
Real-Time PCR-Based Method for Sex Determination in Lake Sturgeon (Acipenser fulvescens)



Abstract
:1. Introduction
2. Materials and Methods
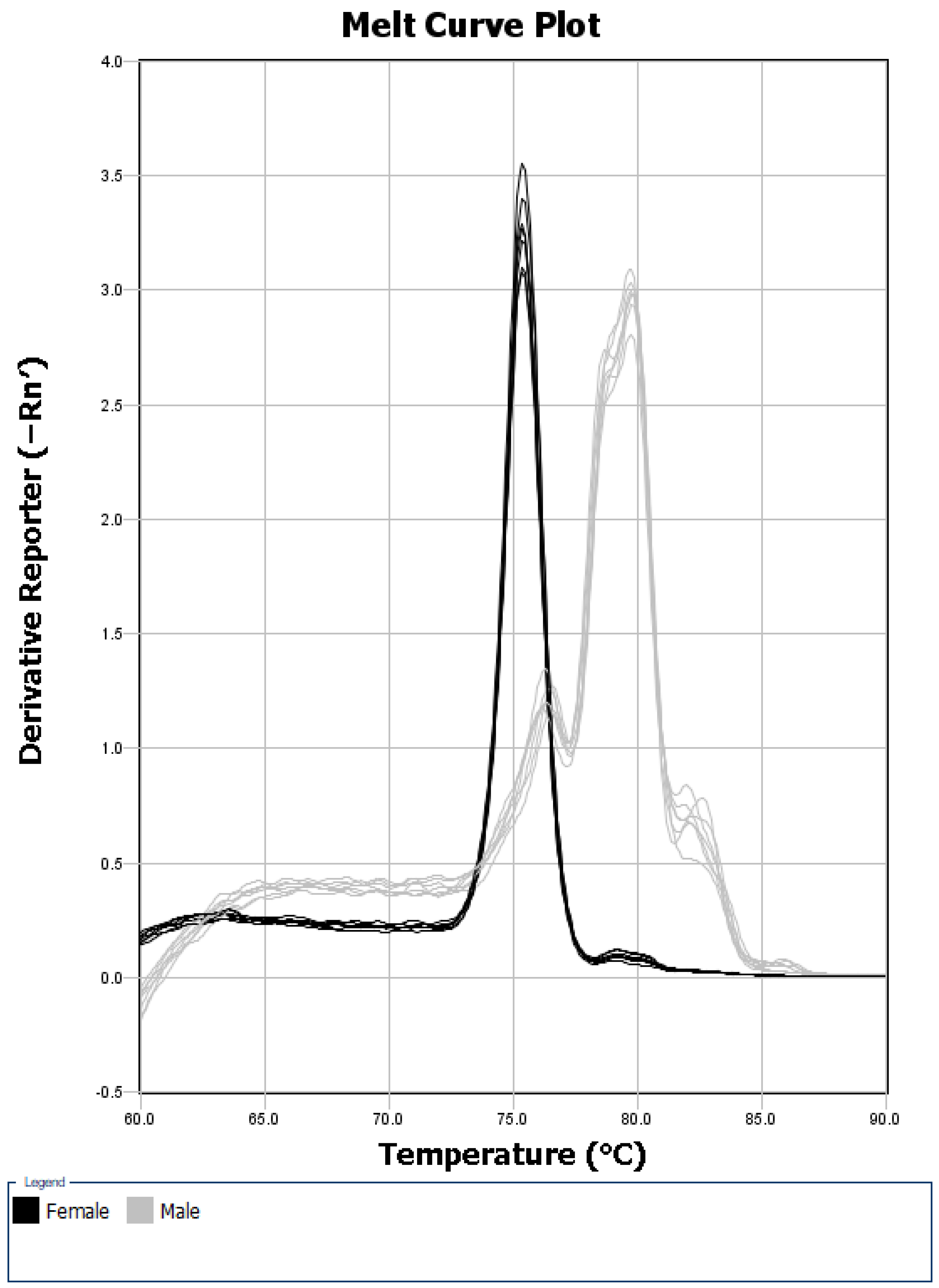
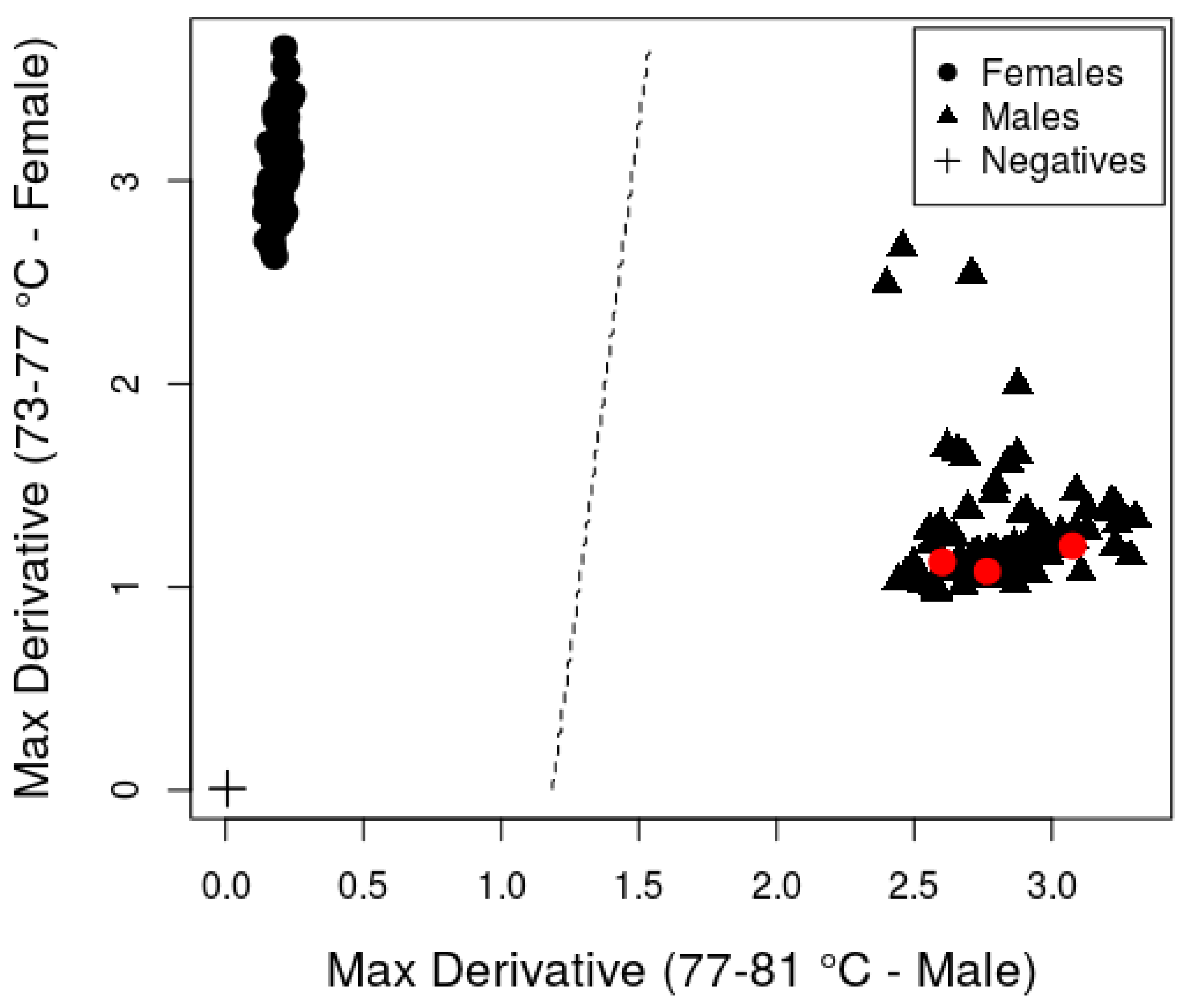
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamel, M.J.; Spurgeon, J.; Steffensen, K.D.; Pegg, M.A. Uncovering unique plasticity in life history of an endangered centenarian fish. Nature Sci. Rep. 2020, 10, 12866. [Google Scholar] [CrossRef] [PubMed]
- Dammerman, K.; Webb, M.; Scribner, K.T. The influence of reproductive timing on female reproductive success and ovarian quality determined using plasma testosterone concentrations in lake sturgeon (Acipenser fulvescens). Can. J. Fish. Aquat. Sci. 2019, 76, 1147–1160. [Google Scholar] [CrossRef] [Green Version]
- Hartl, D.; Clark, A.G. Principles of Population Genetics, 4th ed.; Sinauer Associates: Sunderland, MA, USA, 2006. [Google Scholar]
- Webb MA, H.; Eenennaam JP Van Crossman, J.A.; Chapman, F.A. A practical guide for assigning sex and stage of maturity in sturgeons and paddlefish. J. Appl. Ichthyol. 2019, 35, 169–186. [Google Scholar] [CrossRef] [Green Version]
- Chiotti, J.A.; Boase, J.C.; Hondorp, D.W.; Briggs, A.S. Assigning sex and reproductive stage to adult lake sturgeon using ultrasonography and common morphological measures. Amer. J. Fish. Manag. 2016, 36, 21–29. [Google Scholar] [CrossRef]
- McGuire, J.M.; Bello-Deocampo, D.; Bauman, J.; Baker, E.A.; Scribner, K.T. Histological characteristics of gonadal development of juvenile Lake sturgeon (Acipenser fulvescens). Environ. Biol. Fish. 2019, 102, 969–983. [Google Scholar] [CrossRef]
