
Experimental Warming Effects on Prokaryotic Growth and Viral Production in Coastal Waters of the Northwest Pacific during the Cold Season



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Viral Production and Prokaryotic Growth Rate Experiments
2.3. Determination of Prokaryotic Mortality by Viruses and Burst Size
2.4. Enumeration of Viral and Prokaryotic Abundance by Flow Cytometry (FCM)
2.5. Statistical Analysis
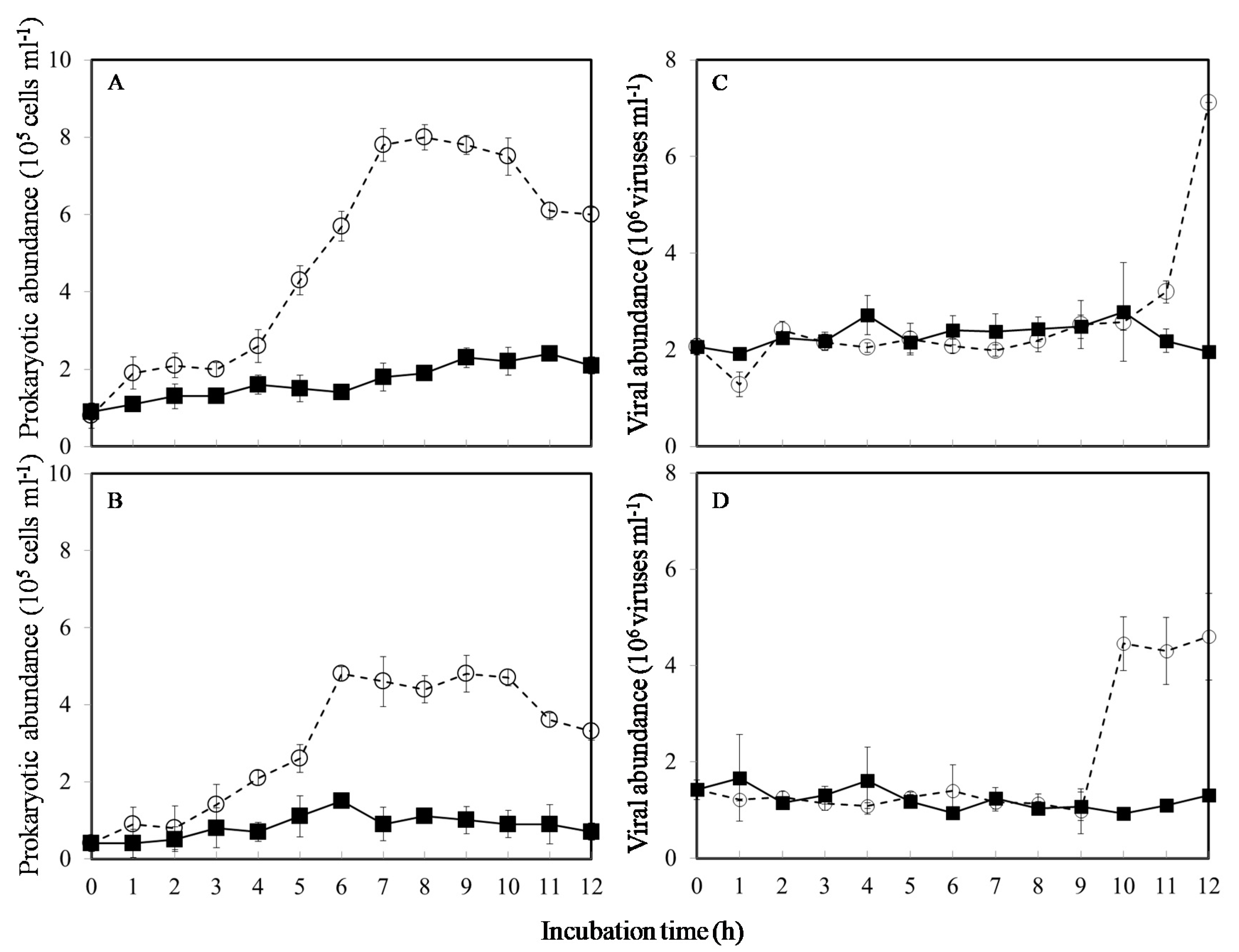
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wommack, K.E.; Colwell, R.R. Virioplankton: Viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 2000, 64, 69–114. [Google Scholar] [CrossRef] [Green Version]
- Rivkin, R.B.; Legendre, L. Biogenic carbon cycling in the upper ocean: Effects of microbial respiration. Science 2001, 291, 2398–2400. [Google Scholar] [CrossRef]
- Rivkin, R.B.; Anderson, M.R.; Lajzerowicz, C. Microbial processes in cold oceans. I. Relationship between temperature and bacterial growth rate. Aquat. Microb. Ecol. 1996, 10, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Domínguez, E.; Vaqué, D.; Gasol, J.M. Temperature effects on the heterotrophic bacteria, heterotrophic nanoflagellates and microbial top predators of the NW Mediterranean. Aquat. Microb. Ecol. 2012, 67, 107–121. [Google Scholar] [CrossRef]
- Mojica, K.D.A.; Huisman, J.; Wilhelm, S.W.; Brussaard, C.P.D. Latitudinal variation in virus-induced mortality of phytoplankton across the North Atlantic Ocean. ISME J. 2015, 10, 500–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendrick, B.J.; DiTullio, G.R.; Cyronak, T.J.; Fulton, J.M.; Van Mooy, B.A.S.; Bidle, K.D. Temperature induced viral resistance in Emiliania huxleyi (Prymnesiophyceae). PLoS ONE 2014, 9, e112134-14. [Google Scholar] [CrossRef] [PubMed]
- Pradeep Ram, A.S.; Boucher, D.; Sime-Ngando, T.; Debroas, D.; Romagoux, J.C. Phage bacteriolysis, protistan bacterivory potential, and bacterial production in a freshwater reservoir: Coupling with temperature. Microb. Ecol. 2005, 50, 64–72. [Google Scholar] [PubMed]
- Tsai, A.Y.; Chiang, K.P.; Chang, J.; Gong, G.C. Seasonal diel variations of picoplankton and nanoplankton in a subtropical Western Pacific coastal ecosystem. Limnol. Oceanogr. 2005, 50, 1221–1231. [Google Scholar] [CrossRef]
- Tsai, A.Y.; Gong, G.C.; Hung, J. Seasonal variations of virus- and nanoflagellate-mediated mortality of heterotrophic bacteria in the coastal ecosystem of subtropical western Pacific. Biogeosciences 2013, 10, 3055–3065. [Google Scholar] [CrossRef] [Green Version]
