



Specific and Intraspecific Diversity of Symphypleona and Neelipleona (Hexapoda: Collembola) in Southern High Appalachia (USA)


Abstract
:1. Introduction
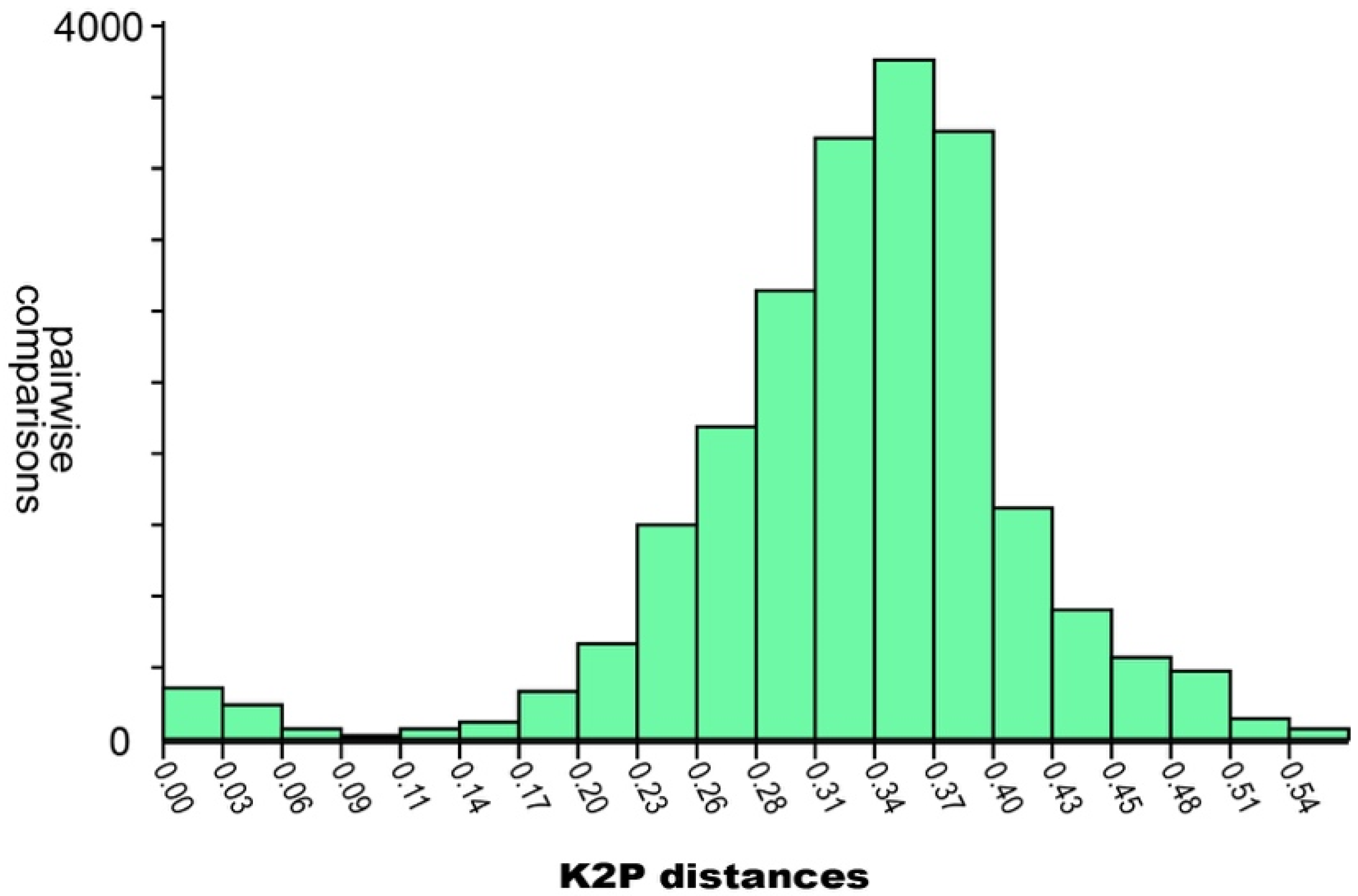
2. Materials and Methods
3. Results and Discussion
3.1. Neelipleona
Neelidae
3.2. Symphypleona
3.2.1. Dicyrtomidae
3.2.2. Arrhopalitidae
3.2.3. Sminthurididae
3.2.4. Katiannidae
3.2.5. Sminthuridae
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barnes, R.; Clark, A.T. Sixty-Five million years of change in temperature and topography explain evolutionary history in eastern North American plethodontid salamanders. Am. Nat. 2017, 190, E1–E12. [Google Scholar] [CrossRef] [PubMed]
- Crespi, E.J.; Rissler, L.J.; Browne, R. Testing Pleistocene refugia theory: Phylogeographical analysis of Desmognathus wrighti, a high-elevation salamander in the southern Appalachians. Mol. Ecol. 2003, 12, 969–984. [Google Scholar] [CrossRef]
- Means, J.C.; Marek, P.E. Is geography an accurate predictor of evolutionary history in the millipede family Xystodesmidae? PeerJ 2017, 5, e3854. [Google Scholar] [CrossRef] [Green Version]
- Carlton, C.; Bayless, V. Documenting Beetle (Arthropoda: Insecta: Coleoptera) Diversity in Great Smoky Mountains National Park: Beyond the Halfway Point. Southeast. Nat. 2007, 6, 182–193. [Google Scholar] [CrossRef]
- Bernard, E.C.; Felderhoff, K.L. Biodiversity explosion: Collembola (springtails) of Great Smoky Mountains National Park. Southeast Nat. 2007, 1, 175–182. [Google Scholar] [CrossRef]
- Barr, T.C. Revision of Appalachian Trechus (Coleoptera: Carabidae). Brimleyana 1979, 2, 29–75. [Google Scholar]
- Gusarov, V.I. A revision of nearctic species of the genus Geostiba Thomson, 1858. Zootaxa 2002, 81, 1–88. [Google Scholar] [CrossRef] [Green Version]
- Peck, S.B. A systematic revision and the evolutionary biology of the Ptomaphagus (Adelops) beetles of North America with emphasis on cave-inhabiting species (Leiodidae). Bull. Mus. Comp. Zool. 1973, 145, 29–162. [Google Scholar]
- Peck, S.B. Systematics and evolution of forest litter Adelopsis in the southern Appalachians (Coleoptera: Leiodidae; Catopinae). Psyche A J. Entomol. 1978, 85, 355–382. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, Q.D.; McHugh, J.V. A New Southern Appalachian Species, Dasycerus Bicolor (Coleoptera: Staphylinidae: Dasycerinae), from Declining Endemic Fir Forests. Coleopt. Bull. 1994, 48, 265–271. [Google Scholar]
- Smolis, A.; Bernard, E.C. Intricatonura fjellbergi, a new peculiar genus and species of Neanurini (Collembola: Neanuridae: Neanurinae) from Great Smoky Mountains National Park. Florida Entomol. 2017, 100, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Keith, R.; Hedin, M. Extreme mitochondrial population subdivision in southern Appalachian paleoendemic spiders (Araneae: Hypochilidae: Hypochilus), with implications for species delimitation. J. Arachnol. 2012, 40, 167–181. [Google Scholar] [CrossRef]
