
Near-Term Changes to Reach Scale Habitat Features Following Headwater Stream Restoration in a Southeastern Massachusetts Former Cranberry Bog


Abstract
:1. Introduction
2. Materials and Methods
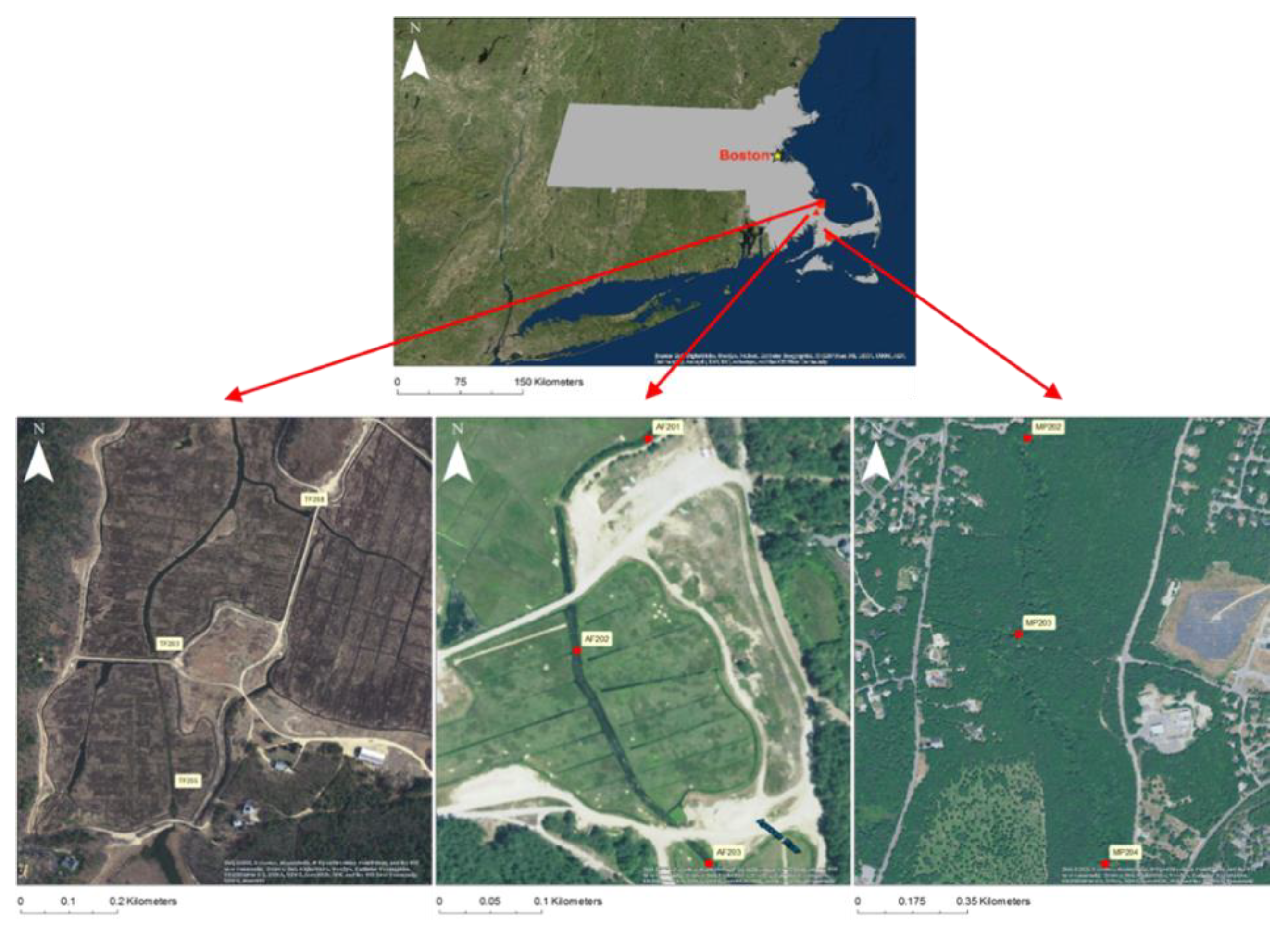
2.1. Study Area
2.2. Restoration Description
2.3. Local-Scale Habitat Analysis
2.4. Statistical Analysis
3. Results
3.1. Habitat Characterization PCA
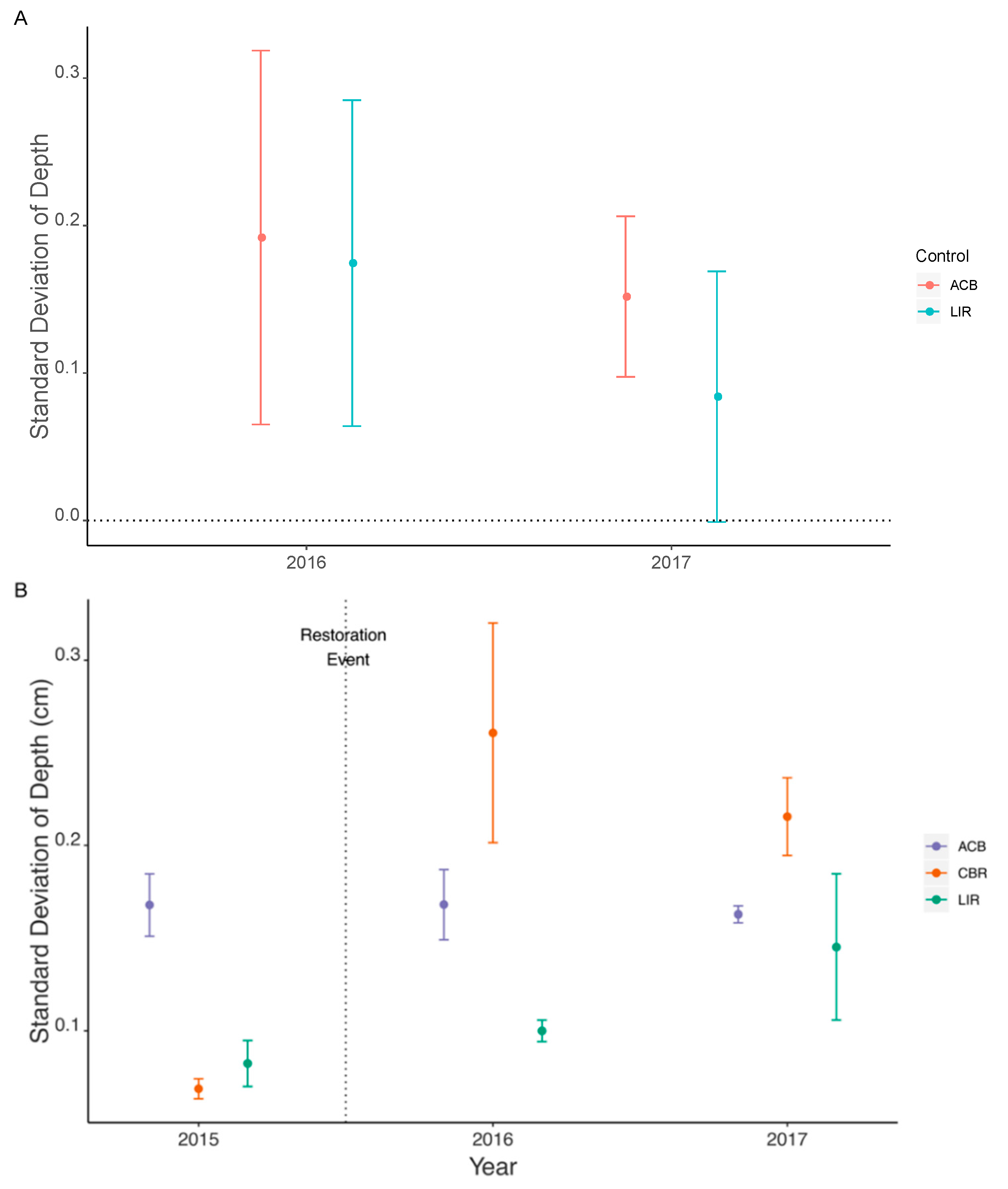
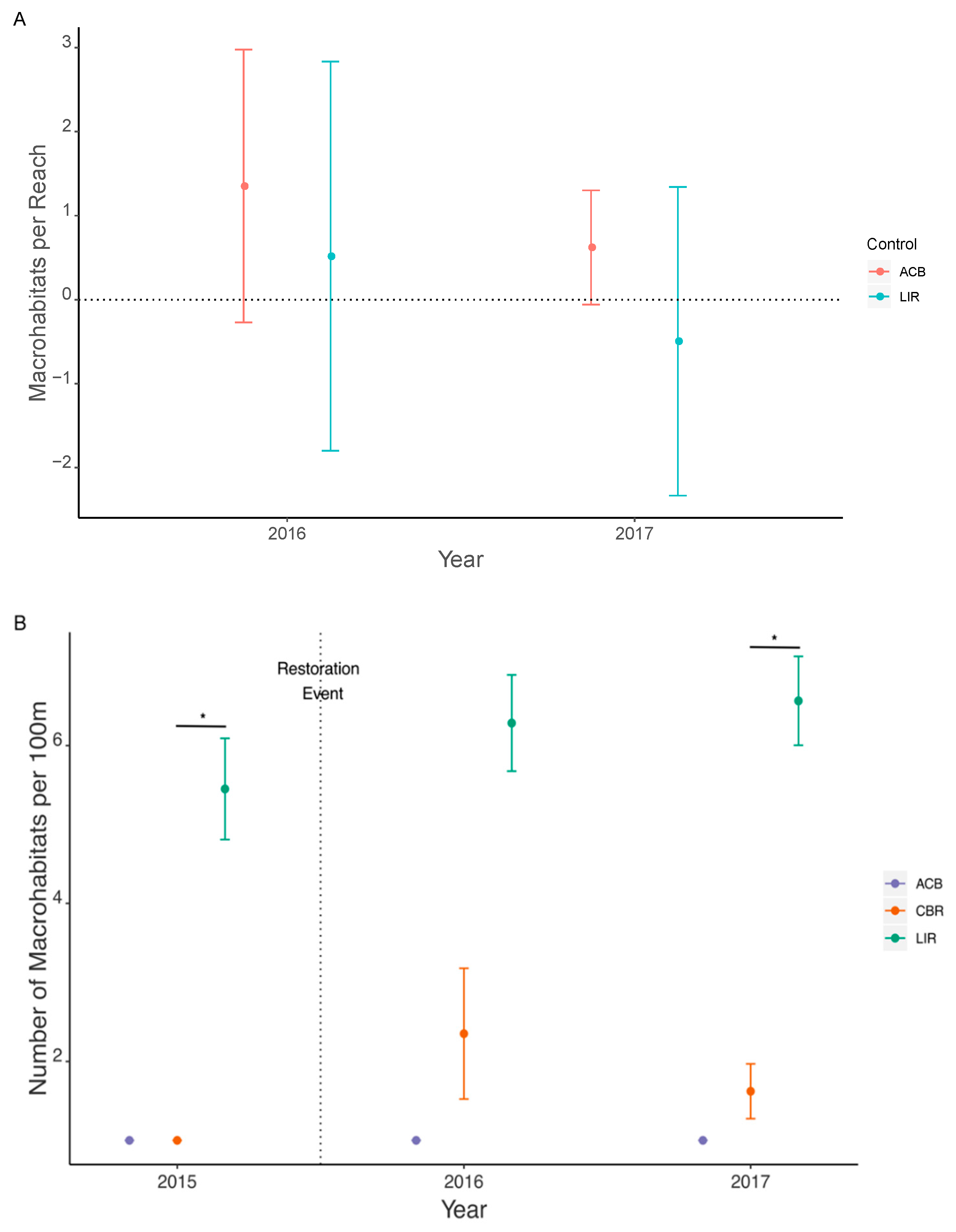
3.2. Habitat Heterogeneity
3.3. Sinuosity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Janse, J.H.; Kuiper, J.J.; Weijters, M.J.; Westerbeek, E.P.; Jeuken, M.H.J.L.; Bakkenes, M.; Alkemade, R.; Mooij, W.M.; Verhoeven, J.T.A. GLOBIO-Aquatic, a Global Model of Human Impact on the Biodiversity of Inland Aquatic Ecosystems. Environ. Sci. Policy 2015, 48, 99–114. [Google Scholar] [CrossRef] [Green Version]
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Biodiversity—Global Biodiversity Scenarios for the Year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Bierschenk, A.M.; Mueller, M.; Pander, J.; Geist, J. Impact of Catchment Land Use on Fish Community Composition in the Headwater Areas of Elbe, Danube and Main. Sci. Total Environ. 