
The Herbaceous Understory Plant Community in the Context of the Overstory: An Overlooked Component of Tropical Diversity


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Data Collection
2.3. Statistical Analyses
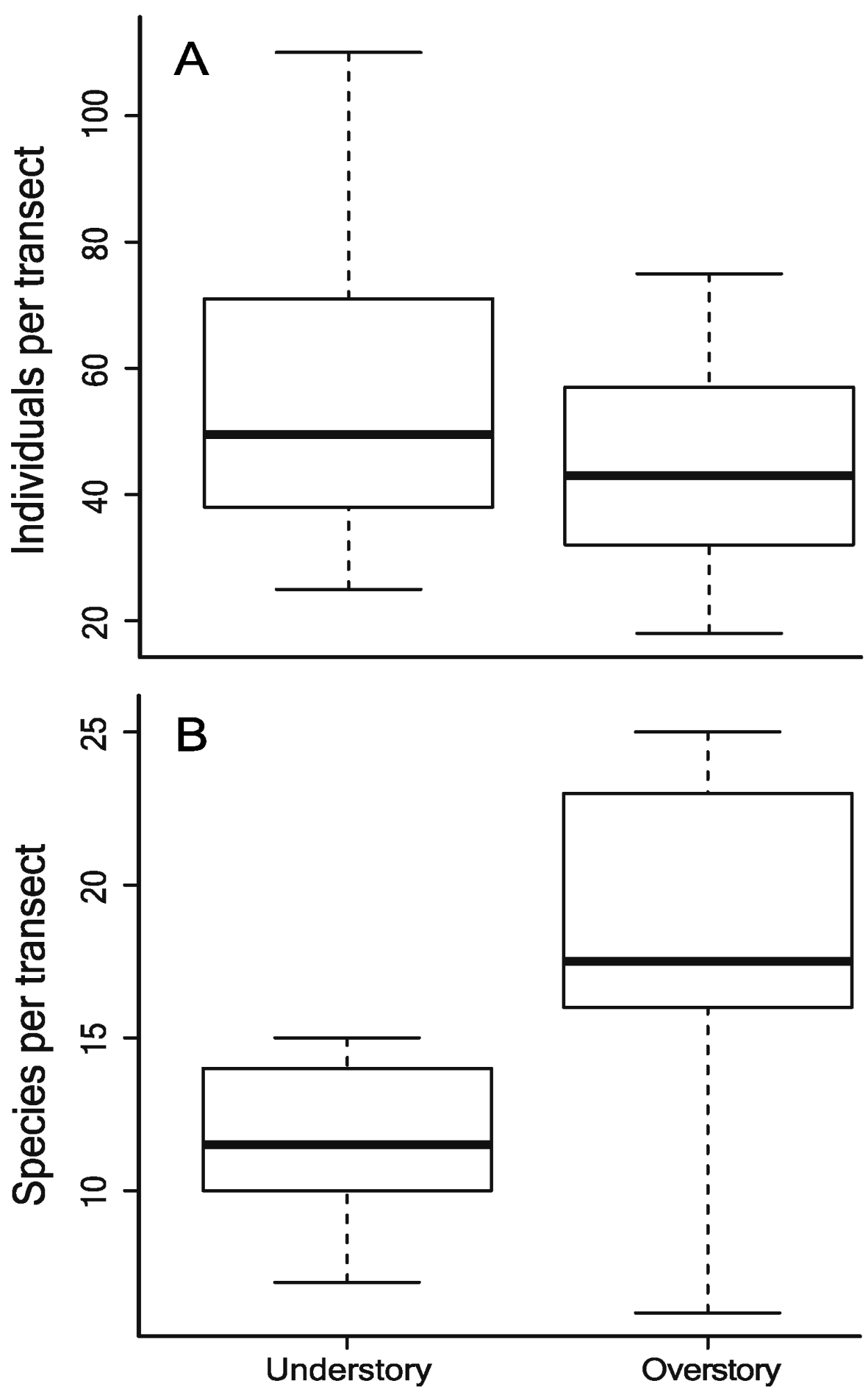
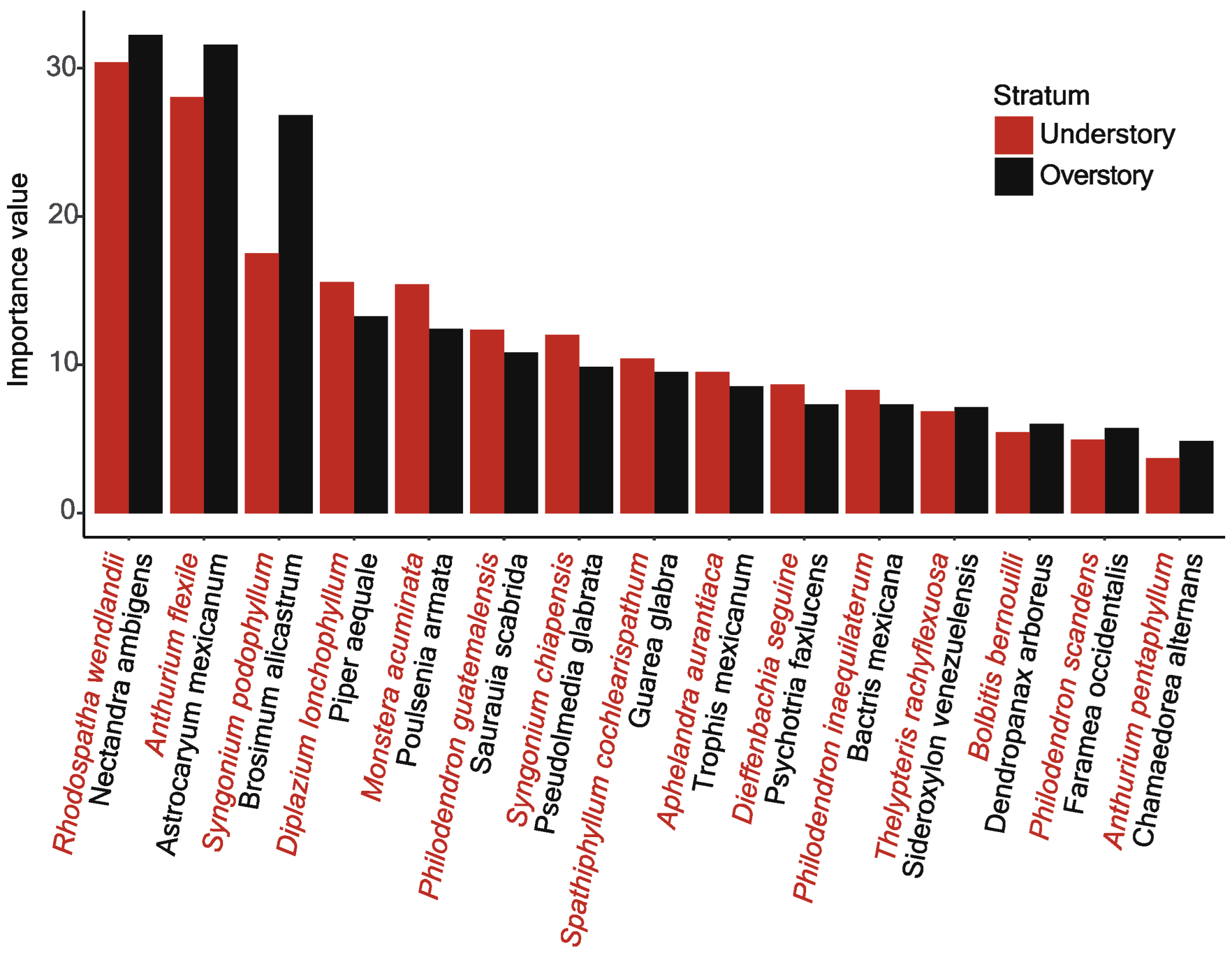
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gentry, A.H. Changes in Plant Community Diversity and Floristic Composition on Environmental and Geographical Gradients. Ann. Mo. Bot. Gard. 1988, 75, 1–34. [Google Scholar] [CrossRef]
- Ashton, P. Species Richness in Plant Communities. In Conservation Biology; Fiedler, P., Jain, S., Eds.; Springer: Boston, MA, USA, 1992. [Google Scholar]
- Dirzo, R.; Raven, P. Global Biodiversity and Loss. Annu. Rev. Environ. Nat. Resour. 2003, 28, 137–167. [Google Scholar] [CrossRef]
- Wright, S.J. Plant Diversity in Tropical Forests: A Review of Mechanisms of Species Coexistence. Oecologia 2002, 130, 1–14. [Google Scholar] [CrossRef]
- Raven, P.H.; Gereau, R.E.; Phillipson, P.B.; Chatelain, C.; Jenkins, C.N.; Uloa Uloa, C. The Distribution of Biodiversity Richness in the Tropics. Sci. Adv. 2020, 6, eabc6228. [Google Scholar] [CrossRef] [PubMed]
- Condit, R. Research in a Large, Long-Term Tropical Forest Plot. Trends Ecol. Evol. 1995, 10, 18–22. [Google Scholar] [CrossRef]
- Valencia, R.; Balslev, H. High Tree Alpha-Diversity in Amazonian Ecuador. Biodivers. Conserv. 1994, 3, 21–28. [Google Scholar] [CrossRef]
- Gilliam, F.S. The Ecological Significance of the Herbaceous Layer in Temperate Forest Ecosystems. BioScience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Croat, T.B. Flora of Barro Colorado Island; Stanford University Press: Stanford, CA, USA, 1978. [Google Scholar]
- Janzen, D.H.; Leisner, R. Annotated Check-List of Plants of Lowland Guanacaste Province, Costa Rica, Exclusive of Grasses and Non-Vascular Cryptogams. Brenesia 1980, 18, 15–90. [Google Scholar]
- Gentry, A.H.; Dodson, C. Contribution of Nontrees to Species Richness of a Tropical Rainforest. Biotropica 1987, 19, 149–156. [Google Scholar] [CrossRef]
- Cicuzza, D.; Krömer, T.; Poulsen, A.D.; Abrahamczyk, S.; Delhotal, T.; Piedra, H.M.; Kessler, M. A Transcontinental Comparison of the Diversity and Composition of Tropical Forest Understory Herb Assemblages. Biodivers. Conserv. 2013, 22, 755–772. [Google Scholar] [CrossRef]