- Hedin, M.; Carlson, D.; Coyle, F. Sky island diversification meets the multispecies coalescent—Divergence in the spruce-fir moss spider (Microhexura montivaga, Araneae, Mygalomorphae) on the highest peaks of southern Appalachia. Mol. Ecol. 2015, 24, 3467–3484. [Google Scholar] [CrossRef]
- Moskwik, M. Recent elevational range expansions in plethodontid salamanders (Amphibia: Plethodontidae) in the southern Appalachian Mountains. J. Biogeogr. 2014, 41, 1957–1966. [Google Scholar] [CrossRef]
- Thomas, S.M.; Hedin, M. Natural history and distribution of the enigmatic southern Appalachian opilionid, Fumontana deprehendor Shear (Laniatores: Triaenonychidae), with an assessment of morphological variation. Zootaxa 2006, 1242, 21–36. [Google Scholar]
- Ulrey, C.; Quintana-Ascencio, P.F.; Kauffman, G.; Smith, A.B.; Menges, E.S. Life at the top: Long-term demography, microclimatic refugia, and responses to climate change for a high-elevation southern Appalachian endemic plant. Biol. Conserv. 2016, 200, 80–92. [Google Scholar] [CrossRef]
- White, P.B.; Van de Gevel, S.L.; Soule, P.T. Succession and disturbance in an endangered red spruce− Fraser fir forest in the southern Appalachian Mountains, North Carolina, USA. Endanger. Species Res. 2012, 18, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, M.A. Impact of the Balsam Woolly Adelgid (Adelges piceae Ratz.) on an Abies fraseri (Pursh) Poir. Dominated Stand near the Summit of Mount LeConte, Tennessee. Castanea 2003, 68, 109–118. [Google Scholar]
- Rusek, J. Biodiversity of Collembola and their functional role in the ecosystem. Biodivers. Conserv. 1998, 7, 1207–1219. [Google Scholar] [CrossRef]
- Filho, L.C.I.O.; Baretta, D. Why should we care about edaphic springtails? Sci. Agrar. 2016, 17, 21–40. [Google Scholar] [CrossRef]
- Hopkin, S.P. Biology of the Springtails; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Christiansen, K.; Bellinger, P. The Collembola of North America, North of the Rio Grande; Grinnell College: Grinnell, IA, USA, 1998. [Google Scholar]
- Folsom, J.W. The distribution of Holarctic Collembola. Psyche 1901, 9, 159–162. [Google Scholar] [CrossRef] [Green Version]
- Ávila-Jiménez, M.L.; Coulson, S.J. A holarctic biogeographical analysis of the collembola (Arthropoda, Hexapoda) unravels recent post-glacial colonization patterns. Insects 2011, 2, 273–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wray, D.L. Collembola of the Great Smoky Mountains. J. Tenn. Acad. Sci. 1963, 38, 85–86. [Google Scholar]
- Snider, R.J. Sminthurus bivittatus, new species from the southeastern United States (Collembola: Sminthuridae). Florida Entomol. 1985, 68, 574–582. [Google Scholar] [CrossRef]
- Snider, R.J. Dicyrtoma (Ptenothrix) renateae, New Species from the Savannah River Plant and Georgia (Collembola: Dicyrtominae). Florida Entomol. 1985, 68, 561–566. [Google Scholar] [CrossRef]
- Snider, R.J. Redescription of Sminthurus floridanus Macgillivray, 1893 (Collembola: Sminthuridae). Florida Entomol. 1982, 65, 221–227. [Google Scholar] [CrossRef]
