
Integrative Descriptions of Two New Tardigrade Species along with the New Record of Mesobiotus skorackii Kaczmarek et al., 2018 from Canada


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Microscopy and Imaging
2.3. Morphometrics and Morphological Nomenclature
2.4. Comparative Material
2.5. Genotyping
2.6. Comparative Genetic Analysis
3. Results
Taxonomic Account
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramazzotti, G.; Maucci, W. Il Phylum Tardigrada. III edizione riveduta e aggiornata. Mem. Ist. Ital. Idrobiol. Pallanza 1983, 41, 1–1012. [Google Scholar]
- Beasley, C.W. The Phylum Tardigrada, 3rd ed.; English Translation; Ramazzotti, G., Maucci, W., Eds.; Clark Beasley: Abilene, TX, USA, 1995; pp. 1–1014. [Google Scholar]
- Nelson, D.R.; Guidetti, R.; Rebecchi, L.; Kaczmarek, Ł.; McInnes, S. Phylum Tardigrada. In Thorp and Covich’s Freshwater Invertebrates; Elsevier: Amsterdam, The Netherlands, 2020; pp. 505–522. [Google Scholar]
- Guidetti, R.; Bertolani, R.B. Tardigrade Taxonomy: An updated check list of the taxa and a list of characters for their identification. Zootaxa 2005, 845, 1–46. [Google Scholar] [CrossRef]
- Degma, P.; Guidetti, R. Notes to the current checklist of Tardigrada. Zootaxa 2007, 1579, 41–53. [Google Scholar] [CrossRef]
- Vicente, F.; Bertolani, R. Considerations on the taxonomy of the Phylum Tardigrada. Zootaxa 2013, 3626, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Degma, P.; Bertolani, R.; Guidetti, R. Actual Checklist of Tardigrada Species. Available online: https://iris.unimore.it/retrieve/handle/11380/1178608/357802/Actual%20checklist%20of%20Tardigrada%2040th%20Edition%2015-07-21.pdf (accessed on 15 July 2021).
- Thulin, G. Über die phylogenie und das system der tardigraden. Hereditas 1928, 11, 207–266. [Google Scholar] [CrossRef]
- Kristensen, R.M.; Michalczyk, Ł.; Kaczmarek, Ł. The first record of the genus Bryodelphax (Tardigrada: Heterotardigrada: Echiniscidae) from Easter Island, Rapa Nui (Pacific Ocean, Chile) with the description of a new species, Bryodelphax aaseae. Zootaxa 2010, 2343, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Gąsiorek, P.; Suzuki, A.C.; Kristensen, R.M.; Lachowska-Cierlik, D.; Michalczyk, Ł. Untangling the Echiniscus Gordian knot: Stellariscus gen. nov. (Heterotardigrada: Echiniscidae) from Far East Asia. Invert. Syst. 2018, 32, 1234–1247. [Google Scholar] [CrossRef]
- Schultze, C.A.S. Macrobiotus Hufelandii, animal e crustaceorum classe novum, reviviscendi post diuturnam asphyxiam et ariditatem potens; Berolini Apud Carolum Curths: Berlin, Germany, 1834; pp. 1–8. [Google Scholar]
- Stec, D.; Dudziak, M.; Michalczyk, Ł. Integrative descriptions of two new Macrobiotidae species (Tardigrada: Eutardigrada: Macrobiotoidea) from French Guiana and Malaysian Borneo. Zool. Stud. 2020, 59, e23. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.; Goudie, I. The tardigrade and associated micrometazoa of the textured lungwort lichen, Lobaria scrobiculata, in Eastern Newfoundland, Canada. J. Agric. Life Sci. 2020, 7. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Michalczyk, Ł.; Mcinnes, S.J. Annotated zoogeography of non-marine Tardigrada. Part III: North America and Greenland. Zootaxa 2016, 4203, 1–249. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Zawierucha, K.; Buda, J.; Stec, D.; Gawlak, M.; Michalczyk, Ł.; Roszkowska, M. An integrative redescription of the nominal taxon for the Mesobiotus harmsworthi group (Tardigrada: Macrobiotidae) leads to descriptions of two new Mesobiotus species from Arctic. PLoS ONE 2018, 13, e0204756. [Google Scholar] [CrossRef] [PubMed]
- Stec, D.; Smolak, R.; Kaczmarek, Ł.; Michalczyk, Ł. An integrative description of Macrobiotus paulinae sp. nov. (Tardigrada: Eutardigrada: Macrobiotidae: Hufelandi Group) from Kenya. Zootaxa 2015, 4052, 501–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roszkowska, M.; Stec, D.; Gawlak, M.; Kaczmarek, Ł. An integrative description of a new tardigrade species Mesobiotus romani sp. nov. (Macrobiotidae: Harmsworthi group) from the Ecuadorian Pacific coast. Zootaxa 2018, 4450, 550–564. [Google Scholar] [CrossRef] [PubMed]
- Dastych, H. A new species of the genus Mopsechiniscus Du Bois-Reymond Marcus, 1944 (Tardigrada) from the Venezuelan Andes. Acta Biol. Ben. 1999, 10, 91–101. [Google Scholar]
