
Physiological and Biochemical Characterization of Isolated Bacteria from a Coccolithophore Chrysotila dentata (Prymnesiophyceae) Culture



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Collection and Culture Conditions
2.2. Isolation and Purification of Culturable Bacteria
2.3. Physiological and Biochemical Characterization
2.4. DNA Extraction
2.5. PCR Amplification
2.6. Construction of Phylogenetic Trees
3. Results
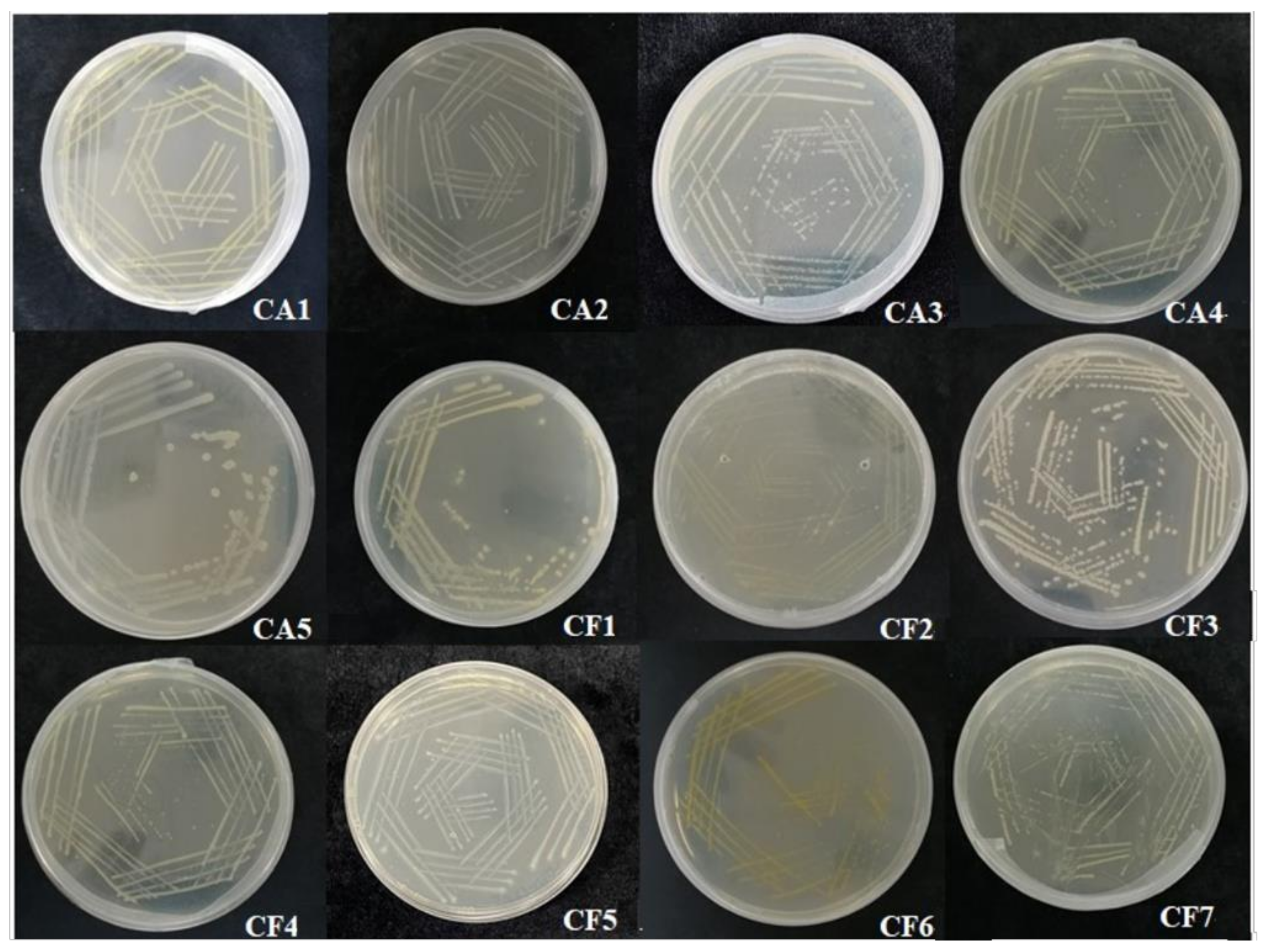
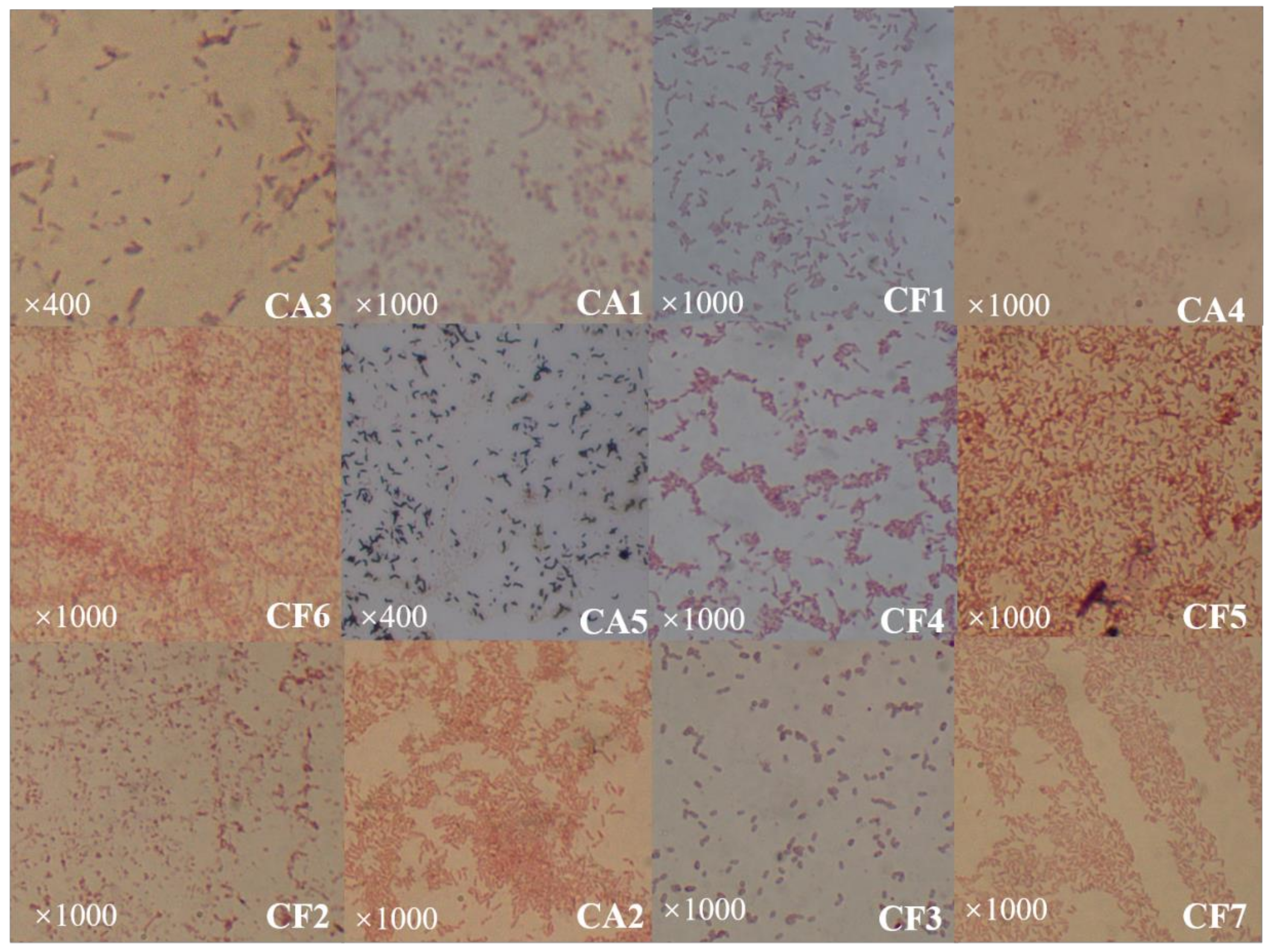
3.1. Identification of Culturable Phycosphere Bacteria
3.1.1. Isolation and Identification of Culturable Phycosphere Bacteria
3.1.2. Identification of Physiological and Biochemical Characteristics of Phycosphere Bacteria in C. dentata
3.2. Phylogenetic Tree of Culturable Phycosphere Bacteria from C. dentata
3.3. Phenotype Charateristics of Culturable Phycosphere Bacteria
3.4. Physiological and Biochemical Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, J.; Jin, S.F. Species diversity of living coccolithophores in Chinese sea waters. Biodivers. Sci. 2011, 19, 787–797. (In Chinese) [Google Scholar]
- Krumhardt, K.M.; Lovenduski, N.S.; Iglesias-Rodriguez, M.D.; Kleypas, J.A. Coccolithophores growth and calcification in a changing ocean. Prog. Oceanogr. 2017, 159, 276–295. [Google Scholar] [CrossRef]
- Weiner, S.; Dove, P.M. An Overview of Biomineralization Processes and the Problem of the Vital Effect. Rev. Mineral. Geochem. 2003, 54, 1–29. [Google Scholar] [CrossRef]
- Matoo, O.B.; Ivanina, A.V.; Ullstad, C.; Beniash, E.; Sokolova, I.M. Interactive effects of elevated temperature and CO2 levels on metabolism and oxidative stress in two common marine bivalves (Crassostrea virginica and Mercenaria mer-cenaria). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 164, 545–553. [Google Scholar] [CrossRef]
- Sun, J. Organic carbon pump and carbonate counter pump of living coccolithophorid. Adv. Earth Sci. 2007, 22, 23–31. [Google Scholar]
- Miao, Z.; Du, Z.J.; Li, H.R.; Lou, Y.Y.; Luo, W. Analysis of bacterial diversity in the phycosphere of five Arcticmi-croalgae. Acta Ecol. Sin. 2015, 35, 1587–1600. [Google Scholar]
