
MtDNA D-Loop Diversity in Alpine Cattle during the Bronze Age

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
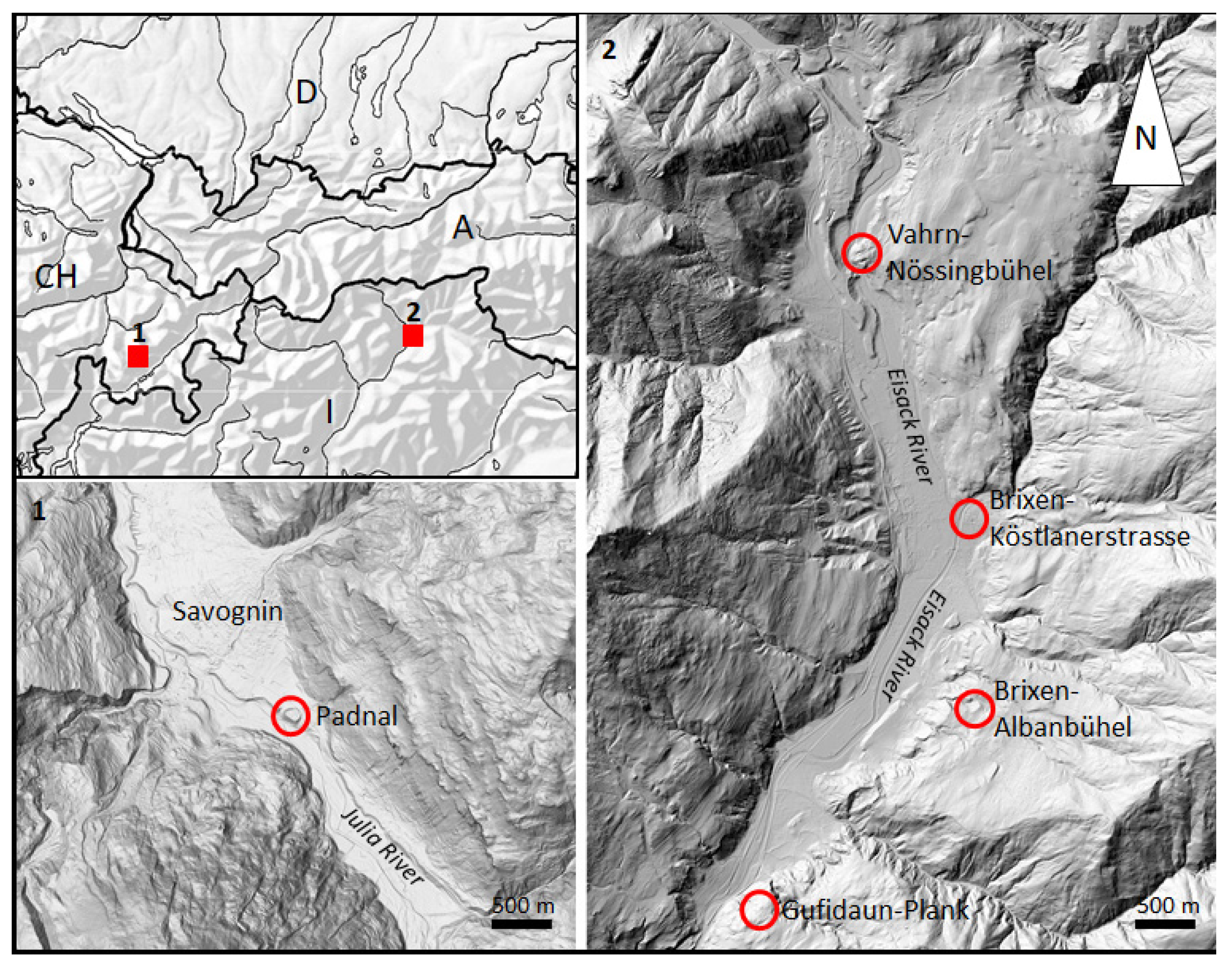
2.1. Samples for Archaeogenetic Analyses
2.2. The Archaeological Sites in Switzerland and Northern Italy
2.2.1. Northern Italy, Brixen Albanbühel 46°37′20″ N 11°53′40″ E
2.2.2. Northern Italy, Brixen Köstlanerstrasse 46°42′51″ N 11°39′48″ E
2.2.3. Northern Italy, Vahrn Nössingbühel 46°45′0.94″ N, 11°38′38.12″ E
2.2.4. Northern Italy, Klausen Gufidaun-Plank: 46°38′54.65″ N, 11°35′55.46″ E
2.2.5. Switzerland, Savognin Padnal 46 35′51.98″ N, 9°35′59.97″ E
2.3. DNA Extraction, PCR, and Sequencing
2.4. Authenticity
2.5. Statistical Analyses
3. Results
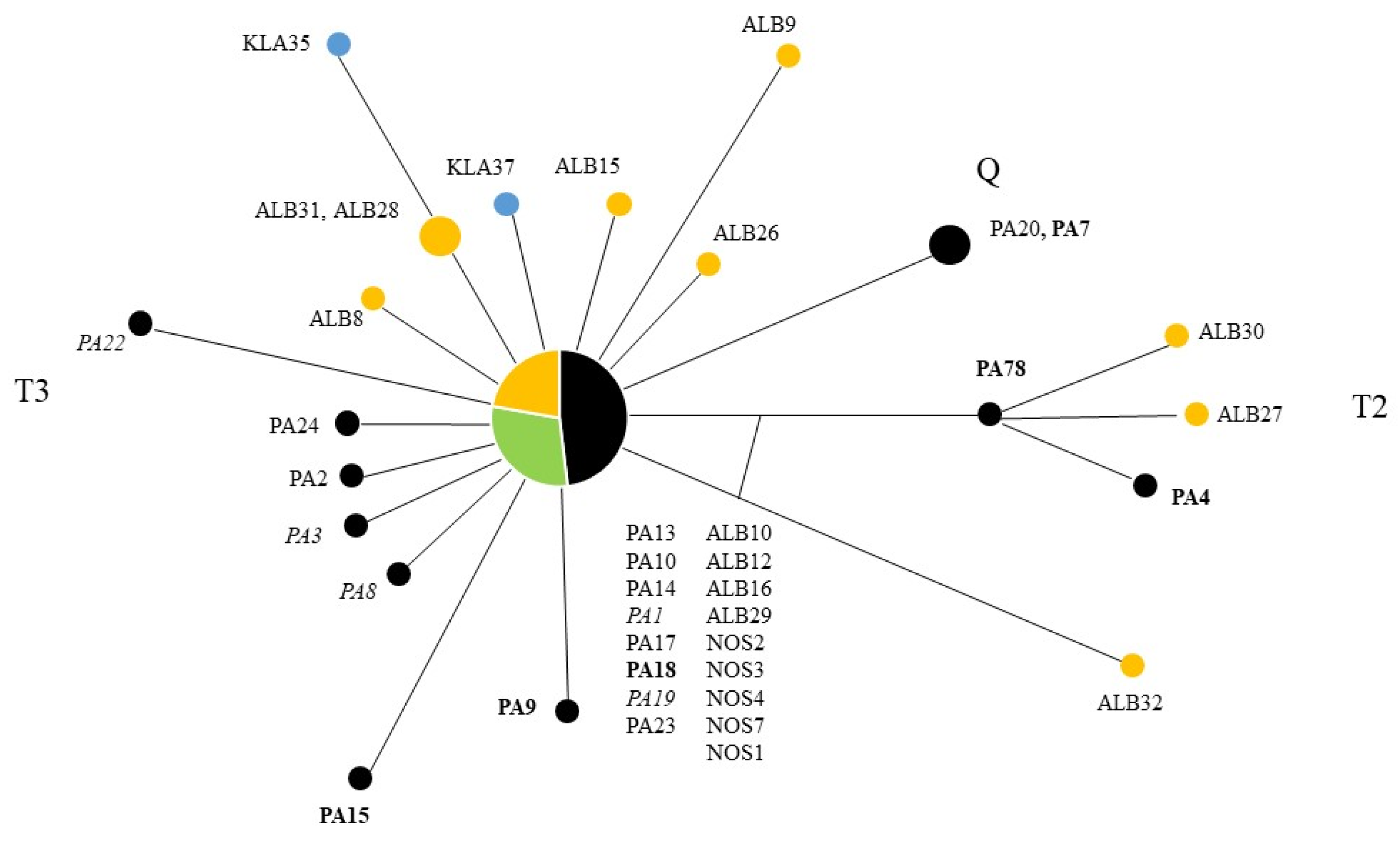
3.1. mtDNA D-Loop Haplotypes, Genetic Diversity, and Relationship between Haplotypes
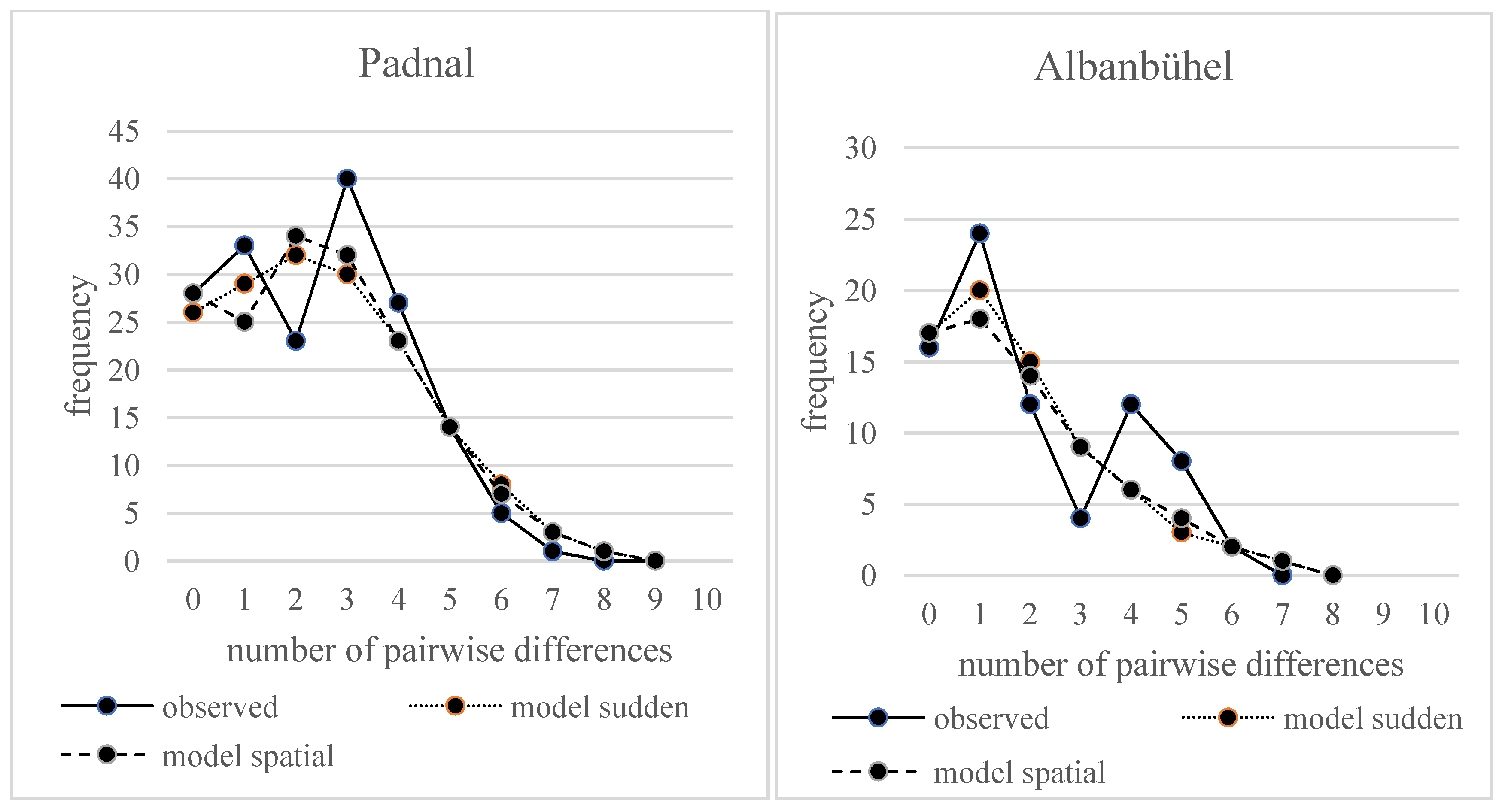
3.2. Population Demographic Estimates
3.3. Chronological Changes in Diversity in Cattle from Savognin Padnal
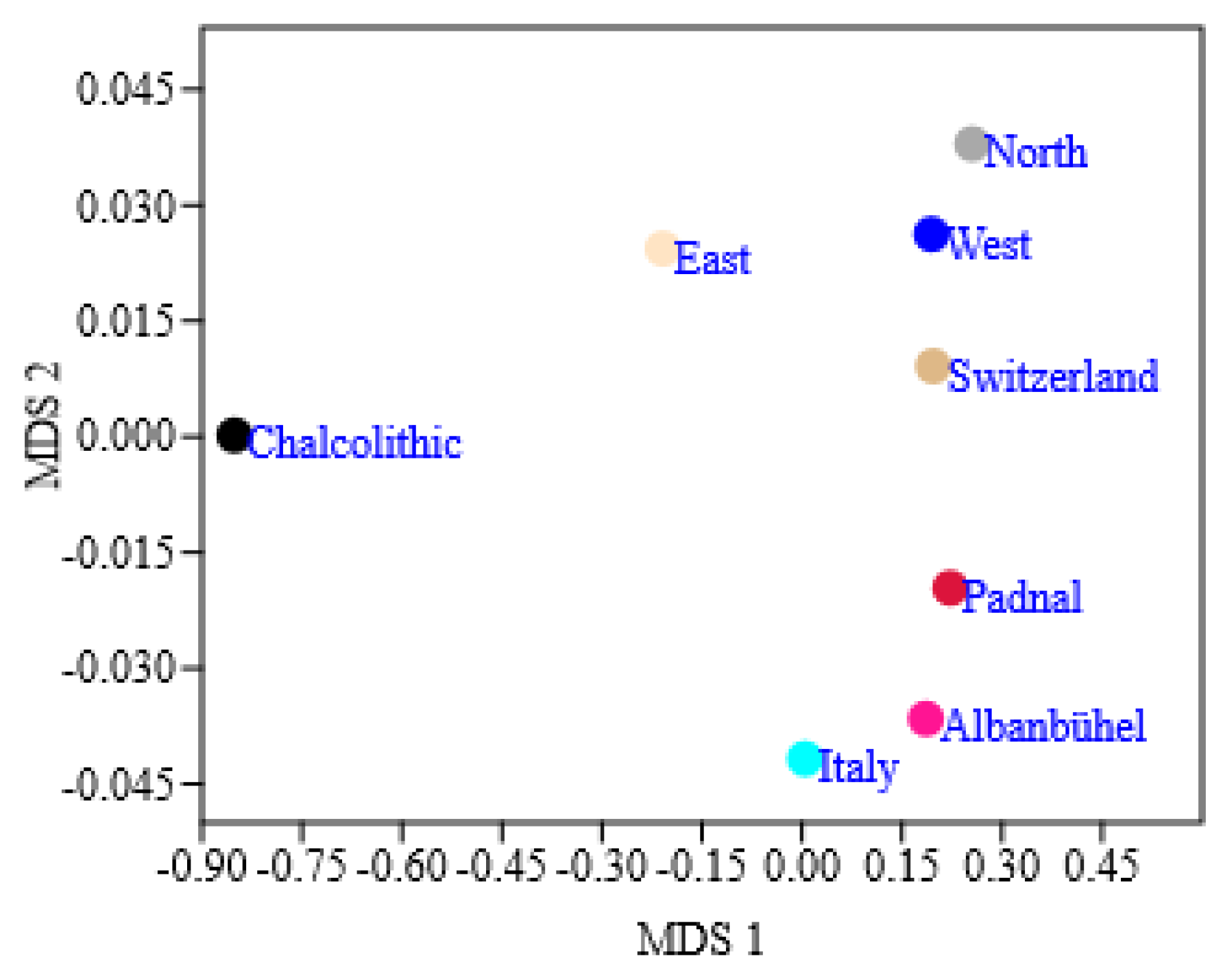
3.4. Bronze Age Cattle Populations in Relation to Modern Cattle Breeds
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fokkens, H.; Harding, A. (Eds.) The Oxford Handbook of the European Bronze Age; Oxford University Press: Oxford, UK, 2013. [Google Scholar] [CrossRef]
- Varalli, A.; Desideri, J.; David-Elbiali, M.; Goude, G.; Honegger, M.; Besse, M. Bronze Age innovations and impact on human diet: A multi-isotopic and multi-proxy study of western Switzerland. PLoS ONE 2021, 16, e0245726. [Google Scholar] [CrossRef] [PubMed]
- Allentoft, M.; Sikora, M.; Sjögren, K.-G.; Rasmussen, S.; Rasmussen, M.; Stenderup, J.; Damgaard, P.D.B.; Schroeder, H.; Ahlström, T.; Vinner, L.; et al. Population genomics of Bronze Age Eurasia. Nat. Cell Biol. 2015, 522, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Jennings, B.R. Exploring Late Bronze Age systems of bronzework production in Switzerland through Network Science. STAR Sci. Technol. Archaeol. Res. 2016, 2, 90–112. [Google Scholar] [CrossRef] [Green Version]
- Stika, H.-P.; Heiss, A. Plant Cultivation in the Bronze Age. In Oxford Handbook of the European Bronze Age; Fokkens, H., Harding, A., Eds.; Oxford University Press: Oxford, UK, 2013. [Google Scholar] [CrossRef]
- Bartosiewicz, L. Animals in Bronze Age Europe. In Oxford Handbook of European Bronze Age; Fokkens, H., Harding, A., Eds.; Oxford University Press: Oxford, UK, 2013. [Google Scholar] [CrossRef]
- Festi, D.; Putzer, A.; Oeggl, K. Mid and late Holocene land-use changes in the Ötztal Alps, territory of the Neolithic Iceman “Ötzi”. Quat. Int. 2014, 353, 17–33. [Google Scholar] [CrossRef]
