1. Introduction
Caves are mostly associated with sedimentary terrains subjected to karst processes, mainly in carbonate rocks but also in gypsum, rock salt, conglomerate, etc. [
1]. These solution caves are the result of a chemical erosive process that progresses slowly (millions of years) and can result in important cavity dimensions, such as the 667 km of Mammoth Cave (USA) or the 371 km of the Sac Actun/Dos Ojos System (Mexico) [
2]. Volcanic terrains typically lack limestone and other sedimentary rocks, at least with sufficient thickness and age to originate karst caves, with a few exceptions when the reef limestone has been uplifted and later karstified [
3]. However, other kinds of caves are frequent in volcanic terrains, formed in a completely different way and different type of rocks, usually fluid basalts but sometimes also phonolites [
4]. Volcanic caves, most of them lava tubes, have a constructive rather than erosive origin, are exactly the same age as their host rock, and become destroyed in a very short geological time span compared to karst ones [
5]. This, together with the fact that the whole volcanic areas have much smaller extension than karstifiable sedimentary rocks, means that volcanic caves are much less abundant worldwide and, in general, of considerably smaller dimensions than karst ones [
6]. It had been claimed that volcanic caves did not have any adapted cave-dwelling fauna [
7]. However, this was later disproved for Hawaiian lava tubes and later for many other tropical and non-tropical volcanic areas [
5,
8,
9,
10,
11]. Conversely, some lava tubes are particularly rich in cave-adapted fauna and feature prominently in global cave rankings. Thus, among the 27 caves richest in cave-adapted species worldwide in 2019, 23 were karst caves, and only four were lava tubes: one in Australia (Bayliss Cave) and three on the Canary Islands [
12]. However, the lava tubes are surprisingly well ranked: Jameos del Agua lava tube for groundwater animals, and Cueva de Felipe Reventón and Cueva del Viento for underground terrestrial richness. Cueva de Felipe Reventón and Cueva del Viento are lava tubes located in Tenerife, Canary Islands, and have been the most attractive caves for scientific studies from this archipelago, together with the anchialine part of Cueva de la Corona lava tube on the island of Lanzarote [