- Timmermann, A.; Oberhuber, J.; Bacher, A.; Esch, M.; Latif, M.; Roeckner, E. Increased El Niño frequency in a climate model forced by future greenhouse warming. Nature 1999, 398, 694–697. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Brigden, S.M.; Suttle, C.A. A dilution technique for the direct measurement of viral production: A comparison in stratified and tidally mixed coastal waters. Microb. Ecol. 2002, 43, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Shelford, E.J.; Middelboe, M.; Møller, E.F.; Suttle, C.A. Virus-driven nitrogen cycling enhances phytoplankton growth. Aquat. Microb. Ecol. 2012, 66, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Mei, M.L.; Danovaro, R. Virus production and life strategies in aquatic sediments. Limnol. Oceanogr. 2004, 49, 459–470. [Google Scholar] [CrossRef]
- Danovaro, R.; Corinaldesi, C.; Filippini, M.; Fischer, U.R.; Gessner, M.R.; Jaqchet, S.; Magagnini, M.; Velimirov, B. Viriobenthos in freshwater and marine sediments: A review. Freshw. Biol. 2008, 53, 1186–1213. [Google Scholar] [CrossRef]
- Brussaard, C.P.D. Optimization of procedures for counting viruses by flow cytometry. Appl. Environ. Microbiol. 2004, 70, 1506–1513. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Lu, J.R.; Binder, B.J.; Liu, Y.C.; Hodson, R.E. Application of digital image analysis and flow cytometry to enumerate marine viruses stained with SYBR Gold. Appl. Environ. Microbiol. 2001, 67, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Hammes, F.; Egli, T. Cytometric methods for measuring bacteria in water: Advantages, pitfalls and applications. Anal. Bioanal. Chem. 2010, 397, 1083–1095. [Google Scholar] [CrossRef]
- Pomeroy, L.R.; Wiebe, W.J. Temperature and substrates as interactive limiting factors for marine heterotrophic bacteria. Aquat. Microb. Ecol. 2001, 23, 187–204. [Google Scholar] [CrossRef] [Green Version]
- Li, W.K.W. Annual average abundance of heterotrophic bacteria and Synechococcus in surface ocean waters. Limnol. Oceanogr. 1998, 43, 1746–1753. [Google Scholar] [CrossRef]
- Christoffersen, K.; Andersen, N.; Søndergaard, M.; Liboriussen, L.; Jeppesen, E. Implications of climateenforced temperature increases on freshwater pico- and nanoplankton populations studied in artificial ponds during 16 months. Hydrobiologia 2006, 560, 259–266. [Google Scholar] [CrossRef]
- Tsai, A.Y.; Gong, G.C.; Shiau, W. Viral lysis and nanoflagellate grazing on prokaryotes: Effects of short-term warming in a coastal subtropical marine system. Hydrobiologia 2015, 751, 43–54. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Rowe, J.M.; Wilhelm, S.W. Determining rates of virus production in aquatic systems by the virus reduction approach. Manu. Aquat. Viral Ecol. 2010, 1, 1–8. [Google Scholar]