- Reitmaier, T.; Doppler, T.; Pike, A.; Deschler-Erb, S.; Hajdas, I.; Walser, C.; Gerling, C. Alpine cattle management during the Bronze Age at Ramosch-Mottata, Switzerland. Quat. Int. 2018, 484, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Salvagno, L.; Tecchiati, U. I resti faunistici del villaggio dell’età del Bronzo di Sotćiastel. Economia e vita di una comunità protostorica alpina (ca. XVII-XIV sec. a.C.). Ladinia Monografica 2011, 3. [Google Scholar]
- Gilck, F.; Poschlod, P. The origin of alpine farming: A review of archaeological, linguistic and archaeobotanical studies in the Alps. Holocene 2019, 29, 1503–1511. [Google Scholar] [CrossRef]
- Bortenschlager, S. The Iceman’s environment. In The Iceman and His Natural Environment. Paleobotanical Results; Bortenschlager, S., Oeggl, K., Eds.; Springer: Vienna, Austria, 2000; Volume 4, pp. 11–27. [Google Scholar]
- Hafner, A.; Schwörer, C. Vertical mobility around the high-alpine Schnidejoch Pass. Indications of Neolithic and Bronze Age pastoralism in the Swiss Alps from paleoecological and archaeological sources. Quat. Int. 2018, 484, 3–18. [Google Scholar] [CrossRef]
- Stopp, B. Animal husbandry and hunting activities in the Late Bronze Age Circum-Alpine region. In The End of the Lake-Dwellings in the Circum-Alpine Region; Menotti, F., Ed.; Oxbow Books: Oxford, UK, 2015; pp. 179–210. [Google Scholar]
- Bopp-Ito, M.; Deschler-Erb, S.; Vach, W.; Schibler, J. Size diversity in Swiss Bronze Age cattle. Int. J. Osteoarchaeol. 2018, 28, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Pucher, E. Wie kam der Auerochse auf die Alm? Forschungsberichte der ANISA für das Internet. 2019, Volume 3, pp. 4–33. Available online: www.anisa.at (accessed on 5 January 2019).
- Putzer, A.; Festi, D.; Oeggl, K. Was the Iceman really a herdsman? The development of a prehistoric pastoral economy in the Schnals Valley. Antiquity 2016, 90, 319–336. [Google Scholar] [CrossRef]
- Pearce, M. Hard cheese: Upland pastoralism in the Italian Bronze and Iron Ages. In Summer Farms: Seasonal Exploitation of the Uplands from Prehistory to the Present; Pearce, J.R.C.M., Nicolis, F., Eds.; JR Collis Publications: Nottingham, UK, 2016; Volume 16, pp. 47–56. [Google Scholar]
- Riedel, A.; Tecchiati, U. Settlements and economy in the Bronze and Iron Age in Trentino-South Tyrol: Notes for an archaeozoological model. Preist. Alp. 2001, 35, 105–113. [Google Scholar]
- Bopp-Ito, M. Animal husbandry in the Bronze Age Alpine settlement “Savognin—Padnal”, Switzerland: A preliminary study. In Proceedings of the General Session of the 11th International Council for Archaeozoology Conference, Paris, France, 23–28 August 2010; Lefèvre, C., Ed.; BAR International Series. BAR Publishing: Oxford, UK, 2012; Volume 2354, pp. 75–85. [Google Scholar]
- Plüss, P. Die Bronzezeitliche Siedlung Cresta bei Cazis (GR): Die Tierknochen; Chronos Verlag: Zürich, Switzerland, 2011; Volume 9. [Google Scholar]
