
Diversity, Phylogenetic Relationships and Distribution of Marsh Frogs (the Pelophylax ridibundus complex) from Kazakhstan and Northwest China

 , ,
, ,  , , , , and
, , , , and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Laboratory Protocol
2.3. Phylogenetic Reconstruction
2.4. Bayesian Molecular Dating
2.5. Population Structure and Demographic History
3. Results
3.1. Data Analysis
3.1.1. Sequence Characteristics
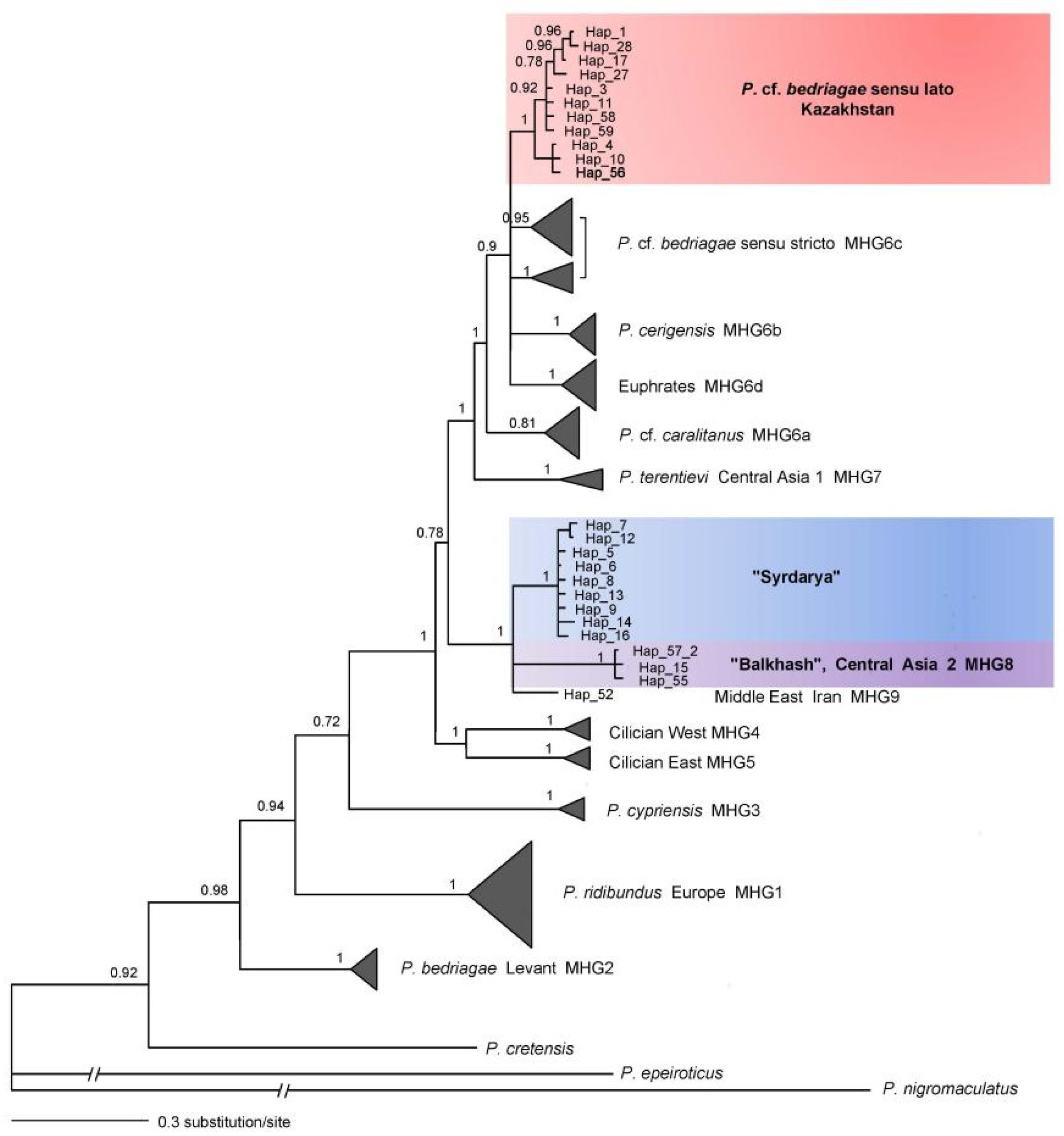
3.1.2. Combined Mitochondrial DNA Phylogeny
3.1.3. Nuclear DNA Phylogeny
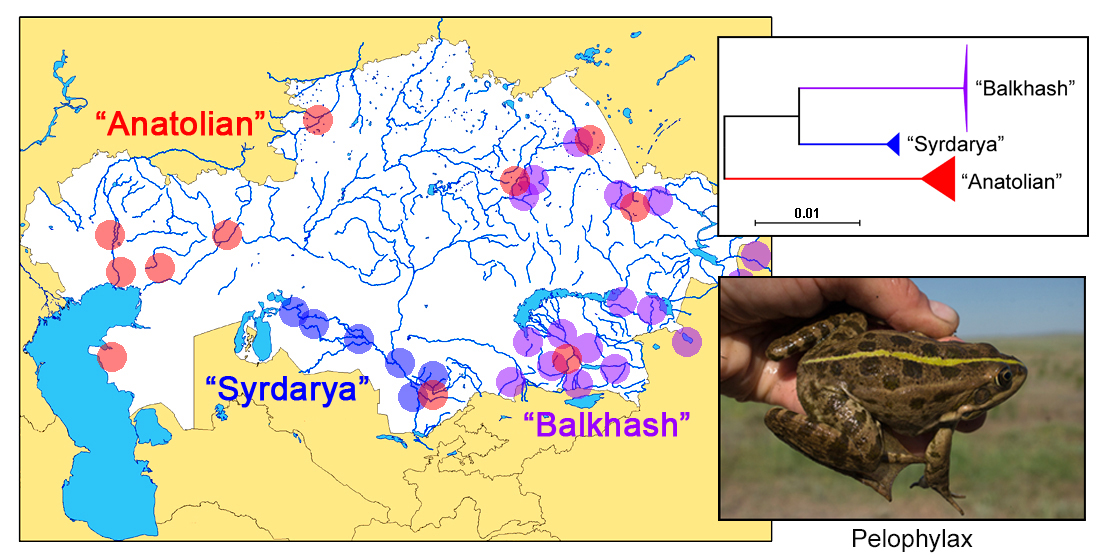
3.2. Distributional Pattern
3.3. Molecular Dating
3.4. Genetic Structure and Demographic History According to mtDNA Data
4. Discussion
4.1. Species Distributional Ranges
4.2. Balkhash and Syrdarya—mtDNA Clades of P. sp. novum
4.3. Anatolian Marsh Frog (P. cf. bedriagae)
4.4. Taxonomic and Conservation Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frost, D. Amphibian Species of the World: An Online Database. 2021. Available online: https://amphibiansoftheworld.amnh.org/ (accessed on 1 January 2022).
- Zhao, E.M.; Adler, K. Herpetology of China; Society for the Study of Amphibians and Reptiles: Oxford, OH, USA, 1993. [Google Scholar]
- Borkin, L.Y.; Litvinchuk, S.N.; Rozanov, Y.M.; Skorinov, D.V. Cryptic species (a case study of amphibians). Entomol. Rev. 2004, 84, 75–98. [Google Scholar]
- Plötner, J. Die Westpaläarktische Wasserfrösche; Laurenti-Verlag: Bielefeld, Germany, 2005; p. 160. [Google Scholar]
- Hotz, H.; Beerli, P.; Uzzell, T.; Guex, G.-D.; Pruvost, N.B.M.; Schreiber, R.; Plötner, J. Balancing a cline by influx of migrants: A genetic transition in water frogs of Eastern Greece. J. Hered. 2012, 104, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Plötner, J. Genetic diversity in mitochondrial 12S of western Palearctic water frog (Anura, Ranidae) and implications for their systematics. J. Zool. Syst. Evol. Res. 1998, 36, 191–201. [Google Scholar] [CrossRef]