13,
14,
15,
16,
17].
2. Location and General Characteristics
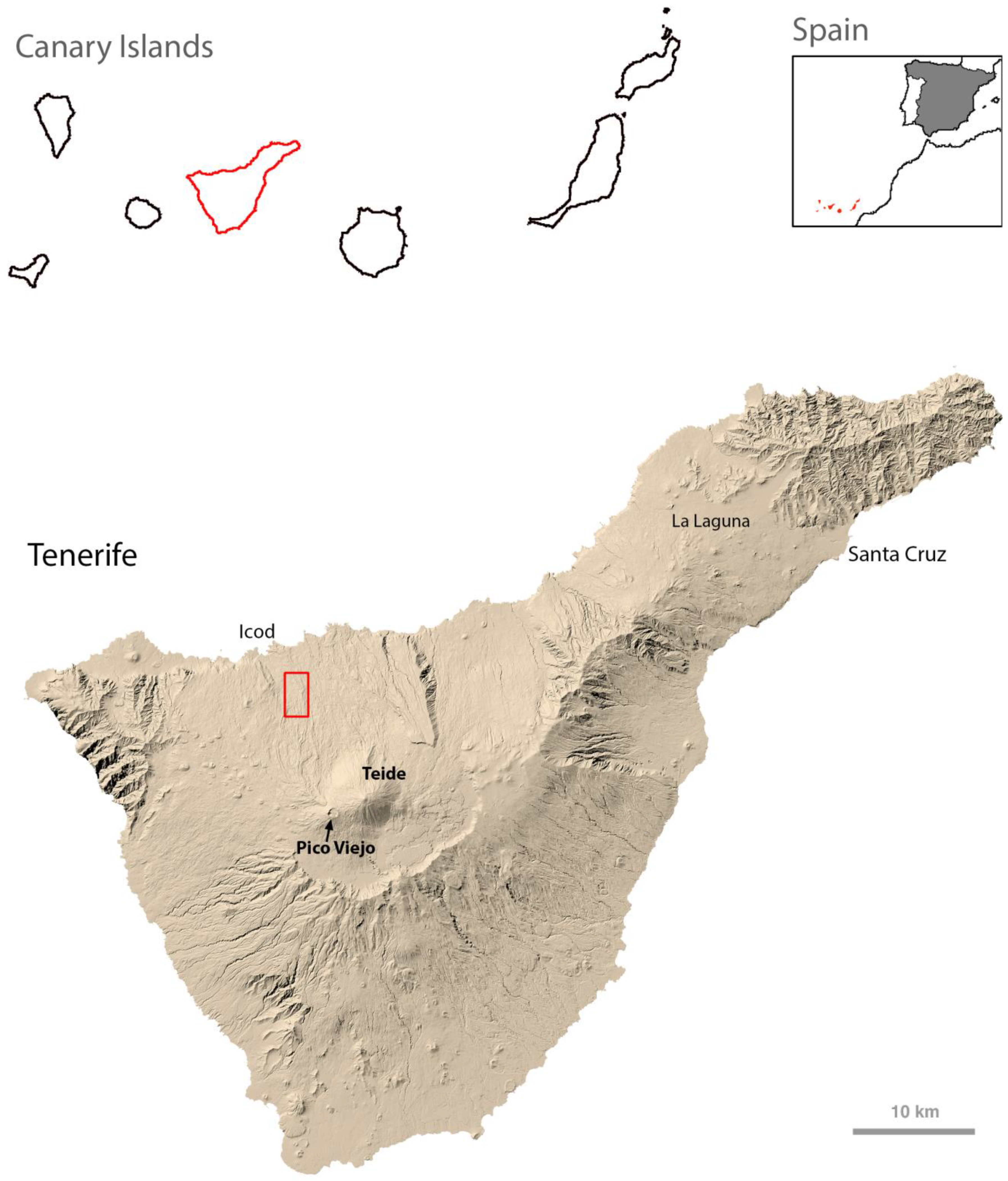
The Canary archipelago is located in the eastern Atlantic between 27°37′ to 29°25′ N and 13°20′ to 18°10′ W, off the coast of south Morocco. It comprises eight main inhabited islands of volcanic origin, with geological ages as emerged land decreasing from 21 Ma for the eastern Fuerteventura to 1 Ma for the westernmost El Hierro. A remarkable variety of volcanic rocks, type of eruptions, local climates and landscapes is found across this archipelago. Tenerife is in the center of the island chain, with an intermediate age of 12 Ma [
18]. It is the largest (2034 km
2), the highest (3714 m at Teide Peak) and the most diverse island both in terms of ecosystems and of plant and animal species [
19]. The abundance of subrecent and recent basaltic lavas allows the existence of many lava tube caves and other subterranean environments that harbor a rich cave-adapted fauna [
20]. The highest density of lava tubes is found around Icod de Los Vinos, in the northwest of the island, an area covered by the lavas of Teide and Pico Viejo stratovolcanos from the central-western part of the island [
21] (
Figure 1). In these lava flows is the Cueva del Viento System, which includes Cueva de Felipe Reventón and Cueva del Viento, as well as other minor lava tubes. The two main caves are situated close to each other with entrances between 572 and 847 m a.s.l.; but the extreme lowest and highest internal galleries of Cueva del Viento are at 395 and 880 m, respectively.
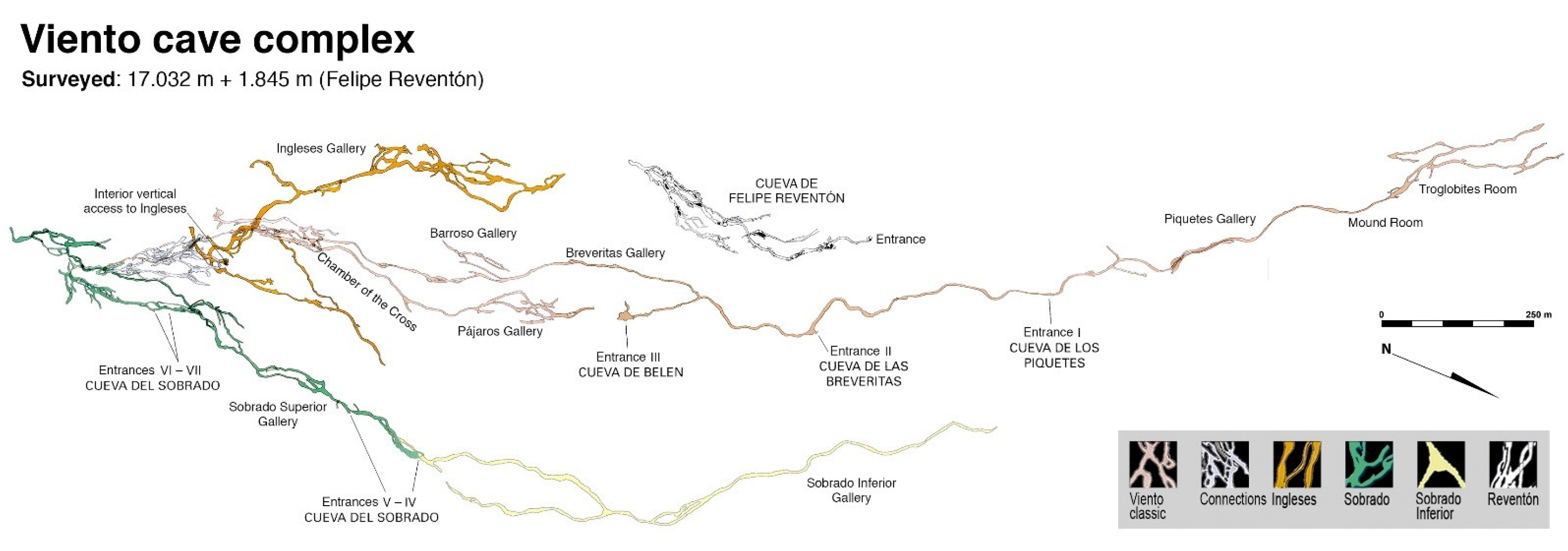
Both caves originated in the same lava flow and probably simultaneously, but they are not connected and have traditionally been considered as independent caves. The maps of their surveys show that Cueva de Felipe Reventón is roughly aligned with one branch of Cueva del Viento, as if it were an extension of the latter (
Figure 2). The corresponding ends facing each other are blocked by solidified lava siphons, leaving an unknown 90 m long stretch that had to connect the two caves during their formation. The access between caves is impossible for humans, but not for small animals through the cracks of the same lava flow. From the biospeleological point of view, they would be part of the same Cueva del Viento System.
