
Effects of Terrestrial Inputs on Mesozooplankton Community Structure in Bohai Bay, China



Abstract
:1. Introduction
2. Materials and Methods
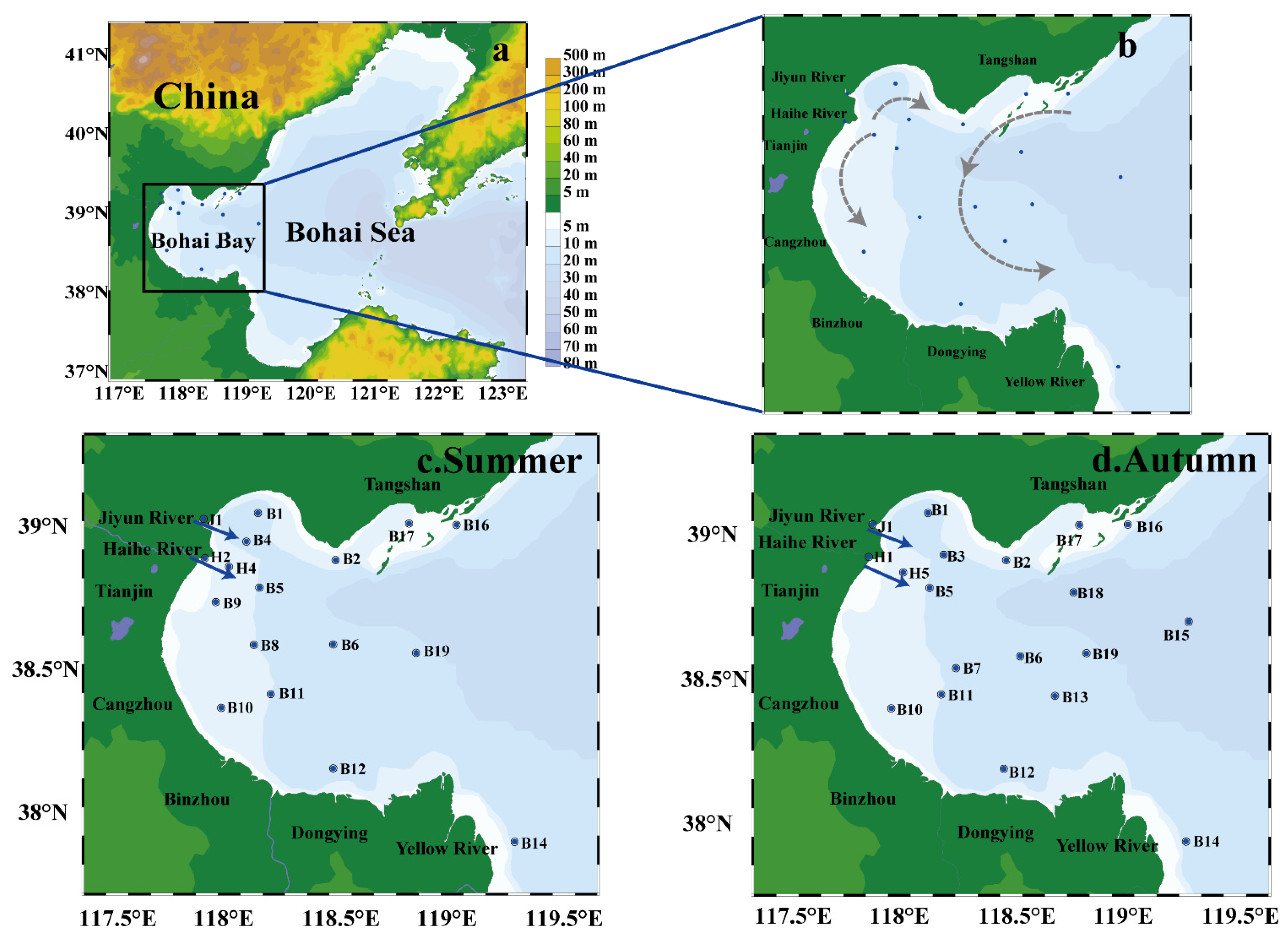
2.1. Study Area and Sampling Stations
2.2. Sample Collection and Analysis
2.3. Statistical Analysis
3. Results
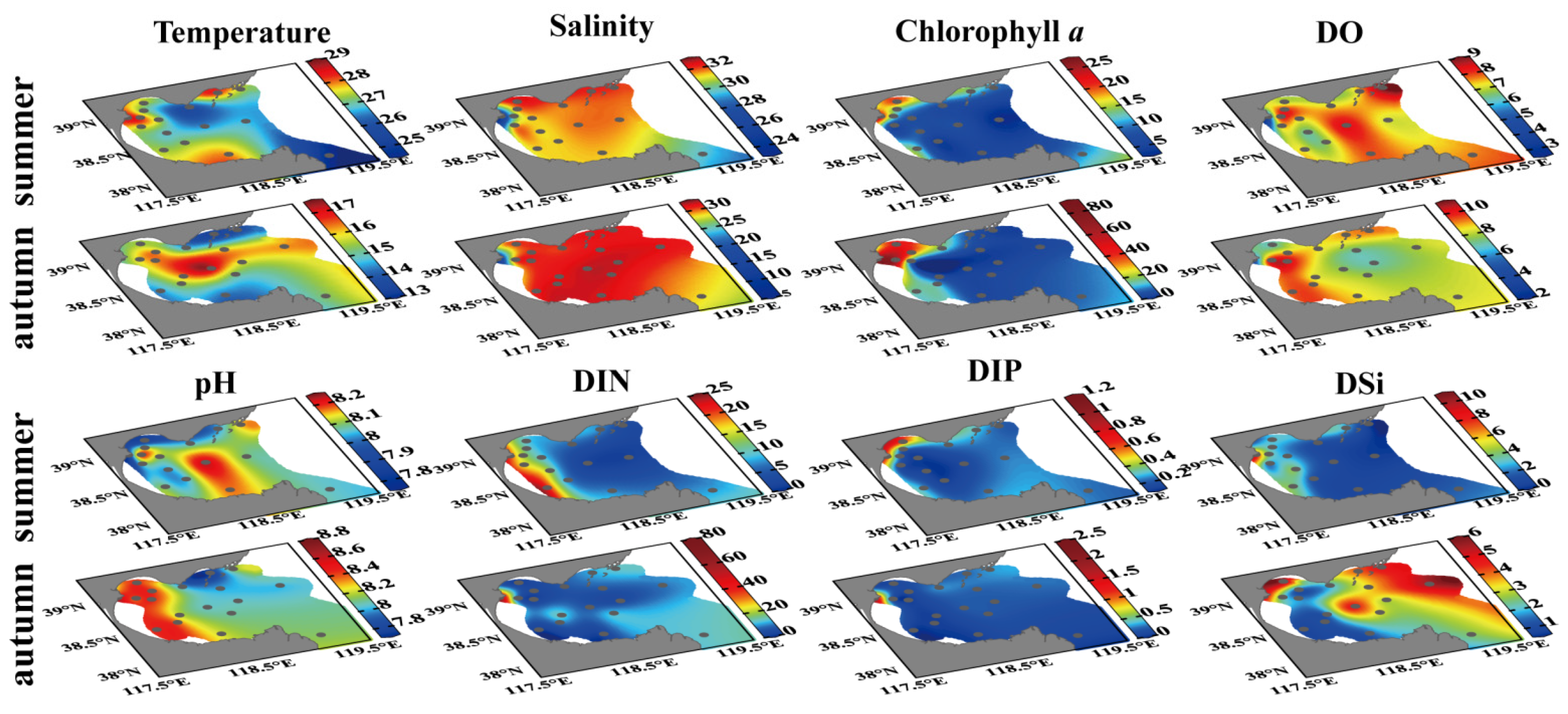
3.1. Environmental Variables
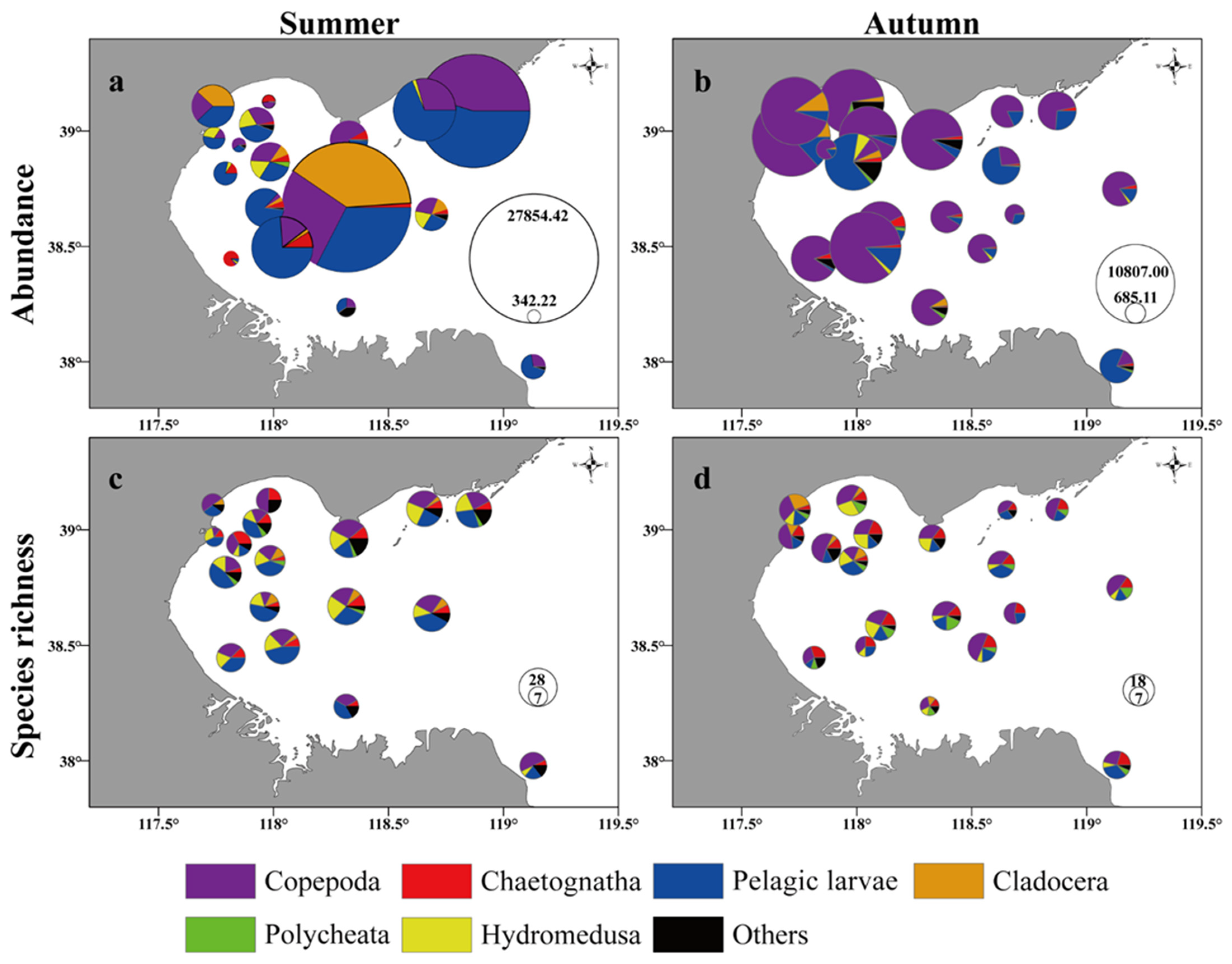
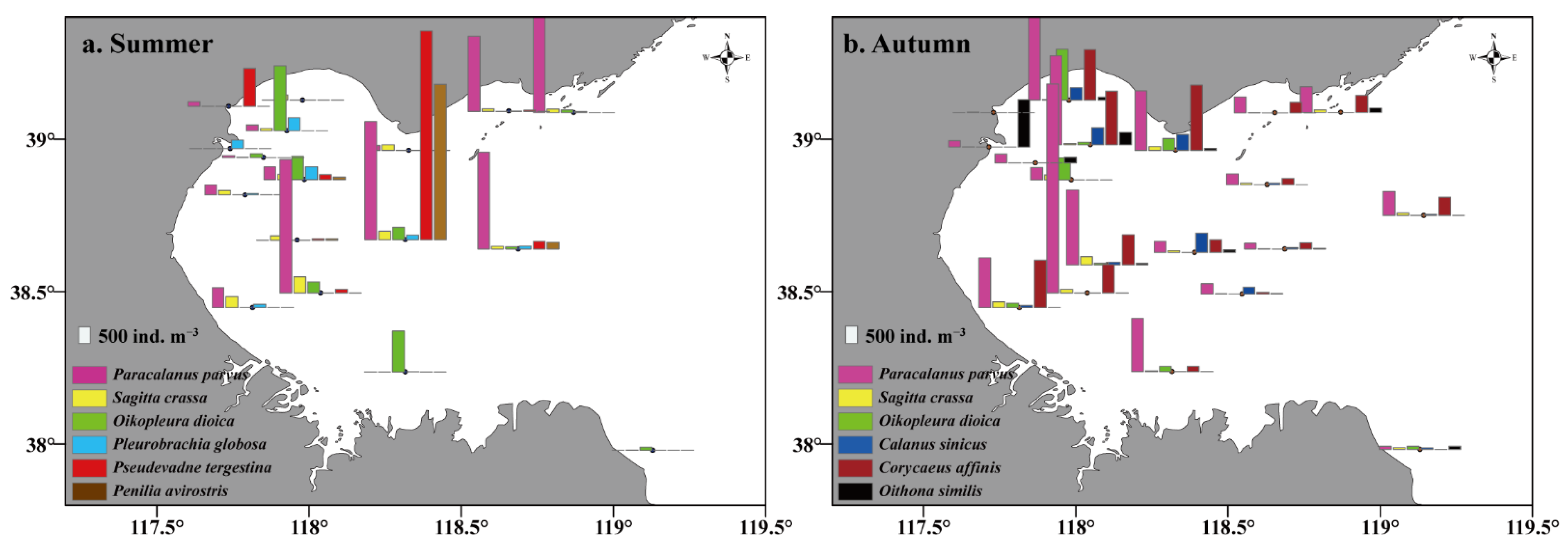
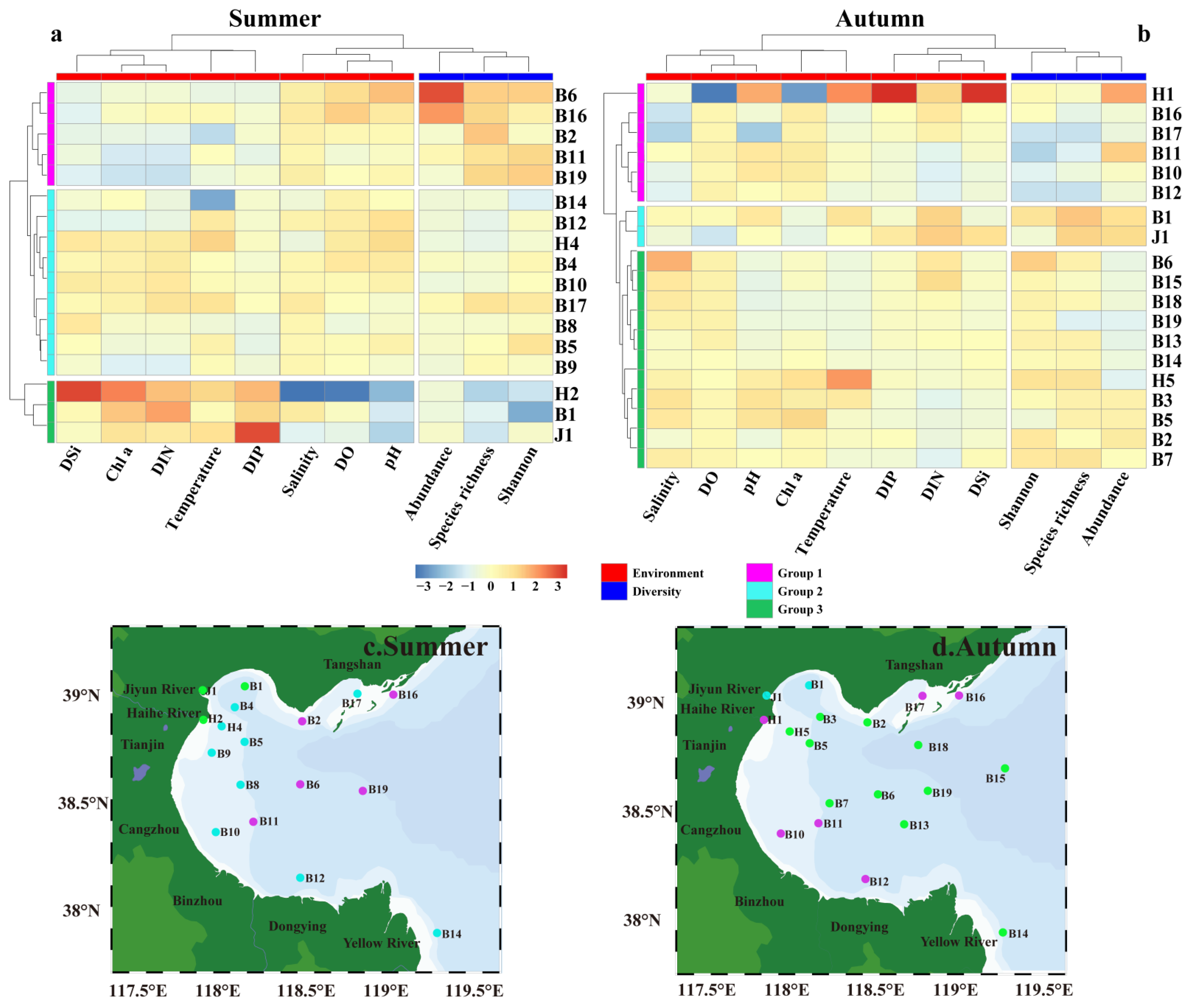
3.2. Mesozooplankton Composition and Abundance
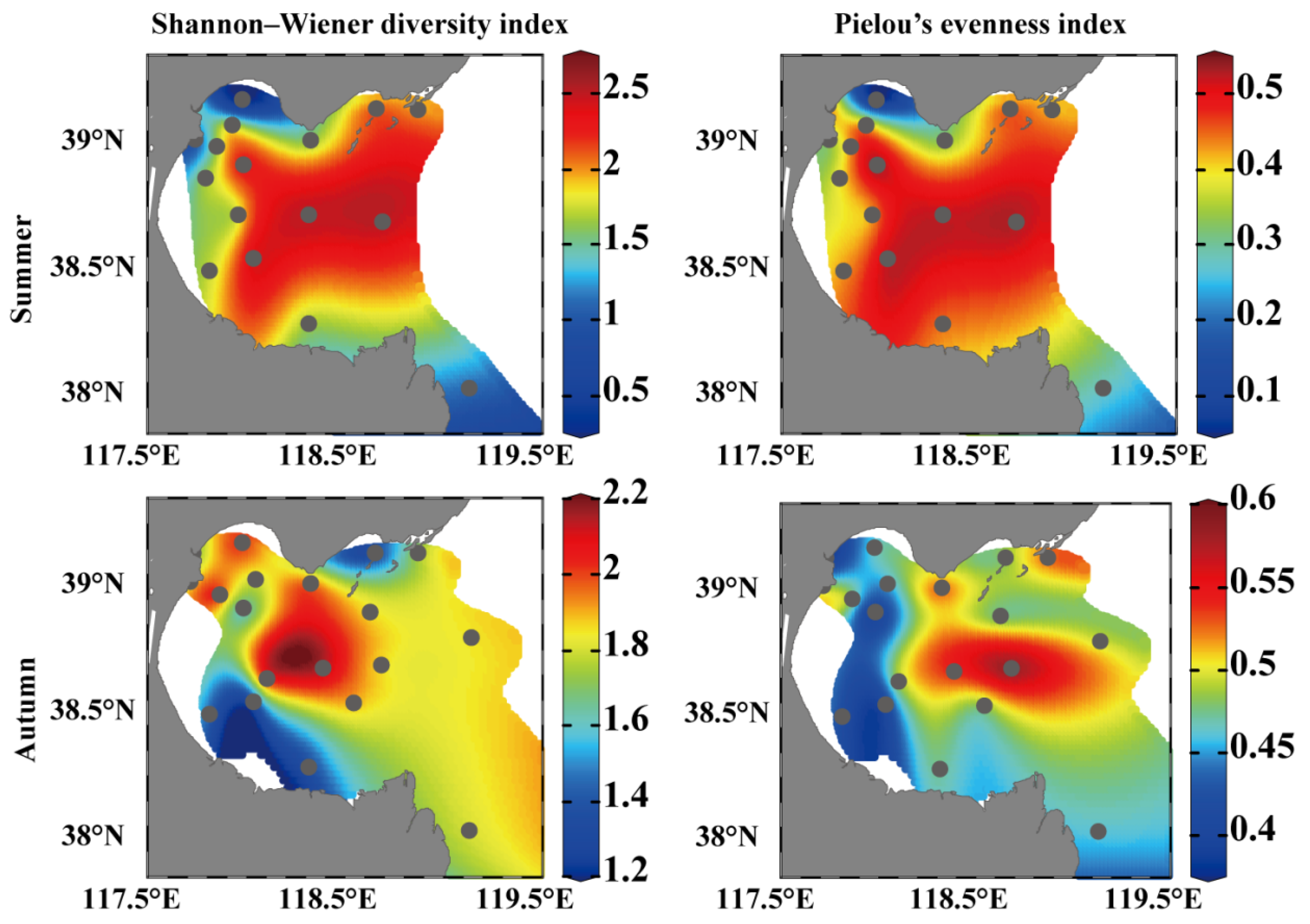
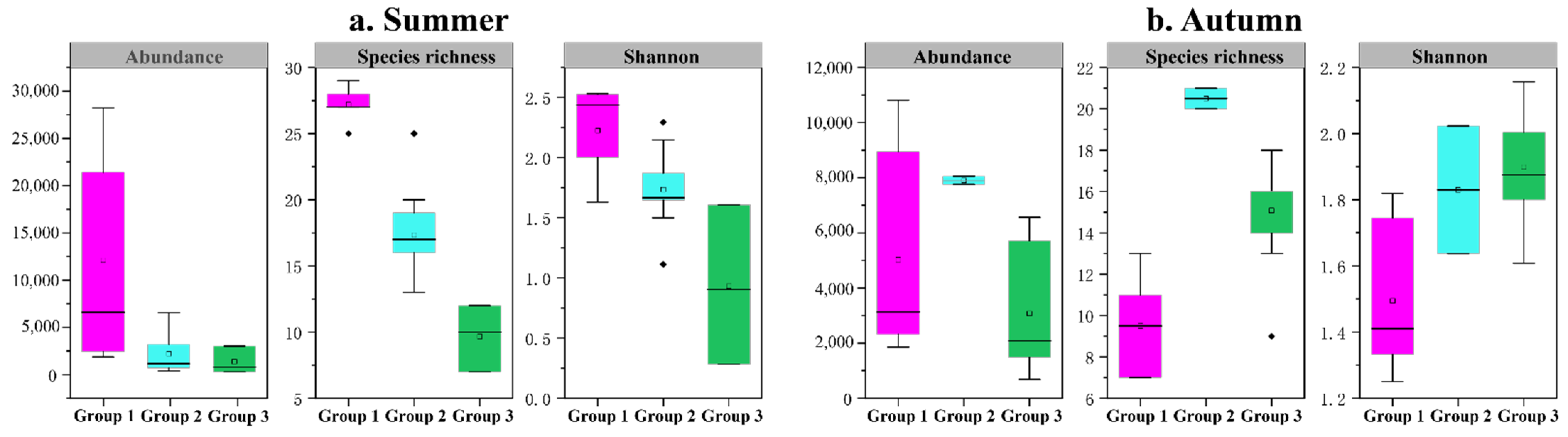
3.3. Diversity Indices of the Mesozooplankton Community
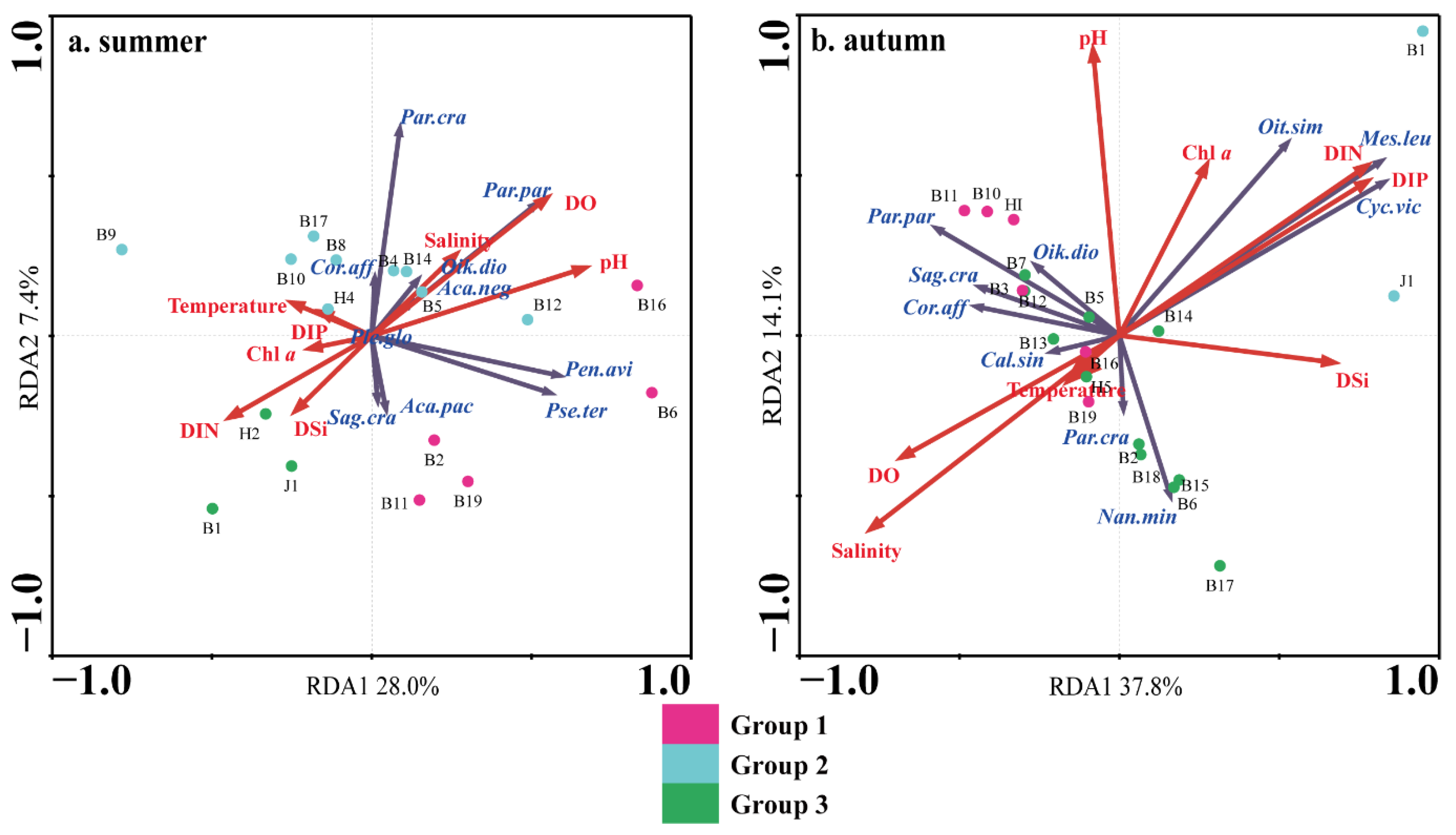
3.4. Relationship between Mesozooplankton Community and Environmental Factors
4. Discussion
4.1. Comparison with Historical Data
4.2. Comparison with Other Bay Ecosystems
4.3. Effects of Terrestrial Inputs on Mesozooplankton Communities
4.4. Effects of Seawater Intrusion on Mesozooplankton Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steinberg, D.; Landry, M. Zooplankton and the Ocean Carbon Cycle. Annu. Rev. Mar. Sci. 2017, 9, 413–444. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z. New progress in marine zooplankton research in China. J. Xiamen Univ. Nat. Sci. Ed. 2006, 45, 8. [Google Scholar]
- Hobday, A.J.; Okey, T.A.; Poloczanska, E.S.; Kunz, T.J.; Richardson, A.J. Impacts of Climate Change on Australian Marine Life, Part B: Technical Report. CSIRO Marine and Atmospheric Research; the Australian Greenhouse Office: Canberra, Australia, 2006. [Google Scholar]