- Lymberakis, P.; Poulokakis, N.; Manthalou, G.; Tsigenopoulos, C.S.; Magoulas, A.; Mylonas, M. Mitochondrial phylogeography of Rana (Pelophylax) populations in the Eastern Mediterranean region. Mol. Phylogenet. Evol. 2007, 44, 115–125. [Google Scholar] [CrossRef]
- Akin, Ҫ.; Bilgin, C.C.; Beerli, P.; Westaway, R.; Ohst, T.; Litvinchuk, S.N.; Uzzell, T.; Bilgin, M.; Hotz, H.; Guex, G.D.; et al. Phylogeographic patterns of genetic diversity in eastern Mediterranean water frogs were determined by geological processes and climate change in the Late Cenozoic. J. Biogeogr. 2010, 37, 2111–2124. [Google Scholar] [CrossRef] [Green Version]
- Plötner, J.; Baier, F.; Akın, C.; Mazepa, G.; Schreiber, R.; Beerli, P.; Litvinchuk, S.N.; Bilgin, C.C.; Borkin, L.; Uzzell, T. Genetic data reveal that water frogs of Cyprus (genus Pelophylax) are an endemic species of Messinian origin. Zoosyst. Evol. 2012, 88, 261–283. [Google Scholar] [CrossRef]
- Plötner, J.; Ohst, T. New hypothesis on the systematic of the Palearctic water frog complex (Anura, Ranidae). Zoosyst. Evol. 2001, 77, 5–21. [Google Scholar] [CrossRef]
- Pesarakloo, A.; Rastegar-Pouyani, E.; Rastegar-Pouyani, N.; Kami, H.; Khosravani, A.; Oraie, H. The first taxonomic revaluation of the Iranian water frogs of the genus Pelophylax (Anura: Ranidae) using sequences of the mitochondrial genome. Mitochondrial DNA 2017, 28, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Dufresnes, C.; Litvinchuk, S.N. Diversity, distribution and molecular species delimitation in frogs and toads from the Eastern Palearctic. Zool. J. Linn. Soc. 2022, 195, 695–760. [Google Scholar] [CrossRef]
- Plötner, J.; Uzzell, T.; Beerli, P.; Akin, Ç.; Bilgin, C.C.; Haefeli, C.; Ohst, T.; Köhler, F.; Schreiber, R.; Guex, G.-D.; et al. Genetic divergence and evolution of reproductive isolation in eastern Mediterranean water frogs. In Evolution in Action: Case Studies in Adaptive Radiation and the Origin of Biodiversity; Glaubrecht, M., Schneider, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 373–403. [Google Scholar] [CrossRef]