The climate of this area is mild (annual mean 15.1 °C) and moderately humid by the island standards. However, the potential vegetation is not a laurel forest as in other places at the same altitude and orientation, but a mixed pine forest dominated by the Canary pine (Pinus canariensis) with big heathers (Erica arborea) and some broadleaf trees such as wax myrtle (Morella faya), laurel (Laurus novocanariensis) and Canary holly (Ilex canariensis). This area is moderately inhabited, with many scattered houses and cultivation fields but never with a compact urban structure, and only the upper part of the cave system (Cueva del Sobrado) develops under a fairly well-preserved pine forest.
Cueva del Viento has five easily practicable entrances and two small skylights, all originated by roof collapses. Two of these openings are within a public property belonging to the Cabildo de Tenerife (the local government of the island), the cave sector in between being a show cave; the other five openings are in private land. Four different names are used for the main entries and their corresponding cave sectors: Cueva Piquetes, Cueva de Breveritas, Galería Belén and Cueva del Sobrado, each with their own entrance though Sobrado has four (
Figure 2). Piquetes and Breveritas are artificially separated by a cellar of a house; Belén is a short sector of Breveritas isolated from the rest by two stone collapses; and the formerly considered as independent Cueva del Viento and Cueva del Sobrado are actually connected by a narrow natural passage, and now considered as a single cave. For study purposes the whole cave has been divided into the following sections, from the lowest to the highest and using entrances or inner collapses as limits: Piquetes, Breveritas Inferior, Breveritas Superior, Belén, Ingleses, Sobrado Inferior and Sobrado Superior. Breveritas Inferior, Breveritas Superior and Ingleses would be popularly known as Cueva del Viento proper, with a single entrance called Breveritas. All the main galleries of Cueva del Viento have reasonable large dimensions and are easily accessible (
Figure 3).
The mapped extent of Cueva de Felipe Reventón is 1845 m so far measured, but the complete survey has never been finished and it is probably something over 3000 m in length [
24]. The only entrance to Felipe Reventón is in a public property also belonging to the Cabildo de Tenerife, but most of the upper part of the cave lies under private land with a few houses. This entrance is at 628 m a.s.l. just at the lower end of the cave, and no different sector names are used for partial galleries. It is a rather labyrinthine system of interconnected galleries with a relatively short distance (438 m) between upper and lower extremes (
Figure 2), and the internal dimensions are generally somewhat smaller than the main galleries of Cueva del Viento.
3. Discovery, Exploration and Scientific Study
The first detailed description of a volcanic cave in the Canary Islands was in 1776 by Bethencourt de Castro brothers on Cueva de San Marcos, in the same Pico Viejo lava flow as the Viento System but next to the coast [
21]. The objective of the expedition was to describe and survey the cave, and “to continue to the summit …, where some say it has communication with another cave called del Viento”, then already known by this name.
The first modern expedition was carried out in 1969 by the local Sección de Exploraciones Vulcanoespeleológicas de La Guancha (SEVG). A year later they completed the topography of what they considered the entire cave, but the map was not published. Members of the Club Montañés de Barcelona contacted SEVG and published a geological study and the first topography [
13]. Shortly afterwards, the American vulcanospeleologist W.R. Halliday explored the cave and, after analyzing the survey of SEVG, stated that the known part was 6021 m long and listed it as the longest volcanic cavity in the world [
25].
In 1973 and 1974 the British C. Wood and his team explored the cave and published an extensive geological study of Cueva de San Marcos and Cueva del Viento proper, with a new topography of the latter including a new 2340 m branch that runs through a level lower than the rest of the cave [
14]. The cave reached 10,002 m, but by then it could no longer be claimed as the longest known volcanic cavity in the world as both Leviathan Cave (Kenya) and Kazumura Cave (Hawaii) exceeded 11 km [
26].
Between 1986 and 1988, the GIET (Grupo de Investigaciones Espeleológicas de Tenerife) of the University of La Laguna found several unpublished sections of the deep branch discovered by Wood and Mills (Ingleses gallery), the total length rising to 10,964 m [
22]. The same team carried out the first biological study of the complete Cueva del Viento System [
27].
In 1988, members of the Grupo de Espeleología de Tenerife Benisahare (henceforth Benisahare) discovered a connection with the well-known Cueva del Sobrado whose topography, commissioned by the Cabildo of Tenerife in 1989 and made by J.L. Martín, added another 3570 m in one of the most intricate areas of Cueva del Viento.
During the exhibition “Labyrinths of lava” of the Museum of Natural Sciences of Tenerife, Sergio Socorro and collaborators presented in 1990 a three-dimensional topographic assembly of the whole Cueva del Viento, in which the “axis” was taken as in the Wood and Mills topography given the greater accuracy of the orientation of the sections.
The removal of rubble from the lower entrance of Cueva del Sobrado in 1994, in a work directed by the later J.J. Hernández Pacheco for the Cabildo of Tenerife, uncovered a 17 m deep pit connecting with a new lower branch 2346 m long [
23]. The published topographies total 17,032 m of galleries, but searching, exploration and surveying has been continued by the Benisahare group, with a provisional figure of 18,249 m, although still unpublished. Cueva del Viento is not the first but the seventh longest among volcanic caves [
6], but many specialists agree that its complexity of passageways and its geomorphology are the most outstanding among this type of caves [
14].
The first topographic and biological survey of Cueva de Felipe Reventón was performed by the GIET of La Laguna University [
28], whose members together with Benisahare have continued the task over the years [
16].
