
Stygobiont Diversity in the San Marcos Artesian Well and Edwards Aquifer Groundwater Ecosystem, Texas, USA


Abstract
:1. Introduction
2. Materials and Methods
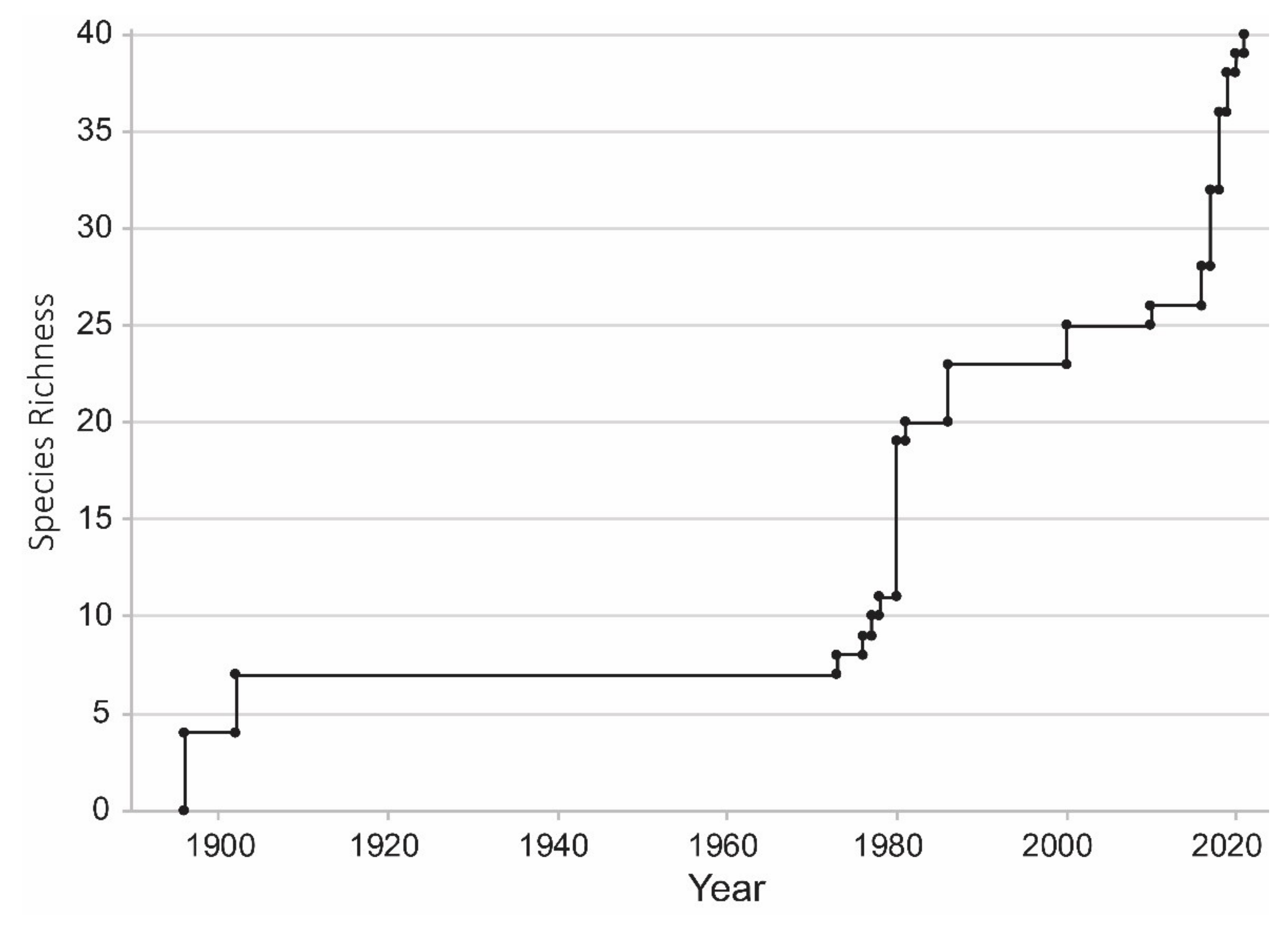
2.1. SMAW Diversity and Temporal Dynamics
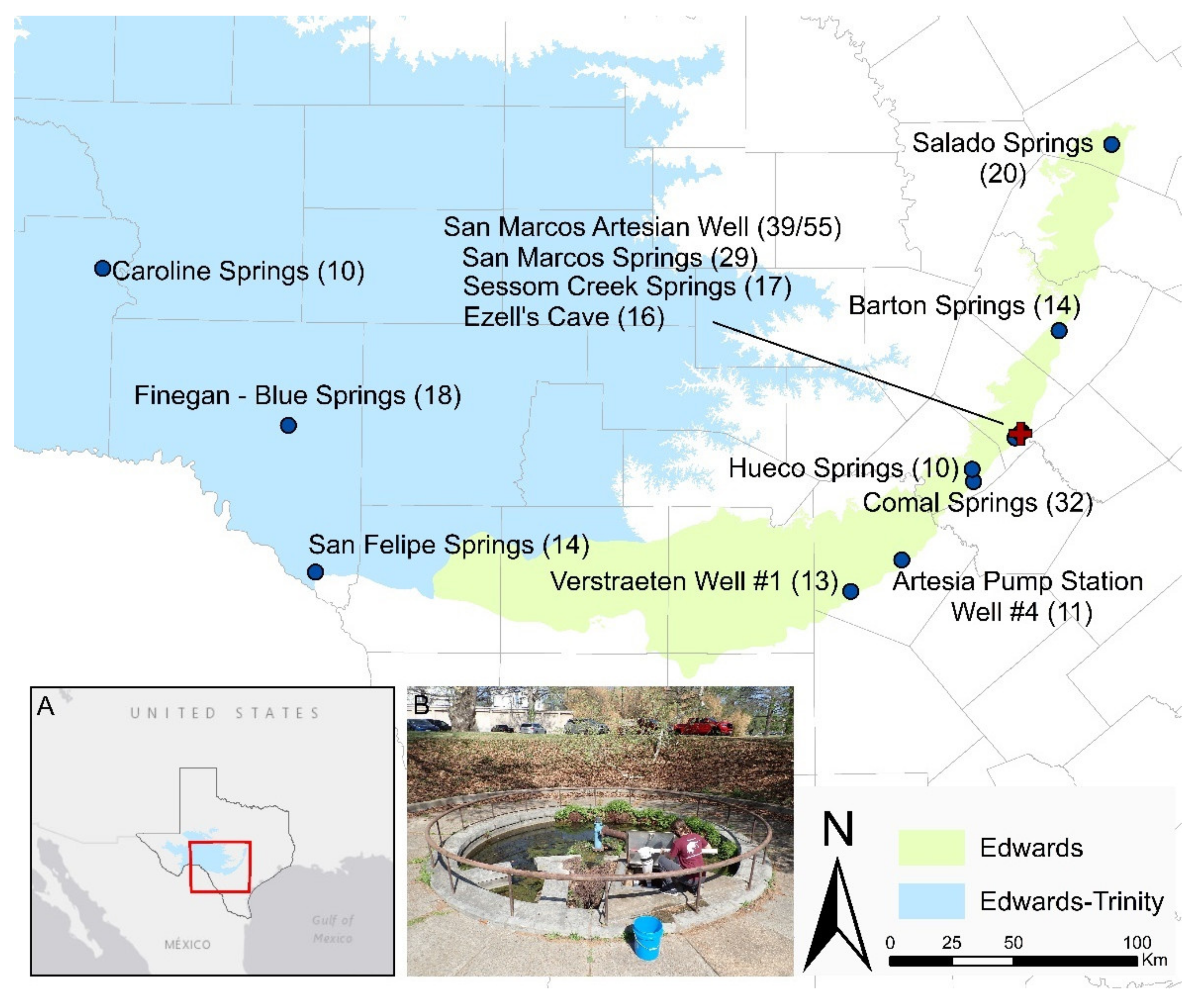
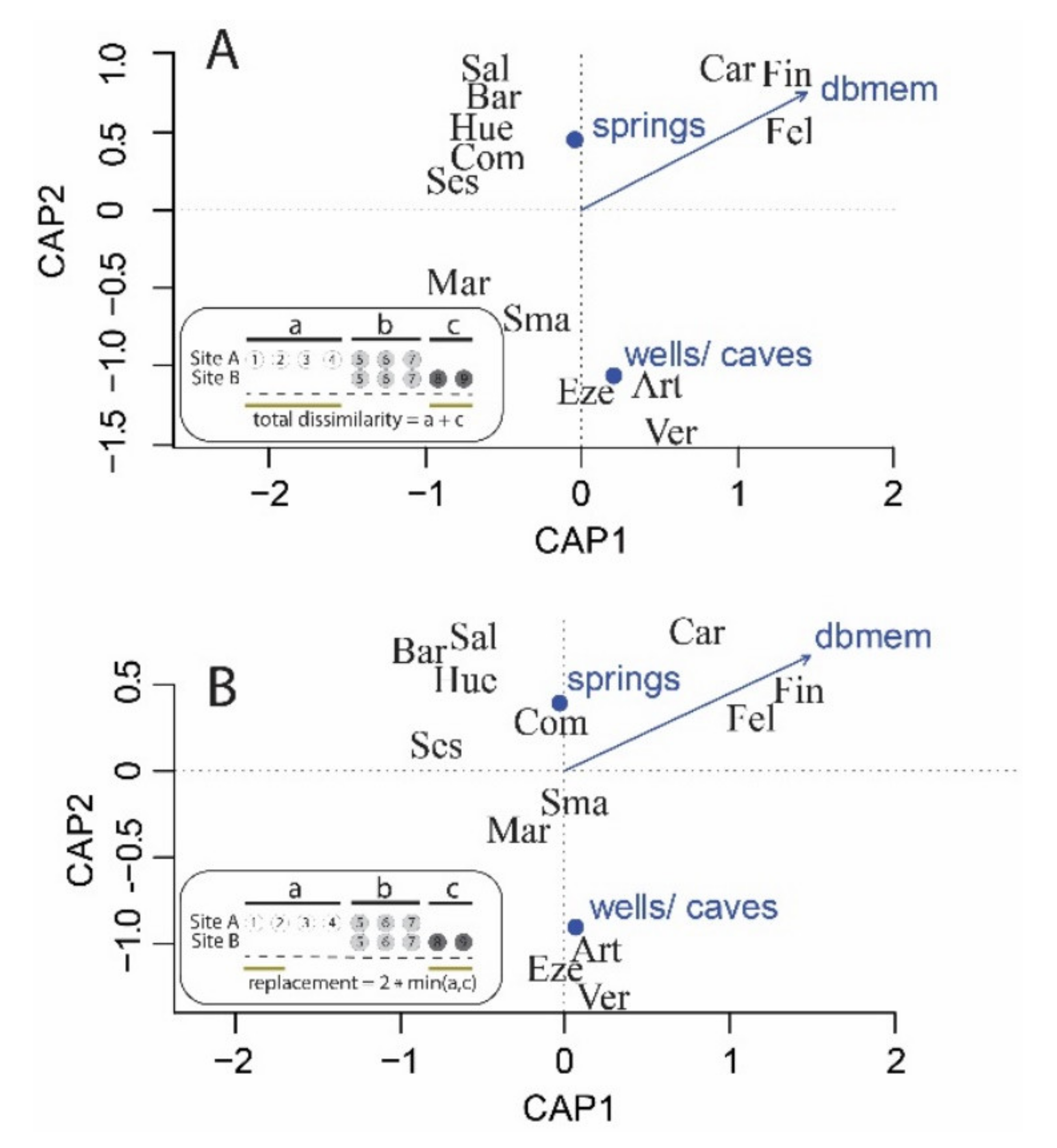
2.2. Edwards Aquifer Groundwater Ecosystem β-Diversity
3. Results
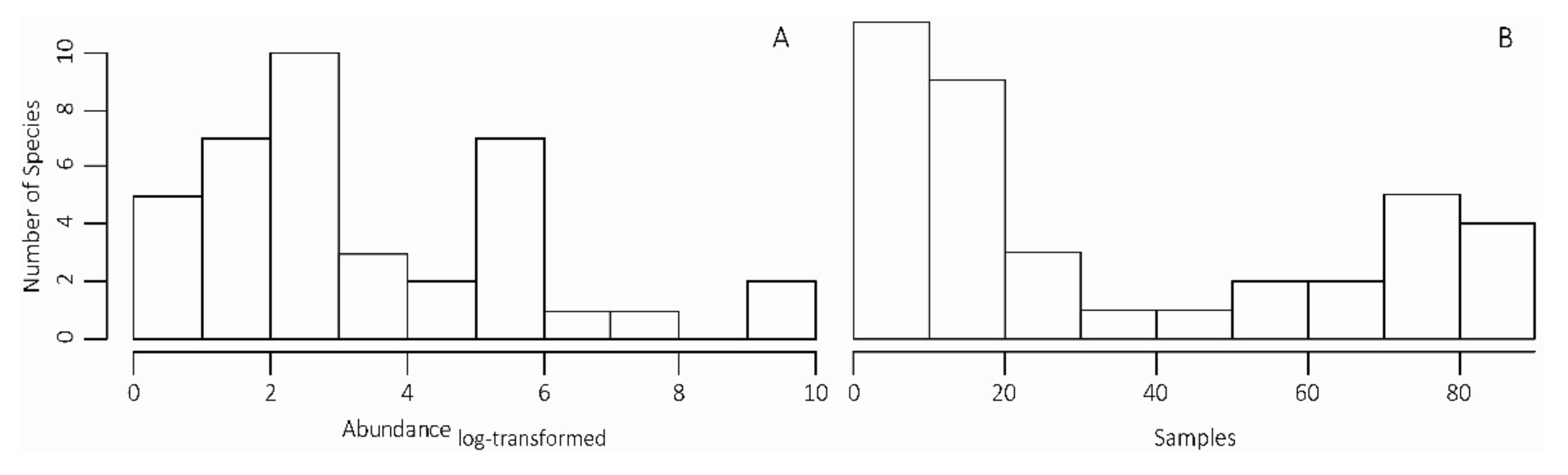
3.1. SMAW Diversity and Temporal Dynamics
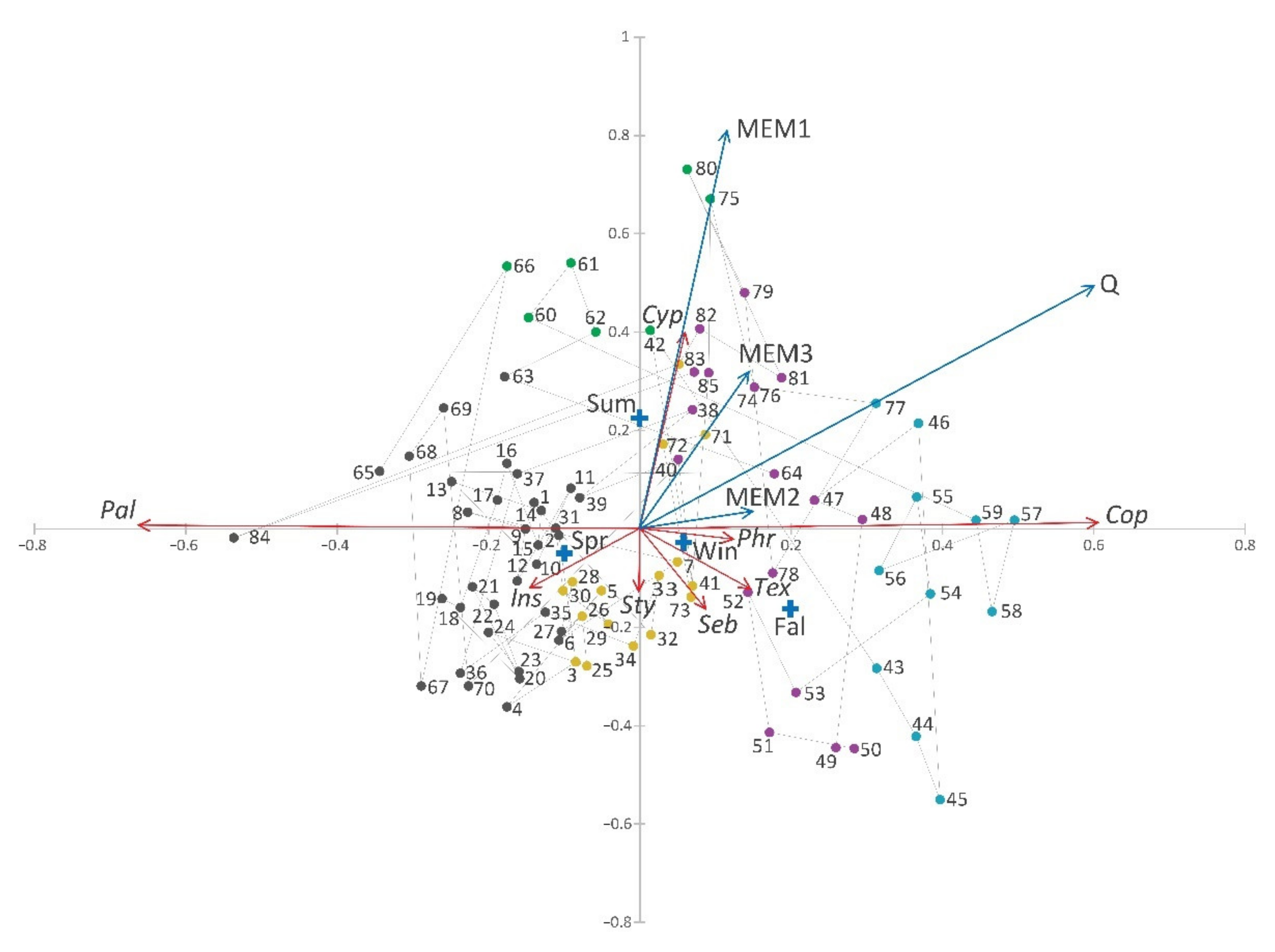
3.2. Edwards Aquifer Groundwater Ecosystem β-Diversity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hinojosa, J.; Green, J.; Estrada, F.; Herrera, J.; Mata, T.; Phan, D.; Tanvir Pasha, A.B.M.; Matta, A.; Johnson, D.; Kapoor, V. Determining the Primary Sources of Fecal Pollution Using Microbial Source Tracking Assays Combined with Land-use Information in the Edwards Aquifer. Water Res. 2020, 184, 116211. [Google Scholar] [CrossRef]
- Longley, G. The Edwards Aquifer: Earth’s Most Diverse Groundwater Ecosystem? Int. J. Speleol. 1981, 11, 123–128. [Google Scholar] [CrossRef]
- Boghici, R. Hydrogeology of Trinity-Edwards Aquifer of Texas and Coahuila in the Border Region. In Aquifers of the Edwards Plateau; Mace, R.E., Angle, E.S., Mullican, W.F., III, Eds.; Texas Water Development Board Report 360: Austin, TX, USA, 2004; pp. 91–114. [Google Scholar]
- Barker, R.A.; Bush, P.W.; Baker, E.T., Jr. Geologic History and Hydrogeologic Setting of the Edwards-Trinity Aquifer System, West-Central Texas. US Geol. Surv. Water Resour. Investig. Rep. 1994, 94, 1–51. [Google Scholar]
- Maclay, R.W. Geology and Hydrology of the Edwards Aquifer in the San Antonio Area, Texas. US Geol. Surv. Water Resour. Investig. Rep. 1995, 95, 1–64. [Google Scholar]
- Sharp, J.M.; Banner, J.L. The Edwards Aquifer: A Resource in Conflict. GSA Today 1997, 7, 1–9. [Google Scholar]
- Schindel, G.M.; Gary, M. Hypogene Processes in the Balcones Fault Zone Segment of the Edwards Aquifer of South-Central Texas. In Hypogene Karst Regions and Caves of the World. Cave and Karst Systems of the World; Klimchouk, A., Palmer, A.N., De Waele, J., Auler, A.S., Audra, P., Eds.; Springer International Publishing AG: Gewerbestrasse, Switzerland, 2017; pp. 511–530. [Google Scholar]
