
Fatty Acid Changes in Nearshore Phytoplankton under Anthropogenic Impact as a Biodiversity Risk Factor for the World’s Deepest Lake Baikal

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Experiment
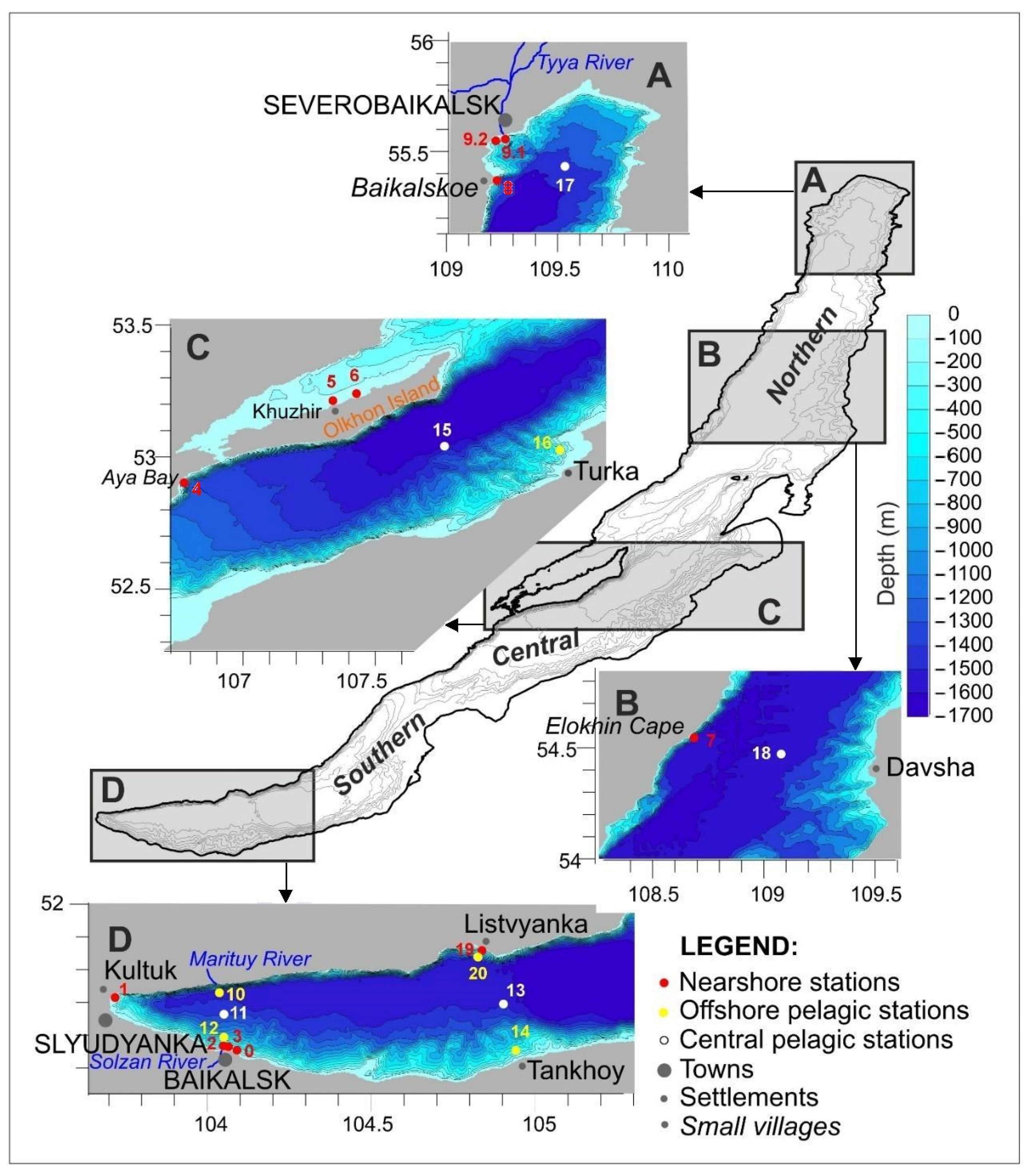
2.1. Sample Collection
2.2. Lipid Extraction and Derivatization of FA to Fatty Acid Methyl Esters (FAMEs)
2.3. FAME Analysis by Gas Chromatography with Flame-Ionization (GC-FID) and Mass-Spectrometric Detection (GC-MS)
2.4. Fatty Acid Peroxidation Estimate by Spectrophotometric Determination of Thiobarbituric acid Reactive Substances (TBARS)
2.5. Statistical Data
2.6. Cultivation of the S. acus
2.7. Anionic Surfactant Analysis by Spectrophotometric Determination
3. Results
3.1. Baikal Phytoplankton Composition
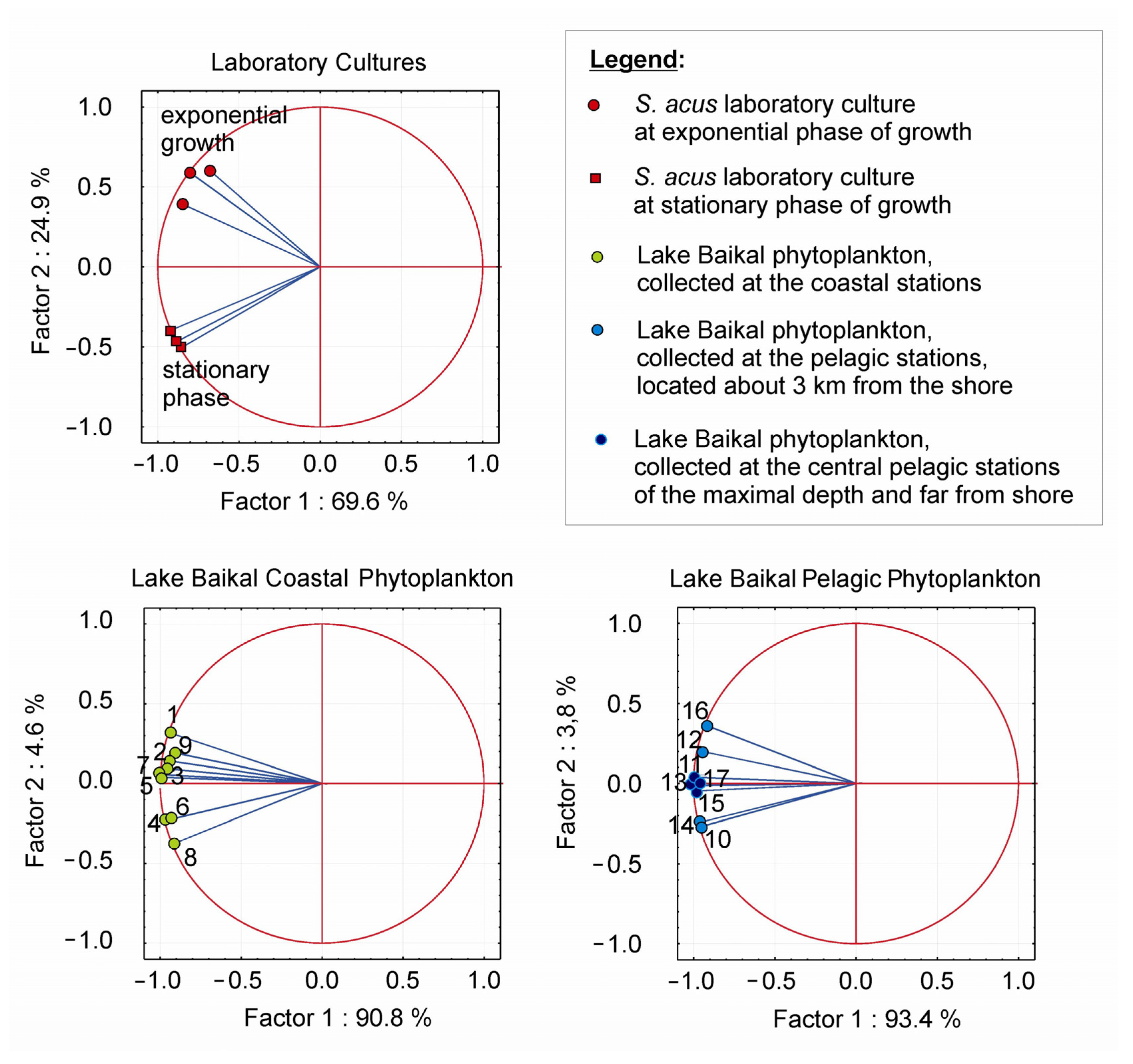