- Mazepa, G. Evolution of Water Frogs Pelophylax in Central Asia: How Hybridization and Mitochondrial Introgression among Ecologically Divergent Species Promote Occupation of Novel Environment. Master’s Thesis, Uppsala University, Uppsala, Sweden, 2013. Available online: https://www.researchgate.net/publication/361727081 (accessed on 19 August 2022).
- Ivanov, A.Y.; Ruchin, A.B.; Fayzulin, A.I.; Chikhlyaev, I.V.; Litvinchuk, S.N.; Kirillov, A.A.; Svinin, A.O.; Ermakov, O.A. The first record of natural transfer of mitochondrial DNA from Pelophylax cf. bedriagae into P. lessonae (Amphibia, Anura). Nat. Conserv. Res. 2019, 4, 125–128. [Google Scholar] [CrossRef]
- Plötner, J.; Köhler, F.; Uzzell, T.; Beerli, P.; Schreiber, R.; Guex, G.-D.; Hotz, H. Evolution of serum albumin intron-1 is shaped by a 50 truncated non-long terminal repeat retrotransposon in western Palearctic water frogs (Neobatrachia). Mol. Phylogenet. Evol. 2009, 53, 784–791. [Google Scholar] [CrossRef] [Green Version]
- Ermakov, O.A.; Fayzulin, A.I.; Zaks, M.M.; Kaibeleva, E.I.; Zaripova, F.F. Distribution of the “western” and “eastern” forms of marsh frog Pelophylax ridibundus s.l. in Samara and Saratov region (on data of analysis of mtDNA and nDNA). Izv. Samara Sci. Cent. RAS 2014, 16, 409–412. (In Russian) [Google Scholar]
- Dufresnes, C.; Mazepa, G. Hybridogenesis in water frogs. eLS 2020, 1, 718–726. [Google Scholar] [CrossRef]
- Kuzmin, S.; Tarkhnishvili, D.; Ishchenko, V.; Dujsebayeva, T.; Tuniyev, B.; Papenfuss, T.; Beebee, T.; Ugurtas, I.H.; Sparreboom, M.; Rastegar-Pouyani, N.; et al. Pelophylax ridibundus; The IUCN Red List of Threatened Species 2009; IUCN: Gland, Switzerland, 2009; p. e.T58705A11825745. [Google Scholar] [CrossRef]
- Mezhzherin, S.V.; Peskov, V.N. Biochemical variability and genetic differentiation of populations of the marsh frog Rana ridibunda Pallas. Tsitol. Genet. 1992, 26, 43–48. [Google Scholar]
- Mezhzherin, S.V. Genetic differentiation and species identity of marsh frog Rana ridibunda (Amphibia, Ranidae) from Eastern Kazakhstan. Zool. Zhurnal 1997, 76, 933–939. (In Russian) [Google Scholar]