- Massante, J.C.; Götzenberger, L.; Takkis, K.; Hallikma, T.; Kaasik, A.; Laanisto, L.; Hutchings, M.J.; Gerhold, P. Contrasting Latitudinal Patterns in Phylogenetic Diversity between Woody and Herbaceous Communities. Sci. Rep. 2019, 9, 6443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentry, A.H. Patterns of Neotropical Plant Species Diversity. Evol. Biol. 1982, 15, 1–84. [Google Scholar]
- Dirzo, R.; Horvitz, C.C.; Quevedo, H.; López, M.A. The Effects of Gap Size and Age on the Understorey Herb Community of a Tropical Mexican Rain Forest. J. Ecol. 1992, 80, 809–822. [Google Scholar] [CrossRef]
- Costa, F.R.C. Structure and Composition of the Ground-Herb Community in a Terra-Firme Central Amazonian Forest. Acta Amaz. 2004, 34, 53–59. [Google Scholar] [CrossRef]
- Linares-Palomino, R.; Cardona, V.; Hennig, E.I.; Henson, I.; Hoffmann, D.; Lendzion, J.; Soto, D.; Herzog, S.K.; Kessler, M. Non-Woody Life-Form Contribution to Vascular Plant Species Richness in a Tropical American Forest. Plant Ecol. 2009, 201, 87–99. [Google Scholar] [CrossRef]
- Vazquez G, J.A.; Givnish, T.J. Altitudinal Gradients in Tropical Forest Composition, Structure, and Diversity in the Sierra de Manantlán. J. Ecol. 1998, 86, 999–1020. [Google Scholar]
- Davies, T.J.; Buckley, L.B. Phylogenetic Diversity as a Window into the Evolutionary and Biogeographic Histories of Present-Day Richness Gradients for Mammals. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 2414–2425. [Google Scholar] [CrossRef]
- Redford, K. The Empty Forest. BioScience 1992, 42, 412–422. [Google Scholar] [CrossRef]
- Dirzo, R.; Miranda, A. El Límite Boreal de La Selva En El Continente Americano: Contracción de La Selva y Solución de Una Controversia. Interciencia 1991, 16, 240–247. [Google Scholar]
- González-Soriano, E.; Dirzo, R.; Vogt, R.C. Historia Natural de Los Tuxtlas; Universidad Nacional Autónoma de México: Mexico City, Mexico, 1997. [Google Scholar]
- Bongers, F.; Popma, J.; Meave-Del Castillo, J.; Carabias, J. Structure and Floristic Composition of the Lowland Rain-Forest of Los-Tuxtlas, Mexico. Vegetatio 1988, 74, 55–80. [Google Scholar] [CrossRef]
- Ibarra-Manríquez, G.; Martínez-Ramos, M.; Dirzo, R.; Nuñez-Farfán, J. La Vegetación. In Historia Natural de Los Tuxtlas.; González-Soriano, E., Dirzo, R., Vogt, R.C., Eds.; Universidad Nacional Autónoma de México: Mexico City, Mexico, 1997; pp. 61–85. [Google Scholar]