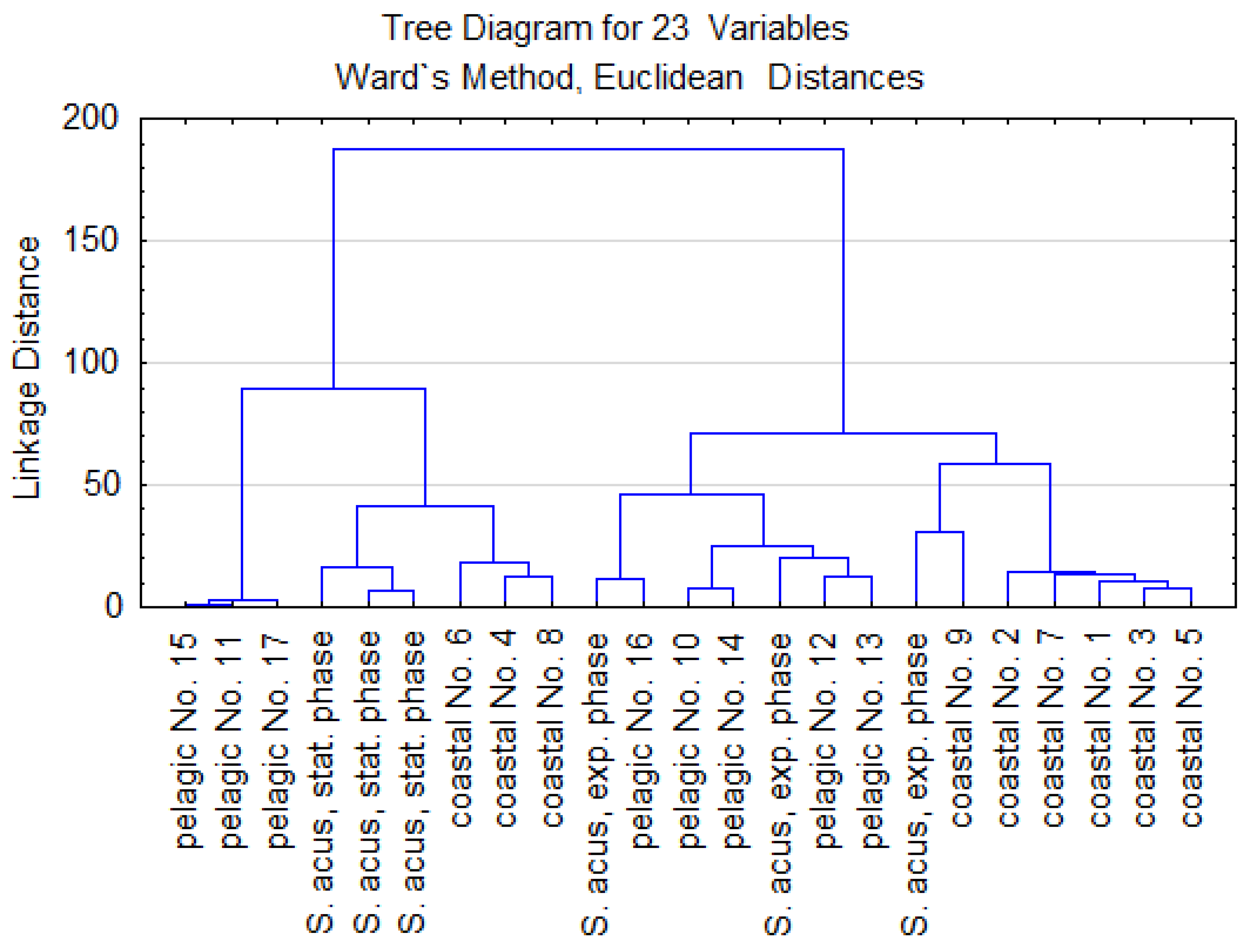
3.2. Statistical Analysis
3.3. FA Composition and FA Peroxidation Product Analysis
4. Discussion
4.1. The Present Composition of Baikal Phytoplankton during the Spring Season
4.2. Phytoplankton FA Composition and FA Peroxidation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmid, M.; De Batist, M.; Granin, N.G.; Kapitanov, V.A.; McGinnis, D.F.; Mizandrontsev, I.B.; Obzhirov, A.I.; Wuest, A. Sources and sinks of methane in Lake Baikal: A synthesis of measurements and modeling. Limnol. Oceanogr. 2007, 52, 1824–1837. [Google Scholar] [CrossRef] [Green Version]
- De Batist, M.; Canals, M.; Sherstyankin, P.; Alekseev, S. The INTAS Project 99-1669 Team, 2002. A New Bathymetric Map of Lake Baikal. Available online: http://www.lin.irk.ru/intas/index.htm (accessed on 6 December 2021).