- Ma, Y.; Ke, Z.; Huang, L.; Tan, Y. Identification of human-induced perturbations in Daya Bay, China: Evidence from plankton size structure. Cont. Shelf Res. 2014, 72, 10–20. [Google Scholar] [CrossRef]
- Simon, N.; Cras, A.-L.; Foulon, E.; Lemée, R. Diversity and evolution of marine phytoplankton. Comptes Rendus Biol. 2009, 332, 159–170. [Google Scholar] [CrossRef]
- Jumars, P.A. Biological oceanography: An introduction (C. M. Lalli and T. R. Parsons). Limnol. Oceanogr. 2003, 39, 982. [Google Scholar] [CrossRef]
- Park, G.S. Estuarine relationships between zooplankton community structure and trophic gradients. J. Plankton Res. 2000, 22, 121–136. [Google Scholar] [CrossRef]
- Ji, D.; Yang, J.; Gao, Z. Eutrophication Assessment of the Western Sea Area in the Laizhou Bay During the Low Water Period. Mar. Environ. Sci. 2007, 26, 81. [Google Scholar]
- Xiang, C.; Ke, Z.; Li, K.; Liu, J.; Zhou, L.; Lian, X.; Tan, Y. Effects of terrestrial inputs and seawater intrusion on zooplankton community structure in Daya Bay, South China Sea. Mar. Pollut. Bull. 2021, 167, 112331. [Google Scholar] [CrossRef]
- Jeyaraj, N.; Ravikumar, S.; Rajthilak, C.; Kumar, P.; Santhanam, P. Abundance and diversity of zooplankton in the Gulf of Mannar region on the southeastern coast of India. Acta Mar. Biol. 2016, 6. [Google Scholar] [CrossRef]
- Davor, L.; Hure, M.; Svjetlana, B. The effect of temperature change and oxygen reduction on zooplankton composition and vertical distribution in a semi-enclosed marine system. Mar. Biol. Res. 2019, 15, 325–342. [Google Scholar] [CrossRef]
