
Streptomyces Application Triggers Reassembly and Optimization of the Rhizosphere Microbiome of Cucumber

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pot Experiment
2.2. Sampling and Biomass Estimation
2.3. DNA Extraction, PCR Amplification, and High-Throughput Sequencing
2.4. Bioinformatics Analysis
2.5. Data Analysis and Statistics
3. Results
3.1. Biomass of Cucumber Plants
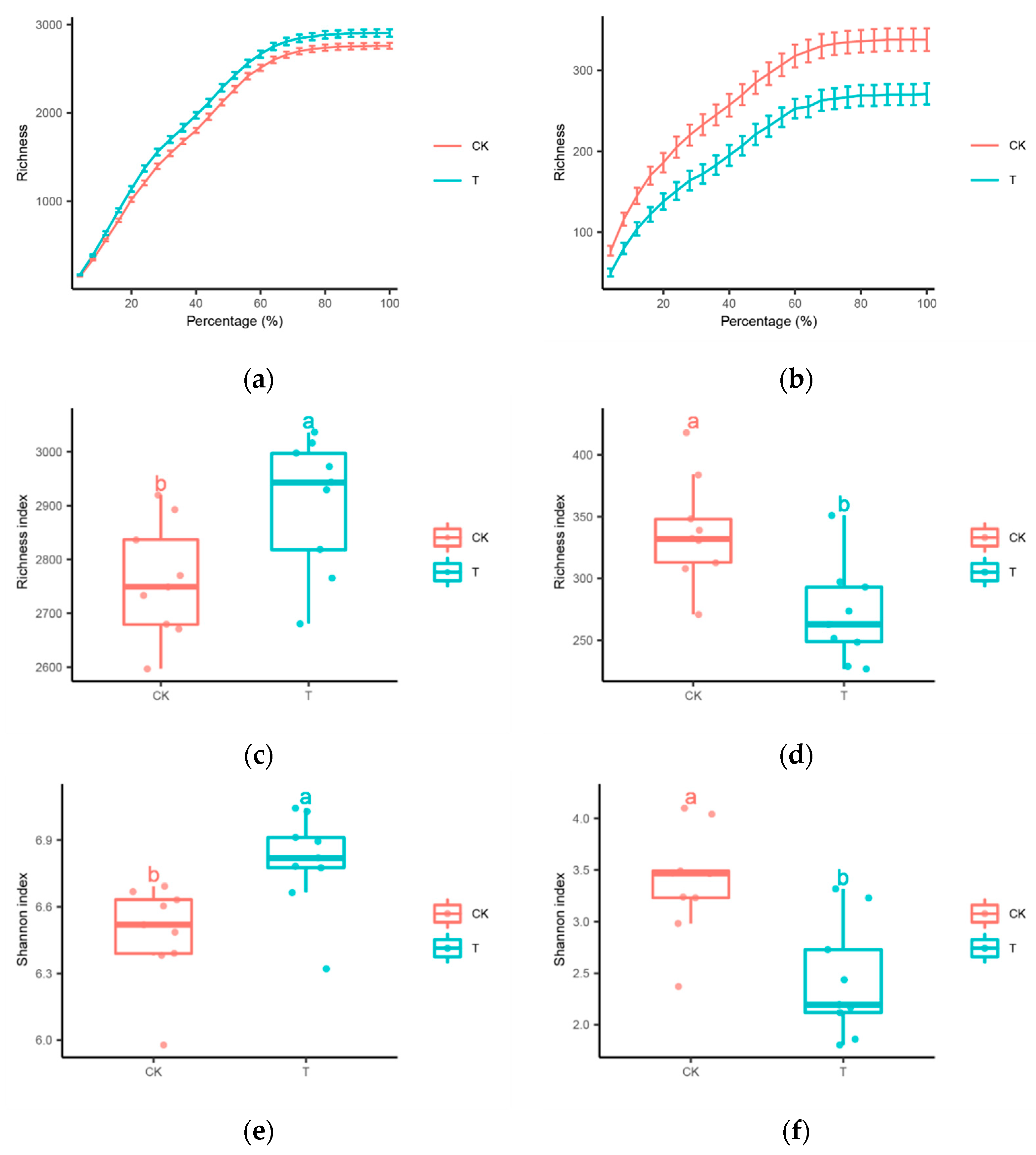
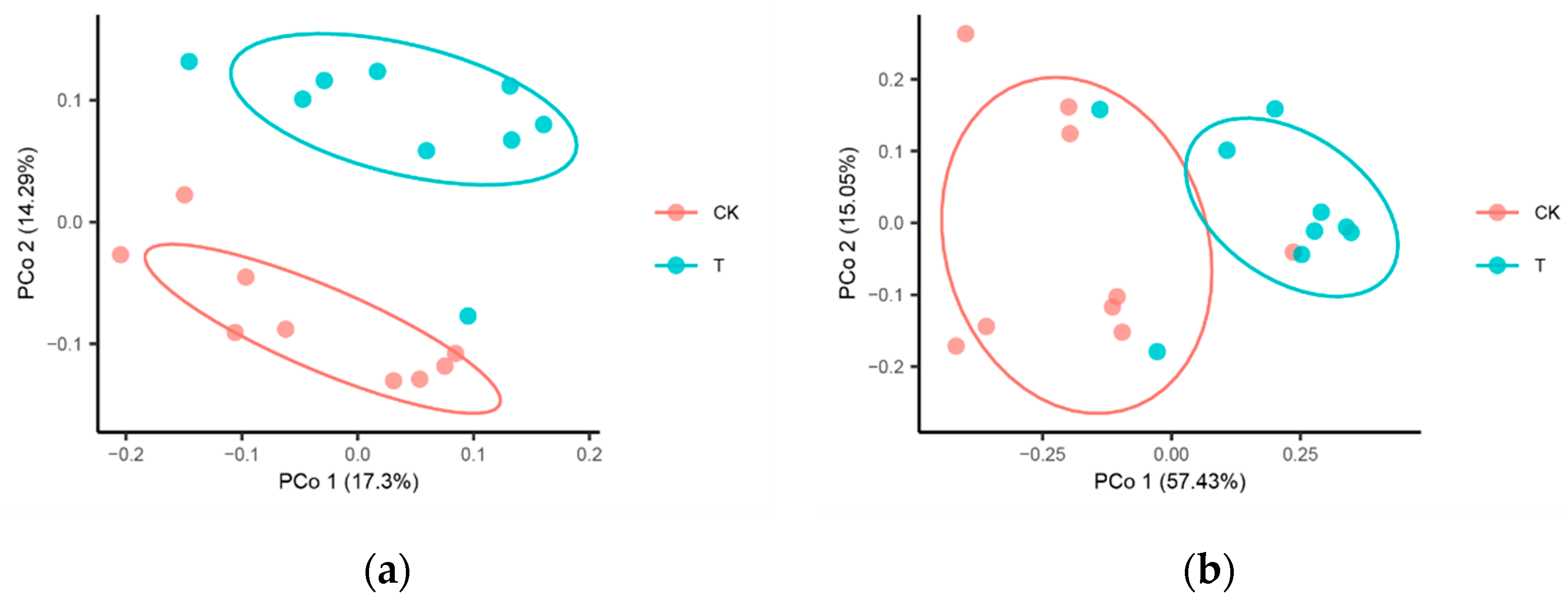
3.2. Distinctive Microbial Community Diversity
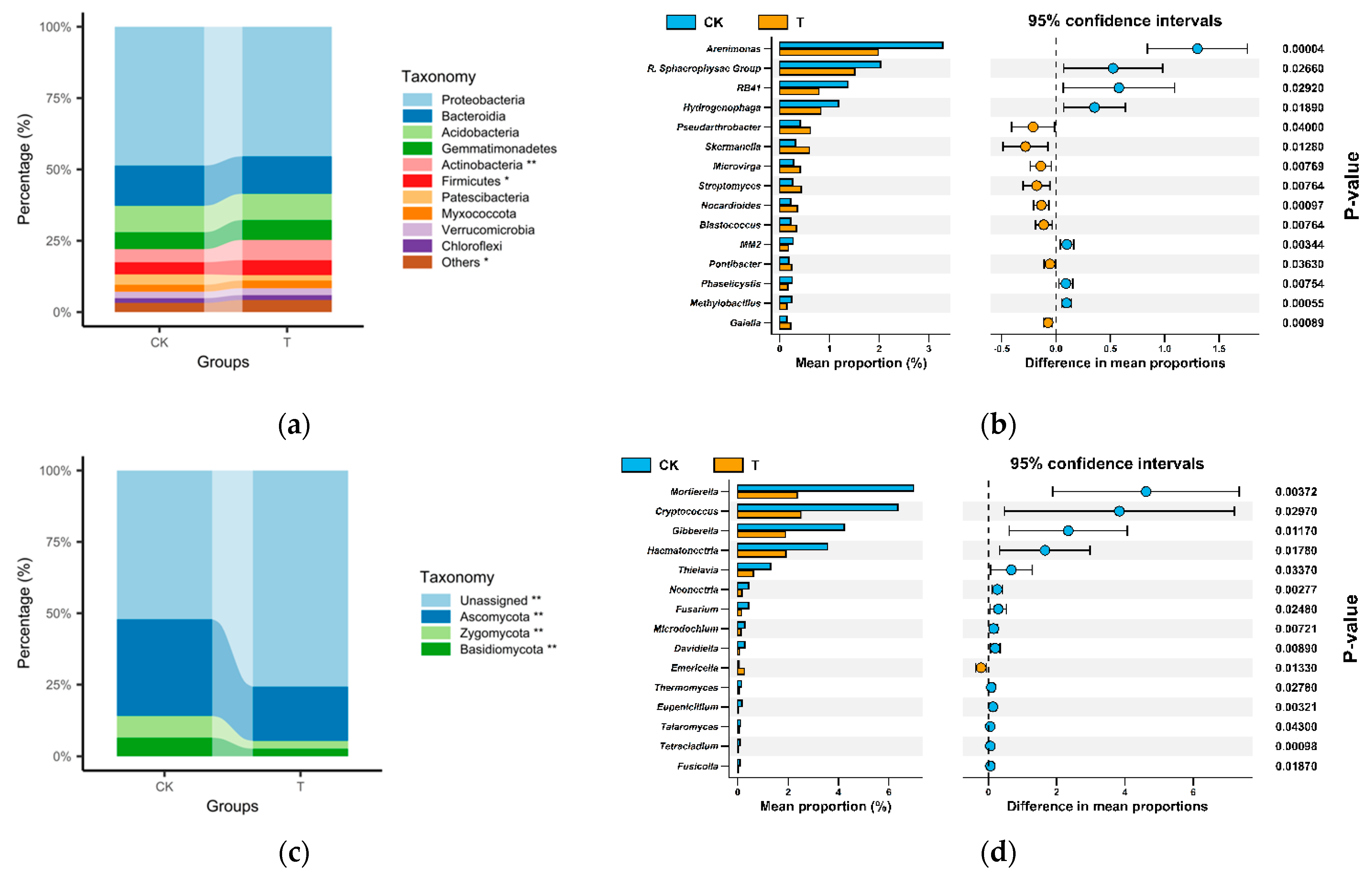
3.3. Shifted Microbial Community Composition
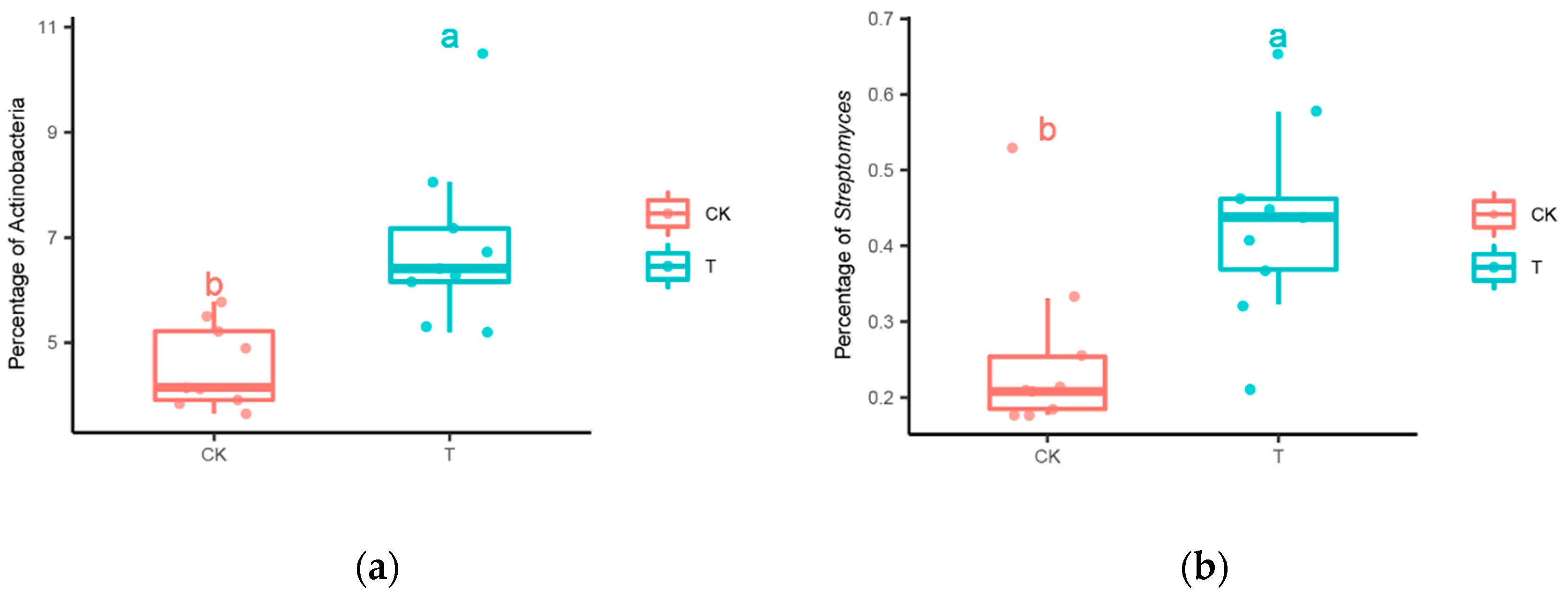
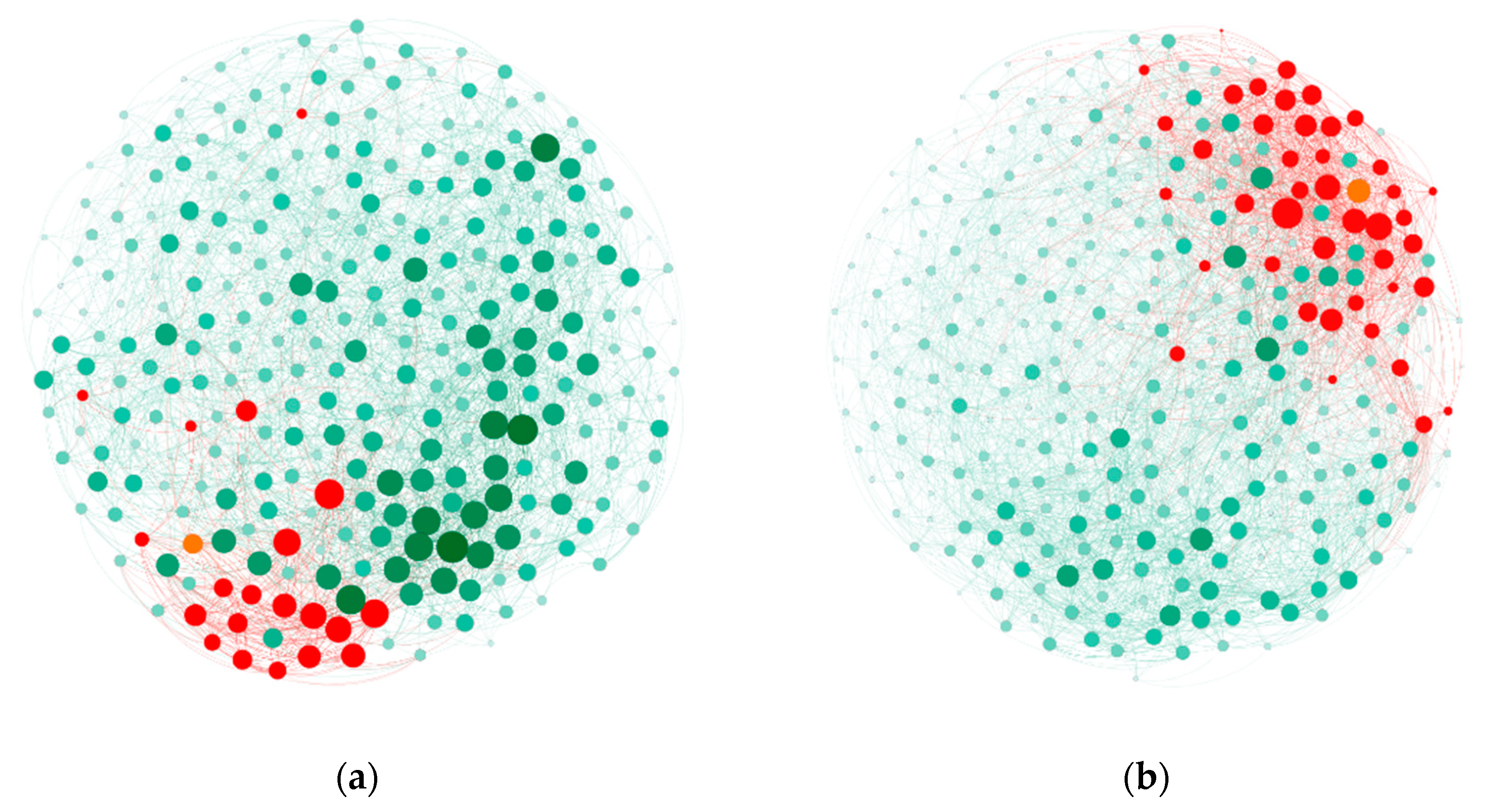
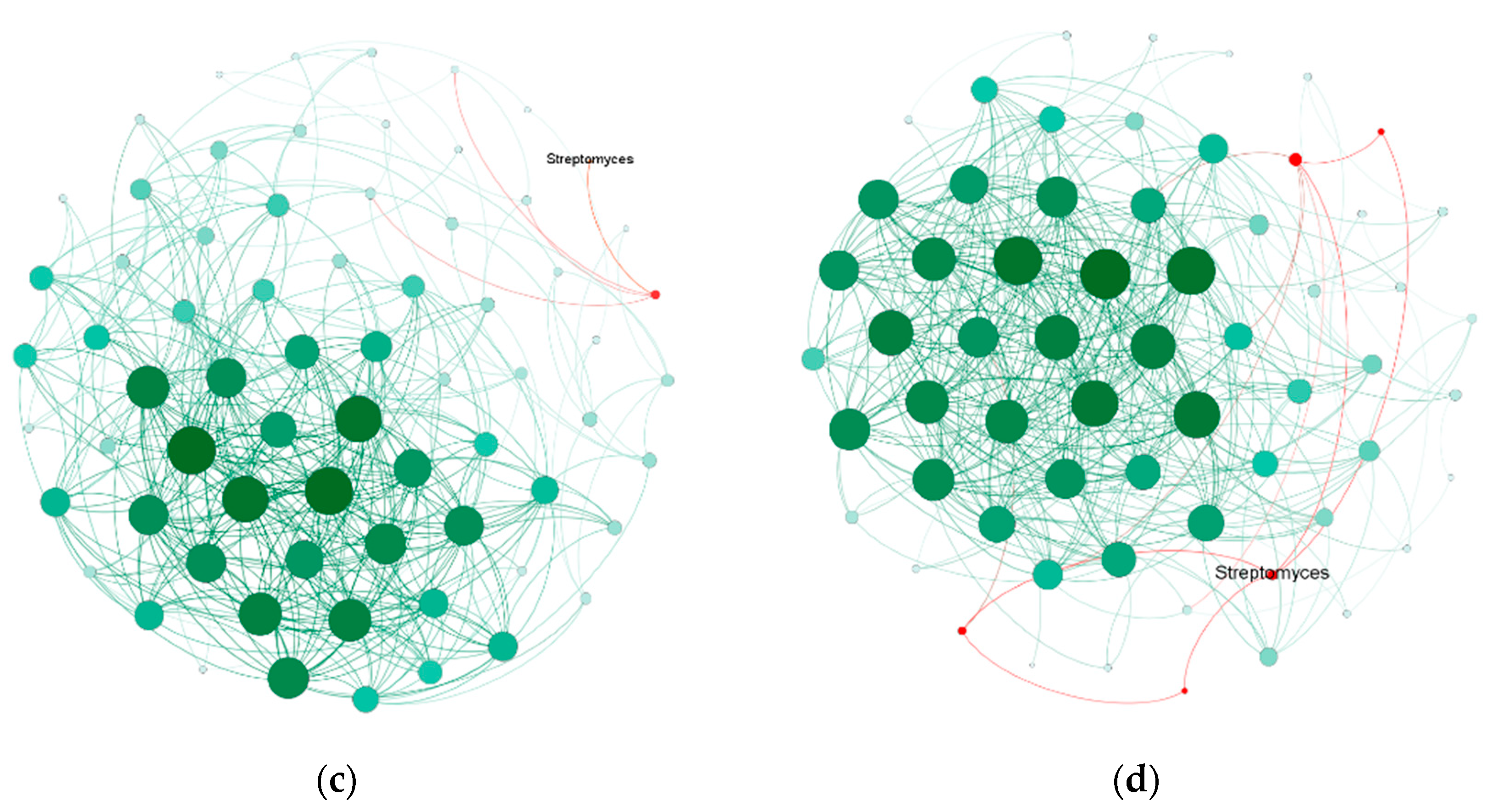
3.4. Increased Streptomyces Abundance and Enriched Bacterial Networks
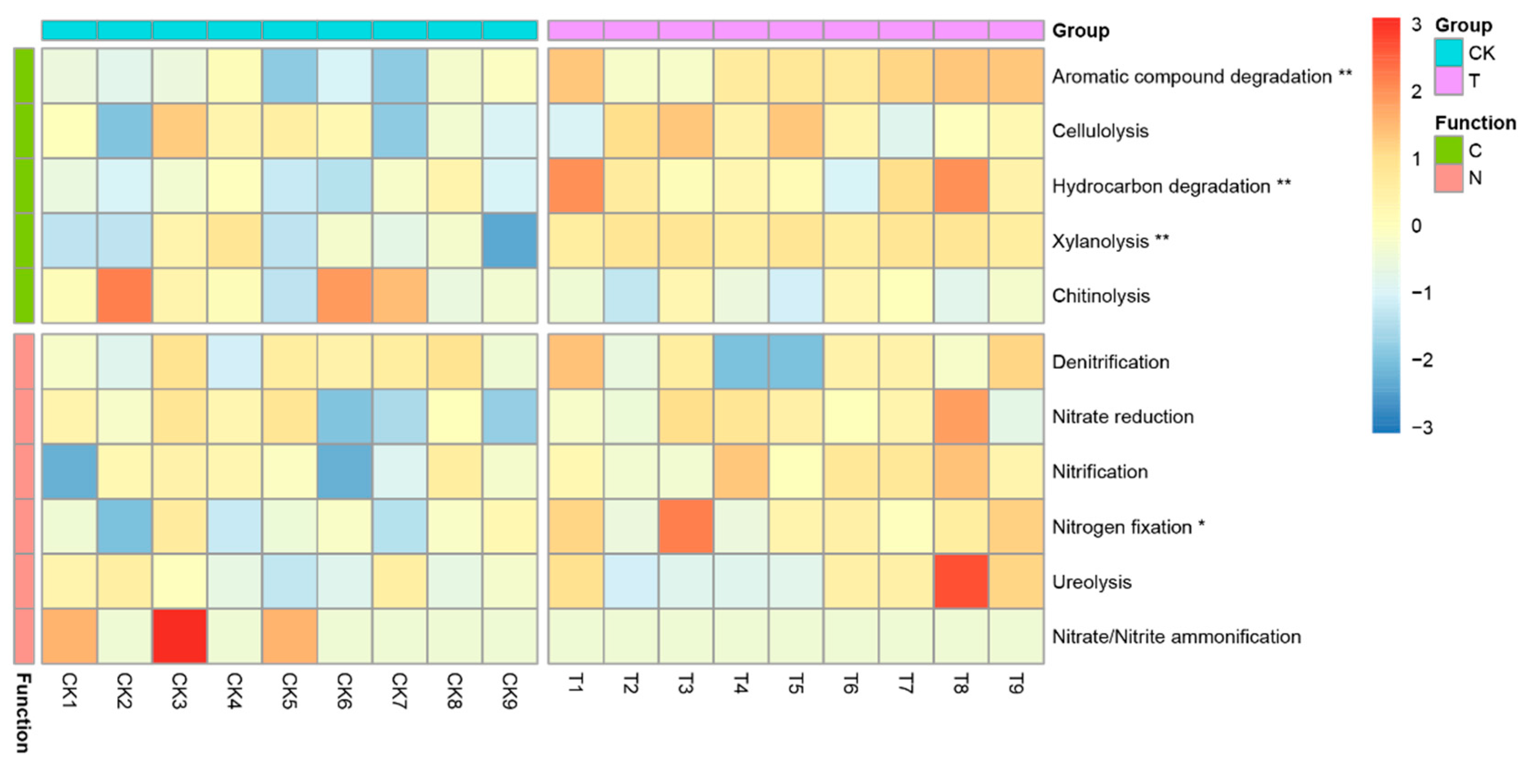
3.5. Potential Functions of Rhizosphere Microbial Communities
4. Discussion
4.1. Rhizosphere Microbiome Diversity and Structure Influenced by Streptomyces Application
4.2. Bacterial Co-Occurrence Enhanced by Streptomyces in the Rhizosphere
4.3. Reassembled Rhizosphere Microbiome Function May Be More Conducive to Plant Survival
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cordovez, V.; Dini-Andreote, F.; Carrion, V.J.; Raaijmakers, J.M. Ecology and evolution of plant microbiomes. Annu. Rev. Microbiol. 2019, 73, 69–88. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellin, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Natl. Acad. Sci. USA 2015, 112, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Zhao, X.; Li, Y.; Xie, Z.; Wang, R. First report of Fusarium tricinctum causing stem and root rot on Lanzhou lily (Lilium davidii var. unicolor) in China. Plant Dis. 2014, 98, 999–1000. [Google Scholar] [CrossRef]