- Vereschagin, G.Y. Baikal; Gosudartvennoye Izdatel’stvo Geograficheskoi Literatury: Moscow, Russia, 1949; 228p. (In Russian) [Google Scholar]
- Galazii, G.I. (Ed.) Baikal; Russian Academy of Sciences, Siberian Branch, Interdepartmental Scientific Board on “Siberia” Program, Federal Service of Geodesy and Cartography of Russia: Moscow, Russia, 1993; 160p. (In Russian) [Google Scholar]
- Wetzel, R.G. Limnology: Lake and River Ecosystems, 3rd ed.; Academic Press: San Diego, CA, USA; San Francisco, CA, USA; New York, NY, USA; Boston, MA, USA; London, UK; Sydney, Australia; Tokyo, Japan, 2001; 1006p. [Google Scholar]
- Nakashizuka, T.; Stork, N. (Eds.) Biodiversity Research Methods: YBOY (International Biodiversity Observation Year) in Western Pacific and Asia; University Press, Trans Pacific Press: Kyoto, Japan, 2002; 216p. [Google Scholar]
- Karabanov, E.B. Podvodnye Ladshafty Baikala [Underwater Landscapes of Lake Baikal]; Nauka Publ.: Novosibirsk, Russia, 1987; p. 183. (In Russian) [Google Scholar]
- Kozhov, M.M. Biologiya Ozera Baikal [Lake Baikal Biology]; Acad. of Sciences of USSR: Moscow, Russia, 1962; p. 316. [Google Scholar]
- Lut, B.F. Morfologiya i morfometriya Baikalskoi vpadiny [Morphology and Morphometry of Baikal Depression], Chapter II. In The Way for knowledge of Baikal; Afanasieva, E.L., Beckman, M.Y., Bezrukova, E.V., Verbolov, V.I., Votintcev, K.K., Galazii, G.I., Goldyrev, G.S., Granina, L.Z., Dryukker, V.V., Ladeishokov, N.P., et al., Eds.; Nauka Publ.: Novosibirsk, Russia, 1987; pp. 34–47. (In Russian) [Google Scholar]
- Votintsev, K.K.; Meshcheryakova, A.I.; Popovskya, G.I. Krugovorot Organicheskogo Veshchestva v Ozere Baikal [Organic Substance Cycle in Lake Baikal]; Nauka Publ.: Novosibirsk, Russia, 1975; 190p. (In Russian) [Google Scholar]
- Vadeboncoeur, Y.; McIntyre, P.B.; Vander Zanden, M.J. Borders of Biodiversity: Life at the Edge of the World’s Large Lakes. BioScience 2011, 61, 526–537. [Google Scholar] [CrossRef]
- Popovskaya, G.I. Ecological monitoring of phytoplankton in Lake Baikal. Aquat. Ecosyst. Health Manag. 2000, 3, 215–225. [Google Scholar] [CrossRef]
- Shishlyannikov, S.M.; Klimenkov, I.V.; Bedoshvili, E.D.; Mikhailov, I.S.; Gorshkov, A.G. Effect of mixotrophic growth on the ultrastructure and fatty acid composition of the diatom Synedra acus from Lake Baikal. J. Biol. Res. 2014, 21, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rives, D.B.; Jewson, D.H.; Sturm, M.; Battarbee, R.; Flower, R.G.; Mackay, A.W.; Granin, N.G. Quantitative and qualitative relationships between planktonic diatom communities and diatom assemblages in sedimenting material and surface sediments in Lake Baikal, Siberia. Limnol. Oceanogr. 2003, 48, 1643–1661. [Google Scholar] [CrossRef] [Green Version]
- Behrenfeld, M.J.; O’Malley, R.T.; Siegel, D.A.; McClain, C.R.; Sarmiento, J.L.; Feldman, G.C.; Milligan, A.J.; Falkowski, P.G.; Letelier, R.; Boss, E. Climate-driven trends in contemporary ocean productivity. Nature 2006, 444, 752–755. [Google Scholar] [CrossRef]
- Ansari, A.A.; Gill, S.S.; Lanza, G.R.; Rast, W. Eutrophication: Causes, Consequences and Control; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2011; 394p. [Google Scholar]
- Häder, D.P.; Gao, K. Interactions of anthropogenic stress factors on marine phytoplankton. Front. Environ. Sci. 2015, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Echeveste, P.; Galbán-Malagón, C.; Dachs, J.; Berrojalbiz, N.; Agustí, S. Toxicity of natural mixtures of organic pollutants in temperate and polar marine phytoplankton. Sci. Total. Environ. 2016, 571, 34–41. [Google Scholar] [CrossRef]
- Grachev, M.; Bukin, Y.; Blinov, V.; Khlystov, O.; Firsova, A.; Bashenkhaeva, M.; Kamshilo, O.; Titova, L.; Bairamova, E.; Bedoshvili, Y.; et al. Is a High Abundance of Spring Diatoms in the Photic Zone of Lake Baikal in July 2019 Due to an Upwelling Event? Diversity 2021, 13, 504. [Google Scholar] [CrossRef]
- Odum, E.P.; Barrett, G.W. Fundamentals of Ecology, 5th ed.; Cengage Learning: Boston, UK, 2004; 624p. [Google Scholar]
- Votintsev, K.K.; Glazunov, I.V. Hydrochemical regime of Lake Baikal near settlement of Listvennichnyi//Transactions of the Limnological Institute. V. III (XXIII). In Hydrochemical Investigations of Lake Baikal; Galazii, G.I., Ed.; AS RAS Publ.: Moscow, Russia, 1963; pp. 3–56. (In Russian) [Google Scholar]