- Zheng, C.; Li, S.; Lian, G. Biology of Marine Copepods; Xiamen University Press: Xiamen, China, 1992. [Google Scholar]
- Zheng, Z.; Cao, W. Studies on marine cladocerans in China—II. Distribution. Chin. J. Oceanogr. (Chin. Version) 1982, 81–92. [Google Scholar]
- Froneman, P. Zooplankton community structure and biomass in a southern African temporarily open/closed estuary. Estuar. Coast. Shelf Sci. 2004, 60, 125–132. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, H.; Liu, G. The characteristics and influencing factors of zooplankton community in Shacheng Port, Fujian. Journal of Ocean University of China. Nat. Sci. Ed. 2008, 38, 8. [Google Scholar] [CrossRef]
- Du, M.; Liu, Z.; Wang, C.; Dong, Z.; Jing, Z. Community structure and seasonal changes of zooplankton in offshore China. Chin. J. Ecol. 2013, 33, 12. [Google Scholar] [CrossRef]
- Huang, J.; Zhu, Y.; Wang, Y.; Wu, Y. Quantitative driving analysis on temporal and spatial variations of mesozooplankton community structures in Xiangshan Bay. Ecol. Sci. 2014, 33, 713–722. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Zheng, B.; Zhu, Y.; Wang, X. Analysis of the bacterial community in the two typical intertidal sediments of Bohai Bay, China by pyrosequencing. Mar. Pollut. Bull. 2013, 72, 181–187. [Google Scholar] [CrossRef]
- Liu, J.; Feng, Y.; Zhang, Y.; Liang, N.; Wu, H.; Liu, F. Allometric releases of nitrogen and phosphorus from sediments mediated by bacteria determines water eutrophication in coastal river basins of Bohai Bay. Ecotoxicol. Environ. Saf. 2022, 235, 113426. [Google Scholar] [CrossRef]
