.4 times as long as broad, dorsal margin smooth, with 2–3 small granules on distal surface, very few small granules on inside palm surface; carpus about 0.3 × palm length, narrowing proximally, without spines; merus three-fourth length of palm, 5 times longer than broad in lateral view, proximally compressed laterally, margins entirely smooth; ischium about as long as carpus, laterally compressed, unarmed. Basis and coxa short; coxa mesially granulate. Epipod absent.
Fourth and fifth pereiopods long and slender, similar, sparsely setose; dactylus (including spines) about one-fourth the length of propodus, biungulate, both unguis clearly demarcated, with small, simple or distally faintly bifurcate accessory tooth on proximal ventral margin; propodus not subdivided, about 0.4 × carpal length, pereiopods 4 and 5 propodi similar in width, 5.7 times longer than wide, with single row of 16–20 movable spine along entire flexor margin, both margins with few long, simple and very few plumose setae; carpus longest, 2.2–2.5 × propodus, with movable spine at distoventral angle, one median suture present; merus about 0.8 × carpal length, unarmed; ischium less than half length of merus, unarmed. Basis and coxa short, coxa mesially granular but unarmed. Epipod absent.
First pleopod uniramous, second to fifth biramous, all lacking appendices, unarmed.
Uropod well developed, about as long as telson. Protopod stout, with sharp posterolateral spine. Exopod broader than endopod; lateral margin slightly convex, with 12–13 small teeth along distal three-fourth of margin, distal margin shallowly convex, dorsal surface with two distinct longitudinal ridges. Endopod simple, unarmed, surface with one longitudinal ridge. Exopod and endopod fringed with dense, plumose seta.
Eggs: The ovigerous female retained 10 sub-oval eggs of diameters 1.5 × 2.0 mm.
Body smaller than female holotype.
Rostrum proportionally longer, 0.5 × PCL, distinctly overreaching the antennular peduncle, dorsal margin with 8 small teeth in addition to proximal shallow rounded eminence at level of posterior orbital margin; ventral margin with two teeth; with 3 small ventrolateral teeth.
The carapace is slightly less inflated, the distribution of spines is similar, the numbers vary slightly: 6–7 spines along the branchiostegal groove, three small postrostral spines, 3–5 anterolateral marginal spines. The two prominent postorbital spines match the holotype and are considered diagnostic.
Thoracic sternites 6–8 anteriorly produced, with spines, the coxa of the pereiopods and furnished with denticulate processes.
Pleonal somites similar to holotype; posteroventral margins of tergite 4 with 1–2 spines, tergite 5 with 3–4 small spines, posterior margin with rows of small spines; dorsal surfaces smooth. Fifth pleomere with small median spine ventrodistally, directed posteriorly.
Telson similarly shaped, dorsal ridges with 5–6 spines, laterally also with 6 spines; distal margin with 6 small spines (median spine with 2 and 3 lateral spines).
Antennule and antenna of similar shape and proportions; antennal basicerite with additional distoventral and distolateral spine, mesial angle unarmed. Antennal scale slightly more slender at 2.8 (length-width ratio), with 6 lateral spines.
Epistome similarly inflated, smooth.
Mouthparts as for female holotype.
Pereiopod 1 and 2 as for female holotype. Pereiopod 3 slightly more robust, similar length with 2.0 × CL; dactylus 0.6 × palm; palm slightly more massive at 2.8–2.9 times longer than wide; merus with distoventral spine and regular dorsal row of granules and small spines; ischium smooth as for holotype. Single loose pereiopod 4 or 5 retained for allotype has a simple, sharp accessory tooth on dactylus; propodus with 14 spines along flexor margin; carpus with two sutures.
Uropods as for holotype, except for protopod distally with pair of distolateral spines exopod with 12 and 14 teeth along lateral margins.
Measurements. CL: 9.3–12.3 [10.3] mm (females, this does not include the largest female that has a broken rostrum), 7.4–11.2 mm (males), PCL: 6.0–12.9 [7.7] mm (females) 5.2–8.7 mm (males), TL: 25–37 [25.5] mm (females), 19.5–30.7 (males). Measurements for the holotype are given in square brackets.
Distribution. Southern Norfolk Ridge, West Norfolk Ridge, eastern North Island, New Zealand; 382–1225 m (
Figure 1).
Coloration. Collection note with NMNZ CR.016806 “white shrimp from inside glass sponge”. Live coloration was captured for NORFANZ specimens NMNZ CR.019259 and CR.019494: the body is pale apricot, carapace and most appendages transparent, palm of third pereiopod orange (
Figure 22).
Hosts. Two specimens were collected from inside the euplectellid glass sponge
Regadrella okinoseana (Hexactinellida, Lyssacinosida, Euplectellidae) (NMNZ Cr. 09494, det. M Kelly,
Figure 22). Other collection records retained are: “from
Euplectella” (NMNZ CR.05952) and “from inside glass sponge” (NMNZ CR.016806).
Parasites. One male (NMNZ CR.016806) bears a sacculinid rhizocephalan under the pleon.
Remarks. Multi-gene sequencing revealed a clade that resolved most of the New Zealand specimens of
Spongiocaris as an undescribed species, while it united the type specimens of both
S. yaldwyni and
S. neocaledonensis in a well-supported clade, requiring that the latter be synonymized with the former (see below,
Figure 2). Morphologically, the variability of characters across the material examined rendered it difficult to establish fixed diagnostic characters.
Spongiocaris species are currently separated using differences in the length of the rostrum, distribution of spines on the carapace surface and anterior margin, pleomere and telson, as well as the pereiopods 3–5 [
11]. Examination of a total of 21 specimens assigned to
S. antipodes sp. nov. indicate the following variability: the rostrum is slightly shorter to longer than the antennular peduncle (not reaching to overreaching article 3), 0.3–0.5 × PCL, with 5–11 dorsal, 1–3 ventral and 0–3 lateral teeth; the carapace has 2–3 pairs of small postrostral spines always present, at least one but typically two distinct postorbital spines, 4–10 pairs of postcervical spines are always distinct, occasionally followed by a few scattered dorsal cardiac spinules (
Figure 20A,B), a field of 4–10 distinct anterolateral spines, a few small hepatic and/or branchial spines are usually present (
Figure 20B); an antennal spine on the anterior carapace margin may be absent, minute or well-developed and can vary from side to side (e.g.,
Figure 20B), a small branchiostegal spine may be present or absent and the pterygostomian angle with 1–3 distinct spines; the dorsal surfaces of the pleonites are always unarmed and smooth, the ventrodistal margins of pleura 4–6 usually bear some spines (pleuron 4 may be smooth, pleuron 5 always bears multiple spines and pleuron 6 has at least an acute angle); the telson shape is variable (
Figure 18C,
Figure 20C and
Figure 21L–N), 6–8 lateral spines in addition to subproximal spine, 5–8 dorsal spines and 3–8 small spines along the distal margin, usually arranged with a median spine separated from pairs of laterally-placed spines (with the exception of the larger male, NMNZ CR.019491, that has a continuous row of 6 spines); the third cheliped is about twice as long as CL with limited sexual dimorphism apparent, the fingers are 0.5–0.6 × palm length, margins unarmed, with dense setal brushes distally, along about the distal two-thirds of the margin of the fixed finger, the palm length-width ratio ranges from 2.8 in small to 3.4 in larger specimens, the proximal portion always has 2–7 small granules, the merus is smooth (
Figure 19B) or serrated (
Figure 20O), the ischium is always unarmed with smooth margins; pereiopods 4–5 carpi with 1–3 sutures, propodi with 14–23 movable spines along the entire flexor margin and dactyli biungulate with ventral unguis bearing a variable accessory tooth, from simple hook-shaped (
Figure 21G) to bifurcate (
Figure 21F,I).
Further notable variation is the presence of an anterior spine on the epistome of a male of NMNZ CR.019491, the shape is otherwise the same as in all others, anteriorly inflated and round. The P4–5 propodi usually have between 17–22 spines along the flexor margin, but these can be as few as 12 (e.g., NMNZ CR.019491 and CR.019492)
Based on the combination of these characteristics, S. antipodes closely aligns with S. yaldwyni (to include S. neocaledonensis, see below). These two species appear to be difficult to separate morphologically. Diagnostic characters proposed are:
- -
the presence of postrostal and, typically, at least two pairs of postorbital carapace spines (the postrostral area is usually smooth in
S. yaldwyni but see Goy ([
13]:
Figure 12) and the carapace only has a single postorbital spine). This character can vary from side to side and some small specimens (e.g., NMNZ CR.019494, CR.019491, NIWA 135621) have only a single spine.
- -
the dorsal surface of pleonal tergite 6 is always smooth (S. yaldwyni often has 2–3 median spines, see Figures 24C and 25A, but is smooth in holotype, Figure 23B).
- -
the dense setal brushes along the distal fixed finger of the third pereiopod is distinct along about two-thirds in S. antipodes but reach to 0.7–0.8 portion in S. yaldwyni.
- -
the fourth and fifth pereiopod dactyli are never triungulate in S. antipodes but can be both biungulate and triungulate in S. yaldwyni.
There may be a degree of overlap in these characters and DNA sequencing for confirmation of identification is advised.
DNA sequence data. Between six and nine specimens were sequenced across the genes examined (
Figure 2). In all cases they formed a monophyletic clade with some indication of further intraspecific structure that might warrant further investigation in the future.
CO1: intra-specific divergences between 0.2–2.3%; intra-generic divergences between 4.9–6.7% (Spongiocaris yaldwyni).
16S rRNA: intra-specific divergences between 0.0–1.6%; intra-generic divergences range from 1.9% (Spongiocaris yaldwyni) to 11.4% (S. koehleri).
3.2.6. Spongiocaris yaldwyni Bruce & Baba, 1973
Material examined. Holotype: F (PCL 7.6 mm); Motunau Island (Plate Island), Bay of Plenty, 37.50° S, 176.55° E, 585–622 m; 29 Sep 1962; N.Z. Marine Department Haul 12; NMNZ CR.001888 (collection note: “venus flower basket sponge”).
Other material: Kermadec Ridge. 1 F ov. (PCL: 7.6 mm), 1 M (PCL: 6.6 mm); 41 km NNE of Macauley Island, 29.8617 S, 178.1817 W, 965 m, 27 Jul 1974; NZOI Stn K831; Agassiz trawl; NIWA 10483. 1 F (PCL: 5.0 mm); NE of summit, Clark Seamount, Southern Kermadec Ridge, 36.445–36.442° S, 177.839–177.840° E, 850–927 m; 24 Apr 2012; NIWA Stn. TAN1206/99; Sled, epibenthic; NIWA 83102. Bay of Plenty. 1 M (PCL: 12.7 mm), Whakatane Seamount, 36.790–36.790° S, 177.454–177.456° E, 1160–1155 m; 23 Apr 2012; epibenthic sled; NIWA Stn. TAN1206/90; NIWA 82821. 1 F (crushed carapace, TL~25 mm); ca. 22 km E of Aldermen Islands, 36.96° S, 176.36° E, 803–846 m; 24-Jan-79; RV Tangaroa; NMNZ Stn. 79760; NMNZ CR.009840.
Diagnosis. Small commensal spongicolid shrimp so far known to be associated with euplectellid glass sponges and gorgonian corals; body slightly depressed. Carapace with distinct cervical groove furnished with 4–10 small spines; small branchiostegal spine and antennal spine present or absent; one pair of supraorbital spines; pterygostomian angle furnished with a few small spines; a few anterolateral spines always present; hepatic spines typically present. Rostrum length reaching to distinctly overreaching last antennular peduncle segment, entire dorsal margin with spines with 4–8 dorsal, 0–4 ventral, 0–3 lateral spines; only proximal part of cornea pigmented. Antennal scale subrectangular, with 4–7 spines along the lateral margin. Epistome unarmed; endopod of maxillule unarmed. Third maxilliped setiferous organ covering nearly entire length of propodal ventral margin; epipod present or absent. First pereiopod with well-developed setiferous organ. Third pereiopod robust; carpi unarmed; meri with row of small spines along dorsal margin; palm with a few scattered spines along distal portion of outer surface, at least 3 times longer than wide; fixed finger with dense fringe of setal brushes along more than two-thirds of ventral margin, unarmed. Fourth and fifth pereiopods with carpi distinctly or partly segmented, full suture typically present at approximately mid-point; coxa armed with short distomesial spinose ridge; all pereiopods lack epipods. Posterior margin of fifth pleonal tergite ending in four or five spines, posterior margins of pleura irregular to spinose; sixth tergite with 3–4 posterior dorsal spines, surface smooth or with two or more small spines along mid-line, posterior margin of pleura with 1–2 spines. Telson with 4–10 spines along each lateral margin. Uropodal endopodite with two terminal dorsal hairs.