- Wang, T.; Hao, Y.; Zhu, M.; Yu, S.; Ran, W.; Xue, C.; Ling, N.; Shen, Q. Characterizing differences in microbial community composition and function between Fusarium wilt diseased and healthy soils under watermelon cultivation. Plant Soil 2019, 438, 421–433. [Google Scholar] [CrossRef]
- Yuan, J.; Wen, T.; Zhang, H.; Zhao, M.; Penton, C.R.; Thomashow, L.S.; Shen, Q. Predicting disease occurrence with high accuracy based on soil macroecological patterns of Fusarium wilt. ISME J. 2020, 14, 2936–2950. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Zhao, J.; Wen, T.; Zhao, M.; Li, R.; Goossens, P.; Huang, Q.; Bai, Y.; Vivanco, J.M.; Kowalchuk, G.A.; et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome 2018, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bakker, P.A.H.M.; Pieterse, C.M.J.; de Jonge, R.; Berendsen, R.L. The soil-borne legacy. Cell 2018, 172, 1178–1180. [Google Scholar] [CrossRef] [Green Version]
- Viaene, T.; Langendries, S.; Beirinckx, S.; Maes, M.; Goormachtig, S. Streptomyces as a plant’s best friend? FEMS Microbiol. Ecol. 2016, 92, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Guo, T.; Tan, X.; Zhu, W.; Wei, X.; Wang, D.; Xue, Q. Comparison of microecological characterization in rhizosphere soil between healthy and un-healthy plants in continuous cropping potato fields. Acta Agron. Sin. 2013, 39, 2055–2064. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, K.; Ma, J.; Lai, H.; Xue, Q. Effects of two Streptomyces strains on growth and induced resistance of wheat seedlings. Acta Agric. Boreali-Occident. Sin. 2018, 27, 658–666. [Google Scholar] [CrossRef]
- Ma, J.; Liu, Y.; Li, Y.; Sun, Y.; Yang, B.; Lai, H.; Xue, Q. Effects and mechanism of two Streptomyces strains on promoting plant growth and increasing grain yield of maize. Chin. J. Appl. Ecol. 2017, 28, 315–326. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Q.; Li, Y.; Sun, Y.; Xue, Q.; Lai, H. Streptomyces pactum Act12 controls tomato yellow leaf curl virus disease and alters rhizosphere microbial communities. Biol. Fertil. Soils 2019, 55, 149–169. [Google Scholar] [CrossRef]
- Li, H.; Guo, Q.; Jing, Y.; Liu, Z.; Zheng, Z.; Sun, Y.; Xue, Q.; Lai, H. Application of Streptomyces pactum Act12 enhances drought resistance in wheat. J. Plant Growth Regul. 2020, 39, 122–132. [Google Scholar] [CrossRef]
- Cao, S.; Wang, W.; Wang, F.; Zhang, J.; Wang, Z.; Yang, S.; Xue, Q. Drought-tolerant Streptomyces pactum Act12 assist phytoremediation of cadmium-contaminated soil by Amaranthus hypochondriacus: Great potential application in arid/semi-arid areas. Environ. Sci. Pollut. Res. 2016, 23, 14898–14907. [Google Scholar] [CrossRef]
- Mao, N.; Xue, Q.; Tang, M.; Wang, L.; Zhao, J.; Duan, C. Degradation of para-hydroxybenzoic acid by Actinomyces and its effects on strawberry growth. J. Agric. Sci. Tech. China 2010, 12, 103–108. [Google Scholar] [CrossRef]