- Bell, W.; Mitchell, R. Chemotactic and growth responses of marine bacteria to algal extracellular products. Biol. Bull. 1972, 143, 265–277. [Google Scholar] [CrossRef]
- Li, Y.; Liu, F.L.; Wang, Q.H.; Liu, Y.; Li, J.Y.; Ging, Q.L. Effects of cultivable phycosphere bacteria on growth of Haematococcus pluvialis and its astaxanthin accumulation. Period. Ocean Univ. China 2020, 50, 44–55. (In Chinese) [Google Scholar]
- Anesio, A.; Abreu, P.; Biddanda, B. The role of free and attached microorganisms in the decomposition of estuarine macrophyte detritus. Estuar. Coast. Shelf Sci. 2003, 56, 197–201. [Google Scholar] [CrossRef]
- Cao, X.; Li, H.; Zhou, Y.; Song, C. The shift of mutualistic relationships among algae, free-living and attached bacteria through different nutrient addition mode: A mesocosm study. J. Freshw. Ecol. 2020, 35, 535–548. [Google Scholar] [CrossRef]
- Fandino, L.; Riemann, L.; Steward, G.; Long, R.; Azam, F. Variations in bacterial community structure during a dinoflagellate bloom analyzed by DGGE and 16S rDNA sequencing. Aquat. Microb. Ecol. 2001, 23, 119–130. [Google Scholar] [CrossRef]
- 1Schäfer, H.; Abbas, B.; Witte, H.; Muyzer, G. Genetic diversity of ‘satellite’ bacteria present in cultures of marine diatoms. FEMS Microbiol. Ecol. 2002, 42, 25–35. [Google Scholar]
- Grossart, H.-P.; Levold, F.; Allgaier, M.; Simon, M.; Brinkhoff, T. Marine diatom species harbour distinct bacterial communities. Environ. Microbiol. 2005, 7, 860–873. [Google Scholar] [CrossRef] [PubMed]
- Rooney-Varga, J.; Giewat, M.; Savin, M.; Sood, S.; LeGresley, M.; Martin, J. Links between Phytoplankton and Bacterial Community Dynamics in a Coastal Marine Environment. Microb. Ecol. 2005, 49, 163–175. [Google Scholar] [CrossRef]
- Amin, S.A.; Green, D.H.; Hart, M.C.; Kupper, F.C.; Sunda, W.G.; Carrano, C.J. Photolysis of iron-siderophore chelates promotes bacterial-algal mutualism. Proc. Natl. Acad. Sci. USA 2009, 106, 17071–17076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, X.; Sun, J.; Satheeswaran, T.; Zhang, G.; Li, H.; An, X. Morphology, phylogenetic position, and ecophysiological features of the coccolithophore Chrysotila dentata (Prymnesiophyceae) isolated from the Bohai Sea, China. Phycologia 2019, 58, 628–639. [Google Scholar] [CrossRef]