- Fontoura, P.; Morais, P. Assessment of traditional and geometric morphometrics for discriminating cryptic species of the Pseudechiniscus suillus complex (Tardigrada, Echiniscidae). J. Zool. Syst. Evol. Res. 2011, 49, 26–33. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Zawierucha, K.; Smykla, J.; Michalczyk, Ł. Tardigrada of the Revdalen (Spitsbergen) with the descriptions of two new species: Bryodelphax parvuspolaris (Heterotardigrada) and Isohypsibius coulsoni (Eutardigrada). Polar Biol. 2012, 35, 1013–1026. [Google Scholar] [CrossRef] [Green Version]
- Pilato, G.; Binda, M.G. Definition of families, subfamilies, genera and subgenera of the Eutardigrada, and keys to their identification. Zootaxa 2010, 2404, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, Ł.; Michalczyk, Ł. The Macrobiotus hufelandi group (Tardigrada) revisited. Zootaxa 2017, 4363, 101–123. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Bartylak, T.; Stec, D.; Kulpa, A.; Kepel, M.; Kepel, A.; Roszkowska, M. Revisiting the genus Mesobiotus Vecchi et al., 2016 (Eutardigrada, Macrobiotidae)—Remarks, updated dichotomous key and an integrative description of new species from Madagascar. Zool. Anz. 2020, 287, 121–146. [Google Scholar] [CrossRef]
- Michalczyk, Ł.; Kaczmarek, Ł. A description of the new tardigrade Macrobiotus reinhardti (Eutardigrada: Macrobiotidae, harmsworthi group) with some remarks on the oral cavity armature within the genus Macrobiotus Schultze. Zootaxa 2003, 331, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, Ł.; Cytan, J.; Zawierucha, K.; Diduszko, D.; Michalczyk, Ł. Tardigrades from Peru (South America), with descriptions of three new species of Parachela. Zootaxa 2014, 3790, 357–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilato, G. Analisi di nuovi caratteri nello studio degli eutardigradi. Animalia 1981, 8, 51–57. [Google Scholar]
- Kiosya, Y.; Pogwizd, J.; Matsko, Y.; Vecchi, M.; Stec, D. Phylogenetic position of two Macrobiotus species with a revisional note on Macrobiotus sottilei Pilato, Kiosya, Lisi & Sabella, 2012 (Tardigrada: Eutardigrada: Macrobiotidae). Zootaxa 2021, 4933, 113–135. [Google Scholar] [CrossRef]
- Perry, E.; Miller, W.R.; Kaczmarek, Ł. Recommended abbreviations for the names of genera of the phylum Tardigrada. Zootaxa 2019, 4608, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Michalczyk, Ł.; Kaczmarek, Ł. The Tardigrada Register: A comprehensive online data repository for tardigrade taxonomy. J. Limnol. 2013, 72, 175–181. [Google Scholar] [CrossRef]
- Gąsiorek, P.; Vončina, K.; Degma, P.; Michalczyk, Ł. Small is beautiful: The first phylogenetic analysis of Bryodelphax Thulin, 1928 (Heterotardigrada, Echiniscidae). Zoosystematics Evol. 2020, 96, 217–236. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Michalczyk, Ł. A new species Bryodelphax asiaticus (Tardigrada: Heterotardigrada: Echiniscidae) from Mongolia (Central Asia). Raffles Bull. Zool. 2004, 599–602. [Google Scholar]
- Kaczmarek, Ł.; Michalczyk, Ł.; Degma, P. A new species of Tardigrada Bryodelphax brevidentatus sp. nov. (Heterotardigrada: Echiniscidae) from China (Asia). Zootaxa 2005, 1080, 33–38. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Parnikoza, I.; Gawlak, M.; Esefeld, J.; Peter, H.-U.; Kozeretska, I.; Roszkowska, M. Tardigrades from Larus dominicanus Lichtenstein, 1823 nests on the Argentine Islands (Maritime Antarctic). Polar Biol. 2018, 41, 283–301. [Google Scholar] [CrossRef] [Green Version]
- Roszkowska, M.; Gawlak, M.; Draga, M.; Kaczmarek, Ł. Two new species of Tardigrada from Ecuador (South America). Zootaxa 2019, 4545, 511–530. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Michalczyk, Ł. Two new species of Macrobiotidae, Macrobiotus szeptyckii (harmsworthi group) and Macrobiotus kazmierskii (hufelandi group) from Argentina. Acta Zool. Cracov.Ser. B Invert. 2009, 52, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, Ł.; Michalczyk, Ł. New records of Tardigrada from Cyprus with a description of the new species Macrobiotus marlenae (Eutardigrada: Macrobiotidae, hufelandi group). Genus 2004, 15, 141–152. [Google Scholar]
- Pilato, G.; Kaczmarek, Ł.; Michalczyk, Ł.; Lisi, O. Macrobiotus polonicus, a new species of Tardigrada from Poland (Eutardigrada: Macrobiotidae, ‘hufelandi group’). Zootaxa 2003, 258, 1–8. [Google Scholar] [CrossRef]