- Heisler, J.; Glibert, P.; Burkholder, J.; Anderson, D.; Cochlan, W.; Dennison, W.; Dortch, Q.; Gobler, C.; Heil, C.; Humphries, E.; et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2009, 8, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Rabalais, N.N.; Turner, R.E.; Díaz, R.J.; Justić, D. Global change and eutrophication of coastal waters. ICES J. Mar. Sci. 2009, 66, 1528–1537. [Google Scholar] [CrossRef]
- Song, W. Analysis of the Bohai Sea’s temperature and salt field structure in winter and summer and its current characteristics. Ocean. Univ. China 2009. [Google Scholar] [CrossRef]
- Xie, L.; Xu, H.; Xin, M.; Wang, B.; Tu, J.; Wei, Q.; Sun, X. Regime shifts in trophic status and regional nutrient criteria for the Bohai Bay, China. Mar. Pollut. Bull. 2021, 170, 112674. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, W.; Wang, R. Zooplankton community structure in spring and autumn in the central and southern Bohai Sea. Mar. Sci. Collect. 2002, 9. [Google Scholar]
- Parsons, T.R.; Yoshiaki, M.; Lalli, M.C. Artificial Seawater Media. A Man. Chem. Biol. Methods Seawater Anal. 1984, 158–161. [Google Scholar] [CrossRef]
- Brzezinski, M.A.; Nelson, D.M. A solvent extraction method for the colorimetric determination of nanomolar concentrations of silicic acid in seawater. Mar. Chem. 1986, 19, 139–151. [Google Scholar] [CrossRef]
- Karl, D.M.; Tien, G. MAGIC: A sensitive and precise method for measuring dissolved phosphorus in aquatic environments. Limnol. Oceanogr. 1992, 37, 105–116. [Google Scholar] [CrossRef]