- Sokolnikov, V.M. Flows in deep layers and water cycle in Baikal. In Flows and Diffusion of Baikal Water; Galazii, G.I., Ed.; AS RAS Publ.: Moscow, Russia, 1963; pp. 67–74. (In Russian) [Google Scholar]
- Afanasyev, A.N.; Verbolov, V.I. Techeniya v Baikale [Flows in Baikal]; Nauka: Novosibirsk, Russia, 1977; 160p. (In Russian) [Google Scholar]
- Ladeishchikov, N.P. Izuchenie klimata Baikala [Baikal Climate Investigation]. In the Way for Knowledge of Baikal; Afanasieva, E.L., Beckman, M.Y., Bezrukova, E.V., Verbolov, V.I., Votintcev, K.K., Galazii, G.I., Goldyrev, G.S., Granina, L.Z., Dryukker, V.V., Ladeishokov, N.P., et al., Eds.; Nauka Publ.: Novosibirsk, Russia, 1987; pp. 76–84. (In Russian) [Google Scholar]
- Goldman, C.R.; Elser, J.J.; Richards, R.C.; Reuter, J.E.; Priscu, J.C.; Levin, A.L. Thermal stratification, nutrient dynamics and phytoplankton productivity during the onset of spring phytoplankton growth in Lake Baikal, Russia. Hydrobiologia 1996, 331, 9–24. [Google Scholar] [CrossRef]
- Genkai-Kato, M.; Sekino, T.; Yoshida, T.; Miyasaka, H.; Khoszher, T.; Belykh, O.A.; Melnik, N.G.; Kawabata, Z.; Higashi, M.; Nakanishi, M. Nutritional diagnosis of phytoplankton in Lake Baikal. Ecol. Res. 2002, 17, 135–142. [Google Scholar] [CrossRef]
- Falk-Petersen, S.; Sargent, J.R.; Henderson, J.; Hegseth, E.N.; Hop, H.; Okolodkov, Y.B. Lipids and fatty acids in ice algae and phytoplankton from the Marginal Ice Zone in the Barents Sea. Polar Biol. 1998, 20, 41–47. [Google Scholar] [CrossRef]
- Zhila, N.O.; Kalacheva, G.S. Effect of salinity on the biochemical composition of the alga Botryococcus braunii Kütz IPPAS H-252. J. Appl. Phycol. 2011, 23, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Wagenen, J.V.; Miller, T.W.; Hobbs, S.; Hook, P.; Crowe, B.; Huesemann, M. Effect of Light and Temperature on fatty acid production in Nannochloropsis Salina. Energies 2012, 5, 731–740. [Google Scholar] [CrossRef] [Green Version]
- Costantini, D. Oxidative Stress and Hormesis in Evolutionary Ecology and Physiology; Springer: Heidelberg, Germany; New York, NY, USA; Dordrecht, The Nehterlands; London, UK, 2014; 346p. [Google Scholar]
- Nikonova, A.A.; Shishlyannikov, S.M.; Shishlyannikova, T.A.; Avezova, T.N.; Babenko, T.A.; Belykh, O.I.; Glyzina, O.Y.; Obolkin, V.A.; Pavlova, O.N.; Smagunova, A.N.; et al. Determination of Free and Esterified Fatty Acids in Hydrocoles of Different Content of Polyunsaturated Fatty Acids by Gas–Liquid Chromatography. J. Anal. Chem. 2020, 75, 1310–1321. [Google Scholar] [CrossRef]
- Abedi, E.; Sahari, A.M. Long-chain polyunsaturated fatty acid sources and evaluation of their nutrition and functional properties. Food Sci. Nutr. 2014, 2, 443–463. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry, 5th ed.; W. H. Freeman and Company: New York, NY, USA, 2002; 1208p. [Google Scholar]
- Goncalves, A.M.M.; Marques, J.C.; Goncalves, F. Fatty Acids’ Profiles of Aquatic Organisms: Revealing the Impacts of Environental and Anthropogenic Stressors; Intech Open: London, UK, 2017; Chapter 6; pp. 89–117. [Google Scholar]
- Arts, M.T.; Ackman, R.G.; Holub, B.J. “Essential fatty acids” in aquatic ecosystems: A crucial link between diet and human health and evolution. Can. J. Fish. Aquat. Sci. 2001, 58, 122–137. [Google Scholar] [CrossRef]
- Hampel, M.; Ortiz-Delgado, J.B.; Sarasquete, C.; Blasco, J. Effects of sediment sorbed linear alkylbenzene sulphonate on juveniles of the Senegal sole, Solea senegalensis: Toxicity and histological indicators. Histol. Histopathol. 2008, 23, 87–100. [Google Scholar] [PubMed]
- Katagi, T. Surfactant Effects on Environmental Behavior of Pesticides. Rev. Environ. Contam. Toxicol. 2008, 194, 71–177. [Google Scholar]
- Zelzer, S.; Oberreither, R.; Bernecker, C.; Stelzer, I.; Truschnig-Wilders, M.; Fauler, G. Measurement of total and free malondialdehyde by gas–chromatography mass spectrometry-comparison with high-performance liquid chromatography methology. Free. Radic. Res. 2013, 47, 651–656. [Google Scholar] [CrossRef]
- Janero, D.R. Malondialdehyde and thiobarbituric acidreactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med. 1990, 9, 515–540. [Google Scholar] [CrossRef]