- Craig, J.M.; Papoulias, D.M.; Thomas, M.V.; Annis, M.L.; Boase, J. Sex assignment of lake sturgeon (Acipenser fulvescens) based on plasma sex hormone and vitellogenin levels. J. Appl. Ichthyol. 2009, 25, 60–67. [Google Scholar] [CrossRef]
- Du, K.; Stöck, M.; Kneitz, S. The sterlet sturgeon genome sequence and the mechanisms of segmental rediploidization. Nat. Ecol. Evol. 2020, 4, 841–852. [Google Scholar] [CrossRef] [Green Version]
- Kuhl, H.; Guiguen, Y.; Höhne, C.; Kreuz, E.; Du, K.; Klopp, C.; Lopez-Roques, C.; Santidrian Yebra-Pimentel, E.; Ciorpac, M.; Gessner, J.; et al. A 180 My-old female-specific genome region in sturgeon reveals the oldest known vertebrate sex determining system with undifferentiated sex chromosomes. bioRxiv 2020. [Google Scholar] [CrossRef]
- Scribner, K.T.; Kanefsky, J. Molecular sexing of lake sturgeon. J. Great Lakes Res. 2021, 47, 934–936. [Google Scholar] [CrossRef]
- Sanfilippo, G.E.; Riedy, J.J.; Larson, D.L.; Scribner, K.T. Multi-year evidence of unbiased sex ratios in hatchery and wild-reared age-0 lake sturgeon (Acipenser fulvescens). J. Great Lakes Res. 2022, 48, 1306–1313. [Google Scholar] [CrossRef]
- Anglès d’Auriac, M.B.; Urke, H.A.; Kristensen, T. A rapid qPCR method for genetic sex identification of Salmo salar and Salmo trutta including simultaneous elucidation of interspecies hybrid paternity by high-resolution melt analysis. J. Fish Biol. 2014, 84, 1971–1977. [Google Scholar] [CrossRef]
- Ripley, B.; Venables, B.; Bates, D.M.; Hornik, K.; Gebhardt, A.; Firth, D.; Ripley, M.B. Package ‘mass’. Cran r 2013, 538, 113–120. [Google Scholar]