- Cloern, J.E.; Abreu, P.; Carstensen, J.; Chauvaud, L.; Elmgren, R.; Grall, J.; Greening, H.; Johansson, J.O.R.; Kahru, M.; Sherwood, E.T.; et al. Human activities and climate variability drive fast-paced change across the world’s estuarine–coastal ecosystems. Glob. Change Biol. 2016, 22, 513–529. [Google Scholar] [CrossRef]
- Bi, H.; Sun, S.; Gao, S.; Song, S.; Zhang, Z.T. Ecological characteristics of zooplankton communities in the Bohai Sea Ⅲ. Quantity distribution and seasonal changes of some zooplankton. Chin. J. Ecol. 2001, 21, 9. [Google Scholar] [CrossRef]
- Cheng, F. Exploration on the Formation of Seasonal Population Dominance of Cladocera in Daya Bay; Jinan University: Jinan, China, 2019. [Google Scholar]
- Sun, J.; Liu, D.; Yang, S.; Guo, J.; Qian, S. A preliminary study on the phytoplankton community structure in the central Bohai Sea and the Bohai Strait and adjacent waters. Ocean. Limnol. 2002, 33, 11. [Google Scholar] [CrossRef]
- Sun, J.; Dawson, J.; Liu, D. Microzooplankton grazing on phytoplankton in summer in the Jiaozhou Bay, China. Chin. J. Appl. Ecol. 2004, 15, 1245–1252. [Google Scholar]
- Guo, S.; Li, Y.; Zhang, C.; Zhai, W.D.; Huang, T.; Wang, L.F.; Ma, W.; Sun, H.N.; Jin, J. Phytoplankton community structure and its correlation with environmental factors in the Bohai Sea. Mar. Bull. 2014, 33, 11. [Google Scholar] [CrossRef]
- Wu, D.; Wang, H.; Zhang, Z. Investigation of plankton and community structure changes in Yuqiao Reservoir in Tianjin in summer. Lake Sci. 2013, 25, 735–742. [Google Scholar] [CrossRef]