- Hutchins, B.T.; Schwartz, B.F.; Engel, A.S. Environmental Controls on Organic Matter Production and Transport Across Surface-Subsurface and Geochemical Boundaries in the Edwards Aquifer, Texas, USA. Acta Carsologica 2013, 42, 245–259. [Google Scholar] [CrossRef]
- Holsinger, J.R.; Longley, G. The Subterranean Amphipod Crustacean Fauna of an Artesian Well in Texas. Smithson. Contrib. Zool. 1980, 380, 1–62. [Google Scholar] [CrossRef]
- Ogden, A.E.; Quick, R.A.; Rothermel, S.R. Hydrochemistry of the Comal, Hueco, and San Marcos Springs, Edwards Aquifer, Texas. In The Balcones Escarpment: Geology, Hydrology, Ecology and Social Development in Central Texas; Abbott, P.L., Woodruff, C.M., Jr., Eds.; Geological Society of America Annual Meeting: San Antonio, TX, USA, 1986; pp. 115–130. [Google Scholar]
- Culver, D.C.; Sket, B. Hotspots of Subterranean Biodiversity in Caves and Wells. J. Cave Karst Stud. 2000, 62, 11–17. [Google Scholar]
- Ulrich, C.J. A Contribution to the Subterranean Fauna of Texas. Trans. Am. Microsc. Soc. 1902, 23, 83–101. [Google Scholar] [CrossRef]
- Benedict, J.E. Preliminary Descriptions of a New Genus and Three New Species of Crustaceans from an Artesian Well at San Marcos, Texas. Proc. U.S. Nat. Mus. 1896, 18, 615–617. [Google Scholar] [CrossRef]
- Stejneger, L. Description of a New Genus and Species of Blind Tailed Batrachians from the Subterranean Waters of Texas. Proc. U.S. Nat. Mus. 1896, 18, 619–621. [Google Scholar] [CrossRef]
- Holsinger, J.R. Two New Species of the Subterranean Amphipod Genus Mexiweckelia (Gammaridae) from Mexico and Texas, with Notes on the Origin and Distribution of the Genus. Assoc. Mex. Cave Stud. Bull. 1973, 5, 1–12. [Google Scholar]
- Young, F.L.; Longley, G. A New Subterranean Aquatic Beetle from Texas (Coleoptera: Dytiscidae-Hydroporinae). Ann. Entomol. Soc. Am. 1976, 69, 787–792. [Google Scholar] [CrossRef]
- Karnei, H.S., Jr. A Survey of the Subterranean Aquatic Fauna of Bexar County, Texas. Master’s Thesis, Southwest Texas State University, San Marcos, TX, USA, 1978. [Google Scholar]
- Külköylüoğlu, O.; Yavuzatmaca, M.; Akdemir, D.; Schwartz, B.F.; Hutchins, B.T. Description of a New Tribe Cabralcandonini (Candonidae, Ostracoda) from Karst Aquifers in Central Texas, U.S.A. J. Cave Karst Stud. 2019, 81, 136–151. [Google Scholar] [CrossRef]