2019, 652, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Knott, J.; Mueller, M.; Pander, J.; Geist, J. Effectiveness of Catchment Erosion Protection Measures and Scale-Dependent Response of Stream Biota. Hydrobiologia 2019, 830, 77–92. [Google Scholar] [CrossRef]
- BenDor, T.; Lester, T.W.; Livengood, A.; Davis, A.; Yonavjak, L. Estimating the Size and Impact of the Ecological Restoration Economy. PLoS ONE 2015, 10, e0128339. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.A.; Menninger, H.L.; Bernhardt, E. River Restoration, Habitat Heterogeneity and Biodiversity: A Failure of Theory or Practice? Freshw. Biol. 2010, 55, 205–222. [Google Scholar] [CrossRef]
- Smith, R.F.; Hawley, R.J.; Neale, M.W.; Vietz, G.J.; Diaz-Pascacio, E.; Herrmann, J.; Lovell, A.C.; Prescott, C.; Rios-Touma, B.; Smith, B.; et al. Urban Stream Renovation: Incorporating Societal Objectives to Achieve Ecological Improvements. Freshw. Sci. 2016, 35, 364–379. [Google Scholar] [CrossRef] [Green Version]
- Geist, J.; Hawkins, S.J. Habitat Recovery and Restoration in Aquatic Ecosystems: Current Progress and Future Challenges. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 942–962. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Palmer, M.A. River Restoration: The Fuzzy Logic of Repairing Reaches to Reverse Catchment Scale Degradation. Ecol. Appl. 2011, 21, 1926–1931. [Google Scholar] [CrossRef]
- Palmer, M.A.; Bernhardt, E.S.; Allan, J.D.; Lake, P.S.; Alexander, G.; Brooks, S.; Carr, J.; Clayton, S.; Dahm, C.N.; Follstad Shah, J.; et al. Standards for Ecologically Successful River Restoration. J. Appl. Ecol. 2005, 42, 208–217. [Google Scholar] [CrossRef]
- Palmer, M.A.; Hondula, K.L.; Koch, B.J. Ecological Restoration of Streams and Rivers: Shifting Strategies and Shifting Goals. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 247–269. [Google Scholar] [CrossRef] [Green Version]
- Jähnig, S.C.; Brabec, K.; Buffagni, A.; Erba, S.; Lorenz, A.W.; Ofenböck, T.; Verdonschot, P.F.M.; Hering, D. A Comparative Analysis of Restoration Measures and Their Effects on Hydromorphology and Benthic Invertebrates in 26 Central and Southern European Rivers. J. Appl. Ecol. 2010, 47, 671–680. [Google Scholar] [CrossRef]
- Noss, R.F. Indicators for Monitoring Biodiversity: A Hierarchical Approach. Conserv. Biol. 1990, 4, 355–364. [Google Scholar] [CrossRef]