- Wei, G.; Xu, N.; Li, D.; Wu, M. Karyotypes of two Rana from Xinjiang, China. Asiat. Herpetol. Res. 1992, 4, 141–145. [Google Scholar]
- Fei, L.; Ye, C.Y.; Jiang, J.P. Colored Atlas of Chinese Amphibians and Their Distributions; Sichuan Science and Technology Press: Chengdu, China, 2012. (In Chinese) [Google Scholar]
- Fei, L. Atlas of Amphibians in China; Field Edition; Henan Science Press: Zhengzhou, China, 2020; (In Chinese with Latin). [Google Scholar]
- Ma, J.F. A new record of the genus Rana in Chinese Lake frog. Nat. Hist. 1979, 1, 39. (In Chinese) [Google Scholar]
- Xiang, L.Y.; Huang, R.X. Two new records of amphibians in Xinjiang. J. Xinjiang Univ. (Nat. Sci. Ed.) 1980, 6, 82–85. (In Chinese) [Google Scholar]
- Xu, S.K.; Xiang, L.Y.; Fu, X.N.; Su, F. Preliminary analysis on the feeding habits of three species of anuran amphibians in the Yili area. J. Xinjiang Univ. (Nat. Sci. Ed.) 1983, 3, 68–69. (In Chinese) [Google Scholar]
- Fei, L. Chinese Amphibian Illustrated Guide; Henan Science and Technology Press: Zhengzhou, China, 1999. (In Chinese) [Google Scholar]
- Ye, C.Y.; Fei, L.; Hu, S.Q. Rare and Economical Amphibians of China; Sichuan Science and Technology Press: Chengdu, China, 1993. (In Chinese) [Google Scholar]
- Dubois, A. Notes sur la classification des Ranidae (Amphibiens Anoures). Bull. Mens. Soc. Linn. Lyon 1992, 61, 305–352. [Google Scholar] [CrossRef] [Green Version]
- Fei, L.; Ye, C.Y.; Jian, J.; Xie, F.; Huang, Y. An Illustrated Key to Chinese Amphibians; Sichuan Science and Technology Press: Chengdu, China, 2005. (In Chinese) [Google Scholar]
- Fei, L.; Ye, C.Y.; Jiang, J.P. Colored Atlas of Chinese Amphibians; Sichuan Science and Technology Press: Chengdu, China, 2010. (In Chinese) [Google Scholar]
- Li, D. Wild Life in Xinjiang China; Xinjiang Youth Press: Urumqi, China, 2000. (In Chinese) [Google Scholar]
- AmphibiaChina. The Database of Chinese Amphibians; Kunming Institute of Zoology (CAS): Kunming, China, 2022; Available online: http://www.amphibiachina.org/ (accessed on 12 January 2022)(Citation in English).