- Mann, H.B.; Whitney, D.R. On a Test of Whether One of Two Random Variables Is Stochastically Larger than the Other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Kammesheidt, L. The Role of Tree Sprouts in the Restoration of Stand Structure and Species Diversity in Tropical Moist Forest after Slash-and-Burn Agriculture in Eastern Paraguay. Plant Ecol. 1998, 139, 155–165. [Google Scholar] [CrossRef]
- Zanne, A.E.; Tank, D.C.; Cornwell, W.K.; Eastman, J.M.; Smith, S.A.; FitzJohn, R.G.; McGlinn, D.J.; O’Meara, B.C.; Moles, A.T.; Reich, P.B.; et al. Three Keys to the Radiation of Angiosperms into Freezing Environments. Nature 2014, 506, 89–92. [Google Scholar] [CrossRef]
- Rothfels, C.J.; Li, F.; Sigel, E.M.; Huiet, L.; Larsson, A.; Burge, D.O.; Ruhsam, M.; Deyholos, M.; Soltis, D.E.; Stewart, C.N.; et al. The Evolutionary History of Ferns Inferred from 25 Low-copy Nuclear Genes. Am. J. Bot. 2015, 102, 1089–1107. [Google Scholar] [CrossRef] [Green Version]
- Faith, D.P. Conservation Evaluation and Phylogenetic Diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Community Ecology Package, Version 2.3-5; R Package. 2016. Available online: http://www.cran.r-project.org (accessed on 29 August 2022).
- Braun-Blanquet, J. Planzensoziologie; Springer: Vienna, Austria, 1951. [Google Scholar]
- Pfeifer, D.; Bäumer, H.; Schleier, U. The “Minimal Area” Problem in Ecology: A Spatial Poisson Process Approach. Comput. Stat. 1996, 11, 415–428. [Google Scholar]
- Horsley, S.B. Mechanisms of Interference between Hay-Scented Fern and Black Cherry. Can. J. For. Res. 1993, 23, 2059–2069. [Google Scholar] [CrossRef]
- George, L.O.; Bazzaz, F.A. The Fern Understory as an Ecological Filter: Emergence and Establishment of Canopy-Tree Seedlings. Ecology 1999, 80, 833–845. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the Number of Tree Species in Tropical Forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Mangan, S.A.; Schnitzer, S.A.; Herre, E.A.; Mack, K.M.L.; Valencia, M.C.; Sanchez, E.I.; Bever, J.D. Negative Plant-Soil Feedback Predicts Tree-Species Relative Abundance in a Tropical Forest. Nature 2010, 466, 752–755. [Google Scholar] [CrossRef]