- Death, R.G.; Joy, M.K. Invertebrate Community Structure in Streams of the Manawatu–Wanganui Region, New Zealand: The Roles of Catchment versus Reach Scale Influences. Freshw. Biol. 2004, 49, 982–997. [Google Scholar] [CrossRef]
- Auerswald, K.; Geist, J. Extent and Causes of Siltation in a Headwater Stream Bed: Catchment Soil Erosion Is Less Important than Internal Stream Processes. Land Degrad. Dev. 2018, 29, 737–748. [Google Scholar] [CrossRef]
- Frissell, C.A.; Liss, W.J.; Warren, C.E.; Hurley, M.D. A Hierarchical Framework for Stream Habitat Classification: Viewing Streams in a Watershed Context. Environ. Manag. 1986, 10, 199–214. [Google Scholar] [CrossRef]
- Poff, N.L. Landscape Filters and Species Traits: Towards Mechanistic Understanding and Prediction in Stream Ecology. J. N. Am. Benthol. Soc. 1997, 16, 391–409. [Google Scholar] [CrossRef]
- Living Observatory. Learning from the Restoration of Wetlands on Cranberry Farmland: Preliminary Benefits Assessment; Living Observatory, Inc.: Boston, MA, USA, 2020; p. 21. Available online: http://www.livingobservatory.org/learning-report (accessed on 27 May 2021).