- Roszkowska, M.; Ostrowska, M.; Stec, D.; Janko, K.; Kaczmarek, Ł. Macrobiotus polypiformis sp. nov., a new tardigrade (Macrobiotidae; hufelandi group) from the Ecuadorian Pacific coast, with remarks on the claw abnormalities in eutardigrades. Eur. J. Taxon. 2017, 327. [Google Scholar] [CrossRef] [Green Version]
- Kuzdrowska, K.A.; Mioduchowska, M.; Gawlak, M.; Bartylak, T.; Kepel, A.; Kepel, M.; Kaczmarek, Ł. Integrative description of Macrobiotus porifini sp. nov. (Macrobiotidae) from Madagascar and its phylogenetic position within the hufelandi group. Eur. Zool. J 2021, 88, 375–389. [Google Scholar] [CrossRef]
- Pilato, G.; Kiosya, Y.; Lisi, O.; Sabella, G. New records of Eutardigrada from Belarus with the description of three new species. Zootaxa 2012, 3179, 39–60. [Google Scholar] [CrossRef]
- Kayastha, P.; Berdi, D.; Mioduchowska, M.; Gawlak, M.; Łukasiewicz, A.; Gołdyn, B.; Kaczmarek, Ł. Some tardigrades from Nepal (Asia) with integrative description of Macrobiotus wandae sp. nov. (Macrobiotidae: Hufelandi Group). Ann. Zool. 2020, 70, 121–142. [Google Scholar] [CrossRef]
- Casquet, J.; Thebaud, C.; Gillespie, R.G. Chelex without boiling, a rapid and easy technique to obtain stable amplifiable DNA from small amounts of ethanol-stored spiders. Mol. Ecol. Resour. 2012, 12, 136–141. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Grobys, D.; Kulpa, A.; Bartylak, T.; Kmita, H.; Kepel, M.; Kepel, A.; Roszkowska, M. Two new species of the genus Milnesium Doyère, 1840 (Tardigrada, Apochela, Milnesiidae) from Madagascar. ZooKeys 2019, 884, 1–22. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Roszkowska, M.; Poprawa, I.; Janelt, K.; Kmita, H.; Gawlak, M.; Fiałkowska, E.; Mioduchowska, M. Integrative description of bisexual Paramacrobiotus experimentalis sp. nov. (Macrobiotidae) from Republic of Madagascar (Africa) with microbiome analysis. Mol. Phyl. Evol. 2020, 145, 106730. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 4, 294–299. [Google Scholar]
- Sands, C.J.; McInnes, S.J.; Marley, N.J.; Goodall-Copestake, W.P.; Convey, P.; Linse, K. Phylum Tardigrada: An “individual” approach. Cladistics 2008, 24, 861–871. [Google Scholar] [CrossRef]
- Mironov, S.V.; Dabert, J.; Dabert, M. A new feather mite species of the genus Proctophyllodes Robin, 1877 (Astigmata: Proctophyllodidae) from the Long-tailed Tit Aegithalos caudatus (Passeriformes: Aegithalidae)—Morphological description with DNA barcode data. Zootaxa 2012, 3253, 54–61. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. ISBN 9780123721808. [Google Scholar]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European molecular biology open software suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Goujon, M.; McWilliam, H.; Li, W.; Valentin, F.; Squizzato, S.; Paern, J.; Lopez, R. A new bioinformatics analysis tools framework at EMBL–EBI. Nucleic Acids Res. 2010, 38, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Doyère, M. Mémoire sur les tardigrades. Ann. Sci. Nat. Zool. 1840, 2, 269–362. [Google Scholar]
- Marcus, E. Zur Ökologie und Physiologie der Tardigraden. Zool. Jahrb. Abt. Syst. 1927, 44, 323–370. [Google Scholar]
- Richters, F. Tardigrada. In Handbuch der Zoologie, 3rd ed.; Kükenthal, W., Krumbach, T., Eds.; Walter de Gruyter & Co.: Berlin, Germany; Leipzig, Germany, 1926; Volume 3, pp. 58–61. [Google Scholar]
- Durante Pasa, M.V.; Maucci, W. Tardigradi muscicoli dell’Istria con descrizione di due specie nuove. Mem. Ist. Ital. Idrobiol. Pallanza 1975, 32, 69–91. [Google Scholar]
- McInnes, S.J. Zoogeographic distribution of terrestrial/freshwater tardigrades from current literature. J. Nat. Hist. 1994, 28, 257–352. [Google Scholar] [CrossRef]
- Gąsiorek, P.; Stec, D.; Morek, W.; Marnissi, J.; Michalczyk, Ł. The tardigrade fauna of Tunisia, with an integrative description of Bryodelphax maculatus sp. nov. (Heterotardigrada: Echiniscidae). Afr. Zool. 2017, 52, 77–89. [Google Scholar] [CrossRef]
- Jørgensen, A.; Møbjerg, N.; Kristensen, R.M. Phylogeny and evolution of the Echiniscidae (Echiniscoidea, Tardigrada)–an investigation of the congruence between molecules and morphology. J. Zool. Syst. Evol. Res. 2011, 49, 6–16. [Google Scholar] [CrossRef]