- Schibler, J.; Breitenlechner, E.; Deschler-Erb, S.; Goldenberg, G.; Hanke, K.; Hiebel, G.; Plogmann, H.H.; Nicolussi, K.; Marti-Graedel, E.; Pichler, S.L.; et al. Miners and mining in the Late Bronze Age: A multidisciplinary study from Austria. Antiquity 2011, 85, 1259–1278. [Google Scholar] [CrossRef] [Green Version]
- Itan, Y.; Powell, A.; Beaumont, M.; Burger, J.; Thomas, M.G. The Origins of Lactase Persistence in Europe. PLoS Comput. Biol. 2009, 5, e1000491. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, P.; Lancioni, H.; Ceccobelli, S.; Curcio, L.; Panella, F.; Lasagna, E. Uniparental genetic systems: A male and a female perspective in the domestic cattle origin and evolution. Electron. J. Biotechnol. 2016, 23, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Larson, G.; Albarella, U.; Dobney, K.; Rowley-Conwy, P.; Schibler, J.; Tresset, A.; Vigne, J.-D.; Edwards, C.J.; Schlumbaum, A.; Dinu, A.; et al. Ancient DNA, pig domestication, and the spread of the Neolithic into Europe. Proc. Natl. Acad. Sci. USA 2007, 104, 15276–15281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourier, T.; Ho, S.Y.; Gilbert, M.T.P.; Willerslev, E.; Orlando, L. Statistical Guidelines for Detecting Past Population Shifts Using Ancient DNA. Mol. Biol. Evol. 2012, 29, 2241–2251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheu, A.; Powell, A.; Bollongino, R.; Vigne, J.-D.; Tresset, A.; Çakırlar, C.; Benecke, N.; Bürger, J. The genetic prehistory of domesticated cattle from their origin to the spread across Europe. BMC Genet. 2015, 16, 54. [Google Scholar] [CrossRef] [Green Version]
- Hristov, P.; Spassov, N.; Iliev, N.; Radoslavov, G. An independent event of Neolithic cattle domestication on the South-eastern Balkans: Evidence from prehistoric aurochs and cattle populations. Mitochondrial DNA Part A 2015, 28, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Orton, D.; Gaastra, J.; Linden, M.V. Between the Danube and the Deep Blue Sea: Zooarchaeological Meta-Analysis Reveals Variability in the Spread and Development of Neolithic Farming across the Western Balkans. Open Quat. 2016, 2, 6. [Google Scholar] [CrossRef]
- Lenstra, J.A.; Ajmone-Marsan, P.; Beja-Pereira, A.; Bollongino, R.; Bradley, D.G.; Colli, L.; De Gaetano, A.; Edwards, C.J.; Felius, M.; Ferretti, L.; et al. Meta-Analysis of Mitochondrial DNA Reveals Several Population Bottlenecks during Worldwide Migrations of Cattle. Diversity 2014, 6, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Rageth, J. Die wichtigsten Resultate der Ausgrabung in der bronzezeitlichen Siedlung auf dem Padnal bei Savognin. Jahrbuch der Schweizerischen Gesellschaft für Ur-und Frühgeschichte 1986, 69, 63–103. [Google Scholar] [CrossRef]
- Parnigotto, I.; Pisoni, L.; Tecchiati, U. Nuovi dati e riflessioni sul Bronzo Finale nella conca di Bressanone (BZ): Risultati dello scavo di Via Castellano (Campagne 2002–2003). Studi in Onore di Renato Peroni Firenze All’Insegna del Giglio 2006, 17–29. [Google Scholar]