- Green, D.H.; Llewellyn, L.E.; Negri, A.; Blackburn, S.I.; Bolch, C.J. Phylogenetic and functional diversity of the cultivable bacterial community associated with the paralytic shellfish poisoning dinoflagellate Gymnodinium catenatum. FEMS Microbiol. Ecol. 2004, 47, 345–357. [Google Scholar] [CrossRef]
- Mehta, A.; Bhardwaj, K.K.; Shaiza, M.; Gupta, R. Isolation, characterization and identification of pesticide de-grading bacteria from contaminated soil for bioremediation. Biol. Futur. 2021, 72, 317–323. [Google Scholar] [CrossRef]
- Bosshard, P.P.; Santini, Y.; Grüter, D.; Stettler, R.; Bachofen, R. Bacterial diversity and community composition in the chemocline of the meromictic alpine Lake Cadagno as revealed by 16S rDNA analysis. FEMS Microbiol. Ecol. 2000, 31, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishna, K.P.; Macadangdang, B.R.; Rogers, M.B.; Tometich, J.T.; Firek, B.A.; Baker, R.; Ji, J.; Burr, A.H.P.; Ma, C.; Good, M.; et al. Maternal IgA protects against the development of necrotizing enterocolitis in preterm infants. Nat. Med. 2019, 25, 1110–1115. [Google Scholar] [CrossRef]
- Hagström, Å.; Pinhassi, J.; Zweifel, U.L. Biogeographical diversity among marine bacterioplankton. Aquat. Microb. Ecol. 2000, 21, 231–244. [Google Scholar] [CrossRef]
- Wang, Y.; Coyne, K.J. Immobilization of algicidal bacterium Shewanella sp. IRI-160 and its application to control harmful dinoflagellates. Harmful Algae 2020, 94, 101798. [Google Scholar] [CrossRef] [PubMed]
- Eilers, H.; Pernthaler, J.; Peplies, J.; Glöckner, F.O.; Gerdts, G.; Amann, R. Isolation of Novel Pelagic Bacteria from the German Bight and Their Seasonal Contributions to Surface Picoplankton. Appl. Environ. Microbiol. 2001, 67, 5134–5142. [Google Scholar] [CrossRef] [Green Version]