- Bagchi, R.; Gallery, R.E.; Gripenberg, S.; Gurr, S.J.; Narayan, L.; Addis, C.E.; Freckleton, R.P.; Lewis, O.T. Pathogens and Insect Herbivores Drive Rainforest Plant Diversity and Composition. Nature 2014, 506, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Dirzo, R.; Miranda, A. Altered Patterns of Herbivory and Diversity in the Forest Understory: A Case Study of the Possible Consequences of Contemporary Defaunation. In Plant-Animal Interactions: Evolutionary Ecology in Tropical and Temperate Regions; Price, P.W., Lewinsohn, T.M., Fernandes, G.W., Benson, W.W., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 1991; pp. 273–290. [Google Scholar]
- Stiles, F.G. Coadapted Competitors: The Flowering Seasons of Hummingbird-Pollinated Plants in a Tropical Forest. Science 1977, 198, 1177–1178. [Google Scholar] [CrossRef]
- García-Guzmán, G.; Dirzo, R. Patterns of Leaf-Pathogen Infection in the Understory of a Mexican Rain Forest: Incidence, Spatiotemporal Variation, and Mechanisms of Infection. Am. J. Bot. 2001, 88, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Horvitz, C.; Schemske, D. ST-Seed dispersal of a Neotropical myrmecochore: Variation in removal rates and dispersal distance. Biotropica 1986, 18, 319–323. [Google Scholar] [CrossRef]
- Gilliam, F.; Turrill, N.; Adams, M. Herbaceous-Layer and Overstory Species in Clear-Cut and Mature Central Appalachian Hardwood Forests. Ecol. Appl. 1995, 5, 947–955. [Google Scholar] [CrossRef] [Green Version]
- Levin, D.A.; Wilson, A.C. Rates of Evolution in Seed Plants: Net Increase in Diversity of Chromosome Numbers and Species Numbers through Time. Proc. Natl. Acad. Sci. USA 1976, 73, 2086–2090. [Google Scholar] [CrossRef]
- Spyreas, G.; Matthews, J. Floristic Conservation Value, Nested Understory Floras, and the Development of Second-Growth Forest. Ecol. Appl. 2006, 16, 1351–1366. [Google Scholar] [CrossRef]
- Spicer, M.E.; Radhamoni, H.V.N.; Duguid, M.C.; Queenborough, S.A.; Comita, L.S. Herbaceous plant diversity in forest ecosystems: Patterns, mechanisms, and threats. Plant Ecol. 2022, 223, 117–129. [Google Scholar] [CrossRef]
- Wright, S.J.; Hernandéz, A.; Condit, R. The Bushmeat Harvest Alters Seedling Banks by Favoring Lianas, Large Seeds, and Seeds Dispersed by Bats, Birds, and Wind. Biotropica 2007, 39, 363–371. [Google Scholar] [CrossRef]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef]