- Chen, J.; Xue, Q.H.; McErlean, C.S.P.; Zhi, J.H.; Ma, Y.Q.; Jia, X.T.; Ye, X.X. Biocontrol potential of the antagonistic microorganism Streptomyces enissocaesilis against Orobanche cumana. Biocontrol 2016, 61, 781–791. [Google Scholar] [CrossRef]
- Duan, X.; Zhao, F.; Yan, X.; Xue, Q.; Li, X.; Wen, B.; Jia, L.; Yan, H. Construction of SPA7074-deficient mutant of biocontrol strain Streptomyces pactum Act12 and characterization of its secondary metabolites. Acta Microbiol. Sin. 2016, 56, 1883–1891. [Google Scholar] [CrossRef]
- Arseneault, T.; Pieterse, C.M.J.; Gerin-Ouellet, M.; Goyer, C.; Filion, M. Long-term induction of defense gene expression in potato by Pseudomonas sp. LBUM223 and Streptomyces scabies. Phytopathology 2014, 104, 926–932. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Ferraz, H.G.M.; Resende, R.S.; Moreira, P.C.; Silveira, P.R.; Milagres, E.A.; Oliveira, J.R.; Rodrigues, F.A. Antagonistic rhizobacteria and jasmonic acid induce resistance against tomato bacterial spot. Bragantia 2015, 74, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Salla, T.D.; Astarita, L.V.; Santarem, E.R. Defense responses in plants of Eucalyptus elicited by Streptomyces and challenged with Botrytis cinerea. Planta 2016, 243, 1055–1070. [Google Scholar] [CrossRef]
- Zhang, H.; Xue, Q.; Shen, G.; Wang, D. Effects of Actinomycetes agent on ginseng growth and rhizosphere soil microflora. Chin. J. Appl. Ecol. 2013, 24, 2287–2293. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Q.; He, F.; Li, Y.; Xue, Q.; Lai, H. Biocontrol of root diseases and growth promotion of the tuberous plant Aconitum carmichaelii induced by Actinomycetes are related to shifts in the rhizosphere microbiota. Microb. Ecol. 2020, 79, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Shi, M.; Chen, L.; Zhou, J.; Zhang, L.; Li, Y.; Xue, Q.; Lai, H. The biocontrol agent Streptomyces pactum increases Pseudomonas koreensis populations in the rhizosphere by enhancing chemotaxis and biofilm formation. Soil Biol. Biochem. 2020, 144, 107755. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, J.; Pan, D.; Ge, X.; Chen, S.; Wu, F. P-coumaric can alter the composition of cucumber rhizosphere microbial communities and induce negative plant-microbial interactions. Biol. Fertil. Soils 2018, 54, 363–372. [Google Scholar] [CrossRef]
- Duan, C.; Xue, Q.; Hu, S.; Zhao, J.; Wei, Y.; Wang, L.; Shen, G.; Chen, Q. Microbial ecology of Fusarium wilt infected and healthy cucumber plant in root zone of continuous cropping soil. J. Northwest A F Univ. (Nat. Sci. Ed.) 2010, 38, 143–150. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16s rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2015, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Qin, Y.; Chen, T.; Lu, M.; Qian, X.; Guo, X.; Bai, Y. A practical guide to amplicon and metagenomic analysis of microbiome data. Protein Cell 2020, 1–16. [Google Scholar] [CrossRef]
- Xue, C.; Penton, C.; Zhu, C.; Chen, H.; Duan, Y.; Peng, C.; Guo, S.; Ling, N.; Shen, Q. Alterations in soil fungal community composition and network assemblage structure by different long-term fertilization regimes are correlated to the soil ionome. Biol. Fertil. Soils 2018, 54, 95–106. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Sivasithamparam, K. Non-streptomycete actinomycetes as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters. Soil Biol. Biochem. 2006, 38, 1505–1520. [Google Scholar] [CrossRef]
- Nguyen, N.; Song, Z.; Bates, S.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.; Kennedy, P. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Zhou, Y.; Xue, Q.; Yang, B.; Zhang, X.; Xu, Y.; Guo, Z.; Lin, C. Adjusted effect of inoculating with biocontrol Actinomyces on microbial flora of watermelon rooting zone. J. Northwest A F Univ. (Nat. Sci. Ed.) 2008, 36, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Debruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Appl. Environ. Micro. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Ka, J.O.; Song, H.G. Growth promotion of Xanthium italicum by application of rhizobacterial isolates of Bacillus aryabhattai in microcosm soil. J. Microbiol. 2012, 50, 45–49. [Google Scholar] [CrossRef]
- Chen, J.T.; Lin, M.J.; Huang, J.W. Efficacy of spent blewit mushroom compost and Bacillus aryabhattai combination on con-trol of Pythium damping-off in cucumber. J. Agric. Sci. 2015, 153, 1257–1266. [Google Scholar] [CrossRef]
- Zhang, H.; Xie, X.; Kim, M.; Kornyeyev, D.; Holaday, S.; Pare, P.W. Soil bacteria augment Arabidopsis photosynthesis by decreasing glucose sensing and abscisic acid levels in planta. Plant J. 2008, 56, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ji, Y.; Liu, Y.; Wang, H.; Zhao, H.; Niu, Y. Control of tree peony root rot using 6 species of biocontrol fungi and bac-teria. Shandong For. Sci. Technol. 