- Lopez, C.A.; Mendez, J.C. Anticancer drugs acting via radical species, photosensitizers and photodynamic therapy of cancer. Chapter 4. In Medicinal Chemistry of Anticancer Drugs; Carmen, A., Carlos Menéndez, J., Eds.; Elsevier B.V.: Madrid, Spain, 2008; pp. 93–138. [Google Scholar]
- Marnett, L.J. Lipid peroxidation-DNA damage by malondialdehyde. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 1999, 424, 83–95. [Google Scholar] [CrossRef]
- Zhou, J.; Wu, Z.; Yu, D.; Pang, Y.; Cai, H.; Liu, Y. Toxicity of linear alkylbenzene sulfonate to aquatic plant Potamogeton perfoliatus L. Environ. Sci. Pollut. Res. 2018, 25, 32303–32311. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, N.A.; Logacheva, N.F. Structural Changes in Phytoplankton of the Littoral Zone of Lake Baikal. Hydrobiol. J. 2017, 53, 16–24. [Google Scholar] [CrossRef]
- Popovskaya, G.I.; Usoltseva, M.V.; Domysheva, V.M.; Sakirko, M.V.; Blinov, V.V.; Khodzher, T.V. The spring phytoplankton in the pelagic zone of Lake Baikal during 2007–2011. Geogr. Nat. Resoures 2015, 36, 253–262. [Google Scholar] [CrossRef]
- Timoshkin, O.A.; Samsonov, D.P.; Yamamuro, M.; Moore, M.V.; Belykh, O.I.; Malnik, V.; Sakirko, M.; Shirokaya, A.; Bondarenko, N.; Domysheva, V.; et al. Rapid ecological change in the coastal zone of Lake Baikal (East Siberia): Is the site of the world’s greatest freshwater biodiversity in danger? J. Great Lakes Res. 2006, 42, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Khanaev, I.V.; Kravtsova, L.S.; Maikova, O.O.; Bukshuk, N.A.; Sakirko, M.V.; Kulakova, N.V.; Butina, T.V.; Nebesnykh, I.A.; Belikov, D.I. Current state of the sponge fauna (Porifera: Lubomirskiidae) of Lake Baikal: Sponge disease and the problem of conversation of diversity. J. Great Lakes Res. 2018, 44, 77–85. [Google Scholar] [CrossRef]
- Maikova, O.O.; Kravtsova, L.S.; Khanaev, I.V. Baikal endemic sponges in the system of ecological monitoring. Limnol. Freshw. Biol. 2020, 1, 364–367. [Google Scholar] [CrossRef] [Green Version]
- Moselhy, H.F.; Reid, R.G.; Yousef, S.; Boyle, S.P. A specific, accurate, and sensitive measure of total plasma malondialdehyde by HPLC. J. Lipid Res. 2013, 54, 852–858. [Google Scholar] [CrossRef] [Green Version]
- Al-Rashed, S.A.; Ibrahim, M.M.; El-Gaaly, G.A.; Al-Shehri, S.; Mostafa, A. Evaluation of radical scavenging system in two microalgae in response to interactive stresses of UV-B radiation and nitrogen starvation. Saudi J. Biol. Sci. 2016, 23, 706–712. [Google Scholar] [CrossRef] [Green Version]
- Rakita, S.M.; Colovie, D.S.; Levart, A.R.; Banjac, V.V.; Colovie, R.R.; Dragojlović, D.M.; Đuragić Olivera, M. A rapid spectrophotometric method for determination of thiobarbituric acid reactive substances in rainbow trout feed. Food Feed Res. 2020, 47, 43–53. [Google Scholar] [CrossRef]
- Smagunova, A.N. Metody Matematicheskoi Statistiki v Analiticheskoi Khimii [Methods of Mathematical Statistics in Analytical Chemistry]; Gosudarstvennyi Universitet: Irkutsk, Russia, 2008; 340p. (In Russian) [Google Scholar]
- Mardia, K.V.; Kent, J.T.; Bibby, J.M. Multivariate Analysis; Academic Press Limited Publ.: London, UK, 1979; 100p. [Google Scholar]
- Yule, G.U.; Kendall, M.G. An Introduction to the Theory of Statistics, 14th ed.; Griffin & Company Publ.: Glasgow, UK, 1950; 701p. [Google Scholar]
- Liu, N.; Wu, Z. Toxic effects of linear alkylbenzene sulfonate on Chara vulgaris L. Environ. Sci. Pollut. Res. 2018, 25, 4934–4941. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Tripathi, G.; Mishra, S.; Gupta, G. Copper-induced oxidative stress and responses of antioxidants and phytochelatins in Hydrilla verticillata (L.f.). Royle Aquatic Toxicol. 2006, 8, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Vega-López, A.; Ayala-López, G.; Posadas-Espadas, B.P.; Olivares-Rubio, H.F.; Dzul-Caamal, R. Relations of oxidative stress in freshwater phytoplankton with heavy metals and polycyclic aromatic hydrocarbon. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 165, 498–507. [Google Scholar] [CrossRef]
- Guoa, J.; Penga, J.; Leia, Y.; Kanervab, M.; Lia, Q.; Song, J.; Guo, J.; Sun, H. Comparison of oxidative stress induced by clarithromycin in two freshwater microalgae Raphidocelis subcapitata and Chlorella vulgaris. Aquat. Toxicol. 2020, 219, 105376. [Google Scholar] [CrossRef] [PubMed]
- Nikonova, A.A. Organic synthetic anionic surfactants as persistent organic pollutants of water ecosystems. Limnol. Freshw. Biol. 2020, 4, 620–621. [Google Scholar] [CrossRef]