- Riedel, A.; Tecchiati, U. I resti faunistici dell’abitato d’altura dell’antica e media età del bronzo di Nössing in Val d’Isarco (Com. di Varna, Bolzano). Atti dell’Accademia Roveretana degli Agiati Classe di Scienze a 249 Ser. VII 1999, 9, 285–327. [Google Scholar]
- Tecchiati, U. Die Wallburg Nössing bei Vahrn (Brixen): Neue und alte Fragestellungen zur Frühbronzezeit Südtirols. In Jahresbericht des Institutes für Archäologien der Leopold-Franzens-Universität Innsbruck, Graben-Dokumentieren-Präsentieren 2015; Baur, C., Ed.; Institutes für Archäologien der Leopold-Franzens-Universität Innsbruck: Innsbruck, Austria, 2017; pp. 8–9. [Google Scholar]
- Angelini, I.; Artioli, G.; Pedrotti, A.; Tecchiati, U. La metallurgia dell’età del Rame dell’Italia settentrionale con particolare riferimento al Trentino Alto Adige: Le risorse minerarie e i processi di produzione del metallo. In L’età del Rame: La pianura padana e le Alpi al tempo di Ötzi; de Marinis, R.C., Ed.; La Compagnia Della Stampa Massetti Rodella: Brescia, Italy, 2013; pp. 101–116. [Google Scholar]
- Artioli, G.; Angelini, I.; Tecchiati, U.; Pedrotti, A. Eneolithic copper smelting slags in the Eastern Alps: Local patterns of metallurgical exploitation in the Copper Age. J. Archaeol. Sci. 2015, 63, 78–83. [Google Scholar] [CrossRef]
- Elsner, J.; Schibler, J.; Hofreiter, M.; Schlumbaum, A. Burial condition is the most important factor for mtDNA PCR amplification success in Palaeolithic equid remains from the Alpine foreland. Archaeol. Anthr. Sci. 2014, 7, 505–515. [Google Scholar] [CrossRef] [Green Version]
- Schibler, J.; Elsner, J.; Schlumbaum, A. Incorporation of aurochs into a cattle herd in Neolithic Europe: Single event or breeding? Sci. Rep. 2014, 4, 5798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binladen, J.; Thomas, M.; Gilbert, M.; Campos, P.; Willerslev, E. 5′-Tailed sequencing primers improve sequencing quality of PCR products. Biotechniques 2007, 42, 174–176. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.T.P.; Bandelt, H.-J.; Hofreiter, M.; Barnes, I. Assessing ancient DNA studies. Trends Ecol. Evol. 2005, 20, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, A.; Gandini, F.; Achilli, A.; Fichera, A.; Rizzi, E.; Bonfiglio, S.; Battaglia, V.; Brandini, S.; De Gaetano, A.; El-Beltagi, A.; et al. Mitogenomes from Egyptian Cattle Breeds: New Clues on the Origin of Haplogroup Q and the Early Spread of Bos taurus from the Near East. PLoS ONE 2015, 10, e0141170. [Google Scholar] [CrossRef]