- Snider, R.J. A new species of Ptenothrix and records from the southeastern United States (Dicyrtomidae: Collembola). Florida Entomol. 1990, 73, 248–257. [Google Scholar] [CrossRef]
- Snider, R.J. Denisiella lithophila, a new species from a granite outcrop in Georgia (Collembola: Sminthurididae). Florida Entomol. 1988, 71, 124–129. [Google Scholar] [CrossRef]
- Snider, R.J. Sminthurus carolinensis, new species from South Carolina (Collembola: Sminthuridae). Florida Entomol. 1981, 64, 417–424. [Google Scholar] [CrossRef]
- Snider, R.J. Sminthurus fischeri, new species from Georgia (Collembola: Sminthuridae). Florida Entomol. 1982, 65, 321–326. [Google Scholar] [CrossRef]
- Snider, R.J. Dicyrtoma (Ptenothrix) castanea, New Species from the Savannah River Plant (Collembola: Dicyrtominae). Florida Entomol. 1985, 68, 582–587. [Google Scholar] [CrossRef]
- Snider, R.J. Vesicephalus crossleyi, New Species from the Savannah River Plant and Georgia (Collembola: Sminthurinae). Florida Entomol. 1985, 68, 567–574. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSalle, R.; Goldstein, P. Review and interpretation of trends in DNA barcoding. Front. Ecol. Evol. 2019, 7, 302. [Google Scholar] [CrossRef] [Green Version]
- Porco, D.; Bedos, A.; Greenslade, P.; Janion, C.; Skarzyński, D.; Stevens, M.I.; Jansen Van Vuuren, B.; Deharveng, L. Challenging species delimitation in Collembola: Cryptic diversity among common springtails unveiled by DNA barcoding. Invertebr. Syst. 2012, 26, 470–477. [Google Scholar] [CrossRef]
- Deharveng, L. Recent advances in Collembola systematics, 6th International Seminar on Apterygota, Siena, Italy, 2002. Pedobiologia 2004, 48, 415–433. [Google Scholar] [CrossRef]
- Hutama, A.; Dahruddin, H.; Busson, F.; Sauri, S.; Keith, P.; Hadiaty, R.K.; Hanner, R.; Suryobroto, B.; Hubert, N. Identifying spatially concordant evolutionary significant units across multiple species through DNA barcodes: Application to the conservation genetics of the freshwater fishes of Java and Bali. Glob. Ecol. Conserv. 2017, 12, 170–187. [Google Scholar] [CrossRef]
- Dincǎ, V.; Runquist, M.; Nilsson, M.; Vila, R. Dispersal, fragmentation, and isolation shape the phylogeography of the European lineages of Polyommatus (Agrodiaetus) ripartii (Lepidoptera: Lycaenidae). Biol. J. Linn. Soc. 2013, 109, 817–829. [Google Scholar] [CrossRef] [Green Version]
- Bellinger, P.F.; Christiansen, K.; Janssens, F. Checklist of the Collembola of the World. Available online: www.collembola.org (accessed on 16 May 2022).
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Elbrecht, V.; Leese, F. Validation and Development of COI Metabarcoding Primers for Freshwater Macroinvertebrate Bioassessment. Front. Environ. Sci. 2017, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Meier, R.; Wong, W.; Srivathsan, A.; Foo, M. $1 DNA barcodes for reconstructing complex phenomes and finding rare species in specimen-rich samples. Cladistics 2016, 32, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, B.; Rood, J.; Singer, E. BBMerge—Accurate paired shotgun read merging via overlap. PLoS ONE 2017, 12, e0185056. [Google Scholar] [CrossRef]