- Sakae, Y.; Oikawa, A.; Sugiura, Y.; Mita, M.; Nakamura, S.; Nishimura, T.; Suematsu, M.; Tanaka, M. Starvation causes female-to-male sex reversal through lipid metabolism in the teleost fish, medaka (Olyzias latipes). Biol. Open 2020, 9, bio050054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baroiller, J.F.; Chourrout, D.; Fostier, A.; Jalabert, B. Temperature and sex chromosomes govern sex ratios of the mouthbrooding cichlid fish Oreochromis niloticus. J. Exp. Zool. 1995, 273, 216–223. [Google Scholar] [CrossRef]
- Rzepkowska, M.; Ostaszewska, T.; Gibala, M.; Lukasz Roszko, M. Intersex Gonad Differentiation in Cultured Russian (Acipenser gueldenstaedtii) and Siberian (Acipenser baerii) Sturgeon. Biol. Reprod. 2014, 90, 1–10. [Google Scholar] [CrossRef]
- Rzepkowska, M.; Adamek-Urbańska, D.; Fajkowska, M.; Roszko, M.Ł. Histological Evaluation of Gonad Impairments in Russian Sturgeon (Acipenser gueldenstaedtii) Reared in Recirculating Aquatic System (RAS). Animals 2020, 10, 1439. [Google Scholar] [CrossRef] [PubMed]
- Matsche, M.A.; Rosemary, K.M.; Brundage, H.M., III; O’Herron, J.C. Reproductive demographics, intersex, and altered hormone levels in shortnose sturgeon, Acipenser brevirostrum, from Delaware River, USA. J. Appl. Ichthyol. 2013, 29, 299–309. [Google Scholar] [CrossRef]
- Schooley, J.D.; Geik, A.; Scarnecchia, D.L. First observations of intersex development in paddlefish Polyodon spathula. J. Fish Biol. 2020, 97, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Omoto, N.; Maebayashi, M.; Mitsuhashi, E.; Yoshitomi, K.; Adachi, S.; Yamauchi, K. Effects of estradiol-17β and 17α-methyltestosterone on gonadal sex differentiation in the F2 hybrid sturgeon, the bester. Fish. Sci. 2002, 68, 1047–1054. [Google Scholar] [CrossRef]