- Luan, Q.; Kang, Y.; Wang, J. Long-term changes of phytoplankton communities in the Bohai Sea (1959–2015). Adv. Fish. Sci. 2018, 39, 10. [Google Scholar] [CrossRef]
- Yang, L.; Liu, J.; Zhang, J.; Wang, X.L.; Xu, Y.; Li, X.; He, L. Changes of zooplankton community in Bohai Bay and its relationship with environmental factors. Oceanogr. Res. 2018, 36, 9. [Google Scholar] [CrossRef]
- Fan, K.; Li, Q. Community structure of zooplankton and biological evaluation of water quality in Bohai Bay. Anhui Agric. Sci. 2007, 1697–1699. [Google Scholar] [CrossRef]
- Xu, Z.; Gao, Q. Distribution and response to global warming of Daphnia chinensis in the Yangtze River Estuary. Chin. J. Appl. Ecol. 2009, 6, 1196–1201. [Google Scholar]
- Qi, X. Talking about the Application of Biotechnology in Water Environment Monitoring. Sci. Technol. Innov. 2019, 2. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Z.; Gao, Q. Changes in abundance of strong arrowworms and obese arrowworms in the Yangtze Estuary in response to environmental warming. Chin. J. Ecol. 2009, 29, 8. [Google Scholar] [CrossRef]
- Pang, B.; Lan, W.; Li, M.; Li, T. Structural characteristics and seasonal changes of zooplankton community in the coastal waters of Beibu Gulf. Chin. J. Ecol. 2019, 39, 11. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, G. The latest scientific points on the cognition of global climate change in the Fifth Assessment Report of IPCC Working Group I. Glacier Permafr. 2013, 35, 9. [Google Scholar] [CrossRef]