- Külköylüoğlu, O.; Akdemir, D.; Yavuzatmaca, M.; Schwartz, B.F.; Hutchins, B.T. Cypria lacrima sp. nov. A New Ostracoda (Candonidae, Crustacea) Species from Texas, USA. Zool. Stud. 2017, 56, e15. [Google Scholar] [PubMed]
- Külköylüoğlu, O.; Yavuzatmaca, M.; Akdemir, D.; Schwartz, B.F.; Hutchins, B.T. Lacrimacandona n. gen. (Crustacea: Ostracoda: Candonidae) from the Edwards Aquifer, Texas (USA). Zootaxa 2017, 4277, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Külköylüoğlu, O.; Akdemir, D.; Yavuzatmaca, M.; Schwartz, B.F.; Hutchins, B.T. Rugosuscandona, a New Genus of Candonidae (Crustacea: Ostracoda) from Groundwater Habitats in Texas, North America. Species Divers. 2017, 22, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Külköylüoğlu, O.; Yavuzatmaca, M.; Akdemir, D.; Schwartz, B.F.; Hutchins, B.T. Ufocandona hannaleeae gen. et sp. nov. (Crustacea, Ostracoda) from an Artesian Well in Texas, USA. Eur. J. Taxon. 2017, 372, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, B.F.; Nice, C.; Jenson, A.; Gibson, J.R. Final Report: Molecular and Morphological Analysis of Stygobromus sp. Near San Marcos, TX; Texas Parks and Wildlife Department: Austin, TX, USA, 2016. [Google Scholar]
- Schwartz, B.F.; Nice, C.; Coleman, W. Final Report: Status Assessment and Ecological Characterization of the Texas Troglobitic Water Slate (Lirceolus smithii); Texas Parks and Wildlife Department: Austin, TX, USA, 2018. [Google Scholar]
- Birdwell, J.E.; Engel, A.S. Variability in Terrestrial and Microbial Contributions to Dissolved Organic Matter Fluorescence in the Edwards Aquifer, Central Texas. J. Cave Karst Stud. 2009, 71, 144–156. [Google Scholar]
- Gray, C.J.; Engel, A.S. Microbial Diversity and Impact on Carbonate Geochemistry Across a Changing Geochemical Gradient in a Karst Aquifer. ISME J. 2013, 7, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Hutchins, B.T.; Engel, A.S.; Nowlin, W.H.; Schwartz, B.F. Chemolithoautotrophy Supports Macroinvertebrate Food Webs and Affects Diversity and Stability in Groundwater Communities. Ecology 2016, 97, 1530–1542. [Google Scholar] [CrossRef]
- Hutchins, B.T.; Schwartz, B.F.; Nowlin, W.H. Morphological and Trophic Specialization in a Subterranean Amphipod AssemBlage. Freshw. Biol. 2014, 59, 2447–2461. [Google Scholar] [CrossRef]
- Fišer, C.; Blejec, A.; Trontelj, P. Niche-based Mechanisms Operating within Extreme Habitats: A Case Study of Subterranean Amphipod Communities. Biol. Lett. 2012, 8, 578–581. [Google Scholar] [CrossRef]