- Lewis, A.M. Chronic and sublethal toxicities of surfactants to aquatic animals: A review and risk assessment. Water Res. 1991, 25, 101–113. [Google Scholar] [CrossRef]
- Jorgensen, E.; Christoffersen, K. Short-term effects of linear alkylbenzene sulfonate on freshwater Plankton studied under field conditions. Environ. Toxicol. Chem. 2000, 19, 904–911. [Google Scholar] [CrossRef]
- Stamatelatou, K.; Pakou, C.; Lyberatos, G. 6.37.4.1. Linear alkylbenzene sulfonates (LAS). Occurrence, toxicity, and biodegradation of selected emerging priority pollutants in municipal sewage sludge. In Comprehensive Biotechnology, 2nd ed.; Moo-Young, M., Ed.; Pergamon: Oxford, UK, 2011; Volume 6, pp. 473–484. [Google Scholar]
- Vorobyeva, S.S. Phytoplankton assemblages of the Southern Baikal in 1990–1995 and 2016–2018. Limnol. Freshw. Biol. 2018, 2, 141–143. [Google Scholar] [CrossRef]
- Bondarenko, N.A.; Vorobyova, S.S.; Zhuchenko, N.A.; Golobokova, L.P. Current state of phytoplankton in the littoral area of Lake Baikal, spring 2017. J. Great Lakes Res. 2020, 46, 17–28. [Google Scholar] [CrossRef]
- Vologina, E.G.; Sturm, M.; Vorobyeva, S.S. Present Sedimentation in Lake Baikal. Experimental Results with Sedimentary Traps. Dvenadtsatoe Uralskoe Litilogicheskoe Soveshanie [12th Ural Litologic Workshop]; IGGUrO RAN Publ.: Ekaterinburg, Russia, 2018; pp. 62–63. [Google Scholar]
- Tomberg, I.V.; Sakirko, M.V.; Domysheva, V.M.; Sez’ko, N.P.; Lopatina, I.N.; Bashenkhaeva, N.V.; Philevich, E.A.; Kulikova, N.N.; Popova, O.V.; Malnik, V.V.; et al. First Data about Chemical Composition of Interstitial Water of Lake Baikal Splash Zone; Series “Biology. Ecology”; Izvestiya Irkutskogo Gosudarstvennogo Universiteta [Bull. of Irkutsk State University]: Irkutsk, Russia, 2012; Volume 5, pp. 64–74. (In Russian) [Google Scholar]
- Rusinek, O.T.; Takhteev, V.V.; Gladkochub, D.P.; Khodzher, T.V.; Budnev, N.M.; Bezrukova, E.V.; Galkin, V.V.; Galkina, V.I.; Granina, L.Z.; Goryunova, O.I.; et al. Baikalogy: In 2 books. Book 1; Nauka Publ.: Novosibirsk, Russia, 2012; 468p. [Google Scholar]
- Krotova, V.A. Geostrophic Baikal Water Circulation at the Straight Stratification Period. Trudy Limnologicheskogo Institute SO AN SSSR [Transactions of Limnological Institute SB RAS, USSR]; Nauka Publ.: Leningrad, Russia, 1970; pp. 11–45. (In Russian) [Google Scholar]
- Gagarinova, O.V.; Belozertseva, I.A.; Vorobyeva, I.B.; Vlasova, N.V.; Emelyanova, N.V.; Sorokovoi, A.A. Anthropogenic transformations in the mouth area of tributaries as factors of negative impact on Lake Baikal. Water 2021, 13, 1295. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algae Taxa | Sample Stations in South Basin of the Lake | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Western Flank Nearshore Sampling Site No. 19 | Pelagic Sampling Site No. 20 | Eastern Flank Nearshore Sampling Site No. 14 | ||||||||
| Sampling Depth | ||||||||||
| 1 m | 1 m | 50 m | 400 m | 1 m | ||||||
| Number of cells N, 103 cells L−1 and cell biomass B, mg m−3 | ||||||||||
| N | B | N | B | N | B | N | B | N | B | |
| Class Cyanophyta | ||||||||||
| Anabaena sp. | – | – | – | – | 3.4 | 0.34 | – | – | – | – |
| Oscillatoria sp. | 42 | 5.9 | – | – | – | – | – | – | – | – |
| Lyngbya sp. | – | – | – | – | – | – | – | – | – | – |
| Phormidium sp. | – | – | – | – | – | – | – | – | – | – |
| Total | 42 | 5.9 | – | – | 3.4 | 0.3 | – | – | – | – |
| Class Chrysophyta | ||||||||||
| Dinobryon cylindricum O.E. Imhof | 0.2 | 0.4 | 0.3 | 0.6 | 1.4 | 2.7 | 2.5 | 4.8 | 0.4 | 0.20 |
| Cystes of Chrysophyta | 0.6 | 0.3 | 0.6 | 0.3 | 1.4 | 0.7 | 6.5 | 3.3 | – | – |
| Dinobryon sociale (Ehrenberg) Ehrenberg | – | – | – | – | – | – | – | – | – | – |
| Mallomonas vannigera Asmund | – | – | – | – | – | – | – | – | 98 | 4.9 |
| Chrysochromulina parva Lackey | 58 | 2.9 | 42 | 2.1 | 8.4 | 0.4 | 0.5 | 0.03 | – | – |
| Total | 59 | 3.6 | 43 | 3.0 | 11 | 3.8 | 10 | 8.0 | 98 | 5.1 |
| Class Cryptophyta | ||||||||||
| Rhodomonas pusilla (H. Bachmann) Javornicky | 65 | 16 | 26 | 6.3 | 17 | 4.0 | 2.2 | 0.5 | 145 | 35 |
| Cryptomonas gracilis Skuja | 0.4 | 0.8 | 0.6 | 1.2 | 1.1 | 2.2 | 0.5 | 1.1 | 0.6 | 1.2 |
| Cryptomonas ovata Ehrenberg | – | – | – | – | – | – | – | – | – | – |
| Total | 65 | 16 | 27 | 7.5 | 18 | 6.3 | 2.7 | 1.6 | 146 | 36 |