- Jang, M.-C.; Shin, K.; Jang, P.-G.; Lee, W.-J.; Choi, K.-H. Mesozooplankton community in a seasonally hypoxic and highly eutrophic bay. Mar. Freshw. Res. 2015, 66, 719–729. [Google Scholar] [CrossRef]
- Llope, M.; de Carvalho-Souza, G.F.; Baldó, F.; González-Cabrera, C.; Jiménez, M.P.; Licandro, P.; Vilas, C. Gulf of Cadiz zooplankton: Community structure, zonation and temporal variation. Prog. Oceanogr. 2020, 186, 102379. [Google Scholar] [CrossRef]
- Gong, Y.; Xiao, Y.; Xu, S.; Liu, Y.; Yang, Y.; Huang, Z.; Li, C. The structure of zooplankton community in Hailing Bay and its relationship with major environmental factors. South. Fish. Sci. 2019, 15, 7. [Google Scholar] [CrossRef]
- Liu, A.; Song, X.; Ren, L. Characteristics of zooplankton community in Laizhou Bay in summer. Mar. Sci. 2012, 36, 61–67. [Google Scholar]
- Li, Y.; Feng, H.; Yuan, D.K.; Guo, L.; Mu, D. Mechanism Study of Transport and Distributions of Trace Metals in the Bohai Bay, China. China Ocean. Eng. 2019, 33, 73–85. [Google Scholar] [CrossRef]
- Lei, T.; Jian, S.; Haiying, L.; Yuanyi, L.; Jiancheng, S.; Binliang, L.; Zhenhua, N.; Dekui, Y. Research on water exchange in Bohai Bay under the action of tide and monsoon. J. Hydroelectr. Power Gener. 2020, 39, 99–107. [Google Scholar] [CrossRef]
- Liu, H. Study on the Effect of Water Eutrophication and Fish Medicine on Zooplankton in Stitute of Hydrobiology, Chinese Academy of Sciences, Hubei. 2003. Available online: https://ir.ihb.ac.cn/handle/342005/19159 (accessed on 1 June 2003).