4. Geology
The Teide volcano represents the culmination of the evolution of an intraplate oceanic island. The Canary Islands are located on a thick oceanic crust close to a continent. This results in subsidence (sinking of the crust due to the weight of an island) being minimal, which means that these islands can remain emerged for more than 20 million years. In turn, this allows long periods of magmatic evolution and successive eruptive cycles, producing stratovolcanos like Teide. In Tenerife, during the last three million years, two previous stratovolcanos have already grown up and eventually slid down to the sea. At present, after 200,000 years of the third cycle of construction of a stratovolcano, the current Teide is still active in the center of the island [
29].
When an island is located right on the hot spot in its initial formation, basaltic magmas coming directly from the earth’s mantle predominate. This is called a basaltic shield island; most of the Hawaiian Islands do not go beyond that phase (while active). Conversely, Tenerife is already in a mature state, away from the hot spot, such that in its entrails various magmatic chambers can interact with each other [
29].
All the caves of the Cueva del Viento System are volcanic cavities framed in this context. They were formed 27,030 ± 430 years ago in lavas from the first eruptive phase of the Pico Viejo volcano, located on the slopes of Teide [
29] (see
Figure 1).
The composition of lava in the Cueva del Viento System does not correspond to a basalt in the strict sense, which would be the fluid lavas that normally develop as a “pahoehoe” flow (a more fluid basaltic lava with a smooth to ropy surface), where lava tubes are usually formed. The so-called plagioclase basalts of Pico Viejo, in which the caves lie, constitute somewhat evolved lavas in their geochemistry. These lavas had already begun magmatic evolution, so that their composition is between tephrites and tephryphonolites, with the particularity of containing a high proportion of plagioclase crystals [
4]. These crystals were already formed in the magma before the eruption and increased the viscosity of the lava. In other words, the fluidity of the lava is diminished by abundant solid particles in suspension that the plagioclases make up. This lack of fluidity could be compensated by the steep slope of the area and decreased eruptive flow or distributed among several emission centers, making them compatible with the pahoehoe flow mechanism that usually forms the lava tube caves.
On the surface, the area where the different entrances of the Cueva del Viento System are distributed is covered by subsequent flows from Teide-Pico Viejo volcanic building. These more evolved lavas (tephryphonolites and phonolites) as well as the more recent phonolites of Roques Blancos (1800 BP) [
29], formed as huge channels of viscous lava now covering most of the flow that originated the caves, so that its outcrops are very scarce in the area concerned.
This succession of flows determines, in part, the surface vegetation. As a deep soil has not developed, most of the recent flows are occupied by a mixed forest dominated by Pinus canariensis, a tree capable of growing under these conditions, unlike the laurel forest species that might be expected with this altitude and climate.
These caves follow the downward slope of the island dropping 485 m at an average inclination of 11°. They have a unique geomorphological complexity among the known volcanic tubes, characterized by the convergence and bifurcation of their numerous galleries [
14]. This type of tube formation is the most difficult to understand since a great complexity is observed in their structure and arrangement of the branches. Both Cueva del Viento and Cueva de Felipe Reventón are good models of this type of lava tubes. The eruption probably lasted for several months producing flows hundreds of meters wide, leading to the formation of this complex cave with branches that run in three superimposed floors, corresponding to various episodes of the initial activity of Pico Viejo volcano [
14].
In these types of tubes, the way in which the pahoehoe spreads greatly influences how the spatial arrangement of the underground network of ducts can be complicated. Furthermore, after a network of ducts is established, a new flow can overlap on top of the former, creating a new system that can become intercommunicating with the underlying passages. In Breveritas Superior and Sobrado galleries there are two vertical pots through which cascades of lava flowed from a higher tube to a lower one.
5. Geomorphology
The complex network of passages in Cueva del Viento proper is distributed in three main levels (
Figure 4):
An upper one (Level 3) that connects the main tubes through narrow shallower passages;
An intermediate one (Level 2) that includes Sobrado Superior (3.57 km) and most of Cueva del Viento (7.82 km);
A deep Level 1 through which Ingleses (3.14 km) and Sobrado Inferior (2.35 km) galleries lie.
The axis Sala de la Cruz–Pájaros Gallery–Belén–Breveritas Entrance–Piquetes (see
Figure 2) probably corresponds to a lava channel that was initially open to the sky in the pahoehoe flow [
14]. In some sectors of this axis, closure sutures are observed on the cave ceiling and bulges on the outer surface of the terrain (
Figure 5).
In general, the internal surfaces within the cavity consist of cornices, terraces, benches, and tiers due to variations and stabilizations that the level of the lava flow undergoes within the already formed tube. The different levels of flow stabilization are marked symmetrically on both walls, sometimes only by flux striations.
Changes in viscosity are also marked. The smooth surfaces correspond to phases in which the lava circulated with great fluidity (
Figure 6); instead, a rough floor would be the last residual flow that was solidifying (
Figure 7).
In the Sobrado Superior sector, the geomorphological beauty of its galleries and its didactic interest are notable. There are side terraces, multiple levels marked by lava when changing height, effects of centrifugal force on lateral benches, etc. The area called the Labyrinth is an intricate network of interconnected tubes of almost circular section and smooth walls, which can rival the other existing labyrinth in Cueva de Felipe Reventón (
Figure 6).