| Class Dinophyta | ||||||||||
| Gyrodinium helveticum (Penard) Y. Takano and T. Horiguchi | 0.4 | 10 | 0.9 | 23 | 2.3 | 58 | 1.1 | 28 | 0.4 | 11 |
| Gymnodinium baicalense N.L. Antipova | 0.2 | 6 | 0.15 | 4.5 | – | – | – | – | 0.20 | 6.0 |
| Peridinium baicalense Kisselev and V. Zvetkov | – | – | – | – | – | – | – | – | – | – |
| Glenodinium sp. | 0.4 | 0.6 | 0.9 | 1.3 | 1.1 | 1.7 | 2.7 | 4.1 | 1.4 | 2.0 |
| Ceratium hirundinella (O.F.Müller) Dujardin | – | – | – | – | – | – | – | – | – | – |
| Total | 1.0 | 17 | 1.9 | 28 | 3.4 | 59 | 3.8 | 32 | 2.0 | 19 |
| Class Bacillariophyta | ||||||||||
| Aulacoseira baikalensis (Wislouch) Simonsen | 0.4 | 6 | – | – | 0.3 | 4.5 | 0.5 | 8.1 | 3.2 | 48 |
| Aulacoseira islandica (O.Müller) Simonsen + spore | 0.8 | 4 | 0.6 | 3 | 2 | 10 | 3.8 | 19 | 34 | 170 |
| Synedra acus subsp. radians (Kützing) Skabichevskij | 131 | 249 | 210 | 399 | 140 | 266 | 65 | 124 | 280 | 532 |
| Synedra ulna var. danica (Kützing) Van Heurck | 0.2 | 0.6 | – | – | – | – | 0.5 | 1.7 | – | – |
| Synedra ulna (Nitzsch) Ehrenberg | 0.1 | 0.22 | – | – | – | – | – | – | – | – |
| Nitzschia graciliformis Lange-Bertalot and Simonsen | – | – | 0.3 | 0.08 | – | – | – | – | – | – |
| Cyclotella minuta (Skvortsov) Antipova | 0.2 | 0.4 | – | – | 0.3 | 0.6 | 0.3 | 0.6 | 0.20 | 0.4 |
| Cyclotella baicalensis Skvortsov and K.I.Meyer | 0.2 | 4.0 | 0.3 | 5.8 | 0.3 | 6 | 0.3 | 6.0 | 0.20 | 4.0 |
| Ellerbeckia teres (Brun) R.M.Crawford | – | – | – | – | – | – | – | – | – | – |
| Stephanodiscus meyeri Genkal and Popovskaya | 0.2 | 0.2 | 0.6 | 0.6 | 0.6 | 0.6 | 0.8 | 0.8 | – | – |
| Asterionella formosa Hassall | – | – | – | – | – | – | – | – | – | – |
| Total | 133 | 264 | 212 | 408 | 144 | 288 | 71 | 160 | 318 | 754 |
| Class Chlorophyta | ||||||||||
| Monoraphidium arcuatum (Korshikov) Hindák | 15 | 1.8 | 8.6 | 1.0 | 13 | 1.6 | 0.8 | 0.10 | 6.2 | 0.7 |
| Monoraphidium contortum (Thuret) Komárková-Legnerová | – | – | – | – | 0.8 | 0.11 | – | – | – | – |
| Koliella longiseta (Vischer) Hindák | 0.3 | 0.08 | 0.6 | 0.15 | 0.3 | 0.08 | 0.8 | 0.20 | 0.7 | 0.18 |
| Elakatothrix genevensis (Reverdin) Hindák | – | – | 0.14 | 0.02 | – | – | – | – | – | – |
| Scenedesmus acuminatus (Lagerheim) Chodat | – | – | – | – | – | – | – | – | – | – |
| Scenedesmus sp. | – | – | – | – | – | – | – | – | – | – |
| Closterium moniliferum Ehrenberg ex Ralfs | – | – | – | – | – | – | – | – | – | – |
| Pediastrum boryanum (Turpin) Meneghini | – | – | – | – | – | – | – | – | – | – |
| Chlamidomonas sp. | – | – | – | – | – | – | 0.3 | 0.15 | – | – |
| Total | 16 | 1.9 | 9.3 | 1.2 | 14.1 | 1.7 | 1.9 | 0.4 | 6.9 | 0.9 |
| Total phytoplankton biomass | 316 | 309 | 293 | 448 | 194 | 359 | 89 | 202 | 571 | 815 |
| Class of Algae | Sample Stations in the Southern Basin of Lake Baikal | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Western Flank Nearshore Station | Central Pelagic Station | Eastern Flank Nearshore Station | ||||||||
| Sampling Depth | ||||||||||
| 1 m | 1 m | 50 m | 400 m | 1 m | ||||||
| Number of cells N (%) and cell biomass B (%) | ||||||||||
| N | B | N | B | N | B | N | B | N | B | |
| Cyanophyta | 13 | 1.9 | – | – | 1.8 | 0.09 | – | – | – | – |
| Chrysophyta | 19 | 1.2 | 15 | 0.7 | 5.8 | 1.1 | 11 | 4.0 | 17.2 | 0.6 |
| Cryptophyta | 21 | 5.3 | 9.2 | 1.7 | 9.3 | 1.7 | 3.1 | 0.8 | 26 | 4.4 |
| Dinophyta | 0.3 | 5.4 | 0.7 | 6.3 | 1.8 | 16 | 4.3 | 16 | 0.4 | 2.3 |
| Bacillariophyta | 42 | 86 | 72 | 91 | 74 | 80 | 80 | 79 | 56 | 93 |
| Chlorophyta | 4.9 | 0.6 | 3.2 | 0.3 | 7.3 | 0.5 | 2.1 | 0.22 | 1.2 | 0.11 |
| Sampling Stations | Northern Latitude (N) | Eastern Longitude (E) | B (%) of S. acus | Other Predominant Bacillariophyta |
|---|---|---|---|---|
| 0 | 51°31′391″ | 104°11′398″ | 91 | A. baicalensis, C. baicalensis, A. islandica |
| 1 | 51°43′147″ | 103°42′964″ | 95 | A. baicalensis, C. baicalensis, A. islandica |
| 2 | 51°31′726″ | 104°08′086″ | 91 | A. baicalensis, C. baicalensis, A. islandica |
| 3 | 51°31′726″ | 104°08′086″ | 91 | A. baicalensis, C. baicalensis, A. islandica |
| 4 | 52°47′332″ | 106°36′445″ | 85 | A. islandica |
| 5 | 53°12′316″ | 107°19′587″ | 87 | A. islandica |
| 6 | 53°13′201″ | 107°22′290″ | 87 | A. islandica |
| 7 | 54°32′285″ | 108°39′814″ | 89 | A. baicalensis, A. islandica |
| 8 | 55°21′428″ | 109°12′713″ | 88 | A. baicalensis, A. islandica |