- Hassanzadeh Saber, M.; Baradaran Noveiri, S.; Pourkazemi, M.; Yarmohammadi, M.; Jalilpour, J.; Nowruz Fashkhami, M.; Falahatkar, B. Masculinization of the gynogenetic juvenile ship sturgeon (Acipenser nudiventris Lovetsky, 1828) using 17α-methyl testosterone. J. Appl. Ichthyol. 2019, 35, 843–849. [Google Scholar]



{kind=link}
{kind=link}
| Population | Year Collected | Total # | # with Field Sex = M | # with Mol Sex = M | # with Field Sex = F | # with Mol Sex = F | # of Mismatches | Percent with Sex Mismatch | Percent with Both Male & Pronounced Female-Like Peaks Present |
|---|---|---|---|---|---|---|---|---|---|
| Menominee River (MI-WI border) | 2019 | 57 | 36 | 38 | 21 | 19 | 2 # | 3.5% | 0% |
| Black River (MI) | 2020 | 23 | 13 | 14 | 10 | 9 | 1 * | 4.4% | 0% |
| Black River (MI) | 2021 | 63 | 36 | 36 | 27 | 27 | 0 | 0% | 19% |
| Total | 143 | 85 | 88 | 58 | 55 | 3 | 2% | 8.4% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanefsky, J.; Smith, S.; Scribner, K.T. Real-Time PCR-Based Method for Sex Determination in Lake Sturgeon (Acipenser fulvescens). Diversity 2022, 14, 839. https://doi.org/10.3390/d14100839
Kanefsky J, Smith S, Scribner KT. Real-Time PCR-Based Method for Sex Determination in Lake Sturgeon (Acipenser fulvescens). Diversity. 2022; 14(10):839. https://doi.org/10.3390/d14100839
Chicago/Turabian StyleKanefsky, Jeannette, Seth Smith, and Kim T. Scribner. 2022. "Real-Time PCR-Based Method for Sex Determination in Lake Sturgeon (Acipenser fulvescens)" Diversity 14, no. 10: 839. https://doi.org/10.3390/d14100839
APA StyleKanefsky, J., Smith, S., & Scribner, K. T. (2022). Real-Time PCR-Based Method for Sex Determination in Lake Sturgeon (Acipenser fulvescens). Diversity, 14(10), 839. https://doi.org/10.3390/d14100839