Unlike the upper galleries, that of Ingleses (Level 1) evidently did not receive solidifying lava streams and the flow was never stabilized for long enough to form side terraces and benches, which are scarce and not very prominent here. Due to the distance from the surface, the lava is “cleaner” with few subsequent mineral coatings deposited by infiltrated water, as it is more likely to settle at higher levels. In addition, several lava flows from later eruptions have covered the ground surface above these galleries, increasing their distance from the exterior.
The spectacular black band that can be seen along the main gallery of Ingleses must have been formed by a pulse of very fluid lava that flooded the conduit to a certain height (
Figure 7 and
Figure 8). The rapid fall in the lava level left a thin layer that quickly cooled and vitrified in contact with the walls.
In various passages of Ingleses one can see how the hot flows through the tube “corroded from the inside” the solidified lava in the pahoehoe flow on the surface that had formed the roof of the tube. In other words, while the flow and configuration of the ducts last, the stream drags along loose pieces detached from the “puff pastry” of the cavity ceiling, which constitutes the pahoehoe flow itself, and that today we see sectioned in walls and roofs (
Figure 8). These detached blocks are also seen welded to the ground in various places throughout the cavity.
Sometimes, the magnitude of this “re-excavation towards the ceiling” is so great that other higher ducts are intercepted from below (
Figure 7). Such conduits have not yet been explored, and they could correspond to Level 2 at which the main galleries of Breveritas Superior formed above Ingleses Gallery. These geological cuts should be like those observed by Ollier and Brown [
30], which inspired their now outdated theory to explain the formation of tubes, based on what they called “layered lava” and related it to a supposed laminar flow; however, it corresponds to the complex movement of pahoehoe flows with various lobes, embryonic tubes, lava channels, and sometimes continuous mantles of lava.
The cylindrical tube configuration only occurs when the current reaches the ceiling persistently, to consolidate a continuous inner lining. Otherwise, as in many sections, the ceiling is irregular and may show pahoehoe “puff pastry” structures, although more or less blurred by mineral deposits in the upper levels of Breveritas.
The main branches of Cueva de Felipe Reventón and the connecting galleries correspond largely to embryonic lava tube systems that develop in the complex advance of a pahoehoe stream. For this reason, these areas are made up of complicated labyrinths and tubes at different heights, where an authentic tangle of lava outbreaks is interwoven that, at the time, constituted the advanced front of the pahoehoe flow (
Figure 6).
6. The Cave Environment
Lava tube caves originate very quickly in geology and remain very close to the surface, usually a few meters deep throughout their length [
31]. This is the start point for their ecological succession, erosion, and final destruction due to collapse or to clay silting; this happens in a much shorter time span than karstic caves: a maximum of 500,000 years for Hawaiian caves [
5]. Some Canary tubes have persisted for longer, especially when they are gently inclined and develop in dry climates, as in the 992,000 ± 21,000 years old Cueva del Llano formed by drainage of a lava lake in the subdesert Fuerteventura island, or the exceptional case of Cueva de Aslobas in Gran Canaria with more than 13 Ma [
32,
33]. The environment inside very recent lava tubes is highly influenced by surface weather, due to the abundant interconnected cracks and voids of the lava and to the absence of covering soil. Therefore, this habitat is not yet suitable for cave-adapted fauna. Ecological succession on the surface will provide a layer of soil and vegetation on the middle-aged lavas over a period dependent on local climate. A parallel, slightly delayed ecological succession will take place inside the tube until it reaches a humid and stable cave environment [
34]. The process is sometimes accelerated by deposits of small-sized pyroclasts (lapilli and cinders) that almost immediately cover the surface providing sufficient isolation for cave conditions, as happens in Cueva de Don Justo on El Hierro island [
34].
In limestone caves ecological succession advances downwards creating new habitats ever deeper, but in lava tubes the situation is reversed, and succession progresses upwards as protective soil and vegetation develop on the surface [
35]. Thus, deeper galleries of the cave (or deeper parts of a big gallery) mature before shallower ones. Usually, in the Canary Islands lava tubes reach this mature ecological stage between many hundreds and some thousand years depending on the local climate, and they become senile and ecologically decaying after a few hundred thousand years. Therefore, Cueva del Viento and Cueva de Felipe Reventón are now in their best stage to host a rich adapted fauna due to their age (ca. 27,000 ybp) [
21]. Their situation in a vast area of moderately recent lavas still with an inner network of cracks and voids allows obligate subterranean fauna to reach the caves from other underground habitats. The moderate altitude of the area is also favorable given the local climate on the north slope of the island, mesic at middle altitudes between 500 and 1200 m a.s.l. due to humid trade winds, but drier both at lower and higher altitudes. Other lava tubes located in the dry lowlands usually harbor a much poorer cave-adapted fauna [
20,
34].
Stable temperatures and high humidity prevail throughout the year in the most suitable caves, a necessary requirement for cave adapted fauna. The deeper a gallery the greater the environmental stability, but with lower availability of trophic resources. The absence of water streams inside the tubes slows down the advance of organic matter in depth. Thus, the deepest passages of lower levels in Cueva del Viento proper differ from those in shallower ones, the former with scarcer food resources, more constant temperature and humidity, and sometimes higher CO
2 concentration [
36]. In addition to trophic scarcity, these parameters are unsuitable for poorly adapted species and lead to the dominance of troglobionts (i.e., obligate inhabitants of subterranean habitats, usually but not necessarily with the morphological syndrome of lack of pigment, eyelessness and elongated appendages) [
1]. This fact was already verified in lava tubes of Australia and Hawaii [
10] and is noticeable in Sobrado Inferior and especially in Ingleses, both at the third lower level at least 15 m below surface with a scarce, almost exclusively troglobiotic fauna [
36]. In Cueva de Felipe Reventón there are no different levels of galleries and environmental features are very similar along the cave. The main contrast of ecological significance between its galleries is their substrate, sometimes bare lava but often with remarkable earth accumulation derived from the overlying soil through very small roof collapses or thick cracks.