- Achilli, A.; Bonfiglio, S.; Olivieri, A.; Malusà, A.; Pala, M.; Kashani, B.H.; Perego, U.A.; Ajmone-Marsan, P.; Liotta, L.; Semino, O.; et al. The Multifaceted Origin of Taurine Cattle Reflected by the Mitochondrial Genome. PLoS ONE 2009, 4, e5753. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.-J.; Macaulay, V.; Richards, M. Median networks: Speedy construction and greedy reduction, one simulation, and two case studies from human mtDNA. Mol. Phylogenet. Evol. 2000, 16, 8–28. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Beja-Pereira, A.; Caramelli, D.; Lalueza-Fox, C.; Vernesi, C.; Ferrand, N.; Casoli, A.; Goyache, F.; Royo, L.J.; Conti, S.; Lari, M.; et al. The origin of European cattle: Evidence from modern and ancient DNA. Proc. Natl. Acad. Sci. USA 2006, 103, 8113–8118. [Google Scholar] [CrossRef] [Green Version]
- Schlumbaum, A.; Turgay, M.; Schibler, J. Near East mtDNA haplotype variants in Roman cattle from Augusta Raurica, Switzerland, and in the Swiss Evolene breed. Anim. Genet. 2006, 37, 373–375. [Google Scholar] [CrossRef]
- Troy, C.S.; MacHugh, D.E.; Bailey, J.F.; Magee, D.A.; Loftus, R.T.; Cunningham, P.; Chamberlain, A.T.; Sykes, B.C.; Bradley, D.G. Genetic evidence for Near-Eastern origins of European cattle. Nat. Cell Biol. 2001, 410, 1088–1091. [Google Scholar] [CrossRef] [PubMed]
- Pellecchia, M.; Negrini, R.; Colli, L.; Patrini, M.; Milanesi, E.; Achilli, A.; Bertorelle, G.; Cavalli-Sforza, L.L.; Piazza, A.; Torroni, A.; et al. The mystery of Etruscan origins: Novel clues from Bos taurus mitochondrial DNA. Proc. R. Soc. Lond. B 2007, 274, 1175–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meiri, M.; Stockhammer, P.W.; Marom, N.; Bar-Oz, G.; Sapir-Hen, L.; Morgenstern, P.; Macheridis, S.; Rosen, B.; Huchon, D.; Maran, J.; et al. Eastern Mediterranean Mobility in the Bronze and Early Iron Ages: Inferences from Ancient DNA of Pigs and Cattle. Sci. Rep. 2017, 7, 701. [Google Scholar] [CrossRef] [Green Version]
- Verdugo, M.P.; Mullin, V.E.; Scheu, A.; Mattiangeli, V.; Daly, K.; Delser, P.M.; Hare, A.J.; Burger, J.; Collins, M.J.; Kehati, R.; et al. Ancient cattle genomics, origins, and rapid turnover in the Fertile Crescent. Science 2019, 365, 173–176. [Google Scholar] [PubMed]
- Ottoni, C.; Flink, L.G.; Evin, A.; Geörg, C.; De Cupere, B.; Van Neer, W.; Bartosiewicz, L.; Linderholm, A.; Barnett, R.; Peters, J.; et al. Pig Domestication and Human-Mediated Dispersal in Western Eurasia Revealed through Ancient DNA and Geometric Morphometrics. Mol. Biol. Evol. 2013, 30, 824–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furtwängler, A.; Rohrlach, A.B.; Lamnidis, T.C.; Papac, L.; Neumann, G.U.; Siebke, I.; Reiter, E.; Steuri, N.; Hald, J.; Denaire, A.; et al. Ancient genomes reveal social and genetic structure of Late Neolithic Switzerland. Nat. Commun. 2020, 11, 1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, W.; Fricke, H.; Halliday, A.N.; McCulloch, M.T.; Wartho, J.-A. Origin and Migration of the Alpine Iceman. Science 2003, 302, 862–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, G.; Wilson, I.; Payne, B.I.; Elson, J.; Samuels, D.C.; Santibanez-Korev, M.; Hall, S.J.; Chinnery, P.F. Unique mitochondrial DNA in highly inbred feral cattle. Mitochondrion 2012, 12, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Bopp-Ito, M. Archaeozoological Study on the Bronze Age Alpine Settlement Savognin-Padnal in the Canton of Grisons, Switzerland. Ph.D. Thesis, University of Basel, Basel, Switzerland, 2019. [Google Scholar]