| 9.1 | 55°35′385″ | 109°20′720″ | 88 | A. baicalensis, A. islandica |
| 9.1 | 55°34′021″ | 109°13′677″ | 88 | A. baicalensis, A. islandica |
| 19 | 51°49′094″ | 104°54′776″ | 95 | A. baicalensis, C. baicalensis, A. islandica |
| FAs | FA Composition, % | ||||||
|---|---|---|---|---|---|---|---|
| Phytoplankton Sampling Stations | Laboratory S. acus Culture | ||||||
| Nearshore Stations Far from Settlements | Central Pelagic Stations | Nearshore Stations Near Settlements | Offshore Pelagic Stations | 4 h | 24 h | 2 Weeks | |
| C16:1 ω9 | 30.4 | 21.7 | 19.2 | 12.2 | 12.0 | 13.3 | 27.0 |
| C16:0 | 17.3 | 11.4 | 17.4 | 13.6 | 37.2 | 36.1 | 21.0 |
| C16:3 ω4 | 5.5 | 14.2 | 4.5 | 10.8 | 4.8 | 6.0 | 8.4 |
| C16:2 ω4 | 3.6 | 4.3 | 2.9 | 4.7 | 2.2 | 2.9 | 3.9 |
| C18:1 ω9 | 2.3 | 1.9 | 2.3 | 0.7 | 5.6 | 5.4 | 1.7 |
| C18:0 | 5.1 | 1.5 | 5.1 | 12.8 | 19 | 16 | 5.5 |
| C18:4 ω3 | 2.4 | 0.2 | 2.4 | 0.8 | 1.65 | 2.11 | 2.4 |
| C18:3 ω3 | 1.7 | 0.3 | 1.7 | 0.5 | 1.55 | 1.33 | 1.04 |
| C20:4 ω6 | 0.3 | 0.3 | 0.1 | 0.3 | 0.08 | 0.05 | 0.02 |
| C20:5 ω3 | 12.9 | 23.0 | 13.3 | 14.6 | 6.6 | 8.9 | 13.0 |
| C22:6 ω3 | 3.8 | 1.7 | 2.0 | 3.7 | 0.59 | 1.02 | 1.27 |
| PUFAs | 30.3 | 44.1 | 27.6 | 35.4 | 10.1 | 14.7 | 31.9 |
| PUFAs + MUFAs * | 65 | 72 | 53 | 50 | 37 | 43 | 68 |
| Samples | Location | Lake Zone | Total FA, mg g−1 | PUFA and MUFA Percentage | TBARS, μg g−1 of d.w. |
|---|---|---|---|---|---|
| 11 | Marituy River—Solzan River (2016) | Pelagic | 15.8 ± 1.4 | 72 | Not found |
| 17 | Baikalskoe settlement—Frolikha River (2016) | Pelagic | 17.0 ± 1.5 | 72 | Not found |
| 18 | Elokhin Cape—Davsha settlement | Pelagic | 24.2 ± 2.2 | 70 | Not found |
| 1 | Kultuk settlement (2016) | Nearshore | 24.0 ± 2.1 | 56 | 80 |
| 1 | Kultuk settlement (2018) | Nearshore | 20.7 ± 1.8 | 56 | 100 |
| 2 | Baikalsk town (2016) | Nearshore | 27.1 ± 2.4 | 58 | 280 |
| 3 | Baikalsk town (2016) | Nearshore | 27.1 ± 2.4 | 58 | 340 |
| 0 | Baikalsk pulp and paper mill (2016) | Nearshore | 26.2 ± 2.3 | 60 | 160 |
| 0 | Baikalsk pulp and paper mill (2018) | Nearshore | 21.3 ± 1.9 | 54 | 164 |
| 4 | Aya Bay (2016) | Nearshore | 28.1 ± 2.5 | 66 | Not found |
| 6 | Shamanka Bay (2016) | Nearshore | 28.7 ± 2.6 | 60 | 14 |
| 7 | Elokhin Cape (2016) | Nearshore | 18.1 ± 1.6 | 55 | 180 |
| 8 | Baikalskoe settlement (2016) | Nearshore | 49 ± 4 | 68 | Not found |
| 9.1 | Tyya River mouth (2016) | Nearshore | 26.7 ± 2.4 | 46 | 100 |
| 9.1 | Tyya River mouth (2018) | Nearshore | 15.6 ± 1.4 | 58 | 11 |
| 9.2 | Senogda Bay (2016) | Nearshore | 14.0 ± 1.3 | 40 | 190 |
| S. acus laboratory culture, +6 °C, DM medium, 12 h | 4.2 ± 0.4 | 48 | Not found | ||
| S. acus laboratory culture, +6 °C, DM medium, 24 h | 35 ± 3 | 74 | Not found | ||
| S. acus laboratory culture, +6 °C, DM medium, 50 days | No data | Not found | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikonova, A.A.; Shishlyannikov, S.M.; Volokitina, N.A.; Galachyants, Y.P.; Bukin, Y.S.; Blinov, V.V.; Gnatovsky, R.Y.; Vorobyeva, S.S. Fatty Acid Changes in Nearshore Phytoplankton under Anthropogenic Impact as a Biodiversity Risk Factor for the World’s Deepest Lake Baikal. Diversity 2022, 14, 55. https://doi.org/10.3390/d14010055
Nikonova AA, Shishlyannikov SM, Volokitina NA, Galachyants YP, Bukin YS, Blinov VV, Gnatovsky RY, Vorobyeva SS. Fatty Acid Changes in Nearshore Phytoplankton under Anthropogenic Impact as a Biodiversity Risk Factor for the World’s Deepest Lake Baikal. Diversity. 2022; 14(1):55. https://doi.org/10.3390/d14010055
Chicago/Turabian StyleNikonova, Alyona Alexandrovna, Sergey Mikhailovich Shishlyannikov, Nadezhda Antonovna Volokitina, Yuri Pavlovich Galachyants, Yuri Sergeevich Bukin, Vadim Vasil’evich Blinov, Ruslan Yur’evich Gnatovsky, and Svetlana Semyonovna Vorobyeva. 2022. "Fatty Acid Changes in Nearshore Phytoplankton under Anthropogenic Impact as a Biodiversity Risk Factor for the World’s Deepest Lake Baikal" Diversity 14, no. 1: 55. https://doi.org/10.3390/d14010055
APA StyleNikonova, A. A., Shishlyannikov, S. M., Volokitina, N. A., Galachyants, Y. P., Bukin, Y. S., Blinov, V. V., Gnatovsky, R. Y., & Vorobyeva, S. S. (2022). Fatty Acid Changes in Nearshore Phytoplankton under Anthropogenic Impact as a Biodiversity Risk Factor for the World’s Deepest Lake Baikal. Diversity, 14(1), 55. https://doi.org/10.3390/d14010055