- Ercoli, F.; Lefebvre, F.; Delangle, M.; Godé, N.; Caillon, M.; Raimond, R.; Souty-Grosset, C. Differing Trophic Niches of Three French Stygobionts and Their Implications for Conservation of Endemic Stygofauna. Aquat. Conserv. 2019, 29, 2193–2203. [Google Scholar] [CrossRef]
- Chávez-Solís, E.M.; Solís, C.; Simões, N.; Mascaró, M. Distribution Patterns, Carbon Sources and Niche Partitioning in Cave Shrimps (Atyidae: Typhlatya). Sci. Rep. 2020, 10, 12812. [Google Scholar] [CrossRef] [PubMed]
- Dole-Olivier, M.-J.; Marmonier, P.; Creuzé des Châtelliers, M.; Martin, D. Interstitial Fauna Associated with the Alluvial FloodPlains of the Rhône River (France). In Groundwater Ecology; Gibert, J., Danielopol, D.L., Stanford, J.A., Eds.; Academic Press: San Diego, CA, USA, 1994; pp. 313–346. [Google Scholar]
- Pipan, T.; Christman, M.C.; Culver, D.C. Dynamics of Epikarst Communities: Microgeographic Pattern and Environmental Determinants of Epikarst Copepods in Organ Cave, West Virginia. Am. Mid. Nat. 2006, 156, 75–87. [Google Scholar] [CrossRef]
- Gibert, J.; Vervier, P.; Malard, F.; Laurent, R.; Reygrobellet, J.-L. Dynamics of Communities and Ecology of Karst Ecosystems: Example of Three Karsts in Eastern and Southern France. In Groundwater Ecology; Gibert, J., Danielopol, D.L., Stanford, J.A., Eds.; Academic Press: San Diego, CA, USA, 1994; pp. 425–450. [Google Scholar]
- Lorenzo, T.D.; Cipriani, D.; Fiasca, B.; Rusi, S.; Galassi, D.M.P. Groundwater Drift Monitoring as a Tool to Assess the Spatial Distribution of Groundwater Species in Karst Waters. Hydrobiologia 2018, 813, 137–156. [Google Scholar] [CrossRef]
- Opalički Slabe, M. Patterns in Invertebrate Drift from an Alpine Karst Aquifer over a One Year Period. Acta Carsologica 2015, 44, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Gibson, J.R.; Harden, S.J.; Fries, J.N. Survey and Distribution of Invertebrates from Selected Springs of the Edwards Aquifer in Comal and Hays Counties, Texas. Southwest. Nat. 2008, 53, 74–84. [Google Scholar] [CrossRef]
- Trontelj, P.; Douady, C.J.; Fišer, C.; Gibert, J.; Gorički, Š.; Lefébure, T.; Sket, B.; Zakšek, V. A Molecular Test for Cryptic Diversity in Ground Water: How Large are Ranges of Macro-stygobionts? Freshw. Biol. 2009, 54, 727–744. [Google Scholar] [CrossRef]