Most lava tubes are shallow, unless they have been covered by later flows belonging to more recent eruptions. This situation close to the surface and the abundant cracks in the lava allow the plant roots to reach the cave, often dangling from the ceiling (
Figure 3 and
Figure 9). This is an important resource for the animal community, especially for root-feeding and sap-sucking species, the latter being markedly abundant in individuals [
31]. These root-dependent species are common in lava tubes around the world, especially from volcanic archipelagos but also in limestone caves, although the latter almost exclusively in the tropics [
37]. As many as 18 cave-adapted planthopper species are found in the non-tropical volcanic Macaronesian Islands (Canary Is., Madeira and Azores) but only four species are known from karst caves in the rest of West Palearctic [
38,
39,
40]. In some sections of the whole Viento cave system, especially in Sobrado Superior, sap-sucking insects are by far the most abundant cave-dwelling animals, represented by the planthopper
Tachycixius lavatubus Remane and Hoch (
Figure 10). However, the roots do not reach all passages of the cave, especially those at the deepest galleries or those in the cave sections underlying thick overlapping modern lavas. Nevertheless, once the organic matter has reached the shallower parts of the lava tube, it can diffuse horizontally (and more slowly vertically) towards the rest of the cave and be integrated in the food chain of the ecosystem.
Besides the main entrances, there are along both caves several partial collapses of the original compact ceiling, facilitating the connection with the overlying soil and sometimes even with the surface. This allows the entry by many trogloxenes (animals not linked to the caves) and subtroglophiles (only linked for part of its life cycle) [
41], which form an important resource for predatory species in many parts of the lava tubes, sometimes far from any large entrance. Abundant flying insects can be found in such spots, like the subtroglophile
Megaselia scuttle flies that probably are common prey for a diversity of troglobiotic web-spiders, which could not survive only feeding on other troglobionts. In the twilight zone close to entrances, these flying insects are preyed on by the abundant spider
Meta bourneti Simon, a typical eutroglophile with permanent populations completing its life cycle inside and rarely outside the caves [
41].
Relative humidity is high and ceiling drip very frequent and even abundant in many parts, but running water is rarely present except just after heavy rains. Almost all water quickly percolates through the abundant cracks of the lava floor connected with a network of voids in the host rock. Thus, no significant water ponds are present, and no groundwater species have ever been found, given that the water table lies much deeper than these caves. Environmental features in Cueva del Viento and Cueva de Felipe Reventón are very similar since they are in the same lava flow, roughly at the same altitude and at an identical stage of maturity, and they share 32 of their troglobiotic species. There are two notable differences between the two caves: (i) the greater depth of the lowest galleries in Cueva del Viento (e.g., Ingleses), with the consequent greater shortage of organic matter input and of troglobiont individuals, and (ii) the greater accumulation of soil in some passages of Cueva de Felipe Reventón, providing a different habitat for the occurrence of some species never found in the rest of the cave system.
7. Animal Ecology and Diversity
The ecological driver for the high diversity in Cueva del Viento System is evidently the high-productivity due to abundant roots because of being very close to the surface [
42]. As many as 79 different troglobionts are known so far from the island of Tenerife. In the northeastern geologically old Anaga peninsula there are nine troglobionts, two occurring in the only existing cave (a volcanic pit), and the rest in the mesocavernous shallow substratum (MSS), a colluvial subterranean habitat from the mountains of old areas [
1]. Eight of these species are exclusive to this particular area and vicariant with respect to closely related ones from the rest of the island. This is probably due to an ancient ecological isolation, given the high proportion of local Anaga endemics, both underground and surface species [
15,
43,
44]. Among the 72 troglobionts occurring in the rest of the island (where the Cueva del Viento System is located) 14 are known only from the MSS and 58 can be found in MSS and/or in caves, 42 of which are present in Cueva de Felipe Reventón and/or Cueva del Viento (38 and 36 respectively) (see
Table 1). Thus, the fauna of these two caves is an important reflection of the troglobiotic fauna of the island. Only 9 troglobionts are exclusive to the Cueva del Viento System, the remaining 33 species being also found in some or many other subterranean places in Tenerife, either caves or MSS. This reflects the importance of MSS and other subsurface habitats of volcanic terrains as a means of dispersal for troglobionts. Some of them can be found in rather distant caves that are unconnected since they occur in lava flows geographically and chronologically separated. Such distant populations of a single species usually show morphological, behavioral or genetic differences that reflect the slow but finally successful dispersal of troglobionts through underground spaces, even without proper caves in between.
Troglobiotic lifestyle does not always imply apparent morphological adaptations (eye and pigment regression, lengthening of appendages). There are different life forms of troglobionts: troglomorphic, with these characters highly accentuated and large body size; euedaphomorphic, with eye and pigment regression but small body and short appendages; and hypogeomorphic, with variable regressive characters but unchanged appendages and body size compared to surface species [
41]. Some eyeless and depigmented species with an euedaphomorphic or hypogeomorphic rather than troglomorphic body pattern are more frequent in Cueva de Felipe Reventón than in Cueva del Viento, especially small carabid and histerid beetles [
45,
46].