- Agogué, H.; Casamayor, E.O.; Bourrain, M.; Obernosterer, I.; Joux, F.; Herndl, G.J.; LeBaron, P.; Agogué, H. A survey on bacteria inhabiting the sea surface microlayer of coastal ecosystems. FEMS Microbiol. Ecol. 2005, 54, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Kimbrel, J.A.; Samo, T.; Ward, C.; Nilson, D.; Thelen, M.P.; Siccardi, A.; Zimba, P.; Lane, T.W.; Mayali, X. Host selection and stochastic effects influence bacterial community assembly on the microalgal phycosphere. Algal Res. 2019, 40, 101489. [Google Scholar] [CrossRef]
- Huang, Y.-J.; Jiang, Z.-B.; Zeng, J.-N.; Chen, Q.-Z.; Zhao, Y.-Q.; Liao, Y.-B.; Shou, L.; Xu, X.-Q. The chronic effects of oil pollution on marine phytoplankton in a subtropical bay, China. Environ. Monit. Assess. 2011, 176, 517–530. [Google Scholar] [CrossRef]
- Leflaive, J.; Danger, M.; Lacroix, G.; Lyautey, E.; Oumarou, C.; Ten-Hage, L. Nutrient effects on the genetic and functional diversity of aquatic bacterial communities. FEMS Microbiol. Ecol. 2008, 66, 379–390. [Google Scholar] [CrossRef]
- Cho, D.H.; Ramanan, R.; Heo, J.; Lee, J.; Kim, B.H.; Oh, H.M.; Kim, H.S. Enhancing microalgal biomass produc-tivity by engineering a microalgal–bacterial community. Bioresour. Technol. 2015, 175, 578–585. [Google Scholar] [CrossRef]
- Le Chevanton, M.; Garnier, M.; Bougaran, G.; Schreiber, N.; Lukomska, E.; Bérard, J.-B.; Fouilland, E.; Bernard, O.; Cadoret, J.-P. Screening and selection of growth-promoting bacteria for Dunaliella cultures. Algal Res. 2013, 2, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Stevens, H.; Stübner, M.; Simon, M.; Brinkhoff, T. Phylogeny of Proteobacteria and Bacteroidetes from oxic habitats of a tidal flat ecosystem. FEMS Microbiol. Ecol. 2005, 54, 351–365. [Google Scholar] [CrossRef]
- Raven, J.A.; Giordano, M. Biomineralization by photosynthetic organisms: Evidence of coevolution of the organisms and their environment? Geobiology 2009, 7, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Middelboe, M.; Letarte, Y.; Borch, N.H. Attached and free-living bacteria: Production and polymer hydrolysis during a diatom bloom. Microb. Ecol. 1995, 29, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.J.; Sun, J.; Feng, Y.Y. Study on modern coccolithophores in coastal region along the east Hainan Island. Haiyang Xuebao 2015, 37, 27–40. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Samples | Accession | Closest Relative in Database | Accession Number | Similarity (%) |