Measurements. CL: 6.7–15.6 [11.8] mm, PCL: 4.6–12.7 [7.6] mm, TL: 18.0–43.0 mm. Holotype measurements are given in square brackets.
Distribution.Spongiocaris yaldwyni was purportedly endemic to New Zealand with a single record from the Bay of Plenty; 585–622 m. Chen et al. [
17] presented gene sequences for a specimen from Tonga (627–656 m, MNHN-IU-2014-12842). New records extend the regional distribution northwards along the Kermadec Ridge, 850–1160 m (
Figure 1). Considering the synonymy of
S. neocaledonensis, the distribution is extended to include New Caledonia, Norfolk Ridge, Loyalty Islands; 460–970 m.
Coloration. Unknown.
Hosts. The holotype was collected from within the hexactinellid sponge
Regadrella okinoseana [
21]. Goy [
13] reported
S. neocaledonensis from the hexactinellid sponge
R. okinoseana and on a gorgonian octocoral.
Remarks. The original description of
Spongiocaris yaldwyni is comprehensive and is not repeated here, the material presented is the first since the species was described from the single specimen [
21]. The holotype (NMNZ CR.001888) is in poor condition and the body is deformed but allowed the verification of some characteristics: the epistome is unarmed, anteriorly truncate, and the labrum rounded (
Figure 23D); all pleonal tergites have smooth surfaces, the posterior margins of pleonites 5–6 bear 3–4 median spines (
Figure 23B), the posterior margins of the pleura bear spines (pleurae 4 and 5) or are angular (pleuron 6) (
Figure 23C); sternites 6–8 are medially acute and the coxa of pereiopods 3–5 bear mesial spines along their anterior corners (
Figure 23F); third maxilliped has a setiferous organ on nearly entire flexor margin of the propodus; and the first pereiopod has a well-developed setiferous organ on the proximal palm and distal carpus (
Figure 23G); the third pereiopod merus is distinctly serrate along the dorsal (extensor) margin and the setal brushes extend along more than 0.6 portion of the fixed finger (
Figure 23H). The fourth and fifth pereiopods carpi clearly have a suture at approximately mid-length, which was not illustrated by Bruce & Baba [
21].
The type description did not mention the sex of the specimen, it bears gonopores on the coxae of the third pereiopods but also possesses the single forward curving ventral medial spine on the fifth pleomere that is typical of males (
Figure 23C,F). The female (NIWA 83102), however, lacks a spine.
Three further samples are presented here for the region that all share the long rostrum (overreaching the second antennular article). The rostral ornamentation can be expanded to 6–[8] dorsal, 2–[6] ventral and [0]–3 lateral spines (holotype details are presented in square brackets); all specimens have a single pair of postorbital spines, NIWA 10483 and 83102 have 2–3 minute postrostral spines (smooth in the holotype), the carapace has a hepatic spine in the holotype and the larger female of NIWA 10483 but is smooth in NIWA 83102 (
Figure 24A); the surface of pleonite 6 bears dorsal spines in all other specimens (
Figure 24B and
Figure 25A), but is smooth in the holotype. The antennal scale is relatively slender in the holotype (3 × longer than wide), similar to NIWA 83102, but is relatively broader in NIWA 10483 with ratios of 2.6 and 2.8, the lateral margin has 3–7 spines; the basicerite has only a single lateral spine in NIWA 83102 (
Figure 24D,E). The sternites of the females are typically rounded and the coxa are produced to distinctly projected and granulate mesial processes in all cases (e.g.,
Figure 24F). The third maxilliped is lacking an epipod in all specimens, the propodus is furnished with a dense fringe of seta (setiferous organ). The third pereiopod is similarly glabrous in both males and females except for the presence of small spines on the outer palm, at the base of the fixed finger, the setal brushes are distinct and extend up to 0.8 × the length of the fixed finger in NIWA 10483 and 83102 (
Figure 24O). The lose walking leg of NIWA 83102 is lacking a suture on its carpus (
Figure 24Q) but 1–3 sutures are present in all pereiopods 4–5 of NIWA 10483, all specimens have dactyli with small accessory teeth on the ventral unguis, either sharp, simple, or divided (
Figure 24Q inset), NIWA 82821 has all pereiopod 4–5 dactyli triungulate (
Figure 25D). The holotype and two most recently collected specimens (NIWA 82821 and 83102) are closely related genetically (p-distances of ≤1.6% for 16S rRNA fragment, see below) and are genetically distinct from
S. antipodes.DNA sequence analyses indicate that the holotype sequence for
S. yaldwyni was in all cases near identical to the type series of
S. neocaledonensis Goy, 2015 [
13] (see
Figure 2 and comments below). Goy [
13] noted the close overall appearance between these two species but separated them based on the presence (
S. neocaledonensis) and absence (
S. yaldwyni) of an epipod on the third maxilliped and proposed
S. neocaledonensis as distinct from
S. yaldwyni (and all other congeners) based on the segmentation of the carpus of the last two pairs of pereiopods 4–5 (present in
S. neocaledonensis and absent in
S. yaldwyni), the presence of the third maxilliped’s setiferous organ (absent in
S. yaldwyni), and the dense fringe of setal brushes on the third pereiopod propodus (fixed finger) (absent in
S. yaldwyni). As noted above, upon re-examination of the holotype and additional material of
S. yaldwyni, characters overlap considering the presence of sutures on the pereiopod 4–5 carpi of nearly all specimens (absent on single remaining leg of NIWA 83102,
Figure 24Q), presence of dense setal brushes on third maxilliped and pereiopod 3. However, none of the material examined had even rudimentary epipods on the Mxp3 and this character remains enigmatic. Nevertheless,
Spongiocaris yaldwyni and
S. neocaledonensis are here considered synonymous.
Most of the New Zealand specimens (the exception being the holotype) bear a median row of two small spines on pleonal tergite 6, none are as spinose as figured for
S. neocaledonensis male holotype (Goy [
13]:
Figure 12). In nearly all New Zealand specimens, the pereiopod 4–5 dactyli are biungulate, with a small accessory tooth on the ventral unguis (the exception being NIWA 82821 which has a triungulate dactylus as illustrated for
S. neocaledonensis,
Figure 25D). This character has been found to be variable in other species, e.g., the stenopodid
Stenopus goyi Saito, Okuno & Chan 2009 [
52], which has the ambulatory dactyli from simple to triungulate, differing even among the pereiopods of the same specimen [
52]. Variation in spination across this species is now considered as follows: the rostrum has 4–8 dorsal, 0–4 ventral and 0–3 ventrolateral spines; the carapace spination is somewhat variable with some hepatic, postrostral, minute cardiac or branchial spine, 4–10 postcervical spines are always distinct, as is one pair of postorbital spines; the telson has 4–10 spines along the lateral margin, 4–9 spines along dorsal carinae and 3–12 posterior teeth; the antennal scale has 4–7 spines along the lateral margin and the pereiopods 4–5 propodi have 8–21 movable spines along the flexor margin.
The combined distribution range now encompasses New Caledonia, the Norfolk and Kermadec Ridges, which aligns with those of other southwestern Pacific subtropical to temperate decapods (e.g., [
53,
54]), with an overall depth range of 460–1160 m. Both species were first described from the same hexactinellid sponge host species (
Regadrella okinoseana).
DNA sequence data. The holotype of
S. yaldwyni and nine type specimens of
S. neocaledonensis were successfully sequenced, including the holotype, allotype and seven paratype samples. The sequences presented by Chen et al. [
17] from Tonga are confirmed as belonging
S. yaldwyni (
Figure 2,
Table 1). Both genes resolved these specimens as belonging to a single clade, distinct from
Spongiocaris antipodes sp. nov., however, substantial internal differences remain within this group that warrant future investigation.
CO1: intra-specific divergences between 0.2–4.2%; intra-generic divergences between 4.9–6.7% (Spongiocaris antipodes sp. nov.).
16S rRNA: intra-specific divergences between 0.0–2.6%; intra-generic divergences range from 1.9% (Spongiocaris antipodes sp. nov) to 11.4% (S. koehleri).
Family STENOPODIDAE Claus, 1872 [
55]
Genus
Stenopus Latreille, 1819 [
56]
3.2.7. Stenopus hispidus (Olivier, 1811)
In (
Figure 26).
Palaemon hispidus Olivier, 1811 [
23]: 666.
Stenopus hispidus.—Latreille 1819 [
56]: 71.—Holthuis 1946 [
57]: 12 (with references and complete synonomy).—Yaldwyn 1968 [
22]: 278.—Doak 1971 [
58]: pl. 39D.—Yaldwyn 1974 [
59]: 1044.—de Saint-Laurent & Cleva 1981 [
60]: 157.—Goy 1992: 100 [
61].—Poore 2004 [
62]: 149.—Saito et al. 2009 [
52]: 1009.—Yaldwyn & Webber 2011 [
63]: 183 (list).—Webber et al. 2010 [
64]: 225 (list).—Goy 2010 [
3]: 215.—De Grave & Fransen 2011 [
42]: 253.—Goy 2015 [
13]: 336.
Material examined. Kingdom of Tonga. 1 M (PCL: 11.5 mm); Tongatapu Island, Stn. I173, 21.130° S, 175.183° W, 0 m; 8 June 1976; underwater hand sampler; NIWA 10308. Lord Howe Island (Australia EEZ). 1 M (PCL: 14.5 mm); Stn. P31, 29.007° S, 167.915° E, 4–14 m; 28 January 1977; scuba; NIWA 10604. Kermadec Islands. 1 M (PCL: 9.8 mm), 1 F ov. (PCL: 12.6 mm); Raoul Island, Stn. TAN1612/37, 29.226° S, 177.880° W, 0–30 m; 25 October 2016; scuba; AWMM MA182117. 1 M (PCL: 15.8 mm); North of Napier Island, Stn. TAN1612/20, 29.230–29.231° S, 177.876–177.874° W, 0–30 m; 24 October 2016; scuba; NIWA 118168. 1 M (PCL: 10.0 mm); Raoul Island, Denham Bay, southern end, 29.288° S, 177.958° W, 3–18 m; 31 August 2017; scuba, col: Severine Hannam, Anna Murray, David Aguirre, Crispin Middleton, Carl Struthers, Matt Hill; AWMM MA168313. Northland. 1 M (PCL: 12.2 mm), 1 F (PCL: 14.0 mm); Bay of Islands, Tapuaetahi, 35.118° S, 173.978° E, 11 m; 4 September 1986; scuba, col: Brett Stephenson; AWMM MA6648. 1 F (poorly preserved); off northern tip of Poor Knights Islands, 35.417° S, 174.058° E, 40 m; scuba; col: Kelly Tarlton; AWMM MA8585. 1 M (PCL: 12.0); Poor Knights, Southern Island Harbour, 15–18 m; 25 May 1969; col: A.N. Baker; NMNZ CR.006006. 1 F (PCL: 14.7 mm); Poor Knights Islands, 24 m; May 1968; NMNZ CR.016812. 1 M (PCL 11.0 mm); North Island, Poor Knights Islands; 10 January 1970; col: Wade Doak; NMNZ CR.016808. 1 M (PCL: 15.3 mm); South Harbour, Poor Knights Islands, 18 m; 5 May 1969; col: B. Palmer; NMNZ CR.016811. 1 F (PCL: 13.6 mm); off Poor Knights, 18 m; 23 April 1967; col: B.B. Anderson; NMNZ CR.001779. Bay of Plenty. 1 M (PCL: 11.9 mm); Alderman Islands, underwater cave, several present on roof; Jan 2001; scuba; col: C.D. Roberts, J. Hope; NMNZ CR.016809.
Diagnosis. Large shrimp (total length 30–67 mm) with a slender, compressed body, densely covered with spines. Carapace and first three pleonites with curved forward-directed spines. Last three pleonites with straight posteriorly directed spines. Rostrum strong, ultimate point reaching to about middle of second article of antennular peduncle with dorsal and lateral spines but no ventral spines. Stylocerite short and pressed against basal antennular segment. Scaphocerite with outer margin entire for considerable distance before final tooth. Third maxilliped with ischium, merus and carpus provided with external row of spinules. Body white with red transverse bands on carapace, third and sixth pleonite, and third pereiopods; base of the third pereiopod deep blue, the remaining appendages are pale.