- Stoch, F.; Galassi, D.M.P. Stygobiontic Crustacean Species Richness: A Question of Numbers, a Matter of Scale. Hydrobiologia 2010, 653, 217–234. [Google Scholar] [CrossRef]
- Trontelj, P. Structure and Genetics of Cave Populations. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 269–296. [Google Scholar]
- Pipan, T.; Culver, D.C.; Papi, F.; Kozel, P. Partitioning Diversity in Subterranean Invertebrates: The Epikarst Fauna of Slovenia. PLoS ONE 2018, 13, e0195991. [Google Scholar] [CrossRef]
- Trajano, E.; de Carvalho, M.R. Towards a Biologically Meaningful Classification of Subterranean Organisms: A Critical Analysis of the Schiner-Racovitza System from a Historical Perspective, Difficulties of Its Application and Implications for Conservation. Subterr. Biol. 2017, 22, 1–26. [Google Scholar] [CrossRef]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer Science+Business Media, LLC.: New York, NY, USA, 2011. [Google Scholar]
- Kollaus, K.A.; Bonner, T.R. Habitat Associations of a Semi-arid Fish Community in a Karst Spring-fed Stream. J. Arid Environ. 2012, 76, 72–79. [Google Scholar] [CrossRef]
- U.S. Geological Survey. National Water Information System Data Available on the World Wide Web (USGS Water Data for the Nation). Available online: https://waterdata.usgs.gov/tx/nwis/uv/?site_no=08170500&PARAmeter_cd=00065,00060 (accessed on 19 November 2020). [CrossRef]
- Zappitello, S.J.; Johns, D.A.; Hunt, B.B. Summary of Groundwater Tracing in the Barton Springs Edwards Aquifer from 1996 to 2017. In Report # DR-19-04; Barton Springs Edwards Aquifer Conservation District: Austin, TX, USA, 2019. [Google Scholar]
- Legendre, P.; Gauthier, O. Statistical Methods for Temporal and Space-time Analysis of Community Composition Data. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, P.; Rigal, F.; Carvalho, J.C. BAT–Biodiversity Assessment Tools, an R Package for the Measurement and Estimation of Alpha and Beta Taxon, Phylogenetic and Functional Diversity. Methods Ecol. Evol. 2015, 6, 232–236. [Google Scholar] [CrossRef]
- Jaume, D. Global Diversity of Spelaeogriphaceans & Thermosbaenaceans (Crustacea; Spelaeogriphacea & Thermosbaenacea) in Freshwater. Hydrobiologia 2008, 595, 219–224. [Google Scholar]
- Sket, B. Hirudinea. In Stygofauna Mundi; Botosaneanu, L., Ed.; E.J. Brill & Dr. W. Backhuys: Leiden, The Netherland, 1986; pp. 250–253. [Google Scholar]