|---|---|---|---|---|
| CFB group | ||||
| CF2 | SRR17041857 | Flavobacteriaceae bacterium LA8 | AF513435.1 | 99.57 |
| CF6 | SRR17041863 | Flavobacterium sp. Strain 7002-040 | KY770468.1 | 99.71 |
| CA1 | SRR17041865 | Flavobacterium sp. Strain 7002-040 | KY770468.1 | 99.64 |
| α-proteobacteria | ||||
| CA2 | SRR17041864 | Stappia sp. strain FG-4 | JF451107.1 | 100.00 |
| CF1 | SRR17041858 | Alpha proteobacterium 45A11 | JF263527.1 | 99.92 |
| CF4 | SRR17041855 | Thalassococcus sp. strain S24 | KX989355.1 | 100.00 |
| CF7 | SRR17041862 | Stappia sp. Strain VB | MH454495.1 | 100.00 |
| γ-proteobacteria | ||||
| CA4 | SRR17041860 | Alteromonas sp. strain KSTI56 | KX989422.1 | 100.00 |
| CA3 | SRR17041861 | Marinobacterhydrocarbonoclasticus strain RMR 21 | KP769865.1 | 99.46 |
| CF5 | SRR17041854 | Marinobacter alkaliphilus strain Ad 1 | MF029656.2 | 99.85 |
| firmicutes | ||||
| CF3 | SRR170418856 | Bacillus firmus strain sctcc471 | HQ622343.1 | 99.86 |
| CA5 | SRR17041859 | Bacillus xiamenensis strain SN3-8 | MT071697.1 | 99.93 |
| Strain Number | Color | Diameter (mm) | Shape | Elevation | Margin | Gram Nature |
|---|---|---|---|---|---|---|
| CA1 | Yellow | 0.7 | Circular | Flat | Entire | Negative |
| CA2 | White | 0.4 | Circular | Flat | Entire | Negative |
| CA3 | White | 0.4 | Circular | Flat | Entire | Negative |
| CA4 | Pale yellow | 0.7 | Circular | Flat | Entire | Negative |
| CA5 | White | 1.2 | Circular | Flat | Entire | Positive |
| CF1 | Pale yellow | 0.6 | Circular | Flat | Entire | Negative |
| CF2 | Bright yellow | 0.4 | Circular | Flat | Entire | Negative |
| CF3 | White | 0.7 | Circular | Flat | Entire | Positive |
| CF4 | Pale yellow | 0.25 | Circular | Flat | Entire | Negative |
| CF5 | Transparent | 0.35 | Circular | Bulge | Extension | Negative |
| CF6 | Bright yellow | 0.45 | Oval | Flat | Entire | Negative |