- Che, J.; Chen, H.M.; Yang, J.X.; Jin, J.Q.; Jiang, K.; Yuan, Z.Y.; Murphy, R.W.; Zhang, Y.P. Universal COI primers for DNA barcoding amphibians. Mol. Ecol. Res. 2012, 12, 247–258. [Google Scholar] [CrossRef]
- Ye, X.F.; Yuan, L.; Lu, X.F.; He, L.Z.; Wang, X.L.; Ji, R. Genetic diversity and phylogeny of frogs in Xinjiang. Biotechnology 2015, 25, 558–563. (In Chinese) [Google Scholar]
- Ualiyeva, D.A.; Ivanov, A.Y.; Ermakov, O.A. A development of a PCR-RFLP test system for the identification of mitochondrial lines of the Pelophylax ridibundus lake frog in Kazakhstan. Peгuoн 2022, 1, 76–84. (In Russian) [Google Scholar] [CrossRef]
- Ohst, T. Genetische Einflüsse Allochthoner Wasserfrösche auf Endemische Wasserfrosch Populationen (R. kl. Esculenta Komplex); Humboldt-Universitat: Berlin, Germany, 2008. [Google Scholar]
- Akin, Ҫ. Molecular Evolution and Phylogeography of the Eastern Mediterranean Water Frog (Pelophylax) Complex. Ph.D. Thesis, School of Natural and Applied Sciences of Middle East Technical University, Ankara, Turkey, 2015. [Google Scholar]
- Dujsebayeva, T.N.; Ivanov, A.Y.; Kaptyonkina, A.G.; Ualiyeva, D.A.; Krainyuk, V.N.; Cherednichenko, A.V.; Khromov, V.A. The marsh frogs (Pelophylax ridibundus complex) in Central Kazakhstan: Expansion and retreat. Russ. J. Ecosyst. Ecol. 2021, 6, 83–100. [Google Scholar] [CrossRef]
- Bellati, A.; Bassu, L.; Nulchis, V.; Corti, C. Detection of alien Pelophylax species in Sardinia (western Mediterranean, Italy). BioInvasions Rec. 2019, 8, 8–25. [Google Scholar] [CrossRef]
- Duysebaeva, T.N.; Berezovikov, N.N.; Brushko, Z.K.; Kubykin, R.A.; Khromov, V.A. Marsh frog (Rana ridibunda Pallas, 1771) in Kazakhstan: Range changing and recent distribution. Curr. Stud. Herpetol. 2005, 3–4, 29–59. (In Russian) [Google Scholar]
- Dubey, S.; Dufresnes, C. An extinct vertebrate preserved by its living hybridogenetic descendant. Sci. Rep. 2017, 7, 12768. [Google Scholar] [CrossRef]
- Dubey, S.; Leuenberger, J.; Perrin, N. Multiple origins of invasive and ‘native’ water frogs (Pelophylax spp.) in Switzerland. Biol. J. Linn. Soc. 2014, 112, 442–449. [Google Scholar] [CrossRef]
- Dufresnes, C.; Leuenberger, J.; Amrhein, V.; Bühler, C.; Thiébaud, J.; Bohnenstengel, T.; Dubey, S. Invasion genetics of marsh frogs (Pelophylax ridibundus sensu lato) in Switzerland. Biol. J. Linn. Soc. 2018, 123, 402–410. [Google Scholar] [CrossRef]
- Hoffmann, A.; Plötner, J.; Pruvost, N.B.M.; Christiansen, D.G.; Röthlisberger, S.; Choleva, L.; Mikulíček, P.; Cogălniceanu, D.; SasKovács, I.; Shabanov, D.; et al. Genetic diversity and distribution patterns of diploid and polyploid hybrid water frog populations (Pelophylax esculentus complex) across Europe. Mol. Ecol. 2015, 24, 4371–4391. [Google Scholar] [CrossRef]
- Holsbeek, G.; Mergeay, J.; Hotz, H.; Plötner, J.; Volckaert, A.M.; De Meester, L. A cryptic invasion within an invasion and widespread introgression in the European water frog complex: Consequences of uncontrolled commercial trade and weak international legislation. Mol. Ecol. 2008, 17, 5023–5035. [Google Scholar] [CrossRef]
