
Genetic Diversity and Population Structure of Mesoamerican Scarlet Macaws in an Ex Situ Breeding Population in Mexico

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction
2.3. Amplification and Genotyping
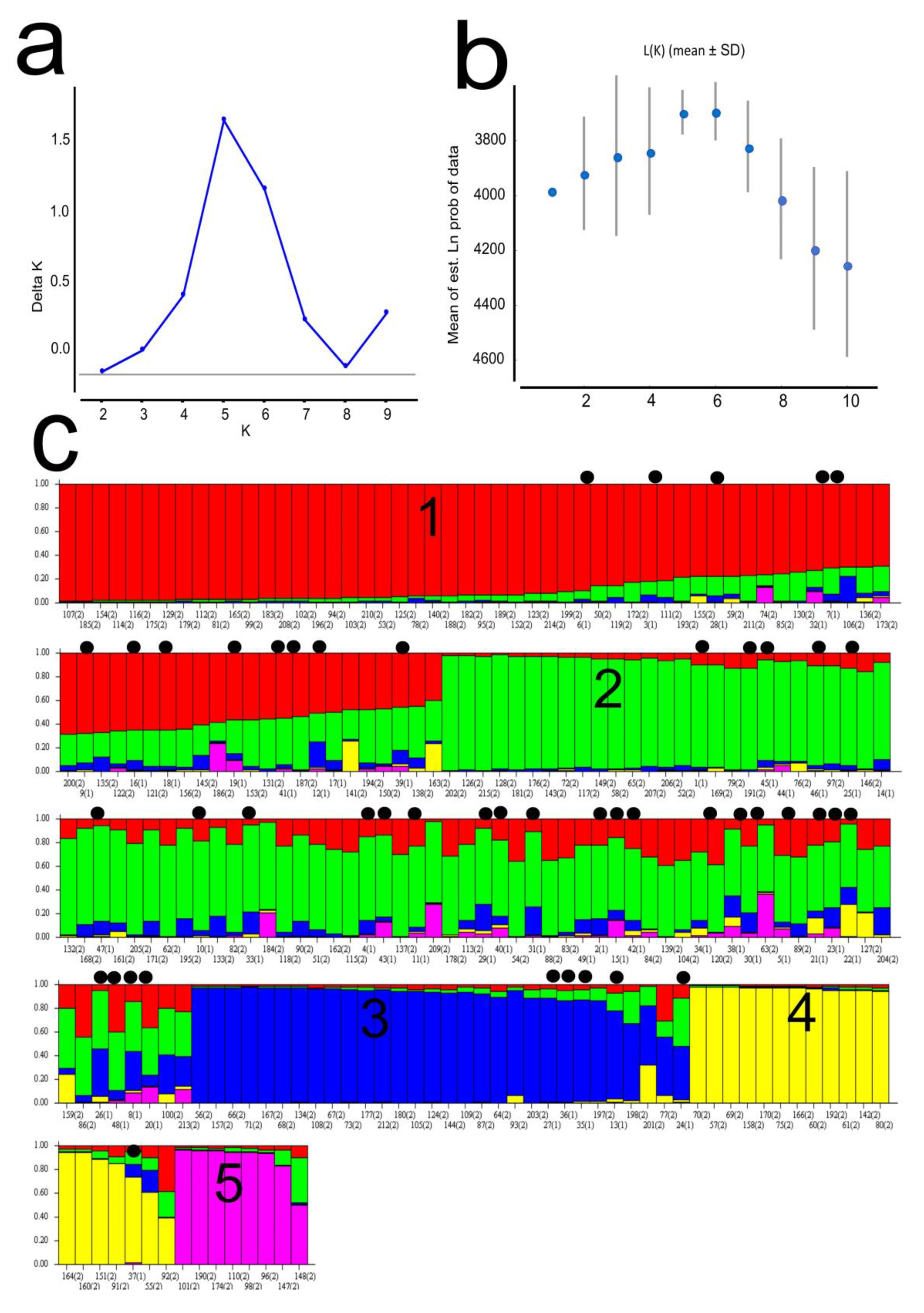
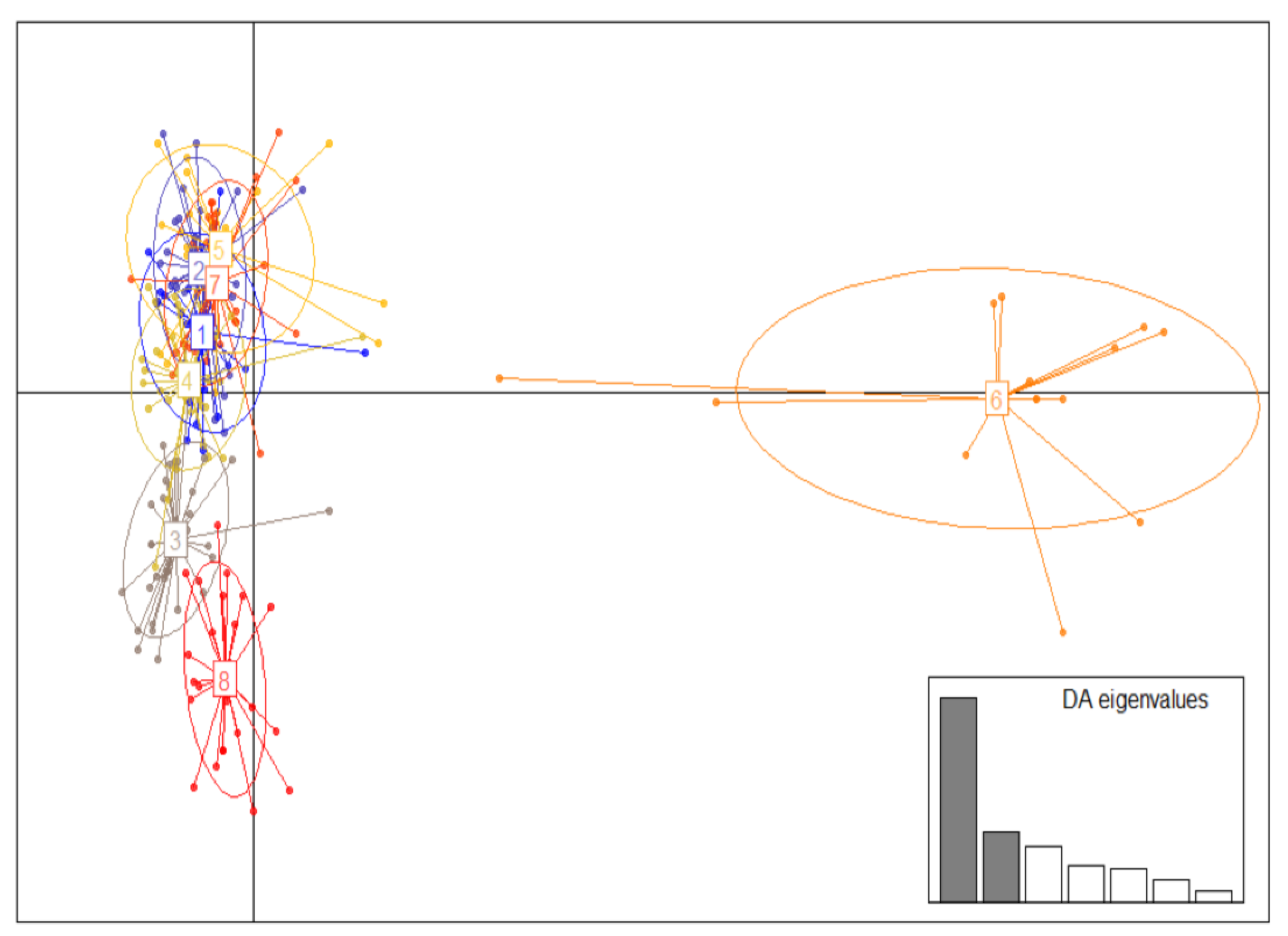
2.4. Population Structure
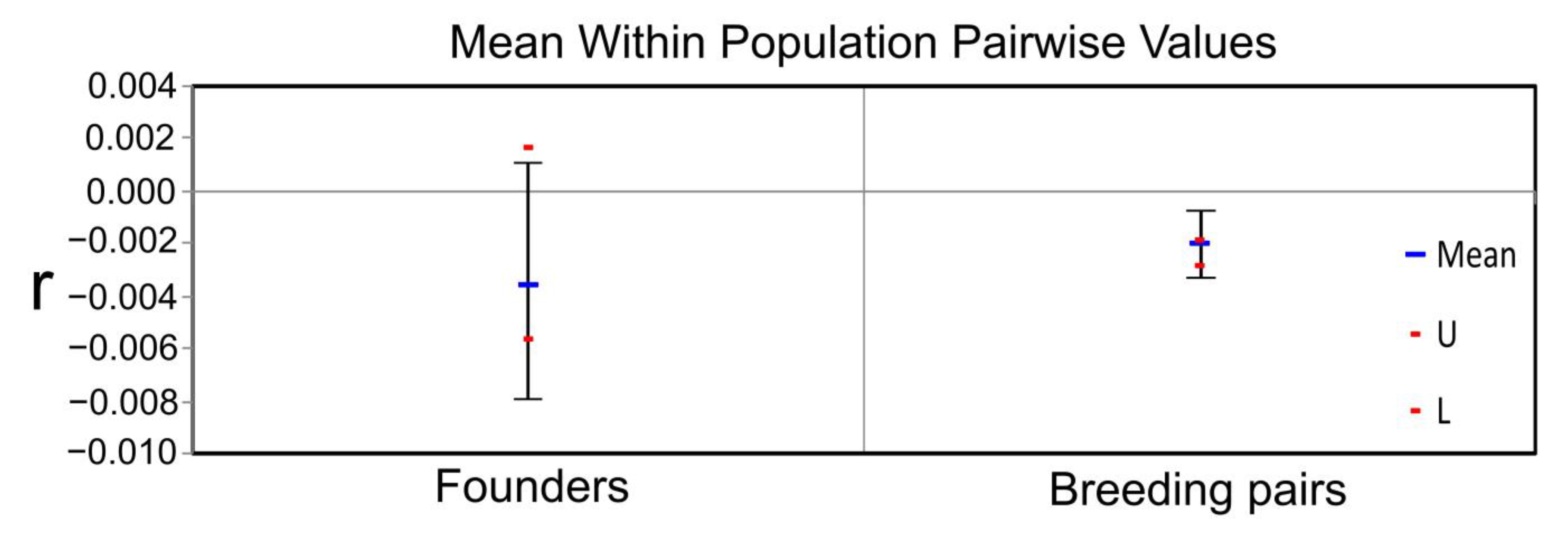
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IUCN/SSC Guidelines for reintroductions and other conservation translocations. In Gland Switz Camb UK IUCNSSC Re-Introd Spec Group; IUCN: Gland, Switzerland; Cambridge, UK, 2013; p. 57.
- Boyce, W.M.; Weisenberger, E.M.; Penedo, M.C.T.; Johnson, C.K. Wildlife translocation: The conservation implications of pathogen exposure and genetic heterozygosity. BMC Ecol. 