- Bopp-Ito, M.; Cucchi, T.; Evin, A.; Stopp, B.; Schibler, J. Phenotypic diversity in Bronze Age pigs from the Alpine and Central Plateau regions of Switzerland. J. Archaeol. Sci. Rep. 2018, 21, 38–46. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code | Genbank Acc. No./Lab Code | Archaeological Code/Horizon | Site | Element | Dating | 14 C Lab Code | Dating | MtDNA Haplogroup | |
|---|---|---|---|---|---|---|---|---|---|
| cal BCE 2 sigma | typologically/cultural period | ||||||||
| 1 | NOS1 | MT423754.1 | 495 | Vahrn, Nössingbühel | M3 | 1949–1776 | MAMS-23501 | Early BA | T3 |
| 2 | NOS2 | MT423755.1 | 1146 | Vahrn, Nössingbühel | M3 | 1737–1541 | MAMS-23502 | Early BA | T3 |
| 3 | NOS3 | MT423756.1 | 1212 | Vahrn, Nössingbühel | M3 | 1875–1687 | MAMS-23503 | Early BA | T3 |
| 4 | NOS4 | MT423757.1 | 376 | Vahrn, Nössingbühel | M3 | 2111–1915 | MAMS-23504 | Early BA | T3 |
| 5 | NOS5 | OEG5 | 1044 | Vahrn, Nössingbühel | M3 | neg | Early BA | neg | |
| 6 | NOS6 | OEG6 | 407 | Vahrn, Nössingbühel | left jawbone with 3 molars | - | Early BA | neg | |
| 7 | NOS7 | MT423758.1 | 470 | Vahrn, Nössingbühel | jawbone with 3 molars | - | Early BA | T3 | |
| 8 | ALB8 | MT423741.1 | M3406 | Brixen, Albanbühel | maxillare | 1602–1438 | MAMS-23506 | T3 | |
| 9 | ALB9 | MT423742.1 | A9717 | Brixen, Albanbühel | maxillare | 1683–1531 | MAMS-23507 | T3 | |
| 10 | ALB10 | MT423767.1 | A3744 | Brixen, Albanbühel | molar | 1863–1642 | MAMS-23508 | T3 | |
| 11 | ALB11 | OEG11 | A5500 | Brixen, Albanbühel | molar | 1613–1502 | MAMS-23509 | neg | |
| 12 | ALB12 | MT423768.1 | A10130 | Brixen, Albanbühel | molar | 1496–1418 | MAMS-23510 | T3 | |
| 13 | ALB13 | OEG13 | A8568 | Brixen, Albanbühel | molar | 1681–1531 | MAMS-23511 | neg | |
| 14 | ALB14 | OEG14 | A7071 | Brixen, Albanbühel | molar | 1732–1533 | MAMS-23512 | neg | |
| 15 | ALB15 | MT423744.1 | 13056 | Brixen, Albanbühel | jawbone with 2 molars | 1518–1424 | MAMS-23513 | T3 | |
| 16 | ALB16 | MT423769.1 | 15278 | Brixen, Albanbühel | jawbone with 2 molars | 1260–1055 | MAMS-23514 | T3 | |
| 17 | ALB25 | OEG25 | 16419 | Brixen, Albanbühel | metatarsus distal | - | Early BA | neg | |
| 18 | ALB26 | MT423743.1 | 13511 | Brixen, Albanbühel | metatarsus proximal | - | Early BA | T3 | |
| 19 | ALB27 | MT423736.1 | 13684 | Brixen, Albanbühel | metarsus distal | - | Early BA | T2 | |
| 20 | ALB28 | MT423739.1 | 14596 | Brixen, Albanbühel | metacarpus proximal | - | Early BA | T3 | |