- Vicente, F.; Fontoura, P.; Cesari, M.; Rebecchi, L.; Guidetti, R.; Serrano, A.; Bertolani, R. Integrative taxonomy allows the identification of synonymous species and the erection of a new genus of Echiniscidae (Tardigrada, Heterotardigrada). Zootaxa 2013, 3613, 557–572. [Google Scholar] [CrossRef] [Green Version]
- Węglarska, B. Die Tardigraden (Tardigrada) Spitzbergens. Acta Zool. Cracov. 1965, 11, 43–52. [Google Scholar]
- Gąsiorek, P.; Stec, D.; Zawierucha, K.; Kristensen, R.M.; Michalczyk, Ł. Revision of Testechiniscus Kristensen, 1987 (Heterotardigrada: Echiniscidae) refutes the polar-temperate distribution of the genus. Zootaxa 2018, 4472, 261–297. [Google Scholar] [CrossRef]
- Schuster, R.O.; Nelson, D.R.; Grigarick, A.A.; Christenberry, D. Systematic criteria of the Eutardigrada. Trans. Am. Microsc. Soc. 1980, 99, 284–303. [Google Scholar] [CrossRef]
- Stec, D.; Krzywański, Ł.; Michalczyk, Ł. Integrative description of Macrobiotus canaricus sp. nov. with notes on M. recens (Eutardigrada: Macrobiotidae). Eur. J. Taxon. 2018, 1–36. [Google Scholar] [CrossRef]
- Nowak, B.; Stec, D. An integrative description of Macrobiotus hannae sp. nov.(Tardigrada: Eutardigrada: Macrobiotidae: Hufelandi group) from Poland. Turk. J. Zool. 2018, 42, 269–286. [Google Scholar] [CrossRef]
- Pilato, G.; Binda, M. Descrizione du una nuova specie di eutardigrado d’Australia Macrobiotus joannae n. sp. Animalia 1983, 10, 262–272. [Google Scholar]
- Coughlan, K.; Stec, D. Two new species of the Macrobiotus hufelandi complex (Tardigrada: Eutardigrada: Macrobiotidae) from Australia and India, with notes on their phylogenetic position. Eur. J. Taxon.. 2019, 573. [Google Scholar] [CrossRef]
- Maucci, W. Prime notizie su Tardigradi “terrestri” del Madagascar con descrizione di tre specie nuove. Boll. Mus. Civ. Stor. Nat. Verona 1993, 17, 381–392. [Google Scholar]
- Roszkowska, M.; Kaczmarek, Ł. Macrobiotus noemiae sp. nov., a new tardigrade species (Macrobiotidae: Hufelandi group) from Spain. Turk. J. Zool. 2019, 43, 331–348. [Google Scholar] [CrossRef]
- Stec, D.; Kristensen, R.M.; Michalczyk, Ł. Integrative taxonomy identifies Macrobiotus papei, a new tardigrade species of the Macrobiotus hufelandi complex (Eutardigrada: Macrobiotidae) from the Udzungwa Mountains National Park (Tanzania). Zootaxa 2018, 4446, 273–291. [Google Scholar] [CrossRef]
- Guidetti, R.; Peluffo, J.R.; Rocha, A.M.; Cesari, M.; de Peluffo, M.C.M. The morphological and molecular analyses of a new South American urban tardigrade offer new insights on the biological meaning of the Macrobiotus hufelandi group of species (Tardigrada: Macrobiotidae). J. Nat. Hist. 2013, 47, 2409–2426. [Google Scholar] [CrossRef]
- Bertolani, R.; Guidetti, R.; Marchioro, T.; Altiero, T.; Rebecchi, L.; Cesari, M. Phylogeny of Eutardigrada: New molecular data and their morphological support lead to the identification of new evolutionary lineages. Mol. Phy. Evol. 2014, 76, 110–126. [Google Scholar] [CrossRef]
- Wełnicz, W.; Grohme, M.A.; Kaczmarek, Ł.; Schill, R.O.; Frohme, M. ITS-2 and 18S rRNA Data from Macrobiotus polonicus and Milnesium tardigradum (Eutardigrada, Tardigrada). J. Zool. Syst. Evol. Res. 2011, 49, 34–39. [Google Scholar] [CrossRef]
- Stec, D.; Morek, W.; Gąsiorek, P.; Blagden, B.; Michalczyk, Ł. Description of Macrobiotus scoticus sp. nov. (Tardigrada: Macrobiotidae: Hufelandi Group) from Scotland by means of integrative taxonomy. Ann. Zool. 2017, 67, 181–197. [Google Scholar] [CrossRef]
- Vecchi, M.; Cesari, M.; Bertolani, R.; Jönsson, K.I.; Rebecchi, L.; Guidetti, R. Integrative systematic studies on tardigrades from Antarctica identify new genera and new species within Macrobiotoidea and Echiniscoidea. Invert. Syst. 2016, 30, 303–322. [Google Scholar] [CrossRef]
- Itang, L.A.M.; Stec, D.; Mapalo, M.A.; Mirano-Bascos, D.; Michalczyk, L. An integrative description of Mesobiotus dilimanensis, a new tardigrade species from the Philippines (Eutardigrada: Macrobiotidae: Furciger group). Raffles Bull. Zool. 2020, 68, 19–31. [Google Scholar]
- Mapalo, M.A.; Stec, D.; Mirano-Bascos, D.; Michalczyk, Ł. Mesobiotus philippinicus sp. nov., the first limnoterrestrial tardigrade from the Philippines. Zootaxa 2016, 4126, 411–426. [Google Scholar] [CrossRef]