Measurements of examined specimens. CL: 13.5–21.3 mm (males), 17.5–20.0 mm (females), PCL: 9.8–15.3 mm (males), 12.6–14.7 mm (females), TL: 34.7–57.0 mm (males), 44.5–51.7 mm (females). Previously reported TL: 30–67 mm.
Distribution.Stenopus hispidus is the only pantropical species in the infraorder Stenopodidea. Previously recorded from the Western Atlantic from Cape Lookout, North Carolina, throughout the Caribbean and Gulf of Mexico to the southern border of Brazil [
65]; central Atlantic from Ascension Island [
66]; Eastern Pacific from Taboga Island, Panama [
61,
67]; and throughout the tropical and warm temperate Pacific; from the intertidal to 210 m [
57]. Specimens presented here are from the northern mainland New Zealand and the islands directly to the North (Lord Howe Island, Kermadec Islands); 3–40 m. Further specimens examined from the NMNZ collection were collected around the Solomon Islands, Niue and Tahiti, but are not presented in detail here.
Colour in life. Good descriptions of the color pattern for this species are previously given [
22,
57]. Considering the key to the species of
Stenopus provided by Saito et al. [
52], the diagnostic combination of coloration for this species are the broad red and white banding of the body and third pereiopod, not uniformly orange (as in
S. spinosus Risso, 1827 [
68]) and carapace not purple (as in
S. tenuirostris de Man, 1888 [
69]). The base of the third pereiopod deep blue, the remaining appendages are pale (the third maxilliped, fourth and fifth pereiopods are not blue as in
S. cyanoscelis Goy, 1984 [
70]) (see
Figure 26).
Remarks. All specimens examined here match the keys provided by Saito et al. [
52] well.
Stenopus hispidus remains the sole pantropical species in this genus, however, recent