- Srivathsan, A.; Lee, L.; Katoh, K.; Hartop, E.; Kutty, S.N.; Wong, J.; Yeo, D.; Meier, R. ONTbarcoder and MinION barcodes aid biodiversity discovery and identification by everyone, for everyone. BMC Biol. 2021, 19, 217. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadler, T. On incomplete sampling under birth-death models and connections to the sampling-based coalescent. J. Theor. Biol. 2009, 261, 58–66. [Google Scholar] [CrossRef]
- Papadopoulou, A.; Anastasiou, I.; Vogler, A.P. Revisiting the insect mitochondrial molecular clock: The mid-Aegean trench calibration. Mol. Biol. Evol. 2010, 27, 1659–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, A.D.; Taylor, S.J.; Davis, M.A. At the confluence of vicariance and dispersal: Phylogeography of cavernicolous springtails (Collembola: Arrhopalitidae, Tomoceridae) codistributed across a geologically complex karst landscape in Illinois and Missouri. Ecol. Evol. 2018, 8, 10306–10325. [Google Scholar] [CrossRef] [Green Version]
- Lukić, M.; Delić, T.; Pavlek, M.; Deharveng, L.; Zagmajster, M. Distribution pattern and radiation of the European subterranean genus Verhoeffiella (Collembola, Entomobryidae). Zool. Scr. 2020, 49, 86–100. [Google Scholar] [CrossRef]
- Ding, Y.H.; Yu, D.Y.; Guo, W.B.; Li, J.N.; Zhang, F. Molecular phylogeny of Entomobrya (Collembola: Entomobryidae) from China: Color pattern groups and multiple origins. Insect Sci. 2019, 26, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Collins, G.E.; Hogg, I.D.; Convey, P.; Sancho, L.G.; Cowan, D.A.; Lyons, W.B.; Adams, B.J.; Wall, D.H.; Green, T.G. Genetic diversity of soil invertebrates corroborates timing estimates for past collapses of the West Antarctic Ice Sheet. Proc. Natl. Acad. Sci. USA 2020, 117, 22293–22302. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapli, P.; Lutteropp, S.; Zhang, J.; Kobert, K.; Pavlidis, P.; Stamatakis, A.; Flouri, T. Multi-rate Poisson tree processes for single-locus species delimitation under maximum likelihood and Markov chain Monte Carlo. Bioinformatics 2017, 33, 1630–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A.P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, T.; Barraclough, T.G. Delimiting species using single-locus data and the generalized mixed yule coalescent approach: A revised method and evaluation on simulated data sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porco, D.; Skarzyński, D.; Decaëns, T.; Hebert, P.D.N.; Deharveng, L. Barcoding the Collembola of Churchill: A molecular taxonomic reassessment of species diversity in a sub-Arctic area. Mol. Ecol. Resour. 2014, 14, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.D.; Giordano, R.; Soto-Adames, F.N. Operational criteria for cryptic species delimitation when evidence is limited, as exemplified by North American Entomobrya (Collembola: Entomobryidae). Zool. J. Linn. Soc. 2015, 173, 818–840. [Google Scholar] [CrossRef] [Green Version]