- Steenhauer, L.M.; Wierenga, J.; Carreira, C.; Limpens, R.W.; Koster, A.J.; Pollard, P.C.; Brussaard, C.P. Isolation of cyanophage CrV infecting Cylindrospermopsis raciborskii and the influence of temperature and irradiance on CrV proliferation. Aquat. Microb. Ecol. 2016, 78, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Maat, D.S.; Prins, M.A.; Brussaard, C.P.D. Sediments from arctic tide-water glaciers remove coastal marine viruses and delay host infection. Viruses 2019, 11, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danovaro, R.; Corinaldesi, C.; Dell’Anno, A.; Fuhrman, J.A.; Middelburg, J.J.; Noble, R.T.; Suttle, C.A. Marine viruses and global climate change. FEMS Microbiol. Rev. 2011, 35, 993–1034. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lymer, D.; Logue, J.B.; Brussaard, C.P.D.; Baudoux, A.C.; Vrede, K.; Lindström, E.S. Temporal variation in freshwater viral and bacterial community composition. Freshw. Biol. 2008, 53, 1163–1175. [Google Scholar] [CrossRef]
- Egilmez, H.I.; Morozov, A.Y.; Clokie, M.R.J.; Shan, J.; Letarov, A.; Galyov, E.E. Temperature-dependent virus lifecycle choices may reveal and predict facets of the biology of opportunistic pathogenic bacteria. Sci. Rep. 2018, 8, 8642. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Temperature | Prokaryotic Growth Rate a | VPR b | Viral Production c | Prokaryotic Mortality c | Burst Size | |
|---|---|---|---|---|---|---|
| Nov | (°C) | (h−1) | (106 Viruses mL−1 h−1) | (105 Cells mL−1 h−1) | (Viruses Prokaryote−1) | |
| In situ | 22 ± 0.5 | 0.08 ± 0.06 | 9.1 ± 1.2 | nd | nd | nd |
| Increased temperature | 24 ± 0.5 | 0.34 ± 0.09 * | 10.3 ± 1.6 | 1.44 | 0.68 | 21 |
| Dec | ||||||
| In situ | 20 ± 0.5 | 0.06 ± 0.04 | 13.2 ± 2.6 | nd | nd | nd |
| Increased temperature | 23 ± 1.0 | 0.41 ± 0.05 * | 10.6 ± 1.8 | 0.77 | 0.56 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, A.-Y.; Gong, G.-C.; Mukhanov, V. Experimental Warming Effects on Prokaryotic Growth and Viral Production in Coastal Waters of the Northwest Pacific during the Cold Season. Diversity 2021, 13, 409. https://doi.org/10.3390/d13090409
Tsai A-Y, Gong G-C, Mukhanov V. Experimental Warming Effects on Prokaryotic Growth and Viral Production in Coastal Waters of the Northwest Pacific during the Cold Season. Diversity. 2021; 13(9):409. https://doi.org/10.3390/d13090409
Chicago/Turabian StyleTsai, An-Yi, Gwo-Ching Gong, and Vladimir Mukhanov. 2021. "Experimental Warming Effects on Prokaryotic Growth and Viral Production in Coastal Waters of the Northwest Pacific during the Cold Season" Diversity 13, no. 9: 409. https://doi.org/10.3390/d13090409
APA StyleTsai, A. -Y., Gong, G. -C., & Mukhanov, V. (2021). Experimental Warming Effects on Prokaryotic Growth and Viral Production in Coastal Waters of the Northwest Pacific during the Cold Season. Diversity, 13(9), 409. https://doi.org/10.3390/d13090409