population genetic studies indicated regional genetic divergences among locations in the Indonesian Archipelago [
71] and between oceans [
72]. Further studies might reveal that this supposedly widespread species indeed represents a species complex but is for now considered as
S. hispidus sensu lato.
DNA sequence data. Three New Zealand specimens (NIWA 118168, AWMM MA182117 and MA168313, recently collected from the Kermadec Islands) unambiguously aligned with conspecific sequences deposited on GenBank with sequence divergences are 0.0% for both CO1 and 16S rRNA genes (
Figure 2,
Table 1). Reference sequences include specimens from across the Indo-Pacific [
17,
72,
73].































{kind=link}
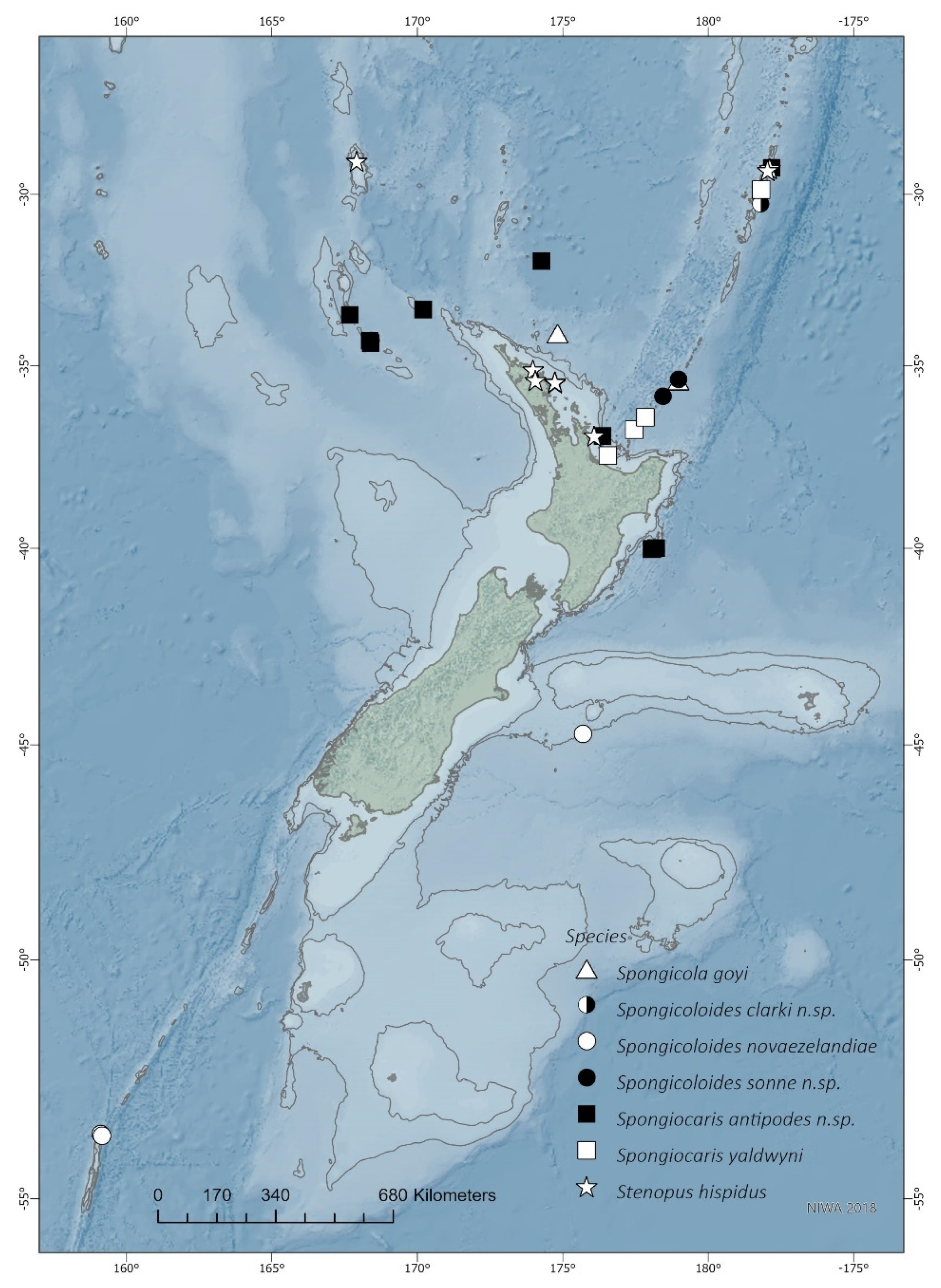{kind=link}
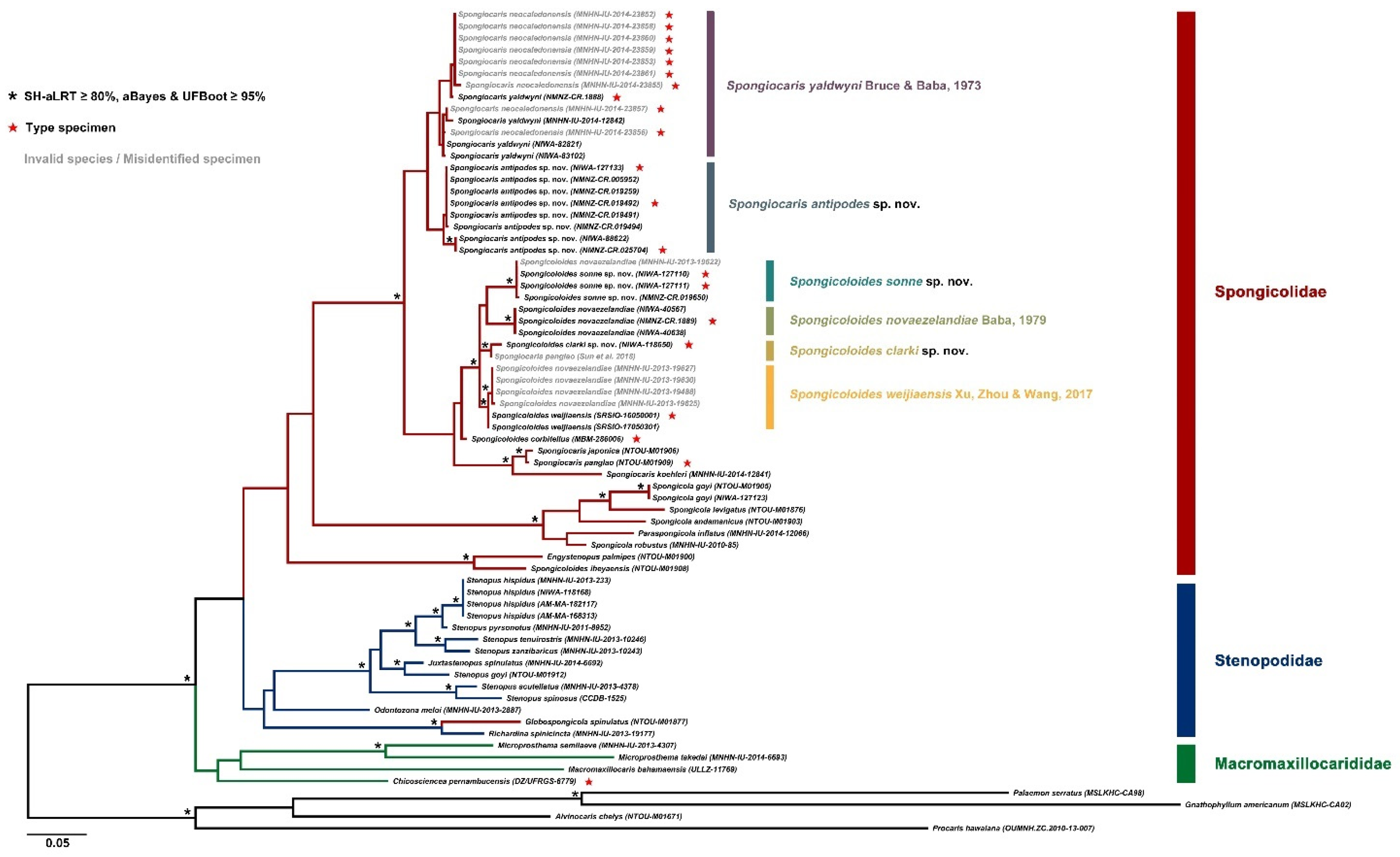{kind=link}
{kind=link}
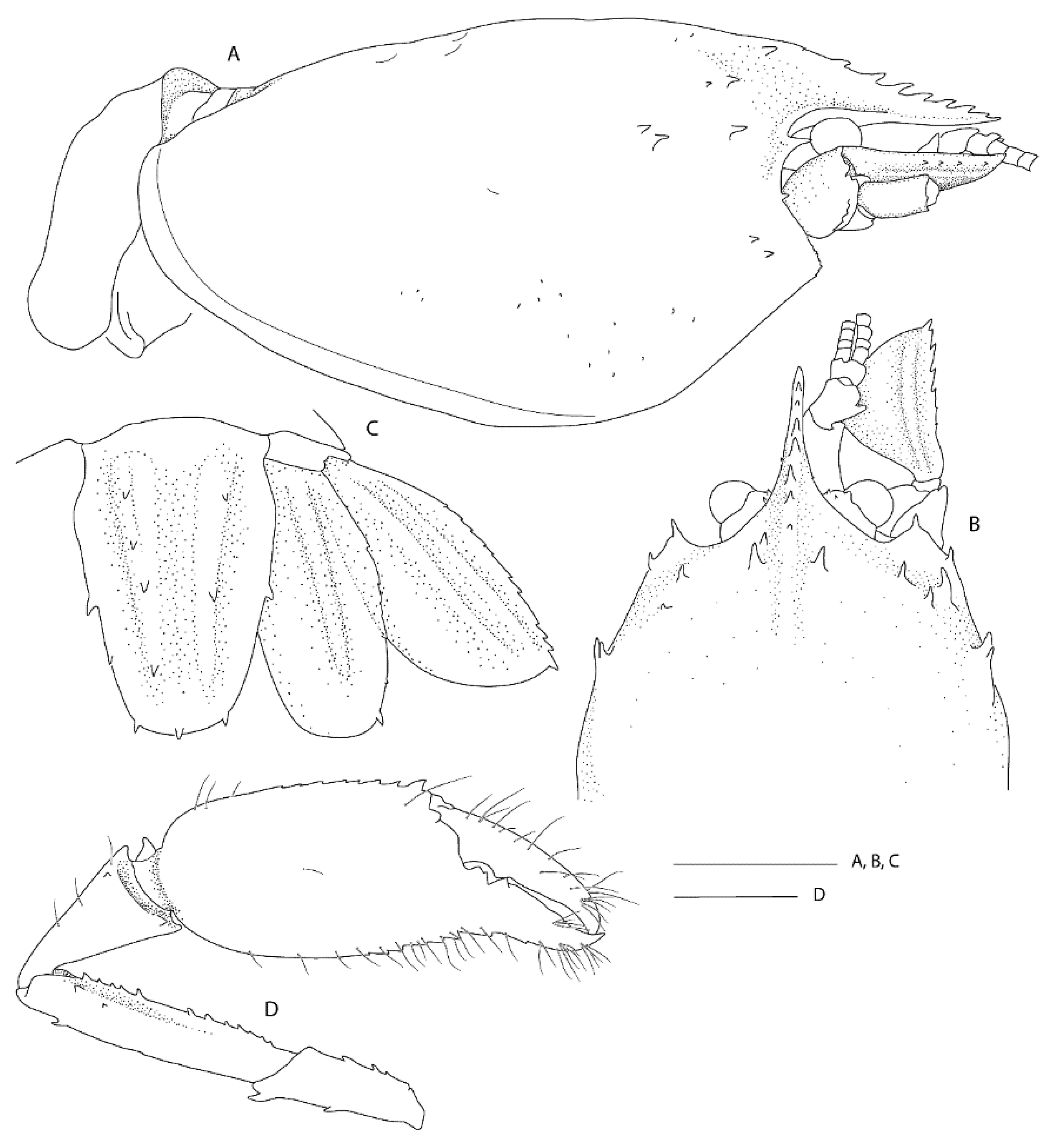{kind=link}
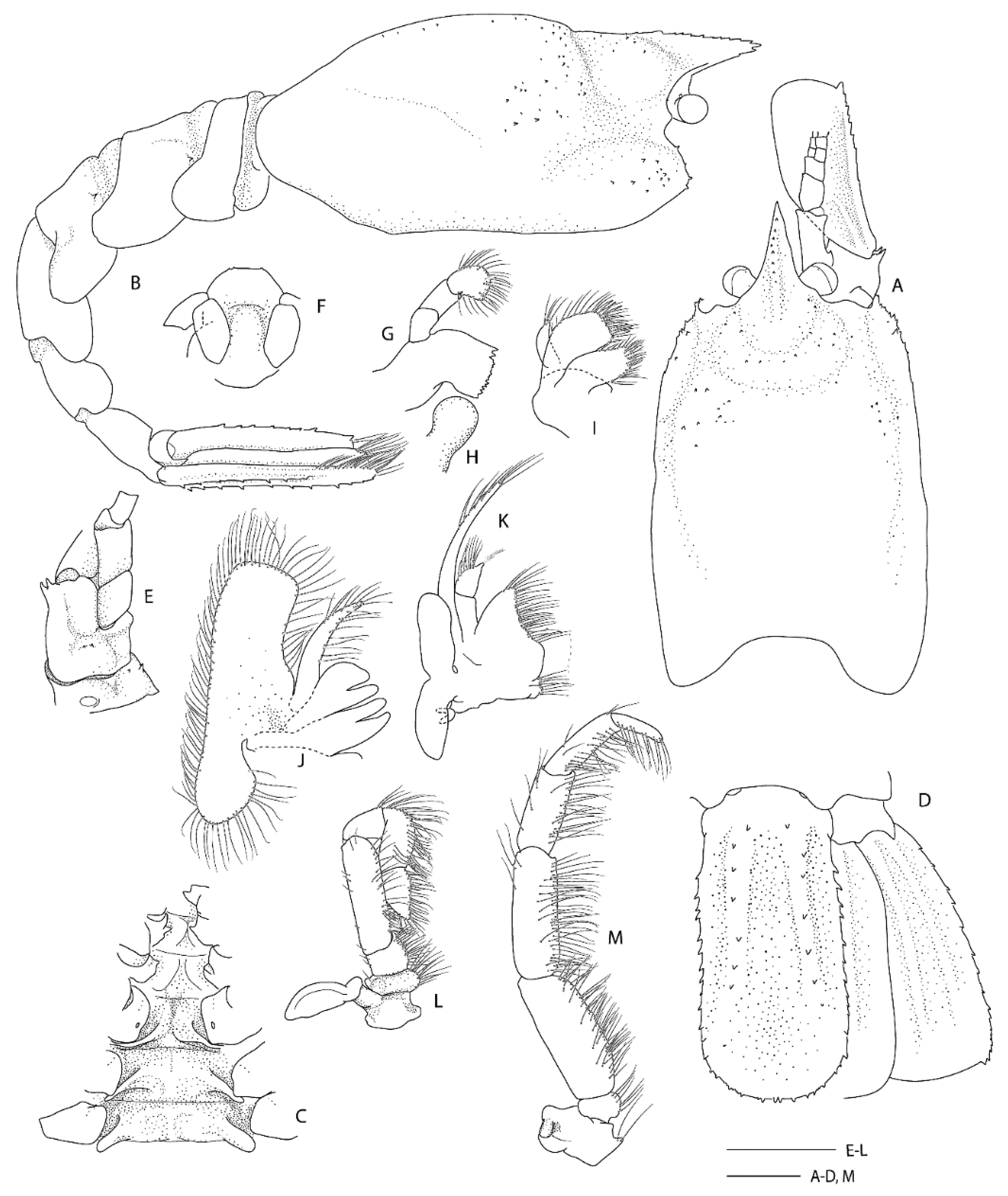{kind=link}
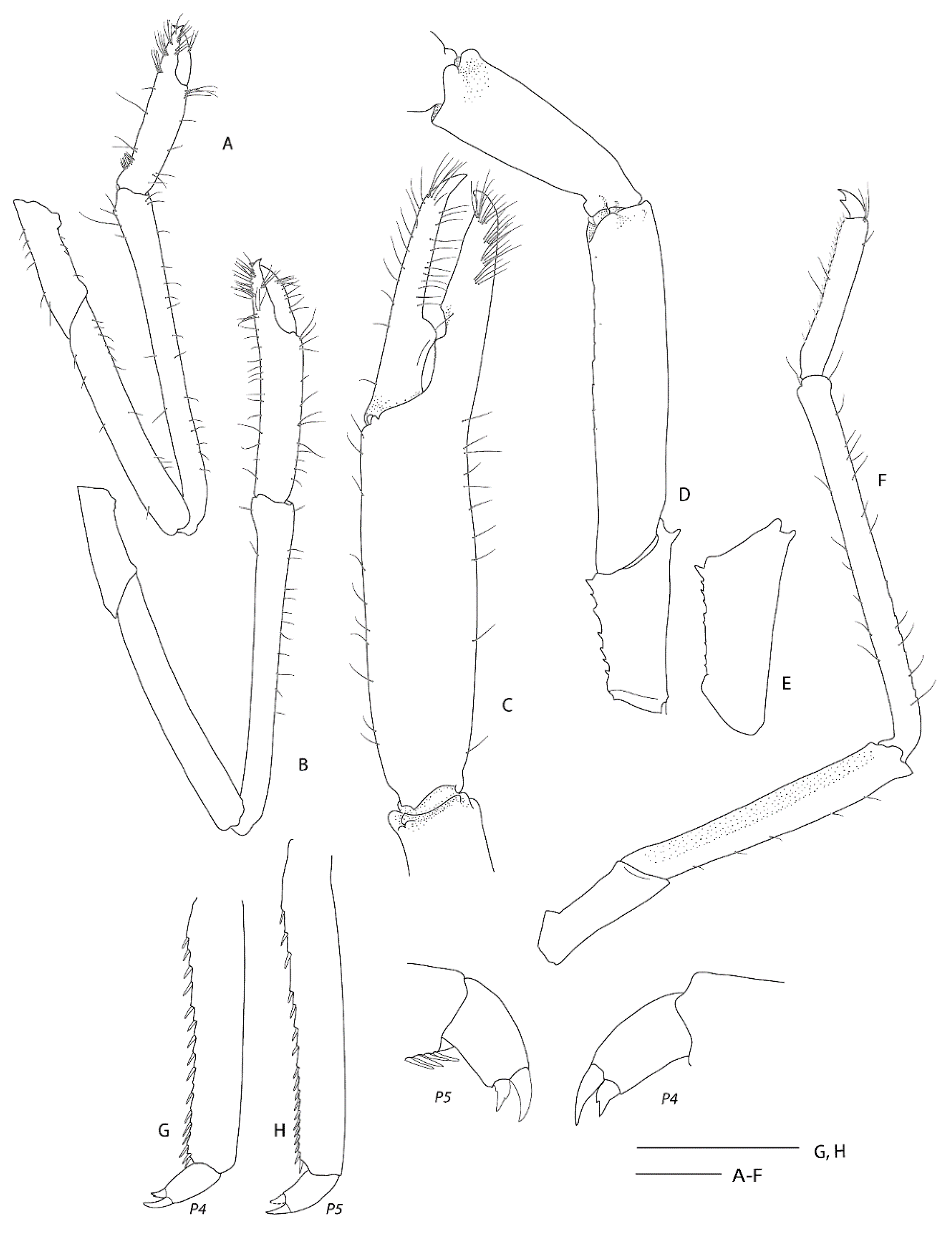{kind=link}
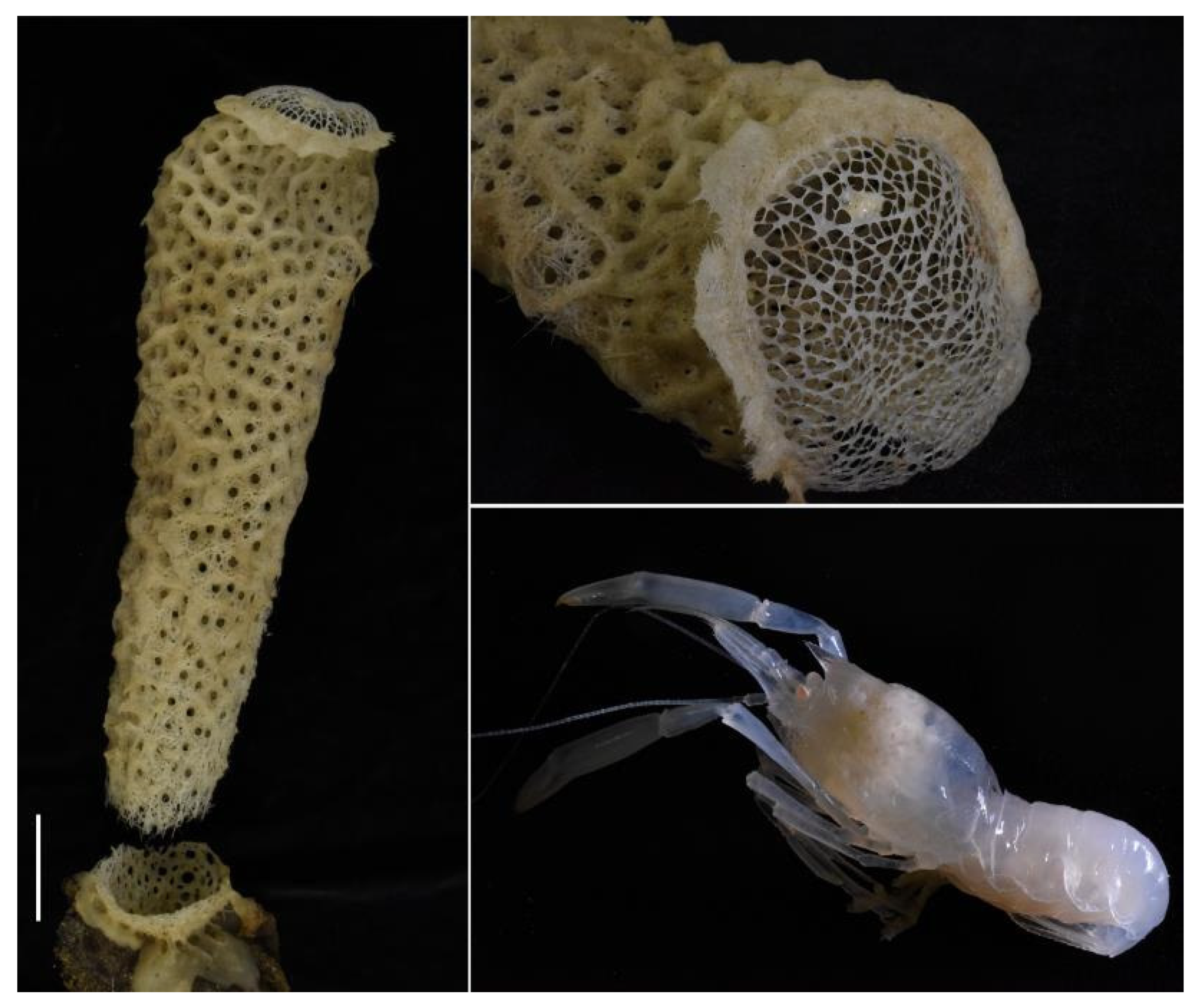{kind=link}
{kind=link}
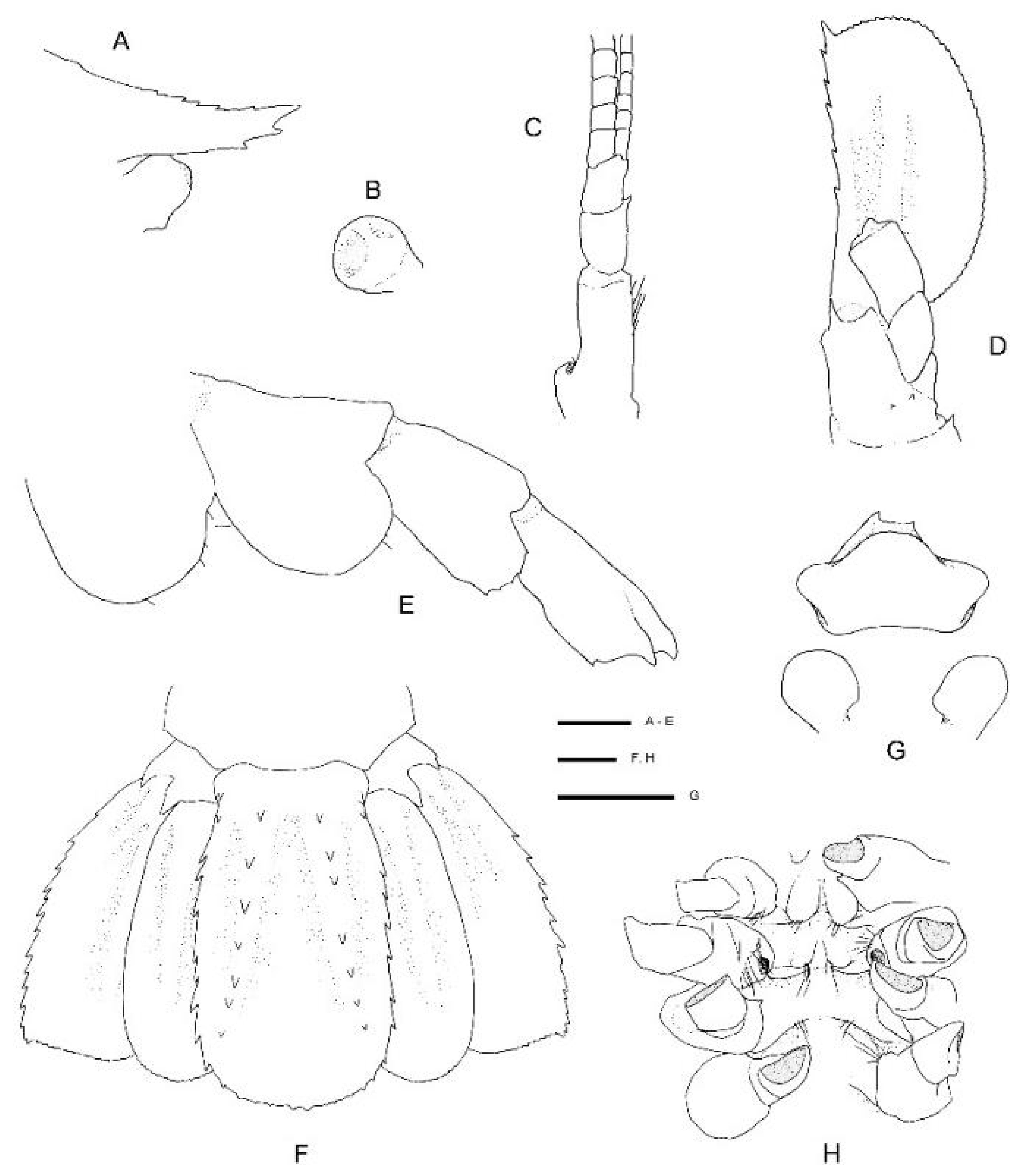{kind=link}
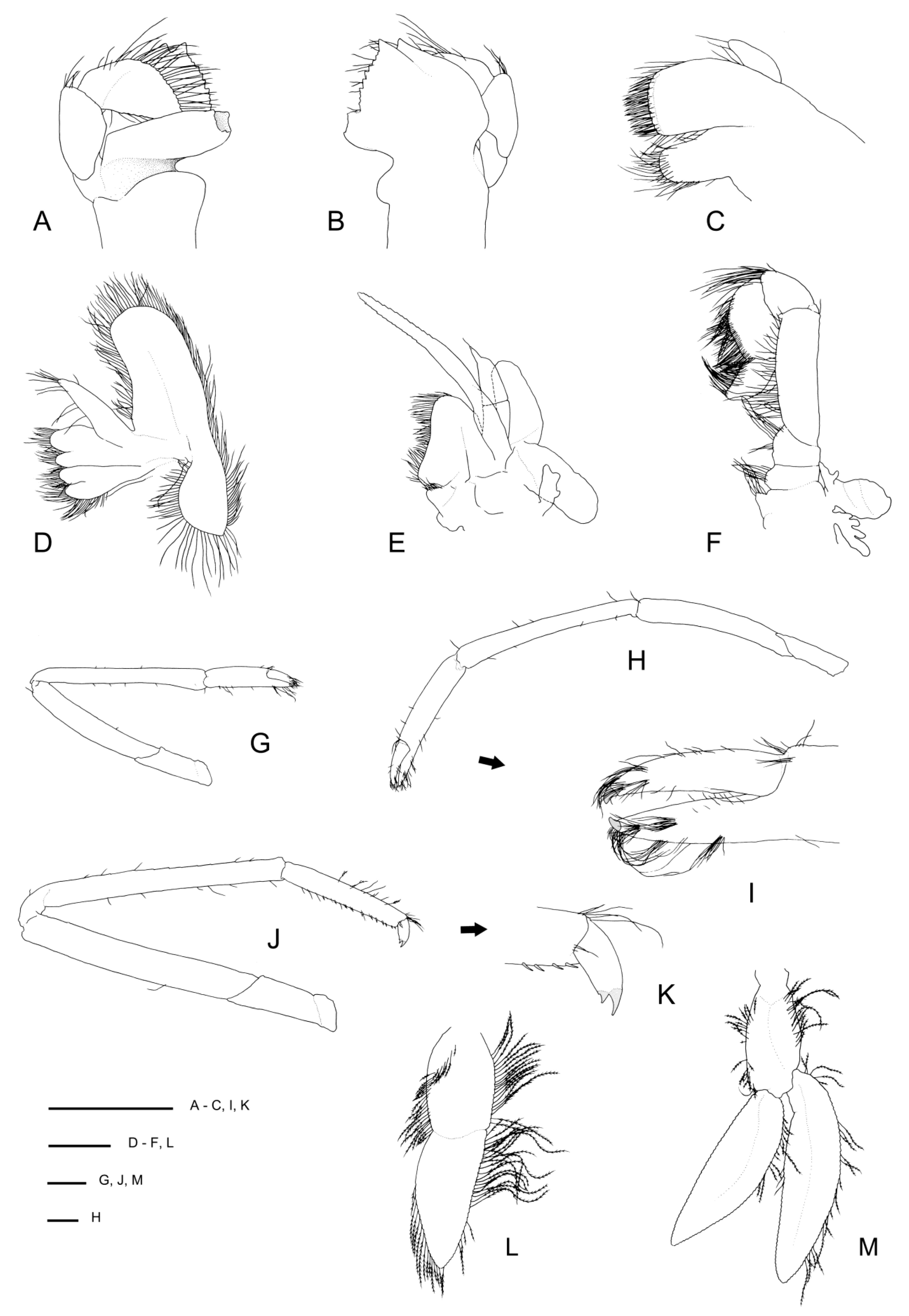{kind=link}
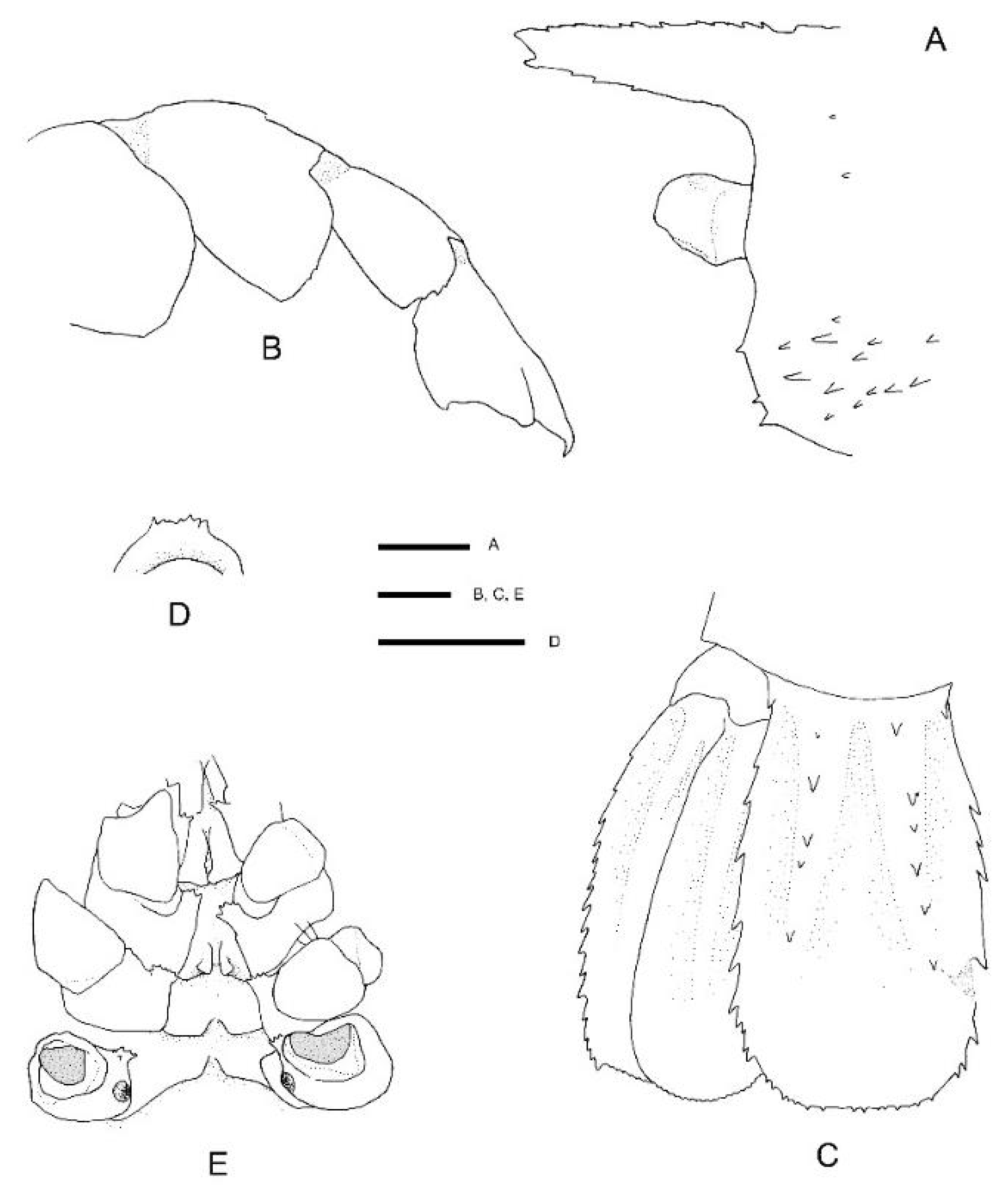{kind=link}