- Vieira, M.C.; Ortega, J.C.G.; Vieira, L.C.G.; Velho, L.F.M.; Bini, L.M. Evidence that dams promote biotic differentiation of zooplankton communities in two Brazilian reservoirs. Hydrobiologia 2022, 849, 697–700. [Google Scholar] [CrossRef]
- Flood, P.R. Yellow-stained oikopleurid appendicularians are caused by bacterial parasitism. Mar. Ecol. Prog. Ser. 1991, 71, 291–295. [Google Scholar] [CrossRef]
- Acuña, J.L.; Bedo, A.W.; Harris, R.P.; Anadón, R. The Seasonal Succession of Appendicularians (Tunicata: Appendicularia) off Plymouth. J. Mar. Biol. Assoc. United Kingd. 1995, 75, 755–758. [Google Scholar] [CrossRef]
- Vásquez-Yeomans, L.; Castellanos, I.; Suarez-Morales, E.; Gasca, R. Variación espacio-temporal de la biomasa de zooplancton en un sistema estuarino del Caribe Occidental durante dos ciclos anuales. 2012, 47, 213–225. Rev. Biol. Mar. Oceanogr. [CrossRef] [Green Version]
- Bian, S.; Han, L.; Mei, P. Characteristics of zooplankton community structure in Liaodong Bay in summer and autumn and its relationship with environmental factors. J. Tianjin Norm. Univ. 2020, 40, 44–49. [Google Scholar] [CrossRef]
- Zhanhui, Q.; Rongjun, S.; Zonghe, Y.; Shumin, X.; Tingting, H.; Shannan, X.; Honghui, H. Research progress on the effects of filter-feeding shellfish culture on plankton. South. Fish. Sci. 2021, 17, 7. [Google Scholar] [CrossRef]
- Liang, D.; Uye, S. Population dynamics and production of the planktonic copepods in a eutrophic inlet of the Inland Sea of Japan. III. Paracalanus sp. Mar. Biol. 1996, 127, 219–227. [Google Scholar] [CrossRef]
- He, C.; Wang, Y.; Lei, Z.; Xu, S. A preliminary discussion on the formation and properties of the cold waters mass in the Yellow Sea. Ocean Lakes 1959, 11–15. [Google Scholar]
- Li, H.; Song, P. Marine Planktology; University of Science and Technology of China Press: Anhui, China, 2012. [Google Scholar]
- Liu, H.; Wang, Y.; Liang, Z.; Gu, B.; Su, J. Newly discovered seagrass beds and their ecological characteristics in Caofeidian, Bohai Sea. J. Ecol. 2016, 35, 7. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dominant Species | Group | Summer | Autumn | ||||
|---|---|---|---|---|---|---|---|
| F | x | Y | F | x | Y | ||
| Paracalanus parvus | Copepoda | 0.76 | 504.97 | 0.15 | 0.95 | 1403.17 | 0.34 |
| Pseudevadne tergestina | Cladoceera | 0.41 | 469.52 | 0.08 | 0.32 | 70.64 | 0.01 |
| Oikopleura dioica | Tunicate | 0.53 | 220.27 | 0.05 | 0.53 | 188.89 | 0.03 |
| Sagitta crassa | Chaetognatha | 0.94 | 118.10 | 0.04 | 1.00 | 77.15 | 0.02 |
| Penilia avirostris | Cladoceera | 0.24 | 283.02 | 0.03 | 0.16 | 41.86 | 0.01 |
| Pleurobrachia globosa | Hydromedusa | 0.71 | 79.85 | 0.02 | 0.58 | 40.53 | 0.01 |
| Corycaeus affinis | Copepoda | 0.41 | 49.87 | 0.01 | 0.79 | 657.61 | 0.13 |
| Calanus sinicus | Copepoda | 0.47 | 13.96 | 0.003 | 0.53 | 123.85 | 0.02 |
| Oithona similis | Copepoda | 0.17 | 4.07 | 0.0008 | 0.58 | 154.77 | 0.02 |
| Time | 2020 This Study | 2013 Wu et al. [35] | 2005–2009 Yang et al. [36] | 2003–2004 Fan et al. [37] | 1959 Bi et al. [29] |
|---|---|---|---|---|---|
| Summer | Paracalanus parvus Pseudevadne tergestina Oikopleura dioica Sagitta crassa Penilia avirostris Pleurobrachia globosa | Paracalanus parvus Acartia bifilosa Acartia pacifica Oithona similis Calanus sinicus Sagitta crassa Paracalanus crassirostris | Sagitta crassa Calanus sinicus Acartia bifilosa Paracalanus parvus | Sagitta crassa Labidocera euchaeta Paracalanus parvus Paracalanus crassirostris Oithona similis | Paracalanus parvus Paracalanus crassirostris Oithona similis Labidocera euchaeta |
| Autumn | Paracalanus parvus Corycaeus affinis Oikopleura dioica Oithona similis Sagitta crassa Calanus sinicus | Sagitta crassa Calanus sinicus Labidocera euchaeta Acartia pacifica Acartia bifilosa Pleurobrachia globosa | Paracalanus parvus Paracalanus crassirostris Oithona similis |
| Time | Research Area | References Method and Mesh Size | Number of Species and Larvae | Abundance (ind. m−3) | Reference | |
|---|---|---|---|---|---|---|
| July–October 2020 | Bohai Bay | 117°72′−119°14′ E 37°98′−139°98′ N | Microscope 200 μm | 105 | 342.22−23,704.17 | This study |
| July–December 2015 | Daya Bay | 114°30′−114°50′ E 22°30′−22°50′ N | Microscope 505 μm | 131 | 56.3−1129.2 | Xiang et al., 2021 [9] |
| April 2004−February 2006 | Masan Bay | 128°34′−128°46′ E 35°00′−35°12′ N | Microscope 200 μm | − | 94−2300 | Jang et al., 2015 [43] |
| 2001–2015 | Gulf of Cadiz | 6°00′−8°00′ W 36°00′−37°00′ N | Microscope 200 μm | 120 | 280.0−30,000.0 | Llope, et al., 2020 [44] |
| February–November 2015 April 2016 | Hailing Bay | 111°43′−111°57′ E 21°28′−21°38′ N | Microscope 200 μm | 132 | 12.88−5652.85 | Gong et al., 2019 [45] |
| August 2009 | Laizhou Bay | 119°10′−120°30′ E 37°10′−37°80′ N | Microscope 200 μm | 38 | 101.3−3620.0 | Liu et al., 2012 [46] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Wen, Y.; Zhang, G.; Zhang, G.; Sun, J.; Xu, W. Effects of Terrestrial Inputs on Mesozooplankton Community Structure in Bohai Bay, China. Diversity 2022, 14, 410. https://doi.org/10.3390/d14050410
Li D, Wen Y, Zhang G, Zhang G, Sun J, Xu W. Effects of Terrestrial Inputs on Mesozooplankton Community Structure in Bohai Bay, China. Diversity. 2022; 14(5):410. https://doi.org/10.3390/d14050410
Chicago/Turabian StyleLi, Danyang, Yujian Wen, Guodong Zhang, Guicheng Zhang, Jun Sun, and Wenzhe Xu. 2022. "Effects of Terrestrial Inputs on Mesozooplankton Community Structure in Bohai Bay, China" Diversity 14, no. 5: 410. https://doi.org/10.3390/d14050410
APA StyleLi, D., Wen, Y., Zhang, G., Zhang, G., Sun, J., & Xu, W. (2022). Effects of Terrestrial Inputs on Mesozooplankton Community Structure in Bohai Bay, China. Diversity, 14(5), 410. https://doi.org/10.3390/d14050410