- Richters, F. Nordische tardigraden. Zool. Anz. 1903, 168–172. [Google Scholar]
- Murray, J. Tardigrada. The British Antarctic Expedition 1907–1909. Rep. Sci. Inv. 1 Biol. 1910, Part V, 83–187. [Google Scholar]
- Murray, J. XXV.—Arctic Tardigrada, collected by Wm. S. Bruce. Earth Environ. Sci. Trans. R. Soc. Edinb 1907, 45, 669–681. [Google Scholar] [CrossRef] [Green Version]
- Binda, M.G.; Rebecchi, L. Precisazioni su Macrobiotus furciger Murray, 1907, e descrizione di Macrobiotus pilatoi n. sp. (Eutardigrada, Macrobiotidae). Animalia 1992, 19, 101–109. [Google Scholar]
- Kaczmarek, Ł.; Roszkowska, M. A new eutardigrade from Costa Rica with taxonomical and zoogeographical remarks on Costa Rican tardigrades. N. Z. J. Zool. 2016, 43, 234–245. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Kosicki, J.; Roszkowska, M. Tardigrada of Bory Tucholskie National Park, Zaborski Landscape Park, and their surroundings (Pomerania Province, Poland). Turk. J. Zool. 2018, 42, 6–17. [Google Scholar] [CrossRef]
- Roszkowska, M.; Ostrowska, M.; Grobys, D.; Kmita, H.; Kaczmarek, Ł. Some tardigrades from Italy, with an updated checklist of limno-terrestrial species from the country. Acta Zool. Bulg. 2019, 71, 167–174. [Google Scholar]
- Guidetti, R.; Gneuß, E.; Cesari, M.; Altiero, T.; Schill, R.O. Life-history traits and description of the new gonochoric amphimictic Mesobiotus joenssoni (Eutardigrada: Macrobiotidae) from the island of Elba, Italy. Zool. J. Linnean Soc. 2019, 188, 848–859. [Google Scholar] [CrossRef] [Green Version]
- Vuori, T.; Massa, E.; Calhim, S.; Vecchi, M. Tardigrades of Finland: New records and an annotated checklist. Zootaxa 2020, 4851, 477–521. [Google Scholar] [CrossRef] [PubMed]
- Bertolani, R.; Rebecchi, L.; Giovannini, I.; Cesari, M. DNA barcoding and integrative taxonomy of Macrobiotus hufelandi CAS Schultze 1834, the first tardigrade species to be described, and some related species. Zootaxa 2011, 2997, 19–36. [Google Scholar] [CrossRef]
- Cesari, M.; Bertolani, R.; Rebecchi, L.; Guidetti, R. DNA barcoding in tardigrada: The first case study on Macrobiotus macrocalix Bertolani & Rebecchi 1993 (Eutardigrada, Macrobiotidae). Mol. Ecol. Res. 2009, 9, 699–706. [Google Scholar] [CrossRef]
- Cesari, M.; Giovannini, I.; Bertolani, R.; Rebecchi, L. An example of problems associated with DNA barcoding in tardigrades: A novel method for obtaining voucher specimens. Zootaxa 2011, 3104, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Coughlan, K.; Michalczyk, Ł.; Stec, D. Macrobiotus caelestis sp. nov., a new tardigrade species (Macrobiotidae: Hufelandi group) from the Tien Shan Mountains (Kyrgyzstan). Ann. Zool. 2019, 69, 499–513. [Google Scholar] [CrossRef]
- Stec, D.; Arakawa, K.; Michalczyk, Ł. An Integrative Description of Macrobiotus shonaicus sp. nov.(Tardigrada: Macrobiotidae) from Japan with notes on its phylogenetic position within the hufelandi group. PLoS ONE 2018, 13, e0192210. [Google Scholar] [CrossRef] [Green Version]
- Stec, D.; Tumanov, D.T.; Kristensen, R.M. Integrative taxonomy identifies two new tardigrade species (Eutardigrada: Macrobiotidae) from Greenland. Eur. J. Taxon. 2020, 614. [Google Scholar] [CrossRef]
- Stec, D.; Roszkowska, M.; Kaczmarek, Ł.; Michalczyk, Ł. An integrative description of a population of Mesobiotus radiatus (Pilato, Binda & Catanzaro, 1991) from Kenya. Turk. Zool. Derg. 2018, 42, 523–540. [Google Scholar] [CrossRef] [Green Version]
- Mapalo, M.; Stec, D.; Mirano-Bascos, D.; Michalczyk, Ł. An integrative description of a limnoterrestrial tardigrade from the Philippines, Mesobiotus insanis, new species (Eutardigrada: Macrobiotidae: Harmsworthi group). Raffles Bull. Zool. 2017, 65, 440–454. [Google Scholar]
- Stec, D.; Kristensen, R.M. An integrative description of Mesobiotus ethiopicus sp. nov. (Tardigrada: Eutardigrada: Parachela: Macrobiotidae: Harmsworthi group) from the northern Afrotropic region. Turk. J. Zool. 2017, 41, 800–811. [Google Scholar] [CrossRef]
- Stec, D. Mesobiotus datanlanicus sp. nov., a new tardigrade species (Macrobiotidae: Mesobiotus harmsworthi group) from Lâm Đồng Province in Vietnam. Zootaxa 2019, 4679, 164–180. [Google Scholar] [CrossRef]
- Jørgensen, A.; Faurby, S.; Hansen, J.G.; Møbjerg, N.; Kristensen, R.M. Molecular phylogeny of Arthrotardigrada (Tardigrada). Mol. Phyl. Evol. 2010, 54, 1006–1015. [Google Scholar] [CrossRef]