- ESRI. ArcGIS Desktop Release 10.4.1; Environmental Systems Research Institute: Redlands, CA, USA, 2016. [Google Scholar]
- McCain, M.; Fuller, D.; Decker, L.; Overton, K. Stream Habitat Classification and Inventory Procedures for Northern California; U.S. Dept. of Agriculture, Forest Service, Pacific Southwest Region: San Francisco, CA, USA, 1990; pp. 1–16. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R package version 2.5-3; 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 27 May 2021).
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Christie, A.P.; Amano, T.; Martin, P.A.; Shackelford, G.E.; Simmons, B.I.; Sutherland, W.J. Simple Study Designs in Ecology Produce Inaccurate Estimates of Biodiversity Responses. J. Appl. Ecol. 2019, 56, 2742–2754. [Google Scholar] [CrossRef] [Green Version]
- Roedenbeck, I.; Fahrig, L.; Findlay, C.S.; Houlahan, J.; Jaeger, J.; Klar, N.; Kramer-Schadt, S.; van der Grift, E. The Rauischholzhausen Agenda for Road Ecology. Ecol. Soc. 2007, 12, 11. [Google Scholar] [CrossRef]
- Kristensen, E.A.; Thodsen, H.; Dehli, B.; Kolbe, P.E.Q.; Glismand, L.; Kronvang, B. Comparison of Active and Passive Stream Restoration: Effects on the Physical Habitats. Geogr. Tidsskr. Dan. J. Geogr. 2013, 113, 109–120. [Google Scholar] [CrossRef]
- Sudduth, E.B.; Hassett, B.A.; Cada, P.; Bernhardt, E.S. Testing the Field of Dreams Hypothesis: Functional Responses to Urbanization and Restoration in Stream Ecosystems. Ecol. Appl. 2011, 21, 1972–1988. [Google Scholar] [CrossRef]
- Maloney, K.O.; Weller, D.E. Anthropogenic Disturbance and Streams: Land Use and Land-use Change Affect Stream Ecosystems via Multiple Pathways. Freshw. Biol. 2011, 56, 611–626. [Google Scholar] [CrossRef]
- Waite, I.R. Agricultural Disturbance Response Models for Invertebrate and Algal Metrics from Streams at Two Spatial Scales within the U.S. Hydrobiologia 2014, 726, 285–303. [Google Scholar] [CrossRef]
- Stanley, E.H.; Powers, S.M.; Lottig, N.R. The Evolving Legacy of Disturbance in Stream Ecology: Concepts, Contributions, and Coming Challenges. J. N. Am. Benthol. Soc. 2010, 29, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Bond, N.R.; Lake, P.S. Ecological Restoration and Large-Scale Ecological Disturbance: The Effects of Drought on the Response by Fish to a Habitat Restoration Experiment. Restor. Ecol. 2005, 13, 39–48. [Google Scholar] [CrossRef]
- Friberg, N.; Kronvang, B.; Ole Hansen, H.; Svendsen, L.M. Long-Term, Habitat-Specific Response of a Macroinvertebrate Community to River Restoration. Aquat. Conserv. Mar. Freshw. Ecosyst. 1998, 8, 87–99. [Google Scholar] [CrossRef]
- Kupilas, B.; Hering, D.; Lorenz, A.W.; Knuth, C.; Gücker, B. Hydromorphological Restoration Stimulates River Ecosystem Metabolism. Biogeosciences 2017, 14, 1989–2002. [Google Scholar] [CrossRef] [Green Version]