2011, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Witzenberger, K.A.; Hochkirch, A. Ex situ conservation genetics: A review of molecular studies on the genetic consequences of captive breeding programmes for endangered animal species. Biodivers. Conserv. 2011, 20, 1843–1861. [Google Scholar] [CrossRef]
- Spielman, D.; Brook, B.; Frankham, R. Most species are not driven to extinction before genetic factors impact them. Proc. Natl. Acad. Sci. USA 2004, 101, 15261–15264. [Google Scholar] [CrossRef] [Green Version]
- SEMARNAT (Secretaría de Medio Ambiente y Recursos Naturales). Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección Ambiental-Especies Nativas de México de Flora y Fauna Silvestres-Categorías de Riesgo y Especificaciones para su Inclusión, Exclusión o Cambio-Lista de Especies en Riesgo. DOF del 30 de Diciembre de 2010. Available online: https://www.dof.gob.mx/nota_detalle.php?codigo=5578808&fecha=14/11/2019 (accessed on 10 September 2021).
- Wiedenfeld, D.A. A new subspecies of Scarlet Macaw and its status and conservation. Una nueva subespecie de la lapa roja y su estado de conservación. Ornitol. Neotrop. 1994, 5, 99–104. [Google Scholar]
- SEMARNAT (Secretaría de Medio Ambiente y Recursos Naturales). Programa de Acción para la Conservación de la Especie Guacamaya Roja (Ara macao cyanoptera). Comisión Nacional de Áreas Naturales, Protegidas, Secretaría del Medio Ambiente y Recursos Naturales. 2009. Available online: https://www.conanp.gob.mx/pdf_especies/Pace_Guacamaya_Roja.pdf (accessed on 10 September 2021).