- Schill, R.O.; Steinbrück, G. Identification and differentiation of Heterotardigrada and Eutardigrada species by riboprinting. J. Zool. Syst. Evol. Res. 2007, 45, 184–190. [Google Scholar] [CrossRef]
- Guil, N.; Giribet, G. A comprehensive molecular phylogeny of tardigrades—Adding genes and taxa to a poorly resolved phylum-level phylogeny. Cladistics 2010, 28, 21–49. [Google Scholar] [CrossRef]
- Tumanov, D.V. Integrative description of Mesobiotus anastasiae sp. nov.(Eutardigrada, Macrobiotoidea) and first record of Lobohalacarus (Chelicerata, Trombidiformes) from the Republic of South Africa. Eur. J. Taxon. 2020, 726, 102–131. [Google Scholar]
- Czechowski, P.; Sands, C.J.; Adams, B.J.; D’Haese, C.A.; Gibson, J.A.; McInnes, S.J.; Stevens, M.I. Antarctic Tardigrada: A first step in understanding molecular operational taxonomic units (MOTUs) and biogeography of cryptic meiofauna. Invertebr. Syst. 2012, 26, 526–538. [Google Scholar] [CrossRef]









{kind=link}
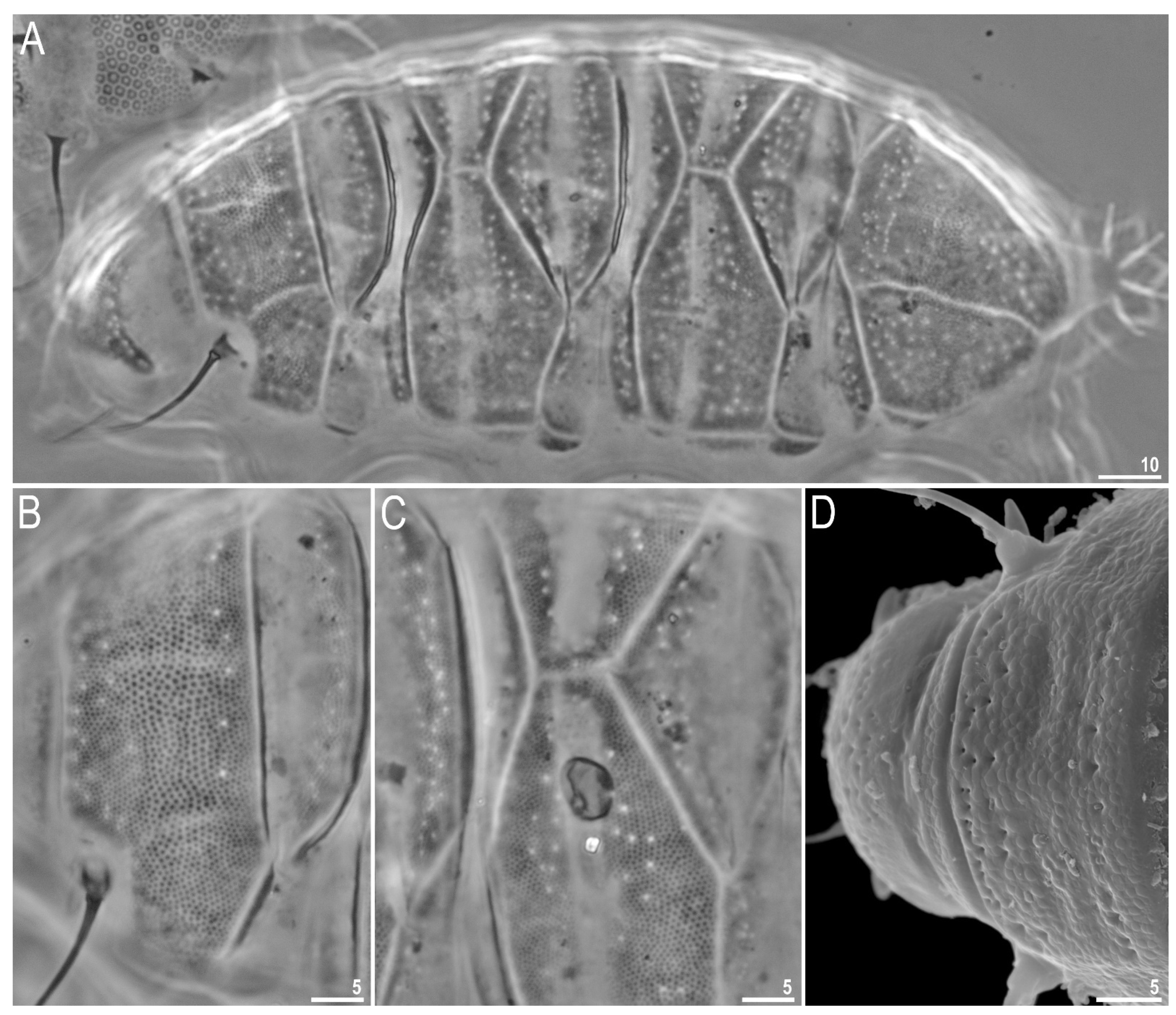{kind=link}
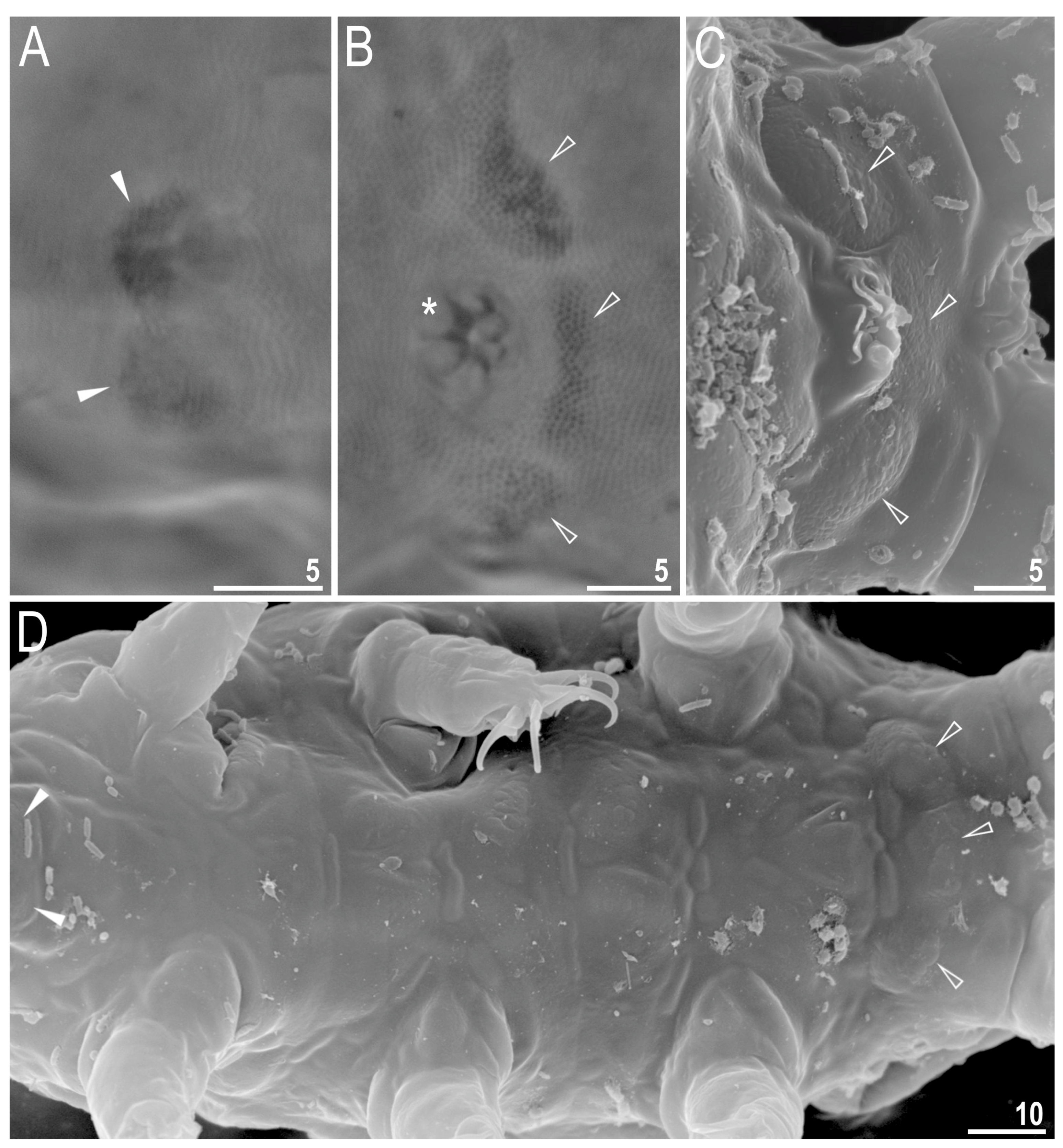{kind=link}
{kind=link}
{kind=link}
{kind=link}
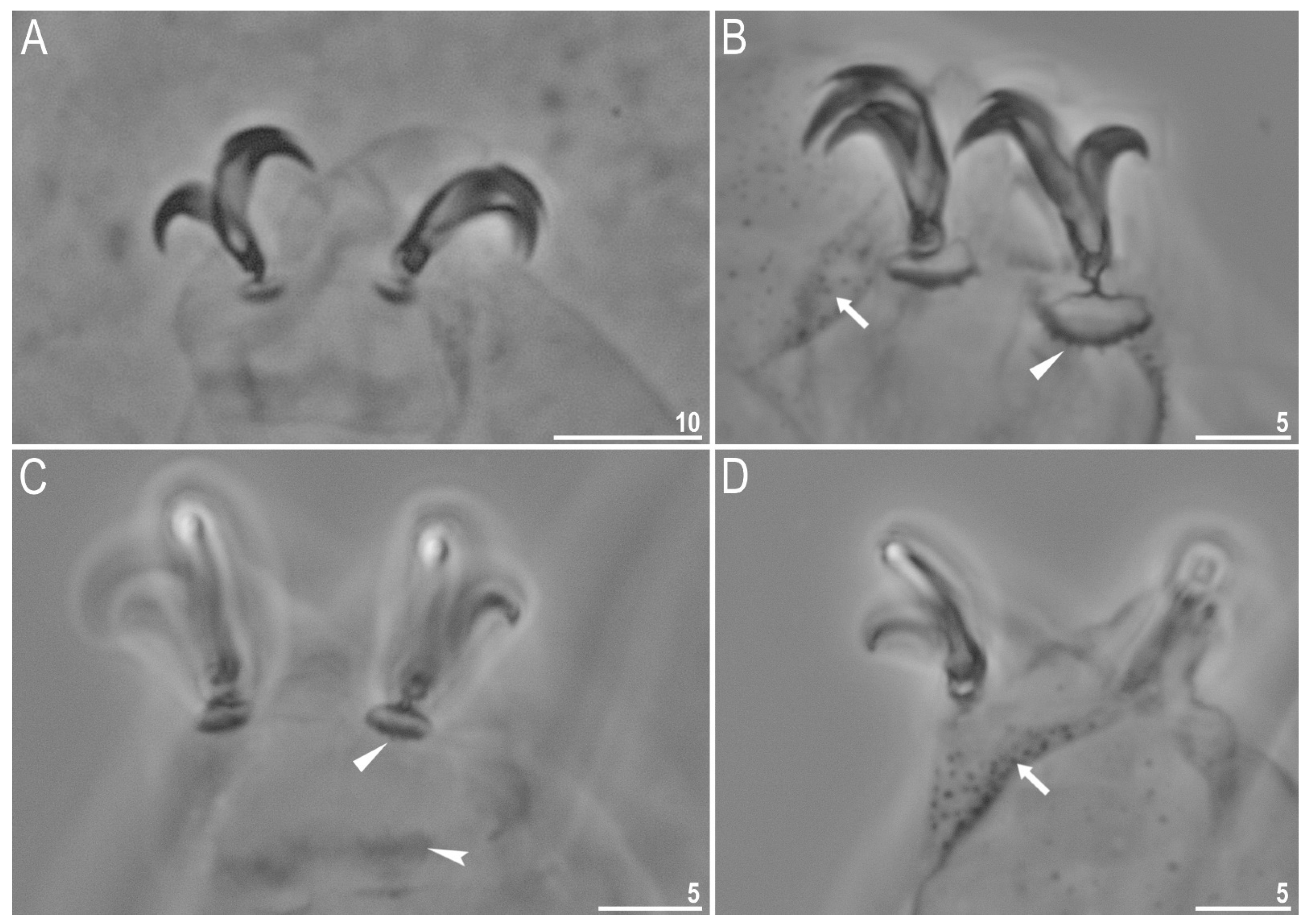{kind=link}
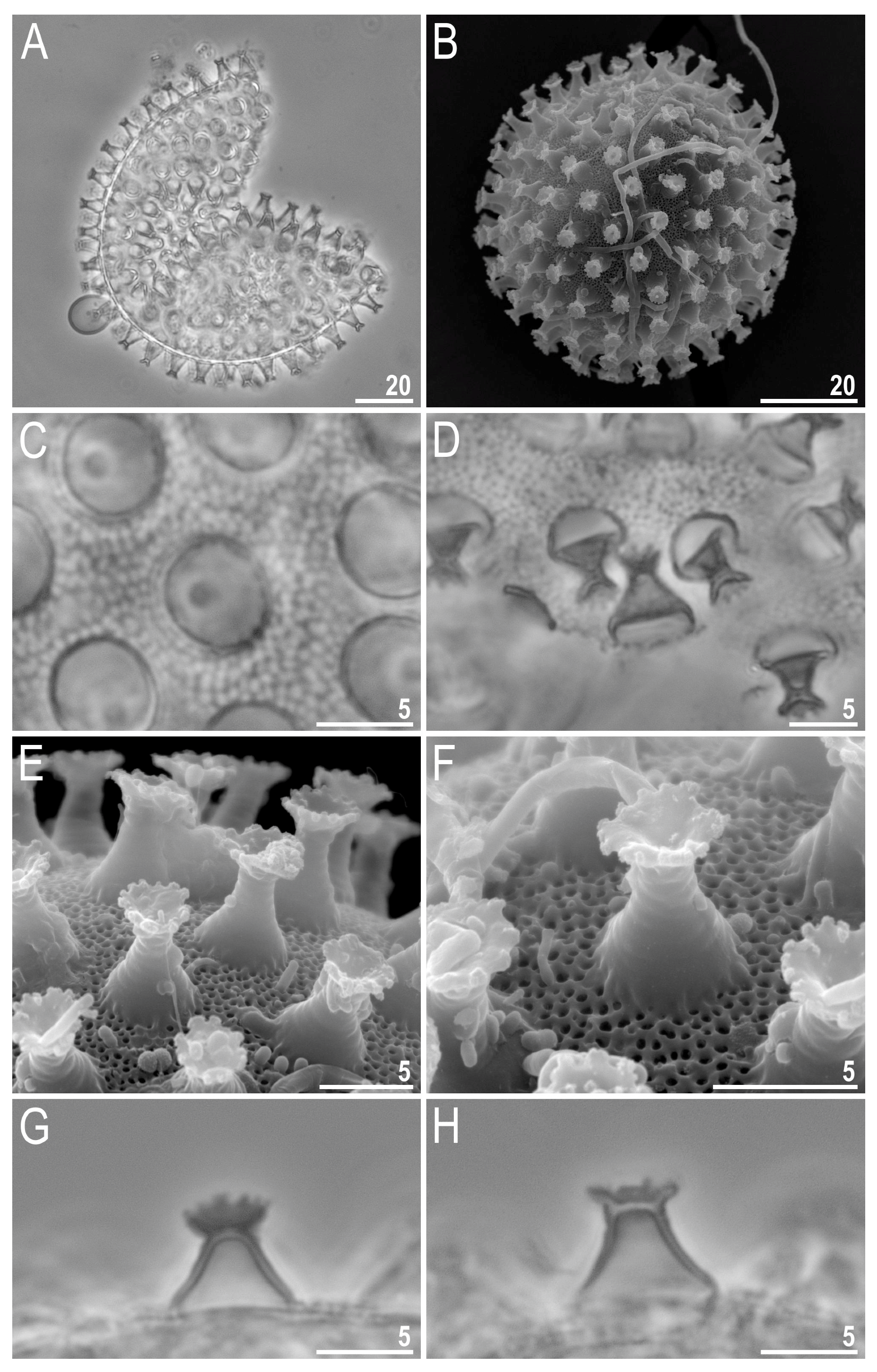{kind=link}
| DNA Fragment | Primer Name/Direction | Primer Sequence (5′-3′) | Source |
|---|---|---|---|
| COI | LCO1490/forward | GGTCAACAAATCATAAAGATATTGG | Folmer et al. [45] |
| HCO2198/reverse | TAAACTTCAGGGTGACCAAAAAATCA | ||
| 18S rRNA | SSU01_F/forward | AACCTGGTTGATCCTGCCAGT | Sands et al. [46] |
| SSU82_R/reverse | TGATCCTTCTGCAGGTTCACCTAC | ||
| 28S rRNA | 28SF0001/forward | ACCCvCynAATTTAAGCATAT | Mironov et al. [47] |
| 28SR0990/reverse | CCTTGGTCCGTGTTTCAAGAC | ||
| ITS-2 | ITS3/forward | GCATCGATGAAGAACGCAGC | White et al. [48] |
| ITS4/reverse | TCCTCCGCTTATTGATATGC |
| Species | GenBank Accession Numbers (Voucher Numbers of Specimens) | |||
|---|---|---|---|---|
| COI | 18S rRNA | 28S rRNA | ITS-2 | |
| Bryodelphax mareki sp. nov. | MW655785-87 (CN8.17/S, CN8.25/S, CN8.28/S) | MW680639-40 (CN8.21/S, CN8.25/S) | MW680637-38 (CN8.21/S, CN8.22/S) | NA |
| Macrobiotus birendrai sp. nov. | MW656266 (CN8.101/S) | MW680641 (CN8.101/S) | MW680644 (CN8.101/S) | MW680418 (CN8.101/S) |
| Mesobiotus skorackii | MW656257 (CN8.115/S) | MW680642-43 (CN8.22/S, CN8.115/S) | MW680636 (CN8.115/S) | NA |
| Character | N | Range | Mean | SD | Holotype | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | sc | µm | sc | µm | sc | µm | sc | ||||||
| Body length | 16 | 122 | – | 181 | 577 | – | 780 | 141 | 689 | 15 | 60 | 146 | 749 |
| Scapular plate length | 16 | 18.4 | – | 24.0 | – | 20.4 | – | 1.7 | – | 19.5 | – | ||
| Head appendages lengths | |||||||||||||
| Cirrus internus | 16 | 5.3 | – | 7.3 | 26.8 | – | 34.6 | 6.2 | 30.6 | 0.5 | 2.7 | 5.3 | 27.0 |
| Cephalic papilla | 15 | 4.1 | – | 5.9 | 21.7 | – | 29.1 | 4.9 | 24.1 | 0.5 | 2.3 | 4.3 | 22.0 |
| Cirrus externus | 16 | 11.1 | – | 13.1 | 53.4 | – | 63.0 | 11.9 | 58.7 | 0.7 | 3.2 | 11.1 | 57.0 |
| Clava | 12 | 4.3 | – | 6.2 | 20.4 | – | 33.0 | 5.2 | 26.1 | 0.5 | 3.7 | 4.8 | 24.6 |
| Cirrus A | 15 | 35.2 | – | 46.7 | 174.1 | – | 212.9 | 39.6 | 193.6 | 3.2 | 10.7 | 36.5 | 186.9 |
| Cirrus A/body length ratio | 15 | 25% | – | 32% | – | 28% | – | 2% | – | 25% | – | ||
| Cirrus int/ext length ratio | 16 | 47% | – | 56% | – | 52% | – | 3% | – | 47% | – | ||
| Body appendages lengths | |||||||||||||
| Papilla on leg IV length | 14 | 1.6 | – | 3.3 | 8.1 | – | 14.3 | 2.3 | 10.9 | 0.5 | 1.9 | 2.2 | 11.3 |
| Claw 1 heights | |||||||||||||
| Branch | 16 | 5.8 | – | 8.4 | 30.3 | – | 42.0 | 7.6 | 37.5 | 0.7 | 2.8 | 7.0 | 35.9 |
| Spur | 13 | 1.3 | – | 1.7 | 5.4 | – | 8.2 | 1.5 | 7.2 | 0.1 | 0.8 | 1.4 | 7.2 |
| Spur/branch length ratio | 13 | 16% | – | 21% | – | 18% | – | 2% | – | 0 | – | ||
| Claw 2 heights | |||||||||||||
| Branch | 16 | 6.7 | – | 8.6 | 30.8 | – | 42.6 | 7.6 | 37.7 | 0.5 | 3.6 | 7.5 | 38.3 |
| Spur | 14 | 1.0 | – | 1.5 | 5.2 | – | 7.2 | 1.3 | 6.6 | 0.1 | 0.7 | 1.4 | 7.2 |
| Spur/branch length ratio | 14 | 15% | – | 20% | – | 18% | – | 1% | – | 0 | – | ||
| Claw 3 heights | |||||||||||||
| Branch | 16 | 5.8 | – | 8.6 | 30.6 | – | 45.1 | 7.5 | 37.3 | 0.8 | 4.0 | 7.0 | 35.6 |