- Cicconardi, F.; Nardi, F.; Emerson, B.C.; Frati, F.; Fanciulli, P.P. Deep phylogeographic divisions and long-term persistence of forest invertebrates (Hexapoda: Collembola) in the North-Western Mediterranean basin. Mol. Ecol. 2010, 19, 386–400. [Google Scholar] [CrossRef] [PubMed]
- Cicconardi, F.; Fanciulli, P.P.; Emerson, B.C. Collembola, the biological species concept and the underestimation of global species richness. Mol. Ecol. 2013, 22, 5382–5396. [Google Scholar] [CrossRef] [Green Version]
- Meier, R.; Zhang, G.; Ali, F. The Use of Mean Instead of Smallest Interspecific Distances Exaggerates the Size of the “Barcoding Gap” and Leads to Misidentification. Syst. Biol. 2008, 57, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Bu, W. Exploring Large-Scale Patterns of Genetic Variation in the COI Gene among Insecta: Implications for DNA Barcoding and Threshold-Based Species Delimitation Studies. Insects 2022, 13, 425. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-Based Registry for All Animal Species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeppelini, D. Phylogeny of Arrhopalites s.l. (Collembola: Symphypleona: Arrhopalitidae): Testing the monophyly of the recently erected genera Arrhopalites s.s. and Pygmarrhopalites. Invertebr. Syst. 2011, 25, 91–105. [Google Scholar] [CrossRef]
- Baquero, E.; Snider, R.; Jordana, R. A revision of the spring-tail genus Vesicephalus Richards (Collembola, Symphypleona, Sminthurinae). Syst. Entomol. 2006, 31, 633–647. [Google Scholar] [CrossRef] [Green Version]
- Banks, N. Two new Smynthurids. J. N. Y. Entomol. Soc. 1897, 5, 33–34. [Google Scholar]
- Borrer, D.J.; Triplehorn, C.A.; Johnson, N.F. An Introduction to the Study of Insects; Saunders College Publishing: Philadelphia, PA, USA, 1989. [Google Scholar]
- Zhang, B.; Chen, T.W.; Mateos, E.; Scheu, S.; Schaefer, I. Cryptic species in Lepidocyrtus lanuginosus (Collembola: Entomobryidae) are sorted by habitat type. Pedobiologia 2018, 68, 12–19. [Google Scholar] [CrossRef]
- Porco, D.; Potapov, M.; Bedos, A.; Busmachiu, G.; Weiner, W.M.; Hamra-Kroua, S.; Deharveng, L. Cryptic diversity in the ubiquist species Parisotoma notabilis (Collembola, Isotomidae): A long-used chimeric species? PLoS ONE 2012, 7, e46056. [Google Scholar] [CrossRef] [Green Version]
- Mateos, E.; Winkler, D.; Riutort, M.; Álvarez-Presas, M. New morphological and molecular data reveal an important underestimation of species diversity and indicate evolutionary patterns in European Lepidocyrtus (Collembola: Entomobryidae). Invertebr. Syst. 2021, 35, 471–492. [Google Scholar] [CrossRef]
- Hartop, E.; Srivathsan, A.; Ronquist, F.; Meier, R. Towards large-scale integrative taxonomy (LIT): Resolving the data conundrum for dark taxa. Syst. Biol. 2022, syac033. [Google Scholar] [CrossRef]