- CITES. Check-List of CITES Species. 2013. Available online: http://checklist.cites.org/# (accessed on 10 September 2021).
- Snyder, N.; McGowan, P.; Gilardi, J.; Grajal, A. (Eds.) Parrots. Status Survey and Conservation Action Plan 2000–2004; IUCN: Gland, Switzerland; Cambridge, UK, 2000; p. 180. [Google Scholar]
- Berkunsky, I.; Quillfeldt, P.; Brightsmith, D.; Abbud, M.; Aguilar, J.; Alemán-Zelaya, U.; Aramburú, R.; Arias, A.A.; McNab, R.B.; Balsby, T.J.S.; et al. Current threats faced by Neotropical parrot populations. Biol. Conserv. 2017, 214, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Boyd, J.D.; McNab, R.B. The Scarlet Macaw in Guatemala and El Salvador: 2008 Status and Future Possibilities. In Findings and Recommendations from a Species Recovery Workshop 9–15 March 2008; Guatemala City and Flores, Petén, Guatemala; Wildlife Conservation Society: Bronx, NY, USA, 2008. [Google Scholar]
- García-Feria, L.M. Un Enfoque Filogeográfico Para la Conservación de Poblaciones de Ara Macao Cyanoptera. Ph.D. Thesis, Instituto de Ecología, AC, Xalapa, Mexico, 2009. [Google Scholar]
- Schmidt, K.L.; Aardema, M.L.; Amato, G. Genetic analysis reveals strong phylogeographical divergences within the Scarlet Macaw Ara macao. Ibis 2020, 162, 735–748. [Google Scholar] [CrossRef]
- Raigoza-Figueras, R. Scarlet macaw Ara macao cyanoptera conservation programme in Mexico. Int. Zoo Yearb. 2014, 48, 48–60. [Google Scholar] [CrossRef]
- Paetkau, D.; Calvert, W.; Stirling, I.; Strobeck, C. Microsatellite analysis of population structure in Canadian polar bears. Mol. Ecol. 1995, 4, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Russello, M.; Calcagnotto, D.; DeSalle, R.; Amato, G. Characterization of microsatellite loci in the endangered St. Vincent Parrot, Amazona guildingii. Mol. Ecol. Notes 2001, 1, 162–164. [Google Scholar] [CrossRef]
- Gebhardt, K.J.; Waits, L.P. Cross-species amplification and optimization of microsatellite markers for use in six Neotropical parrots. Mol. Ecol. Resour. 2008, 8, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Matschiner, M.; Salzburger, W. TANDEM: Integrating automated allele binning into genetics and genomics workflows. Bioinformatics 2009, 25, 1982–1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VAN Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Guo, S.W.; Thompson, E.A. Performing the exact test of Hardy-Weinberg proportion for multiples alleles. Biometrics 1992, 48, 361–372. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goudet, J.F. FSTAT (version 1.2): A computer program to calculate F-statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; vonHoldt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 15 November 2021).