2004, 6, 39–40. [Google Scholar] [CrossRef]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Wei, H.X.; Paré, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Penton, C.R.; Ruan, Y.; Shen, Z.; Xue, C.; Li, R.; Shen, Q. Inducing the rhizosphere microbiome by biofertilizer appli-cation to suppress banana Fusarium wilt disease. Soil Biol. Biochem. 2017, 104, 39–48. [Google Scholar] [CrossRef]
- Gloor, G.B.; Macklaim, J.M.; Vera, P.G.; Egozcue, J.J. Microbiome datasets are compositional: And this is not optional. Front. Microbiol. 2017, 8, 2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galloway-Peña, J.; Guindani, M. Editorial: Novel approaches in microbiome analyses and data visualization. Front. Microbiol. 2018, 9, 2274. [Google Scholar] [CrossRef]
- Araujo, R.; Dunlap, C.; Barnett, S.; Franco, C.M.M. Decoding wheat endosphere–rhizosphere microbiomes in Rhizoctonia solani–infested soils challenged by Streptomyces biocontrol agents. Front. Plant Sci. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wei, Z.; Wang, J.; Jousset, A.; Friman, V.-P.; Xu, Y.; Shen, Q.; Pommier, T. Facilitation promotes invasions in plant-associated microbial communities. Ecol. Lett. 2019, 22, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtz, Z.D.; Müller, C.L.; Miraldi, E.R.; Littman, D.R.; Blaser, M.J.; Bonneau, R.A. Sparse and Compositionally Robust Inference of Microbial Ecological Networks. PLoS Comput. Biol. 2015, 11, e1004226. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment | Fresh Shoot Weight | Fresh Fruit Weight | Dry Fruit Weight |