- Kanda, K.; Gomez, R.A.; Driesche, R.V.; Miller, K.B.; Maddison, D.R. Phylogenetic Placement of the Pacific Northwest Subterranean Endemic Diving Beetle Stygoporus oregonensis Larson & LaBonte (Dytiscidae, Hydroporinae). Zootaxa 2016, 632, 75–91. [Google Scholar]
- Moravec, F.; Huffman, D.G. Three New Helminth Species from Two Endemic Plethodontid Salamanders, Typhlomolge rathbuni and Eurycea nana, in Central Texas. Folia Parasitol. 2000, 47, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Kenk, R. Freshwater Triclads (Turbellaria) of North America, IX: The Genus Sphalloplana. Smithson. Contrib. Zool. 1977, 246, 1–38. [Google Scholar] [CrossRef]
- Alvear, D.; Diaz, P.H.; Gibson, J.R.; Jones, M.; Perez, K.E. An Unusually Sculptured New Species of Phreatodrobia Hershler & Longley (Mollusca: Caenogastropoda: Cochliopidae) from Central Texas. Zootaxa 2020, 4810, 143–152. [Google Scholar]
- Hershler, J.R.; Longley, G. Phreatic Hydrobiids (Gastropoda: Prosobranchia) from the Edwards (Balcones Fault Zone) Aquifer Region, South-Central Texas. Malacologia 1986, 27, 127–172. [Google Scholar]
- Külköylüoğlu, O. A New Genus and Species in the Ostracod Family Candonidae (Crustacea: Ostracoda) from Texas, USA. J. Nat. Hist. 2018, 52, 1295–1310. [Google Scholar] [CrossRef]
- Camacho, A.I.; Hutchins, B.; Schwartz, B.F.; Dorda, B.A.; Casado, A.; Rey, I. Description of a New Genus and Species of Bathynellidae (Crustacea: Bathynellacea) from Texas Based on Morphological and Molecular Characters. J. Nat. Hist. 2018, 52, 29–51. [Google Scholar] [CrossRef]
- Hutchins, B.T.; Schwartz, B.F.; Coleman, W.T. Three New Microcerberids (Isopoda, Microcerberidae) from Subterranean Freshwater Habitats in Texas, U.S.A. (In review)
- Schwartz, B.F.; Hutchins, B.T.; Schwartz, Z.G.; Hess, A.J.; Bonett, R.M. Cirolanides wassenichae sp. nov., a Freshwater, Subterranean Cirolanidae (Isopoda, Cymothoida) with Additional Records of Other Species from Texas, United States. Zootaxa 2019, 4543, 498–514. [Google Scholar] [CrossRef]
- Zara Environmental LLC. Final Report: Hays County Karst Invertebrate Distribution and Cave Development; Texas Parks and Wildlife Department: Austin, TX, USA, 2010. [Google Scholar]
- World of Copepods. Available online: www.marinespecies.org/copepoda (accessed on 11 May 2021).