- Wang, J. Informativeness of genetic markers for pairwise relationship and relatedness inference. Theor. Popul. Biol. 2006, 70, 300–321. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. Coancestry: A program for simulating, estimating and analysing relatedness and inbreeding coefficients. Mol. Ecol. Resour. 2010, 11, 141–145. [Google Scholar] [CrossRef]
- Ritland, K. Estimators for pairwise relatedness and individual inbreeding coefficients. Genet. Res. 1996, 67, 175–185. [Google Scholar] [CrossRef]
- Brouwer, K.; Jones, M.L.; King, C.E.; Schifter, H. Longevity records for Psittaciformes in captivity. Int. Zoo Yearb. 2000, 37, 299–316. [Google Scholar] [CrossRef]
- Gonçalves, E.C.; Ferrari, S.F.; Bastos, H.B.; Wajntal, A.; Aleixo, A.; Schneider, M.P. Comparative genetic diversity of wild and captive populations of the bare-faced curassow (Crax fasciolata) based on cross-species microsatellite markers: Implications for conservation and management. Biochem. Genet. 2010, 48, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Presti, F.T.; Oliveira-Marques, A.R.; Caparroz, R.; Biondo, C.; Miyaki, C.Y. Comparative analysis of microsatellite variability in five macaw species (Psittaciformes, Psittacidae): Application for conservation. Genet. Mol. Biol. 2011, 34, 348–352. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K. Spatial and Temporal Patterns of Genetic Variation in Scarlet Macaws (Ara Macao): Implications for Population Management in La Selva Maya. Ph.D. Thesis, Columbia University, New York, NY, USA, 2013. [Google Scholar]
- Monge, O.; Schmidt, K.; Vaughan, C.; Gutiérrez-Espeleta, G. Genetic patterns and conservation of the Scarlet Macaw (Ara macao) in Costa Rica. Conserv. Genet. 2016, 17, 745–750. [Google Scholar] [CrossRef]
- Caparroz, R.; Miyaki, C.Y.; Bampi, M.I.; Wajntal, A. Analysis of the genetic variability in a sample of the remaining group of Spix’s Macaw (Cyanopsitta spixii, Psittaciformes: Aves) by DNA fingerprinting. Biol. Conserv. 2001, 99, 307–311. [Google Scholar] [CrossRef]
- Dear, F.; Vaughan, C.; Polanco, A.M. Current Status and Conservation of the Scarlet Macaw (Ara macao) in the Osa Conservation Area (ACOSA), Costa Rica. UNED Res. J. 2010, 2, 7–21. [Google Scholar] [CrossRef] [Green Version]
- Myers, M.C.; Vaughan, C. Movement and behavior of scarlet macaws (Ara macao) during the post-fledging dependence period: Implications for in situ versus ex situ management. Biol. Conserv. 2004, 118, 411–420. [Google Scholar] [CrossRef]
- Bourke, B.P.; Frantz, A.C.; Lavers, C.P.; Davison, A.; Dawson, D.A.; Burke, T. Genetic signatures of population change in the British golden eagle (Aquila chrysaetos). Conserv. Genet. 2010, 11, 1837–1846. [Google Scholar] [CrossRef]
- Witzenberger, K.A.; Hochkirch, A. Evaluating ex situ conservation projects: Genetic structure of the captive population of the Arabian sand cat. Mamm. Biol. 2013, 78, 379–382. [Google Scholar] [CrossRef]
- Haig, S.M.; Ballou, J.D.; Derrickson, S.R. Management Options for Preserving Genetic Diversity: Reintroduction of Guam Rails to the Wild. Conserv. Biol. 1990, 4, 290–300. [Google Scholar] [CrossRef]
- Jamieson, I.G. Founder Effects, Inbreeding, and Loss of Genetic Diversity in Four Avian Reintroduction Programs. Conserv. Biol. 2011, 25, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Saura, M.; Pérez-Figueroa, A.; Fernández, J.; Toro, M.A.; Caballero, A. Preserving Population Allele Frequencies in Ex Situ Conservation Programs. Conserv. Biol. 2008, 22, 1277–1287. [Google Scholar] [CrossRef]