|---|---|---|---|---|
| Mean | Mean | Mean | ||
| High | Control | 87.15 ± 5.20 a | 149.44 ± 2.96 b | 5.79 ± 0.56 b |
| With Streptomyces | 93.42 ± 9.51 a | 237.03 ± 16.52 a | 9.57 ± 1.46 a | |
| Middle | Control | 101.97 ± 4.51 a | 56.26 ± 39.43 a | 2.62 ± 1.49 a |
| With Streptomyces | 82.62 ± 17.48 a | 100.02 ± 8.84 a | 4.19 ± 0.77 a | |
| Low | Control | 78.78 ± 3.30 a | 0 | 0 |
| With Streptomyces | 79.05 ± 14.47 a | 69.23 ± 19.15 | 3.16 ± 0.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, T.; Xue, Z.; Liu, Y.; Li, Y.; Li, Y.; Chen, Q. Streptomyces Application Triggers Reassembly and Optimization of the Rhizosphere Microbiome of Cucumber. Diversity 2021, 13, 413. https://doi.org/10.3390/d13090413
Zhang Y, Zhang T, Xue Z, Liu Y, Li Y, Li Y, Chen Q. Streptomyces Application Triggers Reassembly and Optimization of the Rhizosphere Microbiome of Cucumber. Diversity. 2021; 13(9):413. https://doi.org/10.3390/d13090413
Chicago/Turabian StyleZhang, Yanjiang, Tian Zhang, Zhijing Xue, Yifan Liu, Yongzhe Li, Yulong Li, and Qin Chen. 2021. "Streptomyces Application Triggers Reassembly and Optimization of the Rhizosphere Microbiome of Cucumber" Diversity 13, no. 9: 413. https://doi.org/10.3390/d13090413
APA StyleZhang, Y., Zhang, T., Xue, Z., Liu, Y., Li, Y., Li, Y., & Chen, Q. (2021). Streptomyces Application Triggers Reassembly and Optimization of the Rhizosphere Microbiome of Cucumber. Diversity, 13(9), 413. https://doi.org/10.3390/d13090413