- Holsbeek, G.; Maes, G.E.; De Meester, L.; Volckaert, F.A.M. Conservation of the introgressed European water frog complex using molecular tools. Mol. Ecol. 2009, 18, 1071–1087. [Google Scholar] [CrossRef]
- Holsbeek, G.; Mergeay, J.; Volckaert, F.; De Meester, L. Genetic detection of multiple exotic water frog species in Belgium illustrates the need for monitoring and immediate action. Biol. Invasions 2010, 12, 1459–1463. [Google Scholar] [CrossRef]
- Litvinchuk, S.N.; Ivanov, A.Y.; Lukonina, S.A.; Ermakov, O.A. A record of two alien Pelophylax species and widespread mitochondrial DNA transfer in Kaliningradskaya oblast’ (the Baltic coast, Russia). BioInvasions Rec. 2020, 9, 599–617. [Google Scholar] [CrossRef]
- Lyapkov, S.M.; Ermakov, O.A.; Titov, S.V. Distribution and origin of two forms of the marsh frog Pelophylax ridibundus complex (Anura, Ranidae) from Kamchatka based on mitochondrial and nuclear DNA data. Biol. Bull. 2018, 45, 699–705. [Google Scholar] [CrossRef]
- Svinin, A.O.; Dedukh, D.V.; Borkin, L.J.; Ermakov, O.A.; Ivanov, A.Y.; Litvinchuk, J.S.; Zamaletdinov, R.I.; Mikhaylova, R.I.; Trubyanov, A.B.; Skorinov, D.V.; et al. Genetic structure, morphological variation, and gametogenic peculiarities in water frogs (Pelophylax) from northeastern European Russia. J. Zool. Syst. Evol. Res. 2021, 59, 646–662. [Google Scholar] [CrossRef]
- Vershinin, V.L.; Sitnikov, I.A.; Vershinina, S.D.; Trofimov, A.G.; Lebedinsky, A.A.; Miura, I.J. Mitochondrial heteroplasmy in marsh frog (Pelophylax ridibundus Pallas, 1771). Russ. J. Genet. 2019, 55, 1041–1045. [Google Scholar] [CrossRef]
- Ermakov, O.A.; Zaks, M.M.; Titov, S.V. Diagnostics and distribution of “western” and “eastern” forms of the marsh frog Pelophylax ridibundus s. l. in the Penza Province (on data of analysis of the mtDNA cytochrome c oxidase). Vestn. Tambov Univ. 2013, 18, 2999–3002. (In Russian) [Google Scholar]
- Svinin, A.O.; Ivanov, A.Y.; Zaks, M.M.; Litvinchuk, S.N.; Borkin, L.J.; Rosanov, J.M.; Ermakov, O.A. Distribution of the “eastern” and “western” forms of the marsh frog, Pelophylax ridibundus, and their participation in the origin of hemiclonal hybrids, P. esculentus in Mari El Republic. Curr. Stud. Herpetol. 2016, 15, 120–129. (In Russian) [Google Scholar]
- Ivanov, A.Y. Molecular-Genetic and Ecological Features of the Distribution of Cryptic Forms of Marsh Frog in the Eastern Part of the Range. Ph.D. Thesis, Penza State University, Penza, Russia, 2019. (In Russian). [Google Scholar]
- Ermakov, O.A.; Fayzulin, A.I.; Askenderov, A.D.; Ivanov, A.Y. Molecular-genetic characteristics of marsh frog from the Republic of Dagestan (based on mitochondrial and nuclear DNA data). Izv. Samara Sci. Cent. RAS 2016, 18, 94–97. (In Russian) [Google Scholar]
- Ermakov, O.A.; Simonov, E.P.; Ivanov, A.Y.; Zamaletdinov, R.I.; Fayzulin, A.I. Genetic characteristics of marsh frog (Pelophylax ridibundus complex) from the Western Caucasus based on mitochondrial and nuclear DNA data. In Molecular genetics of aquatic organisms. Trans. I.D. Papanin Inst. Biol. Inland Waters RAS 2016, 73, 70–76. (In Russian) [Google Scholar]