| 21 | ALB29 | MT423770.1 | 15722 | Brixen, Albanbühel | metatarsus proximal | - | Early BA | T3 | |
| 22 | ALB30 | MT423737.1 | 15729 | Brixen, Albanbühel | metatarsus distal | - | Early BA | T2 | |
| 23 | ALB31 | MT423738.1 | 13611 | Brixen, Albanbühel | metatarsus mitte | - | Early BA | T3 | |
| 24 | ALB32 | MT423745.1 | 17802 | Brixen, Albanbühel | metatarsus distal | - | Early BA | T3 | |
| 25 | BRI17 | OEG17 | 863 | Brixen, Köstlan | M3 | - | Late BA | neg | |
| 26 | BRI18 | OEG18 | 843 | Brixen, Köstlan | molar | - | Late BA | neg | |
| 27 | BRI19 | OEG19 | 841 | Brixen, Köstlan | molar | 1122–940 | MAMS-23517 | Late BA | neg |
| 28 | BRI20 | OEG20 | 375 | Brixen, Köstlan | molar | - | neg | ||
| 29 | BRI21 | OEG21 | 403 | Brixen, Köstlan | molar | neg | Late BA | neg | |
| 30 | BRI22 | OEG22 | 364 | Brixen, Köstlan | molar | - | Late BA | neg | |
| 31 | BRI23 | OEG23 | 384 | Brixen, Köstlan | molar | - | Late BA | neg | |
| 32 | BRI24 | OEG24 | 382 | Brixen, Köstlan | molar | - | Late BA | neg | |
| 33 | BRI33 | OEG33 | 363 | Brixen, Köstlan | molar | - | Late BA | neg | |
| 34 | KLA34 | OEG34 | 362+368 | Klausen, Gufidaun-Plank | radius proximal | - | Late Chalcolithic | neg | |
| 35 | KLA35 | MT423740.1 | 372 | Klausen, Gufidaun-Plank | tibia proximal | - | Late Chalcolithic | T3 | |
| 36 | KLA37 | MT423746.1 | 451 | Klausen, Gufidaun-Plank | tibia | - | Late Chalcolithic | T3 | |
| 37 | PA1 | MT423762.1 | 73–155 421/D | Savognin-Padnal | M3 | 1450–1350/1300 | Middle BA | T3 | |
| 38 | PA2 | MT423749.1 | 73–93 22/B | Savognin-Padnal | M3 | 1350/1300–900/800 | Late BA | T3 | |
| 39 | PA3 | MT423753.1 | 75–23b 411/D | Savognin-Padnal | M3 | 1450–1350/1300 | Middle BA | T3 | |
| 40 | PA4 | MT423732.1 | 76–9b 54/E | Savognin-Padnal | M3 | 1950/1900–1550/1450 | Early BA/ beginning Middle BA | T2 | |
| 41 | PA6 | MH6 | 72–150 201/B | Savognin-Padnal | M3 | 1350/1300–900/800 | Late BA | neg | |
| 42 | PA7 | MT423733.1 | SP76/126 51 143/E | Savognin-Padnal | M3 | 1950/1900–1550/1450 | Early BA/ beginning Middle BA | Q | |
| 43 | PA8 | MT423750.1 | 55/C | Savognin-Padnal | M3 | 1450–1350/1300 | Middle BA | T3 | |
| 44 | PA9 | MT423752.1 | SP76/189/E | Savognin-Padnal | M3 | 1950/1900–1550/1450 | Early BA/ beginning Middle BA | T3 | |
| 45 | PA10 | MT423760.1 | SP72/120–23/B | Savognin-Padnal | M3 | 1350/1300–900/800 | Late BA | T3 | |
| 46 | PA11 | MH11 | 75–27c 411/D | Savognin-Padnal | M3 | 1450–1350/1300 | Middle BA | neg | |
| 47 | PA12 | MH12 | 56/C | Savognin-Padnal | M3 | 1450–1350/1300 | Middle BA | neg | |
| 48 | PA13 | MT423760.1 | SP73/102–22/B | Savognin-Padnal | M3 | 1350/1300–900/800 | Late BA | T3 | |
| 49 | PA14 | MT423761.1 | SP73/96–201/B | Savognin-Padnal | M3 | 1350/1300–900/800 | Late BA | T3 | |
| 50 | PA15 | MT423747.1 | SP76/9(b) HZ-54/E | Savognin-Padnal | M3 | 1950/1900–1450 BC | Early BA/ beginning Middle BA | T3 | |