Primary consumers are basically those feeding on roots. Two species feed on fresh root tissues: the moth
Schrankia costaestrigalis (Stephens) and the scarce weevil
Oromia hephaestos (
Figure 11). The eutroglophile
Schrankia costaestrigalis is very abundant, especially in Felipe Reventón, where the caterpillars feed on tender roots and pupate in cocoons hanging from the ceiling and covered with dry root pieces. The adult moths reproduce and live permanently in the caves, but unlike
S. howarthi Davis and Medeiros from Hawaiian caves [
47], the females have no troglomorphic adaptations and can fly perfectly.
Oromia hephaestos belongs to an eyeless endemic genus with species occurring either in caves or in the MSS from the islands of La Gomera, Tenerife, and Gran Canaria, with a notable radiation in the latter [
48]. The only sapsucker species is the planthopper
Tachycixius lavatubus, by far the most abundant troglobiont in both caves. This highly troglomorphic species belongs to a genus with other two hypogeomorphic species occurring in colluvial MSS, and a scarce surface-dwelling species. As in other planthoppers, males emit a specific vibration transmitted through the roots, and then answered by females [
49].
Tachycixius lavatubus can be found in almost any cave around the island, although several different allopatric species probably exist, according to unpublished population genetic and bioacoustic studies [
50].
Detritivores are more varied than root-feeding species, but due to the broad spectrum of feeding behavior necessary to survive in the cave, it is often difficult to know the preferences of each millipede, woodlouse, springtail or cockroach. The pill-millipede
Glomeris speobia and the woodlouse
Venezillo tenerifensis are among the most widespread troglobionts on the island and are regularly seen in both the Icod caves discussed here (
Figure 12 and
Figure 13). The millipede genus
Dolichoiulus is a paradigm of island radiation with 21 endemic species to Tenerife, three of them troglobionts [
51] (
Figure 14);
Dolichoiulus labradae and
D. ypsilon are becoming rarer in these caves, probably due to the increasing abundance of the alien, invasive troglophile
Blaniulus guttulatus (F.). The woodlouse
Porcellio martini is also scarce, found here and there in other caves of the island. Another woodlouse
Trichoniscus bassoti and the springtails
Troglopedetes vandeli and
T. cavernicolus may not be correctly identified [
52,
53], since the presence of non-endemic troglobionts on an oceanic island is not conceivable due to their inability to disperse over distances across the sea.
Cockroaches of the genus
Loboptera are among the most characteristic components of the troglobiotic fauna of the western Canary Islands, with eight species on Tenerife, seven allopatric to each other but the eighth (
L. troglobia) widely distributed and sympatric to many others. They are among the biggest local troglobionts, eyeless, wingless, with reduced number of ovarioles and living either in caves or in the MSS [
54]. Two species occur in both Cueva de Felipe Reventón and Cueva del Viento: the big and scarce
L. subterranea which is exclusive to the Icod area, and the smaller and more troglomorphic and widespread
L. troglobia (
Figure 15).
Most predatory troglobionts have reduced populations, some with very few individuals, and many species act also as scavengers. Five different pseudoscorpions are found here, all endemic to the island except the eyeless
Microcreagrina subterranea which also occurs on other islands. The remaining species are Chthoniidae, two of them especially troglomorphic:
Paraliochthonius superstes and
P. setiger; only
Lagynochthonius curvidigitatus is exclusive to Cueva de Felipe Reventón [
55]. The highly diversified spider genus
Dysdera is represented by five species occurring in both caves, one of them (
D. unguimmanis) highly troglomorphic and probably parthenogenetic, since no males have ever been found (
Figure 16). Each one has a different epigean sister species, so they originated by five independent colonization events of the underground environment [
56]. Among troglobiotic web spiders
Troglohyphantes oromii is the most abundant (
Figure 17), not only in these two caves but in the rest of the island except in Anaga; a similar situation is that of the widespread centipede
Lithobius speleovulcanus.
All troglobiont ground beetles are eyeless but otherwise not especially troglomorphic (
Figure 18); Pterostichinae are medium size beetles in two endemic genera (
Wolltinerfia and
Gietopus), while Trechinae are smaller and are represented by the monospecific endemic genus
Spelaeovulcania canariensis and two euedaphomorphic species of
Lymnastis (
Figure 19). The diversity of subterranean rove beetles is remarkable, both in the Canary Islands (27 spp.) and in Icod caves (6 spp.) [
57], and they are usually more troglomorphic than ground beetles, as shown by the marked elongation of body and appendages in
Domene alticola and
D. vulcanica (
Figure 20).
Besides the subterranean obligate fauna, the shallowness of these caves facilitates the input of trogloxene and subtroglophile animals. Thus, the total number of arthropod species found in them roughly doubles the number of troglobionts [
58], but the trogloxenes are almost absent in the deepest levels of Breveritas Superior and Sobrado galleries. Local eutroglophiles are usually widespread species, some introduced like the millipedes
Blaniulus guttulatus and
Choneiulus subterraneus (Silvestri)
, and some probably native species like the spider
Meta bourneti, the psocid
Psyllipsocus ramburii (Selys-Longchamps) and the moth
Schrankia costaestrigalis. The abundant larvae of the subtroglophile winter crane fly
Trichocera maculipennis Meigen capture their preys in sticky silk threads, being important predators in most of the cave communities of Tenerife.