{kind=link}
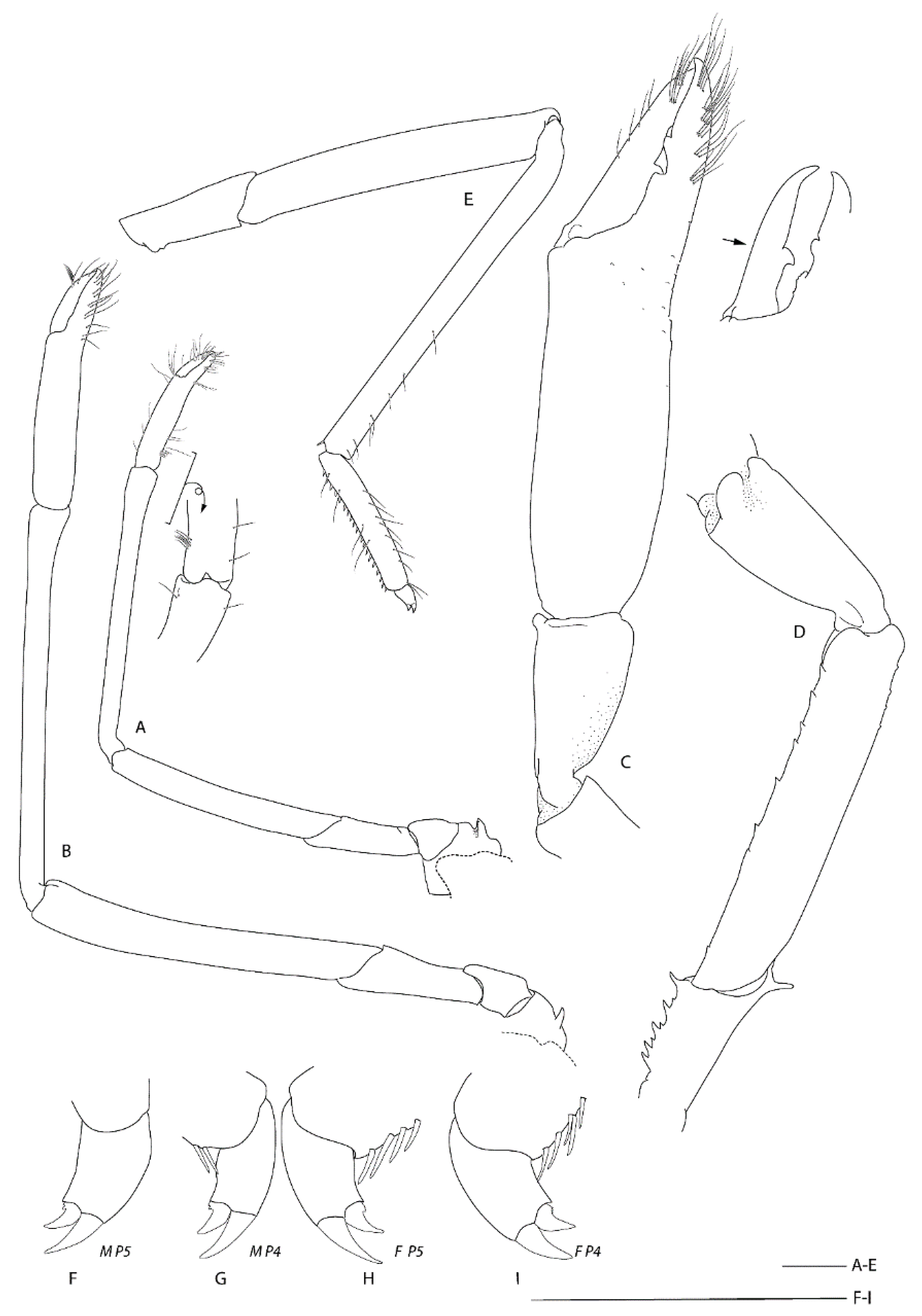{kind=link}
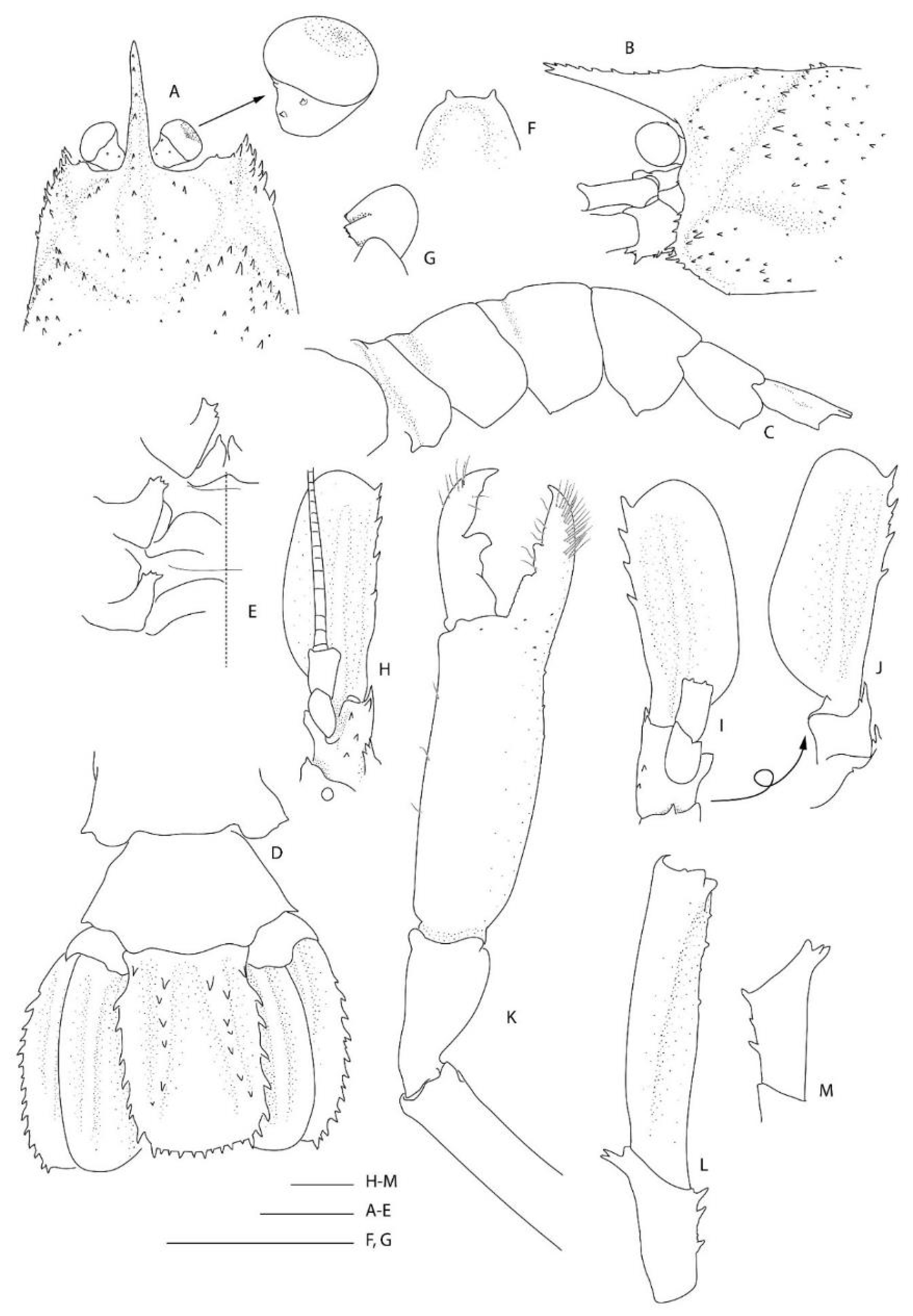{kind=link}
{kind=link}
{kind=link}
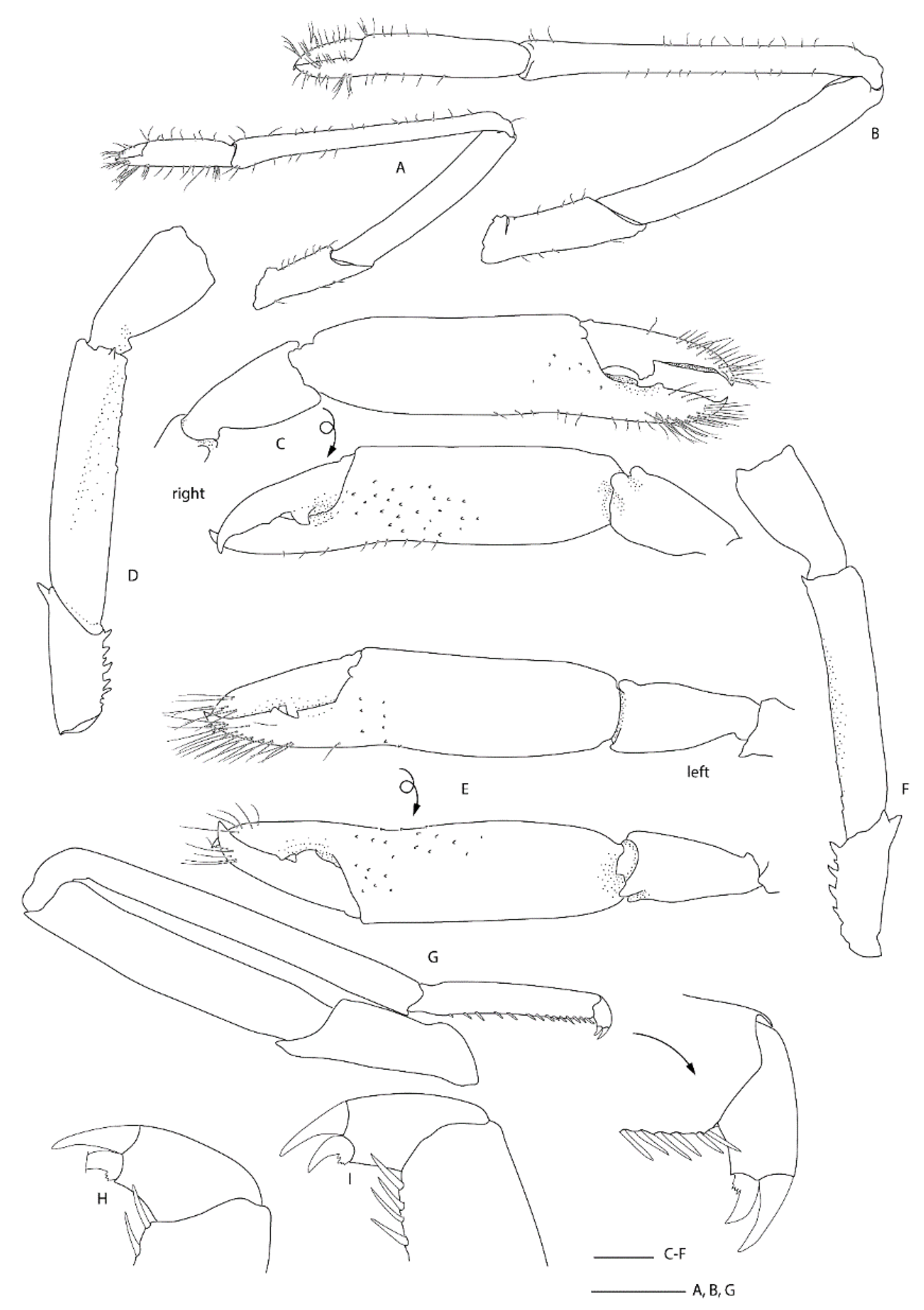{kind=link}
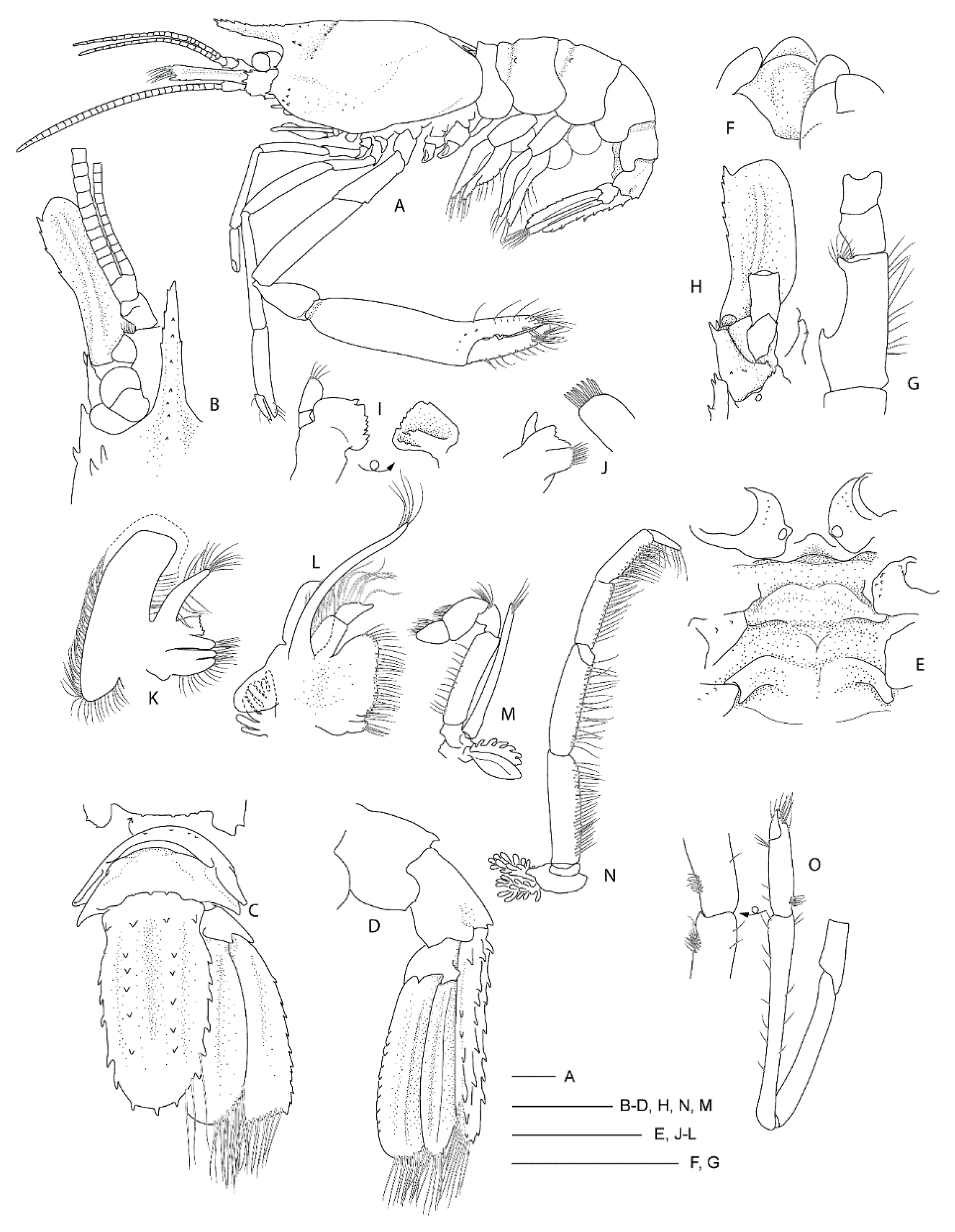{kind=link}
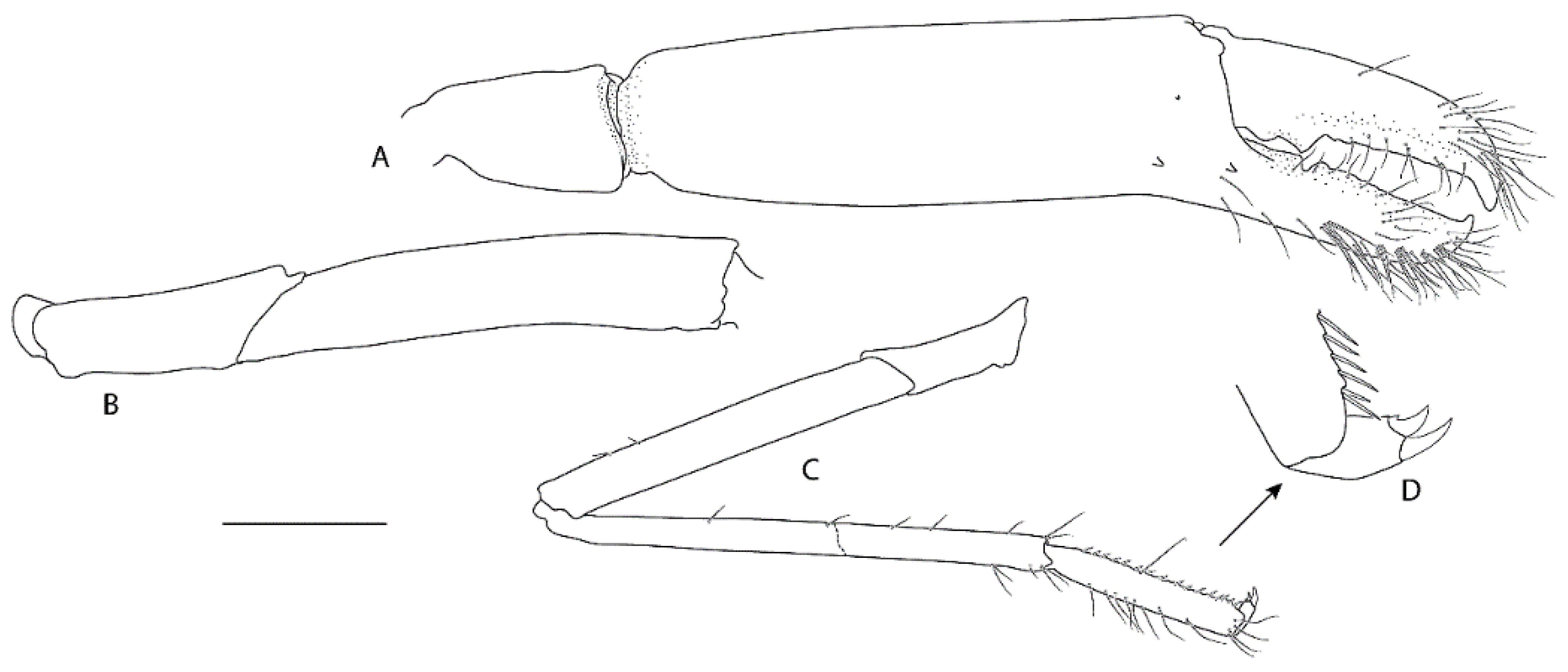{kind=link}
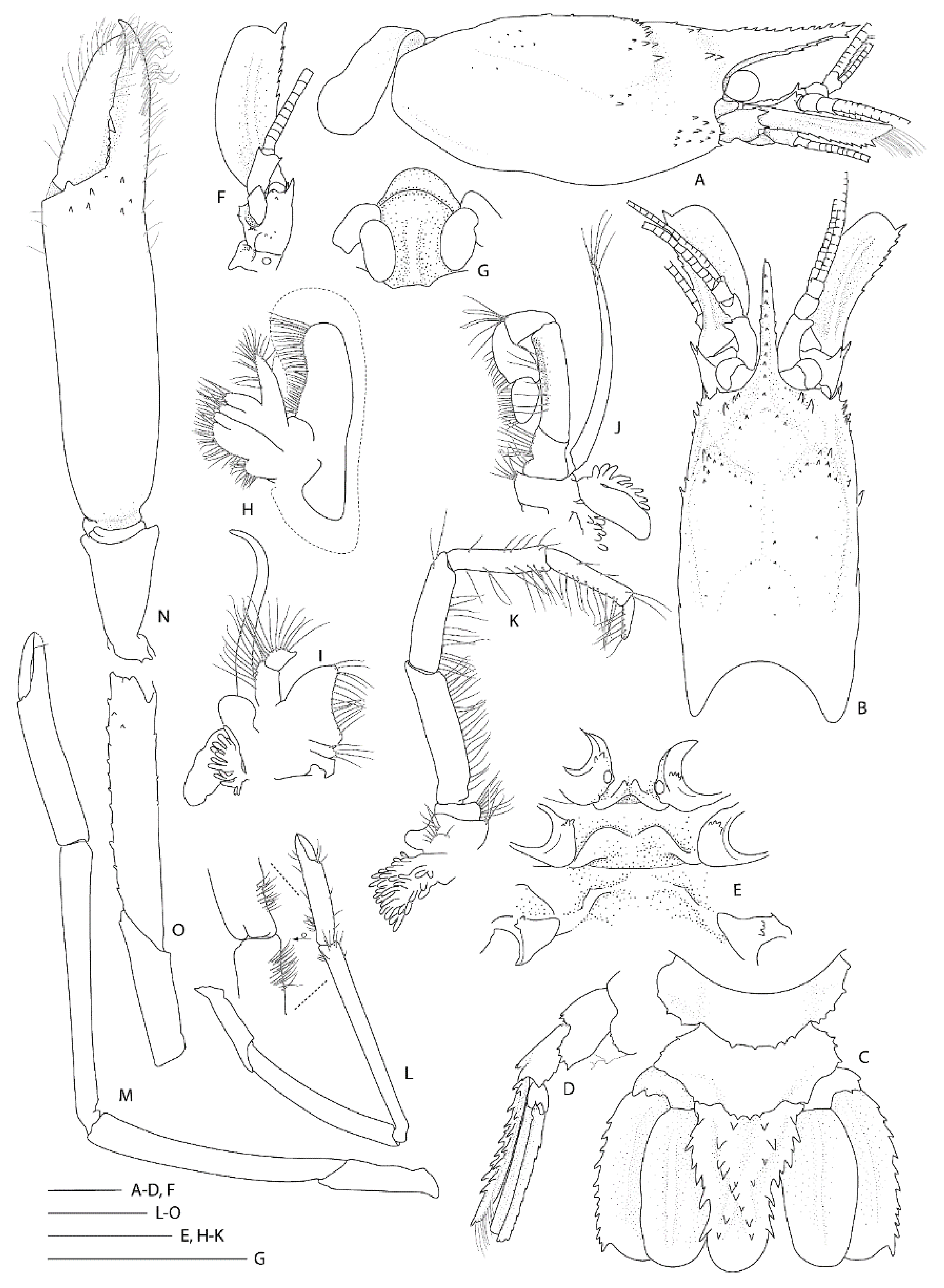{kind=link}
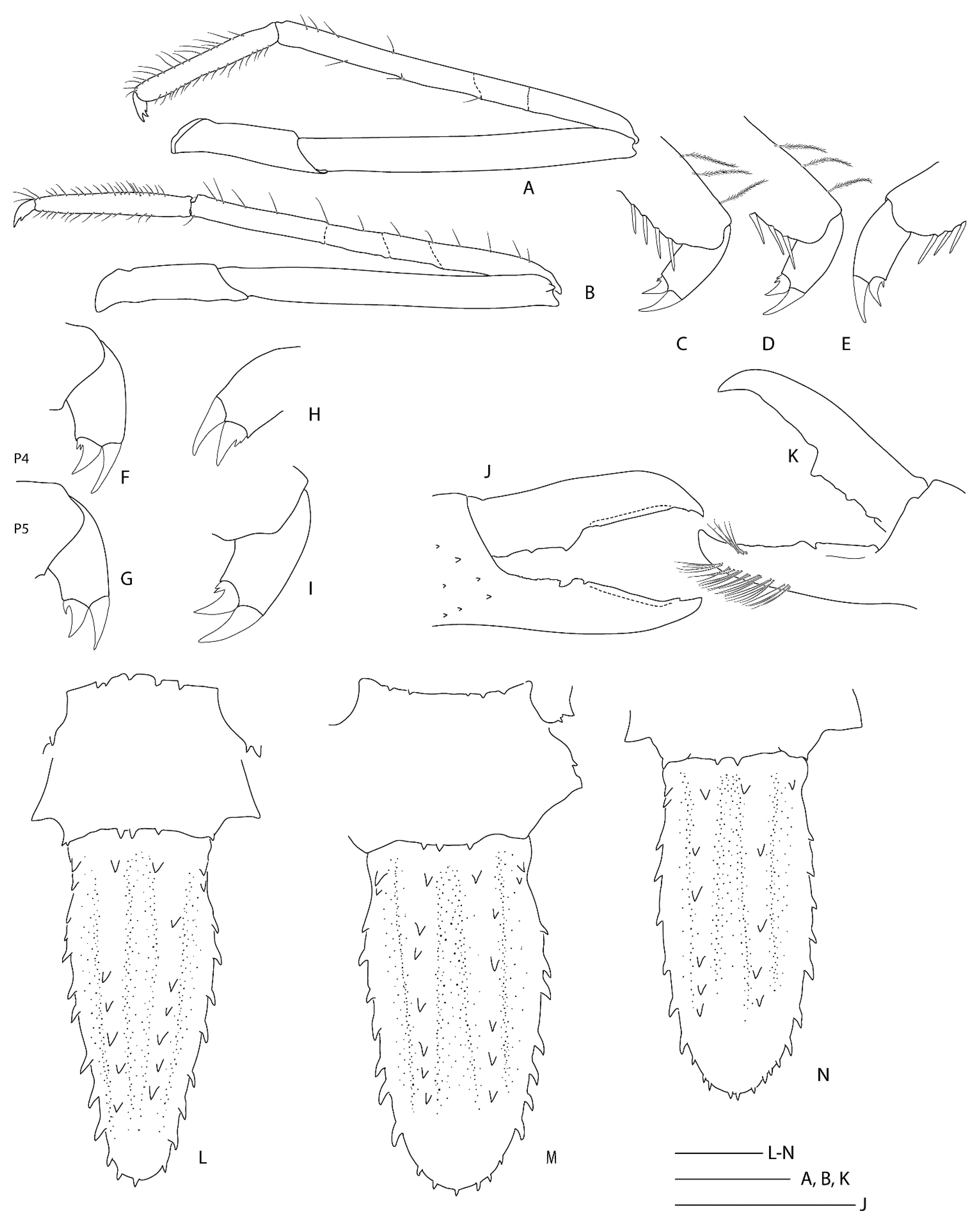{kind=link}
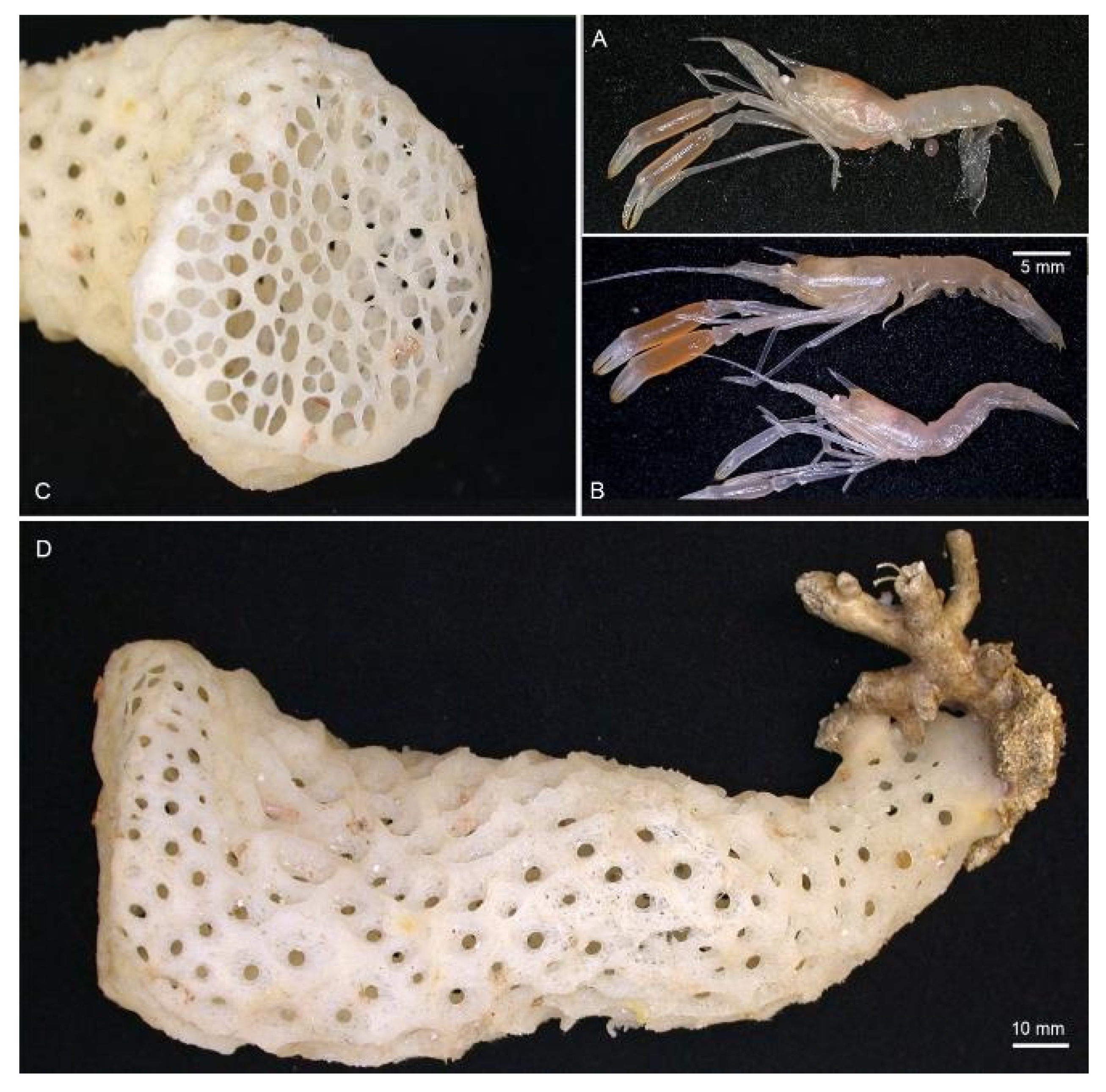{kind=link}
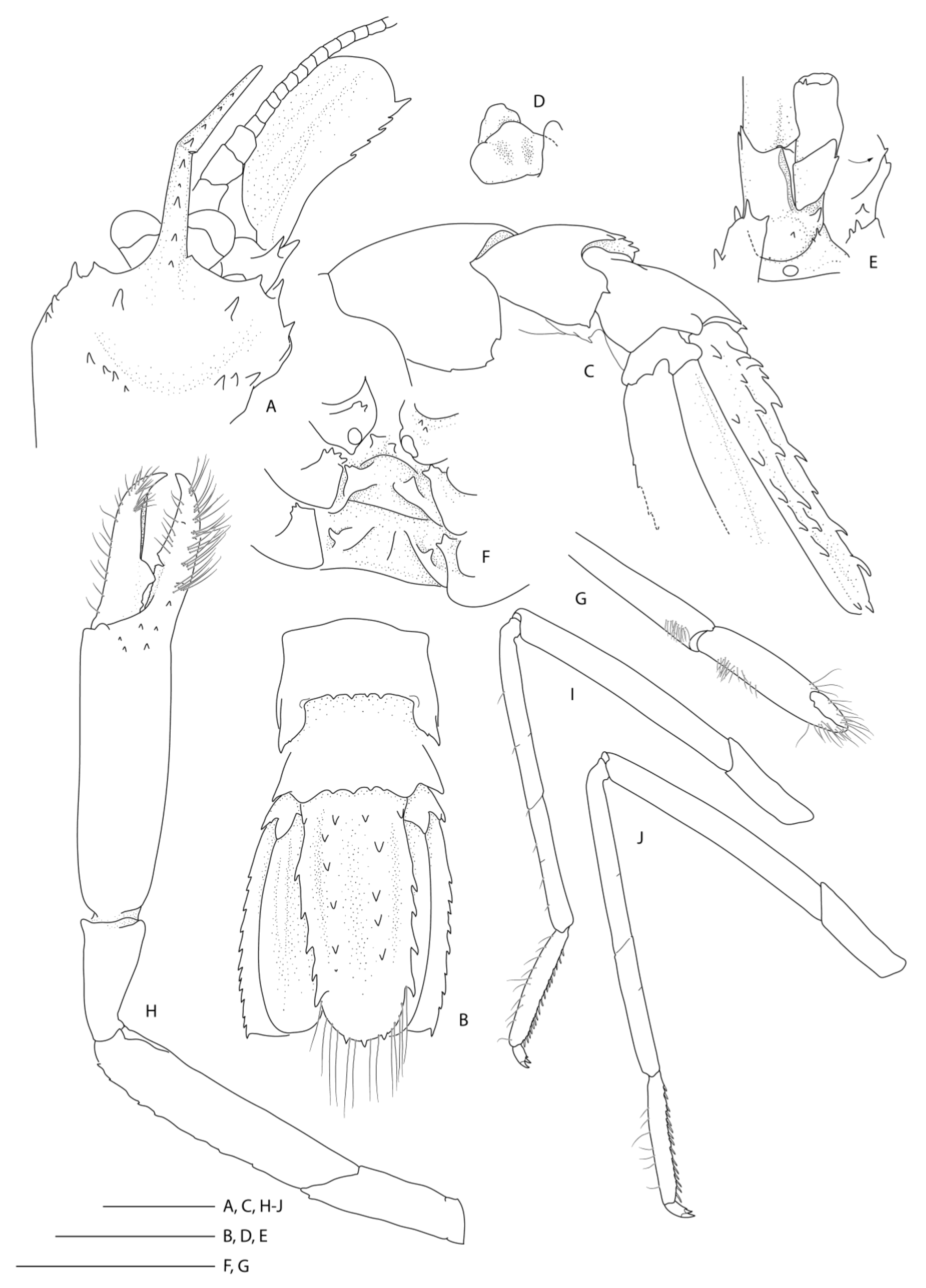{kind=link}
{kind=link}
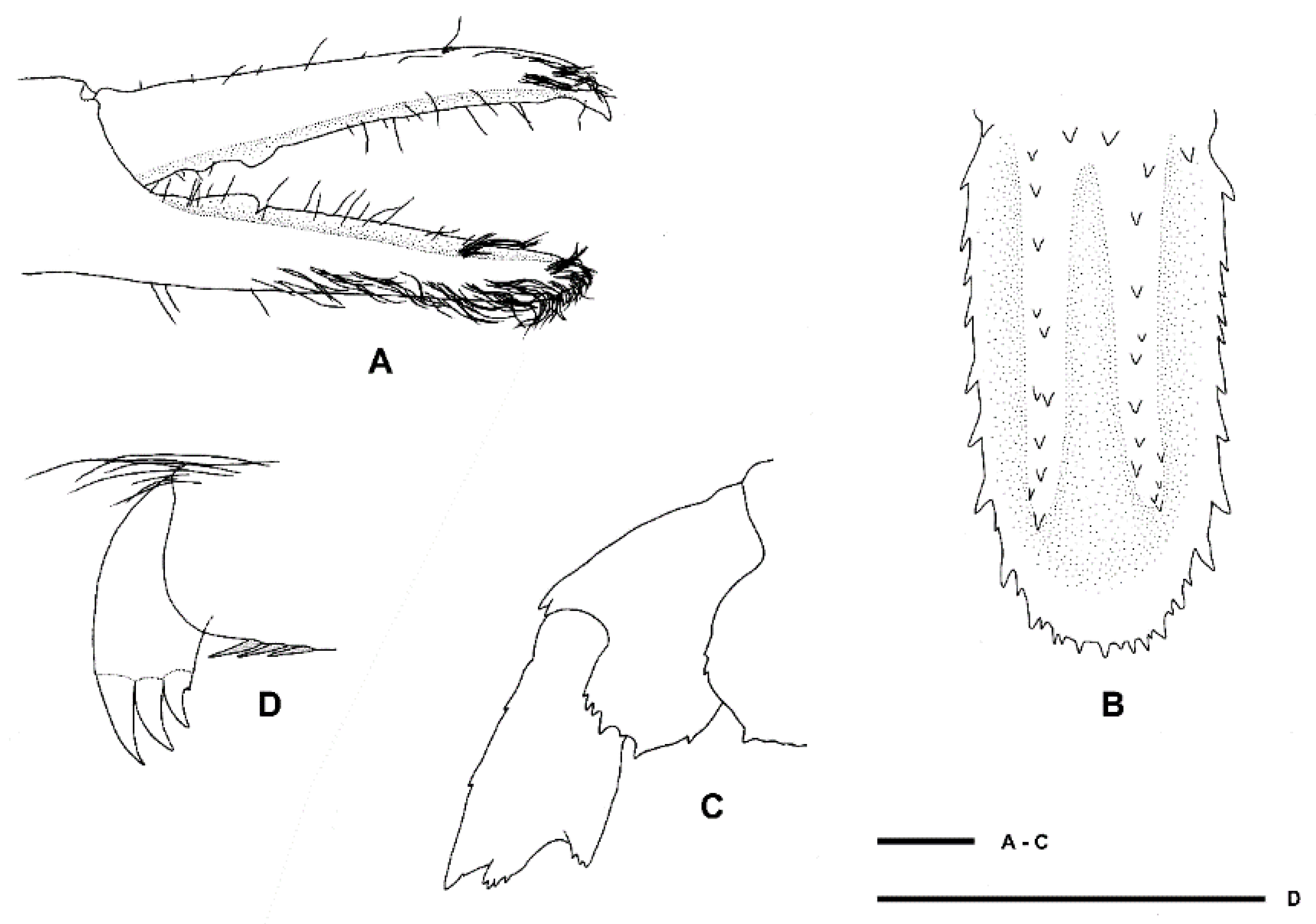{kind=link}
{kind=link}