| Spur | 12 | 1.2 | – | 1.5 | 5.4 | – | 7.5 | 1.4 | 6.6 | 0.1 | 0.8 | 1.3 | 6.7 |
| Spur/branch length ratio | 12 | 16% | – | 20% | – | 18% | – | 1% | – | 0 | – | ||
| Claw 4 lengths | |||||||||||||
| Branch | 16 | 7.3 | – | 9.9 | 38.3 | – | 48.2 | 8.9 | 43.8 | 0.6 | 3.7 | 8.8 | 45.2 |
| Spur | 13 | 1.4 | – | 1.9 | 6.3 | – | 8.9 | 1.6 | 7.8 | 0.1 | 0.7 | 1.6 | 8.0 |
| Spur/branch height ratio | 13 | 15% | – | 21% | – | 18% | – | 2% | – | 0 | – | ||
| Character | N | Range | Mean | SD | Holotype | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | µm | pt | ||||||
| Body length | 15 | 271 | – | 589 | 815 | – | 1291 | 449 | 1055 | 97 | 133 | 368 | 978 |
| Buccopharyngeal tube | |||||||||||||
| Buccal tube length | 16 | 33.2 | – | 52.6 | – | 42.7 | – | 5.9 | – | 37.7 | – | ||
| Stylet support insertion point | 16 | 26.2 | – | 43.0 | 78.0 | – | 82.8 | 34.3 | 80.2 | 5.2 | 1.6 | 29.6 | 78.7 |
| Buccal tube external width | 16 | 5.2 | – | 8.4 | 13.5 | – | 17.0 | 6.6 | 15.3 | 1.0 | 1.1 | 5.5 | 14.7 |
| Buccal tube internal width | 16 | 3.4 | – | 5.6 | 8.7 | – | 11.9 | 4.4 | 10.2 | 0.8 | 1.0 | 3.8 | 10.0 |
| Ventral lamina length | 13 | 19.9 | – | 32.9 | 56.0 | – | 66.2 | 25.7 | 60.9 | 4.2 | 3.2 | 22.9 | 60.8 |
| Placoid lengths | |||||||||||||
| Macroplacoid 1 | 16 | 8.2 | – | 16.0 | 23.0 | – | 31.6 | 11.3 | 26.2 | 2.2 | 2.6 | 8.7 | 23.0 |
| Macroplacoid 2 | 16 | 6.0 | – | 11.2 | 17.2 | – | 22.0 | 8.1 | 18.9 | 1.6 | 1.5 | 6.5 | 16.3 |
| Microplacoid | 16 | 2.4 | – | 4.9 | 6.5 | – | 9.6 | 3.5 | 8.0 | 0.8 | 0.9 | 2.8 | 7.4 |
| Macroplacoid row | 16 | 16.3 | – | 30.2 | 49.1 | – | 57.9 | 22.6 | 52.6 | 4.4 | 3.4 | 18.8 | 49.9 |
| Placoid row | 16 | 19.2 | – | 35.2 | 57.9 | – | 67.9 | 26.9 | 62.7 | 4.9 | 3.4 | 23.2 | 61.5 |
| Claw 1 heights | |||||||||||||
| External primary branch | 16 | 8.5 | – | 13.2 | 20.6 | – | 29.8 | 10.8 | 25.3 | 1.6 | 2.3 | 10.0 | 26.5 |
| External secondary branch | 16 | 6.1 | – | 10.3 | 14.5 | – | 22.1 | 8.1 | 19.0 | 1.4 | 2.2 | 7.8 | 20.7 |
| Internal primary branch | 16 | 8.2 | – | 12.7 | 20.3 | – | 27.7 | 10.3 | 24.2 | 1.5 | 1.8 | 9.3 | 24.8 |
| Internal secondary branch | 16 | 6.2 | – | 10.7 | 16.7 | – | 24.3 | 8.4 | 19.6 | 1.4 | 2.3 | 7.8 | 20.8 |
| Claw 2 heights | |||||||||||||
| External primary branch | 16 | 8.1 | – | 14.0 | 23.8 | – | 29.3 | 11.5 | 26.9 | 1.7 | 1.6 | 10.3 | 27.2 |
| External secondary branch | 16 | 7.0 | – | 11.7 | 17.5 | – | 23.9 | 9.0 | 21.1 | 1.5 | 1.9 | 7.5 | 19.8 |
| Internal primary branch | 16 | 8.6 | – | 13.6 | 23.0 | – | 30.7 | 11.1 | 26.0 | 1.6 | 2.1 | 11.1 | 29.5 |
| Internal secondary branch | 16 | 6.8 | – | 11.0 | 16.5 | – | 23.6 | 8.5 | 20.0 | 1.3 | 2.2 | 8.8 | 23.3 |
| Claw 3 heights | |||||||||||||
| External primary branch | 16 | 8.7 | – | 14.8 | 24.9 | – | 31.7 | 11.7 | 27.4 | 1.7 | 2.1 | 10.4 | 27.5 |
| External secondary branch | 16 | 7.1 | – | 11.7 | 17.5 | – | 25.2 | 9.2 | 21.5 | 1.5 | 1.7 | 7.8 | 20.8 |
| Internal primary branch | 16 | 8.2 | – | 13.5 | 22.7 | – | 30.3 | 10.9 | 25.5 | 1.7 | 1.9 | 9.7 | 25.8 |
| Internal secondary branch | 16 | 6.5 | – | 11.1 | 18.0 | – | 23.2 | 8.8 | 20.6 | 1.5 | 1.7 | 8.3 | 21.9 |
| Claw 4 lengths | |||||||||||||
| Anterior primary branch | 16 | 9.5 | – | 15.8 | 23.9 | – | 34.9 | 12.5 | 29.2 | 1.9 | 2.9 | 11.7 | 30.9 |
| Anterior secondary branch | 16 | 6.7 | – | 13.0 | 18.3 | – | 28.7 | 9.5 | 22.2 | 1.7 | 2.7 | 8.9 | 23.5 |
| Posterior primary branch | 16 | 8.4 | – | 14.2 | 23.7 | – | 32.2 | 12.1 | 28.3 | 1.5 | 2.4 | 11.6 | 30.8 |
| Posterior secondary branch | 16 | 6.5 | – | 12.0 | 18.3 | – | 24.3 | 9.2 | 21.5 | 1.7 | 1.9 | 8.3 | 22.1 |
| Character | N | Range | Mean | SD |
|---|---|---|---|---|
| Egg bare diameter | 5 | 74.9–86.9 | 87.2 | 7.6 |
| Egg full diameter | 5 | 87.8–103.9 | 102.4 | 9.3 |
| Process height | 33 | 5.9–9.4 | 7.9 | 1.1 |
| Process base width | 33 | 5.5–9.0 | 6.6 | 0.8 |
| Process base/height ratio | 33 | 65–100% | 84% | 10% |
| Terminal disc width | 33 | 3.2–6.1 | 4.1 | 0.7 |
| Inter-process distance | 33 | 1.0–3.3 | 2.1 | 0.7 |
| Number of processes on the egg circumference | 5 | 22–26 | 23.6 | 2.0 |
| Character | N | Range | Mean | SD | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | ||||||
| Body length | 11 | 281 | – | 485 | 818 | – | 1102 | 418 | 991 | 62 | 76 |
| Buccopharyngeal tube | |||||||||||