- Carvalho, J.C.; Cardoso, P.; Borges, P.A.V.; Schmera, D.; Podani, J. Measuring Fractions of Beta Diversity and Their Relationships to Nestedness: A Theoretical and Empirical Comparison of Novel Approaches. Oikos 2013, 122, 825–834. [Google Scholar] [CrossRef] [Green Version]
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats, 2nd ed.; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Cardinale, B.J.; Bennett, D.M.; Nelson, C.E.; Gross, K. Does Productivity Drive Diversity or Vice Versa? A Test of the Multivariate Productivity-diversity Hypothesis in Streams. Ecology 2009, 90, 1227–1241. [Google Scholar] [CrossRef]
- Eme, D.; Zagmajster, M.; Fišer, C.; Galassi, D.; Marmonier, P.; Stoch, F.; Cornu, J.-F.; Oberdorff, T.; Malard, F. Multi-causality and Spatial Non-stationarity in the Determinants of Groundwater Crustacean Diversity in Europe. Ecography 2015, 38, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Culver, D.C.; Deharveng, L.; Bedos, A.; Lewis, J.J.; Madden, M.; Reddell, J.R.; Sket, B.; Trontelj, P.; White, D. The Mid-latitude Biodiversity Ridge in Terrestrial Cave Fauna. Ecography 2006, 29, 120–128. [Google Scholar] [CrossRef]
- Toomey, R.S., III; Blum, M.D.; Valastro, S., Jr. Late Quaternary Climates and Environments of the Edwards Plateau, Texas. Glob. Planet. Chang. 1993, 7, 299–320. [Google Scholar] [CrossRef]
- Cardoso, P.; Borges, P.A.V.; Veech, J.A. Testing the Performance of Beta Diversity Measures Based on Incidence Data: The Robustness to Undersampling. Divers. Distrib. 2009, 15, 1081–1090. [Google Scholar] [CrossRef] [Green Version]
- Malard, F.; Boutin, C.; Camacho, A.I.; Ferreira, D.; Michel, G.; Sket, B.; Stoch, F. Diversity Patterns of Stygobiotic Crustaceans Across Multiple Spatial Scales in Western Europe. Freshw. Biol. 2009, 54, 756–776. [Google Scholar] [CrossRef]
- Cáceres, M.D.; Coll, L.; Legendre, P.; Allen, R.B.; Wiser, S.K.; Fortin, M.-J.; Condit, R.; Hubbell, S. Trajectory Analysis in Community Ecology. Ecol. Monogr. 2019, 89, e01350. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S.; Cardoso, P.; Culver, D.C.; Deharveng, L.; Ferreira, R.L.; Fišer, C.; Galassi, D.M.P.; Griebler, C.; Halse, S.; Humphreys, W.H.; et al. Scientists’ Warning on the Conservation of Subterranean Ecosystems. Bioscience 2019, 69, 641–650. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Class | Order | Family | Species | Reference |
|---|---|---|---|---|---|
| Nematoda | Enoplea | Trichocephalida | Capillariidae | Amphibiocapillaria texensis Moravec and Huffman, 2000† | [52] |
| Acanthocephala | Eoacanthocephala | Neoechinorhynchida | Dendronucleatidae | Dendronucleata americana Moravec and Huffman, 2000†§ | [52] |
| Platyhelminthes | Rhabditophora | Tricladida | Kenkiidae | Sphalloplana mohri Hyman, 1938 | [53] |
| Annelida | Clitellata | Arhynchobdellida | Erpobdellidae | Erpobdella sp. | [2] |
| Lumbriculida | Lumbriculidae | gen. sp. undet. | Steve Fend, pers. comm. | ||
| Mollusca | Gastropoda | Littorinimorpha | Cochliopidae | Phreatodrobia micra (Pilsbry and Ferriss, 1906) | [54] |
| Phreatodrobia nugax (Pilsbry and Ferriss, 1906) | [55] | ||||
| Phreatodrobia plana Hershler and Longley, 1986 | [55] | ||||
| Phreatodrobia rotunda Hershler and Longley, 1986 | [55] | ||||
| Arthropoda | Ostracoda | Podocopida | Candonidae | Cabralcandona mixoni Külköylüoğlu, Yavuzatmaca, Akdemir, Schwartz and Hutchins, 2019†§ | [18] |
| Candonopsis sp. | Okan Külköylüoğlu pers. comm. | ||||
| Comalcandona sp. | Okan Külköylüoğlu pers. comm. | ||||
| Cypria lacrima Külköylüoğlu, Akdemir, Yavuzatmaca, Schwartz and Hutchins, 2017†§ | [19] | ||||
| Lacrimacandona wisei Külköylüoğlu, Yavuzatmaca, Akdemir, Schwartz and Hutchins, 2017†§ | [20] | ||||
| Namiotkocypria haysensis Külköylüoğlu, 2018†§ | [56] | ||||
| Rugosuscandona scharfi Külköylüoğlu, Akdemir, Yavuzatmaca, Schwartz and Hutchins, 2017† | [21] | ||||
| Ufocandona hannaleeae Külköylüoğlu, Yavuzatmaca, Akdemir, Schwartz and Hutchins, 2017†§ | [22] | ||||
| Schornikovcandona bellensis Külköylüoğlu, Yavuzatmaca, Akdemir, Diaz and Gibson, 2017 | Okan Külköylüoğlu pers. comm. | ||||
| Darwinulidae | Darwinula sp. | Okan Külköylüoğlu pers. comm. | |||
| Vestalenula sp. | Okan Külköylüoğlu pers. comm. | ||||
| Hexanauplia | Cyclopoida | Cyclopidae | Cyclops cavernarum Ulrich, 1902†§ | [12] | |
| Cyclops learii Ulrich, 1902†§ | [12] | ||||
| Harpacticoida | gen. sp. undet. | Diego Figueroa, pers. comm. | |||
| Malacostraca | Bathynellacea | Bathynellidae | Hobbsinella edwardensis Camacho, Hutchins, Schwartz, Dorda, Casado and Rey, 2017† | [57] | |