| 51 | PA16 | MH16 | 73–151 22/B | Savognin-Padnal | M3 | 1350/1300–900/800 | Late BA | neg | |
| 52 | PA17 | MT423763.1 | SP73/150–201/B | Savognin-Padnal | M3 | 1350/1300–900/800 | Late BA | T3 | |
| 53 | PA18 | MT423764.1 | 75–141b 52/E | Savognin-Padnal | M3 | 1950/1900–1550/1450 | Early BA/ beginning Middle BA | T3 | |
| 54 | PA19 | MT423765.1 | SP75/166–411/D | Savognin-Padnal | M3 | 1450–1350/1300 | Middle BA | T3 | |
| 55 | PA20 | MT423734.1 | SP73/166–202/B | Savognin-Padnal | M3 | 1350/1300–900/800 | Late BA | Q | |
| 56 | PA22 | MT423748.1 | 75–20b 401/D | Savognin-Padnal | M3 | 1450–1350/1300 | Middle BA | T3 | |
| 57 | PA23 | MT423766.1 | SP73–108 202.1 (black)/B | Savognin-Padnal | M3 | 1350/1300–900/800 | Late BA | T3 | |
| 58 | PA24 | MT423751.1 | SP73–108 202.2 (red)/B | Savognin-Padnal | M3 | 1350/1300–900/800 | Late BA | T3 | |
| 59 | PA78 | MT423735.1 | Savognin-Padnal | horncore sin | 1950/1900–1550/1450 | Early BA/ beginning Middle BA | T2 | ||
| 60 | PA178 | MT423735.1 | Savognin-Padnal | horncore dex | 1950/1900–1550/1450 | Early BA/ beginning Middle BA | T2 |
| Parameter | Albanbühel | Padnal | Nössingbühel |
|---|---|---|---|
| N | 13 | 19 | 5 |
| Htdiv | 0.85 ± 0.088 | 0.83 ± 0.086 | 0 |
| MNPD (π) | 2.05 ± 1.22 | 2.42 ± 1.37 | 0 |
| Tajima’s D | −1.45 | −2.01 | nd |
| Tajima’s D p-value | 0.07 | 0.01 * | nd |
| Fu’s Fs | −2.21 | −6.87 | nd |
| Fu’s Fs p-value | 0.057 | 0.0 * | nd |
| Sudden expansion model: Harpending’s raggedness index | 0.064 | 0.29 | nd |
| Harpending’s raggedness index p-value | 0.49 | 0.8 | nd |
| Spatial expansion model: Harpending’s raggedness index | 0.06 | 0.029 | nd |
| Harpending’s raggedness index p-value | 0.69 | 0.84 | nd |
| Period | N | Htdiv | MNDP (π) |
|---|---|---|---|
| Early Bronze Age | 6 | 1 ± 0.009 | 4.33 ± 2.49 |
| Middle Bronze Age | 5 | 0.9 ± 0.16 | 1.6 ± 1.13 |
| Late Bronze Age | 8 | 0.64 ± 0.18 | 1.25 ± 0.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Granado, J.; Harmath, M.; Tecchiati, U.; Oeggl, K.; Schibler, J.; Schlumbaum, A. MtDNA D-Loop Diversity in Alpine Cattle during the Bronze Age. Diversity 2021, 13, 449. https://doi.org/10.3390/d13090449
Granado J, Harmath M, Tecchiati U, Oeggl K, Schibler J, Schlumbaum A. MtDNA D-Loop Diversity in Alpine Cattle during the Bronze Age. Diversity. 2021; 13(9):449. https://doi.org/10.3390/d13090449
Chicago/Turabian StyleGranado, José, Marianna Harmath, Umberto Tecchiati, Klaus Oeggl, Jörg Schibler, and Angela Schlumbaum. 2021. "MtDNA D-Loop Diversity in Alpine Cattle during the Bronze Age" Diversity 13, no. 9: 449. https://doi.org/10.3390/d13090449
APA StyleGranado, J., Harmath, M., Tecchiati, U., Oeggl, K., Schibler, J., & Schlumbaum, A. (2021). MtDNA D-Loop Diversity in Alpine Cattle during the Bronze Age. Diversity, 13(9), 449. https://doi.org/10.3390/d13090449