| CF7 | Pale yellow | 0.15 | Circular | Bulge | Entire | Negative |
| Target | Physiological and Biochemical Characteristics | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CA1 | CA2 | CA3 | CA4 | CA5 | CF1 | CF2 | CF3 | CF4 | CF5 | CF6 | CF7 | |
| Voges–Proskauer | − | − | − | − | − | − | − | − | − | − | − | − |
| Gelatin liquefaction | − | − | − | − | + | − | − | − | − | − | − | − |
| Urease test | − | + | - | − | − | + | − | − | − | − | − | + |
| Maltose | − | − | + | − | − | − | − | − | − | + | − | − |
| Glucose | − | − | − | − | + | − | − | − | − | − | − | − |
| Xylose | − | − | − | − | − | − | − | − | − | − | − | + |
| Arabinose | − | − | + | − | + | − | − | + | − | + | − | + |
| Mannitol | − | − | − | − | + | − | − | − | − | − | − | − |
| Starch hydrolysis | − | − | + | − | + | − | − | + | − | + | + | + |
| Malonate | + | + | − | + | − | + | + | + | + | + | + | + |
| Aesculin | − | − | + | − | + | − | − | + | − | + | + | + |
| Nitrate reduction | − | − | + | − | − | + | − | − | − | − | − | + |
| pH 5.7 | − | − | + | − | + | − | − | + | − | + | − | + |
| 7% NaCl | − | + | + | + | + | + | − | − | − | + | − | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Liu, Y.; Sun, J. Physiological and Biochemical Characterization of Isolated Bacteria from a Coccolithophore Chrysotila dentata (Prymnesiophyceae) Culture. Diversity 2022, 14, 2. https://doi.org/10.3390/d14010002
Wang X, Liu Y, Sun J. Physiological and Biochemical Characterization of Isolated Bacteria from a Coccolithophore Chrysotila dentata (Prymnesiophyceae) Culture. Diversity. 2022; 14(1):2. https://doi.org/10.3390/d14010002
Chicago/Turabian StyleWang, Xueru, Yang Liu, and Jun Sun. 2022. "Physiological and Biochemical Characterization of Isolated Bacteria from a Coccolithophore Chrysotila dentata (Prymnesiophyceae) Culture" Diversity 14, no. 1: 2. https://doi.org/10.3390/d14010002
APA StyleWang, X., Liu, Y., & Sun, J. (2022). Physiological and Biochemical Characterization of Isolated Bacteria from a Coccolithophore Chrysotila dentata (Prymnesiophyceae) Culture. Diversity, 14(1), 2. https://doi.org/10.3390/d14010002