| Thermosbaenacea | Monodellidae | Tethysbaena texana (Maguire, 1965) | [17] | ||
| Isopoda | Microcerberidae | Microcerberidae sp. 1 | [58] | ||
| Microcerberidae sp. 2 | pers. obs. | ||||
| Asellidae | Lirceolus hardeni Lewis and Bowman, 1996 | [24] | |||
| Lirceolus pilus (Steeves, 1968) | [24] | ||||
| Lirceolus smithii (Ulrich, 1902)† | [12] | ||||
| Lirceolus sp. | William Coleman pers. comm. | ||||
| Cirolanidae | Cirolanides texensis Benedict, 1896† | [13] | |||
| Cirolanides wassenichae Schwartz, Hutchins, Schwartz, Hess and Bonett, 2019† | [59] | ||||
| Cirolanides sp. | pers. obs. | ||||
| Amphipoda | Artesiidae | Artesia subterranea Holsinger, 1980† | [9] | ||
| Crangonyctidae | Stygobromus bifurcatus (Holsinger, 1967) | [23] | |||
| Stygobromus flagellatus (Benedict, 1896)† | [13] | ||||
| Stygobromus longipes (Holsinger, 1966) | [23] | ||||
| Stygobromus russelli (Holsinger, 1967) | [9] | ||||
| Hadziidae | Allotexiweckelia hirsuta Holsinger, 1980† | [9] | |||
| Holsingerius samacos (Holsinger, 1980)† | [9] | ||||
| Texiweckelia texensis (Holsinger, 1973)† | [15] | ||||
| Texiweckeliopsis insolita (Holsinger, 1980)† | [9] | ||||
| Parabogidiellidae | Parabogidiella americana Holsinger, 1980† | [9] | |||
| Parabogidiella sp. Holsinger, 1980 | [9] | ||||
| Seborgiidae | Seborgia relicta Holsinger, 1980† | [9] | |||
| Decapoda | Palaemonidae | Calathaemon holthuisi (Strenth, 1976) | [60] | ||
| Palaemon antrorum (Benedict, 1896)† | [13] | ||||
| Arachnida | Trombidiformes | Chappuisididae | Chappuisides sp. | Ian Smith, in prep. | |
| Uchidastygacarus sp. | Ian Smith, in prep. | ||||
| Halacaridae | gen. sp. undet. | Ian Smith, in prep. | |||
| Mideopsidae | gen. sp. undet. | Ian Smith, in prep. | |||
| Nudomideopsidae | Allomideopsis wichitaensis (Smith, 1990) | Ian Smith, in prep. | |||
| Insecta | Coleoptera | Dytiscidae | Haideoporus texanus Young and Longley, 1976† | [16] | |
| Chordata | Amphibia | Urodela | Plethodontidae | Eurycea rathbuni (Stejneger, 1896)† | [14] |
| Positive Association | Negative Association | |||||
| Hydraulic/ environmental effect | → | Community response | Hydraulic/ environmental effect | → | Community response | |
| Increased flow | Activates intermittent/ alternative flowpaths and hydraulic connections | → | Animals transported from typically less-connected aquifer areas to sampling location | Increased flow within conduit | → | Animals within conduit seek refuge in adjacent fracture network |
| Increased hydraulic gradient from fracture network to conduit network | → | Animals transported to sampling location from adjacent fracture network | ||||
| Increased sediment transport and conduit boundary velocities | → | Dislodges benthic/ interstitial organisms already within sampling area | Changes in nutrient availability, community composition, and geochemistry | → | Direct and indirect demographic (e.g., reproduction) or species interaction (e.g., ecologic release) effects | |
| Changes in nutrient availability, community composition, and geochemistry | → | Direct and indirect demographic (e.g., reproduction) or species interaction (e.g., ecologic release) effects | ||||
| Decreased flow | Decreased flow within conduit | → | Animals within fracture network move into conduit | Changes in nutrient availability, community composition, and geochemistry | → | Direct and indirect demographic (e.g., reproduction) or species interaction (e.g., ecologic release) effects |
| Changes in nutrient availability, community composition, and geochemistry | → | Direct and indirect demographic (e.g., reproduction) or species interaction (e.g., ecologic release) effects | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hutchins, B.T.; Gibson, J.R.; Diaz, P.H.; Schwartz, B.F. Stygobiont Diversity in the San Marcos Artesian Well and Edwards Aquifer Groundwater Ecosystem, Texas, USA. Diversity 2021, 13, 234. https://doi.org/10.3390/d13060234
Hutchins BT, Gibson JR, Diaz PH, Schwartz BF. Stygobiont Diversity in the San Marcos Artesian Well and Edwards Aquifer Groundwater Ecosystem, Texas, USA. Diversity. 2021; 13(6):234. https://doi.org/10.3390/d13060234
Chicago/Turabian StyleHutchins, Benjamin T., J. Randy Gibson, Peter H. Diaz, and Benjamin F. Schwartz. 2021. "Stygobiont Diversity in the San Marcos Artesian Well and Edwards Aquifer Groundwater Ecosystem, Texas, USA" Diversity 13, no. 6: 234. https://doi.org/10.3390/d13060234
APA StyleHutchins, B. T., Gibson, J. R., Diaz, P. H., & Schwartz, B. F. (2021). Stygobiont Diversity in the San Marcos Artesian Well and Edwards Aquifer Groundwater Ecosystem, Texas, USA. Diversity, 13(6), 234. https://doi.org/10.3390/d13060234