- Ivanov, A.Y.; Korzikov, V.A.; Alekseev, S.K.; Ermakov, O.A. Molecular genetic characteristics of the marsh frogs Pelophylax ridibundus s.l. from the Upper Oka region. In Modern Problems of Zoology, Ecology and Conservancy, Materials of the Readings and Scientific Conference Devoted to Memory of Professor Andrey Grigoryevich Bannikov, and to the 100 Anniversaries from the Date of Its Birth; Vasilevich, F.I., Spitsin, V.V., Popov, S.V., Eds.; Moscow Zoo: Moscow, Russia, 2015; pp. 228–232. (In Russian) [Google Scholar]
- Faizulin, A.I.; Kukushkin, O.V.; Ivanov, A.Y.; Ermakov, O.A. Preliminary data on the molecular genetic structure of Pelophylax ridibundus (Amphibia: Anura: Ranidae) from the southern part of the Crimean Peninsula, based on mitochondrial and nuclear DNA analysis. Curr. Stud. Herpetol. 2017, 17, 56–65. (In Russian) [Google Scholar] [CrossRef]
- Kukushkin, O.V.; Ivanov, A.Y.; Ermakov, O.A. Genetic heterogeneity of the marsh frog (Pelophylax ridibundus complex; Anura, Ranidae) population in Crimea revealed by mitochondrial and nuclear DNA analyses. Univ. Proc. Volga Reg. 2018, 32–54. (In Russian) [Google Scholar] [CrossRef]
- Kaptyonkina, A.G.; Dujsebayeva, T.N.; Akhmedenov, K.M.; Khromov, V.A.; Krainyuk, V.N.; Sarzhanov, F.; Starikov, S.V.; Tarasovskaya, N.E.; Timoshenko, A.Y.; Titov, S.V. The range of marsh frogs (complex Pelophylax ridibundus, Amphibia, Ranidae) in Kazakhstan: Progressive dispersal or cyclic fluctuations? Proc. Zool. Inst. Russ. Acad. Sci. 2022, 326, 211–237 (In Russian). [Google Scholar] [CrossRef]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt extraction of high-quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Meyer, A. Evolution of mitochondrial DNA in fishes. In Molecular Biology Frontiers, Biochemistry and MOLECULAR Biology of Fishes; Elsevier Press: New York, NY, USA, 1993; Volumn 2, pp. 1–38. [Google Scholar]
- Lissovsky, A.A.; Obolenskaya, E.V.; Abramson, N.I.; Dokuchaev, N.E.; Yakimenko, V.V.; Mal’kova, M.G.; Bogdanov, A.S.; Ivanova, N.V. Geographic variation of Microtus middendorffii (Cricetidae, Arvicolinae, Rodentia) sensu lato studied by craniometrical and mitochondrial features. Russ. J. Theriol. 2010, 9, 71–81. [Google Scholar] [CrossRef]
- Ermakov, O.; Ivanov, A.; Titov, S.; Svinin, A.; Litvinchuk, S. New multiplex PCR method for identification of East European green frog species and their hybrids. Russ. J. Herpetol. 2019, 26, 367–370. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.V.D.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Plötner, J.; Uzzell, T.; Beerli, P.; Spolsky, C.; Ohst, T.; Litvinchuk, S.N.; Guex, G.-D.; Reyer, H.-U.; Hotz, H. Widespread unidirectional transfer of mitochondrial DNA: A case in western Palaearctic water frogs. J. Evol. Biol. 2008, 21, 668–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, G. Estimating the Dimension of a Model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Institute of Evolutionary Biology, University of Edinburgh, Edinburgh. FigTree v1.3.1. 2010. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 June 2022).