- Robert, A. Captive breeding genetics and reintroduction success. Biol. Conserv. 2009, 142, 2915–2922. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Founders | |||||||||||
| Loci | N | Range | Na | Ne | Ho | He | P | D | AR | F | HWE |
| AgGT17 | 48 | 115–137 | 10 | 5.592 | 0.875 | 0.821 | ˗ | 0.832 | 9.997 | −0.066 | */- |
| AgGT19 | 48 | 181–189 | 4 | 2.386 | 0.458 | 0.581 | ˗ | 0.595 | 3.939 | 0.211 | - |
| AgGT21 | 48 | 169–189 | 8 | 5.224 | 0.729 | 0.809 | ˗ | 0.819 | 7.939 | 0.098 | - |
| AgGT42 | 45 | 243–271 | 12 | 6.784 | 0.956 | 0.853 | 1 | 0.863 | 12 | −0.121 | - |
| UnaCT21 | 48 | 166–172 | 4 | 3.022 | 0.646 | 0.669 | ˗ | 0.676 | 4 | 0.035 | - |
| UnaCT74 | 48 | 150–168 | 7 | 2.808 | 0.625 | 0.644 | 2 | 0.661 | 6.936 | 0.029 | */** |
| Mean | 7.5 | 4.303 | 0.717 | 0.729 | 7.466 | 0.031 | |||||
| (SE) | 1.31 | 0.735 | 0.074 | 0.046 | 1.316 | 0.048 | |||||
| Breeding Pairs | |||||||||||
| AgGT17 | 159 | 115–137 | 12 | 6.11 | 0.881 | 0.836 | 2 | 0.839 | 10.508 | −0.053 | */- |
| AgGT19 | 166 | 181–189 | 4 | 2.16 | 0.614 | 0.536 | ˗ | 0.537 | 3.605 | −0.147 | */- |
| AgGT21 | 155 | 169–191 | 9 | 4.87 | 0.858 | 0.795 | 1 | 0.797 | 8.093 | −0.08 | */- |
| AgGT42 | 165 | 243–275 | 14 | 6.87 | 0.873 | 0.854 | 3 | 0.857 | 11.369 | −0.022 | */** |
| UnaCT21 | 165 | 150–174 | 6 | 2.98 | 0.715 | 0.664 | 2 | 0.666 | 5.679 | −0.077 | */** |
| UnaCT74 | 155 | 152–168 | 5 | 2.53 | 0.639 | 0.604 | ˗ | 0.606 | 4.995 | −0.058 | - |
| Mean | 8.3 | 4.250 | 0.763 | 0.715 | 7.375 | −0.073 | |||||
| (SE) | 1.6 | 0.810 | 0.050 | 0.054 | 1.279 | 0.017 | |||||
| Species/Population | Origin | N | Na | He | Ho | Reference |
|---|---|---|---|---|---|---|
| A. macao cyanoptera (Mesoamerican scarlet macaw) | ||||||
| Founders (Xcaret) | Captivity | 49 | 7.5 | 0.715 | 0.729 | This study |
| Breeding individuals | Captivity | 166 | 8.3 | 0.763 | 0.715 | This study |
| La Selva Maya, Guatemala | Wild | 37 | 7.1 | 0.696 | 0.713 | [33] |
| A. m. macao (scarlet macaw) | ||||||
| Costa Rica (CP) | Wild | 41 | 6.7 | 0.63 | 0.61 | [34] |
| Costa Rica (SP) | Wild | 55 | 8.1 | 0.68 | 0.65 | [34] |
| Perú | Wild | 25 | 10.3 | 0.833 | 0.84 | [16] |
| Brazil | Wild | 28 | 11.8 | 0.846 | 0.842 | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escalante-Pliego, P.; Matías-Ferrer, N.; Rosas-Escobar, P.; Lara-Martínez, G.; Sepúlveda-González, K.; Raigoza-Figueras, R. Genetic Diversity and Population Structure of Mesoamerican Scarlet Macaws in an Ex Situ Breeding Population in Mexico. Diversity 2022, 14, 54. https://doi.org/10.3390/d14010054
Escalante-Pliego P, Matías-Ferrer N, Rosas-Escobar P, Lara-Martínez G, Sepúlveda-González K, Raigoza-Figueras R. Genetic Diversity and Population Structure of Mesoamerican Scarlet Macaws in an Ex Situ Breeding Population in Mexico. Diversity. 2022; 14(1):54. https://doi.org/10.3390/d14010054
Chicago/Turabian StyleEscalante-Pliego, Patricia, Noemí Matías-Ferrer, Patricia Rosas-Escobar, Gabriela Lara-Martínez, Karol Sepúlveda-González, and Rodolfo Raigoza-Figueras. 2022. "Genetic Diversity and Population Structure of Mesoamerican Scarlet Macaws in an Ex Situ Breeding Population in Mexico" Diversity 14, no. 1: 54. https://doi.org/10.3390/d14010054
APA StyleEscalante-Pliego, P., Matías-Ferrer, N., Rosas-Escobar, P., Lara-Martínez, G., Sepúlveda-González, K., & Raigoza-Figueras, R. (2022). Genetic Diversity and Population Structure of Mesoamerican Scarlet Macaws in an Ex Situ Breeding Population in Mexico. Diversity, 14(1), 54. https://doi.org/10.3390/d14010054