| Buccal tube length | 11 | 34.3 | – | 47.6 | – | 42.0 | – | 4.4 | – | ||
| Stylet support insertion point | 11 | 25.9 | – | 35.6 | 74.6 | – | 77.0 | 31.7 | 75.5 | 3.2 | 0.8 |
| Buccal tube external width | 11 | 5.9 | – | 8.0 | 15.2 | – | 18.0 | 7.0 | 16.8 | 0.7 | 0.7 |
| Buccal tube internal width | 11 | 3.8 | – | 5.6 | 11.0 | – | 12.7 | 4.9 | 11.5 | 0.6 | 0.6 |
| Ventral lamina length | 11 | 21.0 | – | 29.8 | 54.4 | – | 62.9 | 25.5 | 60.6 | 2.9 | 2.7 |
| Placoid lengths | |||||||||||
| Macroplacoid 1 | 11 | 4.3 | – | 6.7 | 11.6 | – | 14.3 | 5.5 | 13.1 | 0.7 | 1.0 |
| Macroplacoid 2 | 11 | 3.7 | – | 5.4 | 9.8 | – | 12.6 | 4.7 | 11.3 | 0.5 | 0.8 |
| Macroplacoid 3 | 11 | 4.1 | – | 6.4 | 11.1 | – | 13.8 | 5.3 | 12.5 | 0.7 | 0.9 |
| Microplacoid | 11 | 2.5 | – | 4.8 | 6.7 | – | 10.3 | 3.6 | 8.5 | 0.6 | 1.2 |
| Macroplacoid row | 11 | 15.3 | – | 22.9 | 44.4 | – | 48.2 | 19.4 | 46.2 | 2.4 | 1.2 |
| Placoid row | 11 | 19.7 | – | 27.6 | 54.2 | – | 60.5 | 24.4 | 57.9 | 2.9 | 1.7 |
| Claw 1 heights | |||||||||||
| External primary branch | 11 | 6.9 | – | 11.2 | 18.3 | – | 26.2 | 9.5 | 22.5 | 1.4 | 2.1 |
| External secondary branch | 11 | 5.6 | – | 9.5 | 14.5 | – | 20.7 | 7.1 | 16.9 | 1.3 | 2.1 |
| Internal primary branch | 11 | 6.8 | – | 11.4 | 19.8 | – | 25.9 | 9.5 | 22.6 | 1.3 | 1.7 |
| Internal secondary branch | 11 | 5.3 | – | 8.9 | 15.4 | – | 20.6 | 7.7 | 18.2 | 1.0 | 1.5 |
| Claw 2 heights | |||||||||||
| External primary branch | 11 | 6.9 | – | 11.4 | 20.2 | – | 26.7 | 9.9 | 23.6 | 1.4 | 1.8 |
| External secondary branch | 11 | 5.4 | – | 9.7 | 15.8 | – | 22.7 | 8.0 | 19.0 | 1.6 | 2.6 |
| Internal primary branch | 11 | 7.1 | – | 11.0 | 20.1 | – | 25.6 | 9.2 | 21.9 | 1.0 | 1.6 |
| Internal secondary branch | 10 | 5.5 | – | 10.3 | 16.0 | – | 24.2 | 7.6 | 17.9 | 1.3 | 2.6 |
| Claw 3 heights | |||||||||||
| External primary branch | 11 | 7.8 | – | 11.6 | 22.8 | – | 28.1 | 10.4 | 24.7 | 1.2 | 1.8 |
| External secondary branch | 11 | 5.2 | – | 9.9 | 15.2 | – | 23.1 | 8.2 | 19.4 | 1.3 | 2.7 |
| Internal primary branch | 11 | 6.7 | – | 10.7 | 19.6 | – | 27.2 | 9.5 | 22.6 | 1.2 | 1.9 |
| Internal secondary branch | 11 | 5.3 | – | 9.6 | 15.5 | – | 22.2 | 8.0 | 18.9 | 1.1 | 2.0 |
| Claw 4 lengths | |||||||||||
| Anterior primary branch | 11 | 7.2 | – | 13.0 | 21.0 | – | 29.6 | 11.4 | 27.0 | 1.7 | 2.4 |
| Anterior secondary branch | 11 | 5.9 | – | 10.3 | 17.1 | – | 23.7 | 9.0 | 21.3 | 1.3 | 2.1 |
| Posterior primary branch | 11 | 6.5 | – | 13.2 | 18.9 | – | 30.7 | 10.7 | 25.3 | 2.1 | 3.6 |
| Posterior secondary branch | 11 | 5.2 | – | 10.7 | 15.1 | – | 24.4 | 8.5 | 20.1 | 1.5 | 2.6 |
| Character | N | Range | Mean | SD |
|---|---|---|---|---|
| Egg bare diameter | 16 | 62.8–89.5 | 74.1 | 8.1 |
| Egg full diameter | 16 | 87.6–113.6 | 102.2 | 7.6 |
| Process height | 48 | 13.5–20.4 | 17.1 | 1.7 |
| Process base width | 48 | 12.7–19.0 | 16.1 | 1.4 |
| Process base/height ratio | 48 | 72–127% | 95% | 12% |
| Inter-process distance | 48 | 1.3–5.6 | 3.5 | 1.2 |
| Number of processes on the egg circumference | 16 | 9–13 | 11.1 | 1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kayastha, P.; Roszkowska, M.; Mioduchowska, M.; Gawlak, M.; Kaczmarek, Ł. Integrative Descriptions of Two New Tardigrade Species along with the New Record of Mesobiotus skorackii Kaczmarek et al., 2018 from Canada. Diversity 2021, 13, 394. https://doi.org/10.3390/d13080394
Kayastha P, Roszkowska M, Mioduchowska M, Gawlak M, Kaczmarek Ł. Integrative Descriptions of Two New Tardigrade Species along with the New Record of Mesobiotus skorackii Kaczmarek et al., 2018 from Canada. Diversity. 2021; 13(8):394. https://doi.org/10.3390/d13080394
Chicago/Turabian StyleKayastha, Pushpalata, Milena Roszkowska, Monika Mioduchowska, Magdalena Gawlak, and Łukasz Kaczmarek. 2021. "Integrative Descriptions of Two New Tardigrade Species along with the New Record of Mesobiotus skorackii Kaczmarek et al., 2018 from Canada" Diversity 13, no. 8: 394. https://doi.org/10.3390/d13080394
APA StyleKayastha, P., Roszkowska, M., Mioduchowska, M., Gawlak, M., & Kaczmarek, Ł. (2021). Integrative Descriptions of Two New Tardigrade Species along with the New Record of Mesobiotus skorackii Kaczmarek et al., 2018 from Canada. Diversity, 13(8), 394. https://doi.org/10.3390/d13080394