- Page, R.D.M. DNA barcoding and taxonomy: Dark taxa and dark texts. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150334. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Number on Figure 1 | Morphosp Abbrev | State | Lat. | Long. | Elevation (m) |
|---|---|---|---|---|---|---|
| Mount Rogers | 1 | MRg | VA | 36.6602 | −81.5447 | 1746 |
| Whitetop | 2 | WT | VA | 36.6388 | −81.6062 | 1682 |
| Grandfather Mountain | 3 | GrM | NC | 36.1118 | −81.8105 | 1812 |
| Grassy Ridge Bald | 4 | GRB | TN/NC | 36.1001 | −82.0818 | 1878 |
| Roan High Bluff | 5 | RHB | NC | 36.0933 | −82.1447 | 1910 |
| Roan High Knob | 6 | RHK | TN/NC | 36.1045 | −82.1224 | 1916 |
| Big Bald | 7 | BgBld | TN/NC | 35.9893 | −82.4903 | 1681 |
| Celo Knob | 8 | CK | NC | 35.8527 | −82.2487 | 1928 |
| Big Tom | 9 | BT | NC | 35.7798 | −82.2599 | 2006 |
| Mount Mitchell | 10 | MM | NC | 35.7644 | −82.2641 | 2037 |
| Big Cataloochee Mountain | 11 | BCat | NC | 35.6722 | −83.1756 | 1876 |
| Mount Kephart | 12 | MK | NC/TN | 35.6311 | −83.3895 | 1895 |
| Newfound Gap (off Hwy 441) | 13 | Hwy | TN/NC | 35.624 | −83.4163 | 1394 |
| Clingmans Dome | 14 | CD | TN/NC | 35.5623 | −83.5036 | 2025 |
| Browning Knob | 15 | BrK | NC | 35.463 | −83.1319 | 1902 |
| Mount Lyn Lowry | 16 | LL | NC | 35.4635 | −83.1107 | 1902 |
| Richland Balsam | 17 | RB | NC | 35.363 | −82.989 | 1954 |
| Mount Hardy | 18 | MHy | NC | 35.3032 | −82.9276 | 1865 |
| Black Balsam Knob | 19 | BBK | NC | 35.3258 | −82.8777 | 1894 |
| Huckleberry Knob | 20 | HKnb | NC | 35.3216 | −83.9929 | 1683 |
| Cowee Bald | 21 | CB | NC | 35.3287 | −83.3366 | 1506 |
| Brasstown Bald | 22 | BBld | GA | 34.8763 | −83.8107 | 1457 |
| Rabun Cliffs | 23 | RC | GA | 34.9707 | −83.3008 | 1320 |
| Sassafras Mt. | 24 | Sass | SC | 35.0658 | −82.7763 | 1083 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dukes, C.D.; Janssens, F.; Recuero, E.; Caterino, M.S. Specific and Intraspecific Diversity of Symphypleona and Neelipleona (Hexapoda: Collembola) in Southern High Appalachia (USA). Diversity 2022, 14, 847. https://doi.org/10.3390/d14100847
Dukes CD, Janssens F, Recuero E, Caterino MS. Specific and Intraspecific Diversity of Symphypleona and Neelipleona (Hexapoda: Collembola) in Southern High Appalachia (USA). Diversity. 2022; 14(10):847. https://doi.org/10.3390/d14100847
Chicago/Turabian StyleDukes, Caroline D., Frans Janssens, Ernesto Recuero, and Michael S. Caterino. 2022. "Specific and Intraspecific Diversity of Symphypleona and Neelipleona (Hexapoda: Collembola) in Southern High Appalachia (USA)" Diversity 14, no. 10: 847. https://doi.org/10.3390/d14100847
APA StyleDukes, C. D., Janssens, F., Recuero, E., & Caterino, M. S. (2022). Specific and Intraspecific Diversity of Symphypleona and Neelipleona (Hexapoda: Collembola) in Southern High Appalachia (USA). Diversity, 14(10), 847. https://doi.org/10.3390/d14100847