8. Paleontology of the Caves
Like in other volcanic archipelagos, Canary Islands lava tubes and pits are the best paleontological sites, especially for quaternary subfossils of vertebrates. The accumulation of bird bones is usually not far from cave entrances, which sometimes function as traps that allow the birds to come in by accident, and get injured and lost, and soon die. Lizards and small mammals can actively enter and explore the habitat, reaching longer and deeper distances due to their higher resistance to starvation. Actually, in Cueva del Viento it is rather frequent to see lost or recently dead lizards and young rabbits which penetrate through narrow, small roof collapses existing all along the lava tube due to its shallowness. The Cueva del Viento System is one of the most important paleontological sites in the archipelago, where many remains of several already extinct vertebrates have been found. Bones of the Tenerife Giant Lizard
Gallotia goliath (Mertens), over one meter long, were collected abundantly in Piquetes galleries in the lower part of Cueva del Viento [
59], underlying a surface habitat where these reptiles were probably common given the relatively warm climate and the absence of forest. The Tenerife giant rat
Canariomys bravoi Crusafont and Petit was also present in this part of the cave, but particularly more abundant in galleries at higher altitude, especially in Sobrado Inferior passages (
Figure 21).
In 1994 an important paleontological site was discovered at the end of a 600 m long Pájaros Gallery in Breveritas Superior (see
Figure 2), with bone remains of more than 200 vertebrate individuals comprising 6% lizards, 46% mammals (mainly extinct giant rats and recent rabbits and rats) and 48% birds, among which there were some extant species and some others extinct on the island or completely extinct [
60] (
Table 2). Such accumulation of bones away from accessible entrances is explained by the presence of an old collapse that interrupts the gallery just before an entrance of the cave (Galería Belén, see
Figure 2) by which most of the animals could have entered before this erosional event. Two extinct bird species were found at this spot: the Canary Quail (
Coturnix gomerae Jaume, McMinn and Alcover) and the Long-legged Bunting (
Emberiza alcoveri Rando and López), the latter being the only flightless bunting so far known [
61].
The Tenerife giant rat was probably the only one of the abovementioned vertebrates that used the cave as part of its habitat, as the introduced Brown Rat does today [
62]; the other species would be inside just by accident. Bones of the giant rat have also been found in Cueva de Felipe Reventón, though less abundantly. Some of these bones were dated with C
14 with ca. 25,000 BP, the minimum age applicable to the whole cave system [
63]. Extinct species disappeared soon after the arrival of the aboriginal people (ca. 2500 ybp) [
64].
9. Conservation Status
The Cueva del Viento System is in an inhabited area, so it is threatened much depending on the caves and sectors. The main problem is that the whole municipality of Icod de los Vinos lacks a sewage system, and the solution for each house is to have a cesspool or, alternatively and much worse, to drill a hole until reaching the highly porous rock which very often is just a cave. The local people call this “to find the volcano”, quite easy given the abundance of holes and caves in the underground of the area. Thus, the lower part of Cueva del Viento (Piquetes) is polluted by raw sewage and dirty mud, and is almost impassable; Breveritas Inferior is also contaminated with filtered dirty waters and even bad smell, but the central part (Breveritas Superior) is reasonably well preserved though continually endangered given the uncontrolled construction of houses; the relatively short Galería Belén is just below a single house, enough to cause a sewer smell; the access to Sobrado Inferior is through a controlled entrance to Sobrado show cave and the visitors are not allowed, but some traces of pollutants were detected in past analyses [
16]; the upper part (Sobrado Superior) is in a mostly natural area under pine forest and is owned by the local government, double-gated and in very good condition. The only solution for this important contamination would be a sewage system in densely inhabited zones, and individual treatment plants for isolated houses. Unfortunately, the municipality has neither money nor special will to do it, and with time more and more illegal houses have been built. All entrances are gated except Piquetes, Breveritas Inferior and Belén, which are not visited due to unpleasant conditions.
A 150 m long stretch of Sobrado Superior is dedicated to public visits, but this does not cause a noticeable impact to the environment because it is between two large entrances. Ventilation is higher and humidity lower than in the rest of the cave, and 20 years ago, before its adaptation for visits, it had been verified that it lacked troglobionts. The visitors enter the cave in groups of maximum 10 people, each with a personal headlamp since the tube has no electric lighting to prevent the growth of green algae and mosses. The touristic use of a small portion devoid of adapted fauna compensates for the control which is exercised over the adjacent richer parts of Sobrado.
Cueva de Felipe Reventón has a solid gate in the only entrance in public land and only a few people can visit it. However, the upper part of the cave lies under private land and a house is nearby, with danger of sewage filtering. There are no apparent signs of contamination and the troglobiotic fauna is rich, but water analyses made in 2000 revealed the presence of nitrates and nitrites (unpublished own data).
Cavers and conservationists have been for many years arguing for the protection of the area, always without success. A LIFE-Nature project on the biodiversity and conservation of caves in Tenerife, La Palma and El Hierro islands was carried out in 1999–2001 and included a project for a sewage system in the Icod area, but the installation has never been performed [
15]. A project by the Cabildo de Tenerife to declare a protected area around these caves is dragging on for many years without taking place, and it will be increasingly difficult due to pressures by landowners who want to build.

























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}