- Royo, A.A.; Carson, W.P. The Herb Community in a Tropical Forest in Central Panamá: Dynamics and Impact of Mammalian Herbivores. Oecologia 2005, 145, 66–75. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Major Lineage | Order | Family | Species | IVI |
|---|---|---|---|---|
| Monocots | Alismatales | Araceae | Anthurium flexile | 28.1 |
| Anthurium pentaphyllum | 3.7 | |||
| Anthurium schlesei | 1.1 | |||
| Dieffenbachia seguine | 8.7 | |||
| Monstera acuminata | 15.4 | |||
| Philodendron guatemalensis | 12.4 | |||
| Philodendron inaequilaterum | 8.3 | |||
| Philodendron scandens | 4.9 | |||
| Philodendron tripartitum | 1.1 | |||
| Rhodospatha wendlandii | 30.4 | |||
| Spathiphyllum cochlearispathum | 14.4 | |||
| Syngonium chiapensis | 12.0 | |||
| Syngonium podophyllum | 17.5 | |||
| Zingiberales | Costaceae | Costus dirzoii | 1.1 | |
| Costus scaber | 2.1 | |||
| Marantaceae | Calathea microcephala | 1.1 | ||
| Zingiberaceae | Renealmia mexicana | 1.1 | ||
| Eudicots | Lamiales | Acanthaceae | Schaueria parviflora | 2.3 |
| Aphelandra aurantiaca | 9.5 | |||
| Pteridophyta | Polypodiales | Athyriaceae | Diplazium lonchophyllum | 15.6 |
| Lomariopsidaceae | Bolbitis bernouilli | 5.5 | ||
| Thelypteridaceae | Thelypteris rachyflexuosa | 6.9 | ||
| Schizaeales | Lygodiaceae | Lygodium heterodoxum | 1.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perea, R.; Schroeder, J.W.; Dirzo, R. The Herbaceous Understory Plant Community in the Context of the Overstory: An Overlooked Component of Tropical Diversity. Diversity 2022, 14, 800. https://doi.org/10.3390/d14100800
Perea R, Schroeder JW, Dirzo R. The Herbaceous Understory Plant Community in the Context of the Overstory: An Overlooked Component of Tropical Diversity. Diversity. 2022; 14(10):800. https://doi.org/10.3390/d14100800
Chicago/Turabian StylePerea, Ramón, John W. Schroeder, and Rodolfo Dirzo. 2022. "The Herbaceous Understory Plant Community in the Context of the Overstory: An Overlooked Component of Tropical Diversity" Diversity 14, no. 10: 800. https://doi.org/10.3390/d14100800
APA StylePerea, R., Schroeder, J. W., & Dirzo, R. (2022). The Herbaceous Understory Plant Community in the Context of the Overstory: An Overlooked Component of Tropical Diversity. Diversity, 14(10), 800. https://doi.org/10.3390/d14100800