- Dumke, J.D.; Hrabik, T.R.; Brady, V.J.; Gran, K.B.; Regal, R.R.; Seider, M.J. Channel Morphology Response to Selective Wood Removals in a Sand-Laden Wisconsin Trout Stream. N. Am. J. Fish. Manag. 2010, 30, 776–790. [Google Scholar] [CrossRef]
- Lake, P.S. Ecological Effects of Perturbation by Drought in Flowing Waters. Freshw. Biol. 2003, 48, 1161–1172. [Google Scholar] [CrossRef]
- Brookes, A. Restoring the Sinuosity of Artificially Straightened Stream Channels. Environ. Geol. Water Sci. 1987, 10, 33–41. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Brock, W.A. Rising Variance: A Leading Indicator of Ecological Transition. Ecol. Lett. 2006, 9, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Bureau of Geographic Information. Bureau of Geographic Information (MassGIS); Commonwealth of Massachusetts, Executive Office of Technology and Security Services; 2016. Available online: https://www.mass.gov/info-details/massgis-data-layers (accessed on 27 May 2021).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | PC1 (33.04%) | PC2 (10.03%) |
|---|---|---|
| Length of Habitat | −0.7813222 | 0.56131155 |
| Wetted Width | 0.18867467 | 0.54345261 |
| Maximum Depth | −0.4353481 | −0.303593 |
| Minimum Depth | −0.3508264 | 0.03180419 |
| Standard Deviation of Depth | −0.3143273 | −0.3162003 |
| % Boulder (>30 cm diam.) Substrate | 0.03491382 | 0.10338119 |
| % Cobble (8–30 cm diam.) Substrate | 0.0179058 | −0.1769267 |
| % Gravel (1–8 cm diam.) Substrate | 0.3209406 | 0.09894388 |
| % Sand (0.5 mm–1 cm diam.) Substrate | −0.1085014 | −0.0468564 |
| % Fine (<0.5 mm diam.) Substrate | −0.261565 | −0.0232071 |
| % Undercut Bank | 0.1614075 | −0.1441662 |
| % Large Woody (>30 cm diam.) Debris | 0.00951469 | −0.3575576 |
| % Small Woody (<30 cm diam.) Debris | 0.13093508 | −0.2112439 |
| % In-Stream Terrestrial Vegetation | 0.08023346 | −0.0702898 |
| % Algal Cover | −0.7813222 | 0.56131155 |
| % Rooted Aquatic Vegetation | 0.18867467 | 0.54345261 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCanty, S.T.; Dimino, T.F.; Christian, A.D. Near-Term Changes to Reach Scale Habitat Features Following Headwater Stream Restoration in a Southeastern Massachusetts Former Cranberry Bog. Diversity 2021, 13, 235. https://doi.org/10.3390/d13060235
McCanty ST, Dimino TF, Christian AD. Near-Term Changes to Reach Scale Habitat Features Following Headwater Stream Restoration in a Southeastern Massachusetts Former Cranberry Bog. Diversity. 2021; 13(6):235. https://doi.org/10.3390/d13060235
Chicago/Turabian StyleMcCanty, Sean T., Thomas F. Dimino, and Alan D. Christian. 2021. "Near-Term Changes to Reach Scale Habitat Features Following Headwater Stream Restoration in a Southeastern Massachusetts Former Cranberry Bog" Diversity 13, no. 6: 235. https://doi.org/10.3390/d13060235
APA StyleMcCanty, S. T., Dimino, T. F., & Christian, A. D. (2021). Near-Term Changes to Reach Scale Habitat Features Following Headwater Stream Restoration in a Southeastern Massachusetts Former Cranberry Bog. Diversity, 13(6), 235. https://doi.org/10.3390/d13060235