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Meulenkamp, J.E. Aspects of the late Cenozoic evolution of the Aegean region. In Geological Evolution of the Mediterranean Basin; Stanley, D.J., Wezel, F.C., Eds.; Springer: New York, NY, USA, 1985; pp. 307–321. [Google Scholar]
- Dermitzakis, D.M. The colonisation of Aegean islands in relation with the paleogeographic evolution. Biol. Gallo Hell. 1990, 17, 99–130. [Google Scholar]
- Yang, W.; Feiner, N.; Pinho, C.; While, G.M.; Kaliontzopoulou, A.; Harris, D.J.; Salvi, D.; Uller, T. Extensive introgression and mosaic genomes of Mediterranean endemic lizards. Nat. Commun. 2021, 12, 2762. [Google Scholar] [CrossRef] [PubMed]
- Cohen, K.M.; Finney, S.C.; Gibbard, P.L.; Fan, J.-X. The ICS International Chronostratigraphic Chart. Episodes 2013, 36, 199–204. Available online: https://stratigraphy.org/timescale/ (accessed on 10 June 2022). [CrossRef] [PubMed] [Green Version]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Harpending, H. Signature of ancient population growth in a low-resolution mitochondrial DNA mismatch distribution. Hum. Biol. 1994, 66, 591–600. [Google Scholar]
- Ramos-Onsins, S.E.; Rozas, J. Statistical properties of new neutrality tests against population growth. Mol. Biol. Evol. 2002, 19, 2092–2100. [Google Scholar] [CrossRef] [Green Version]
- Stephens, M.; Smith, N.J.; Donnelly, P. A new statistical method for haplotype reconstruction from population data. Am. J. Hum. Genet. 2001, 68, 978–989. [Google Scholar] [CrossRef] [Green Version]
- Stephens, M.; Donnelly, P. A comparison of Bayesian methods for haplotype reconstruction from population genotype data. Am. J. Hum. Genet. 2003, 73, 1162–1169. [Google Scholar] [CrossRef] [Green Version]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar]
- Kukla, G.J. Pleistocene land–sea correlations I. Europe. Earth. Sci. Rev. 1977, 13, 307–374. [Google Scholar] [CrossRef]
- Naidina, O.; Richards, K. The Akchagylian stage (late Pliocene-early Pleistocene) in the North Caspian region: Pollen evidence for vegetation and climate change in the Urals-Emba region. Quat. Int. 2018, 540, 22–37. [Google Scholar] [CrossRef]
- Kostenko, N.P. Relief Development of the Highland (on the Example of Middle Asia); Mysl’: Moscow, Russia, 1970. (In Russian) [Google Scholar]
- Aubekerov, B.; Gorbunov, A.P. Quaternary permafrost and mountain glaciation in Kazakhstan. Permafr. Perigl. Processes 1999, 10, 65–80. [Google Scholar] [CrossRef]
- Trifonov, V.G.; Ivanova, T.P.; Bachmanov, D.M. Recent transformation of the Central Alpine-Himalayan belt. Geotektonika 2012, 5, 3–21, (In Russian with English abstract). [Google Scholar]
- Svitoch, A.A. The Great Caspian Region: Its Structure and History of Development; Moscow State Univ. Press: Moscow, Russia, 2014; p. 272, (In Russian with English Abstract). [Google Scholar]
- Krijgsman, W.; Tesakov, A.; Yanina, T.; Lazarev, S.; Danukalova, G.; Van Baak, C.G.C.; Agustí, J.; Alçiçek, M.C.; Aliyeva, E.; Bista, D.; et al. Quaternary time scales for the Pontocaspian domain: Interbasinal connectivity and faunal evolution. Earth. Sci. Rev. 2019, 188, 1–40. [Google Scholar] [CrossRef]
- Kollár, J.; Poulíčková, A.; Dvořák, P. On the relativity of species, or the probabilistic solution to the species problem. Mol. Ecol. 2021, 31, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Dufresnes, C.; Brelsford, A.; Jeffries, D.L.; Mazepa, G.; Suchan, T.; Canestrelli, D.; Nicieza, A.; Fumagalli, L.; Dubey, S.; Martínez-Solano, I.; et al. Mass of genes rather than master genes underlie the genomic architecture of amphibian speciation. Proc. Natl. Acad. Sci. USA 2021, 118, e2103963118. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| 2 | 0.047 | ||||||||
| 3 | 0.036 | 0.031 | |||||||
| 4 | 0.067 | 0.079 | 0.070 | ||||||
| 5 | 0.068 | 0.080 | 0.074 | 0.063 | |||||
| 6 | 0.061 | 0.079 | 0.073 | 0.072 | 0.075 | ||||
| 7 | 0.042 | 0.055 | 0.049 | 0.071 | 0.070 | 0.069 | |||
| 8 | 0.013 | 0.046 | 0.036 | 0.064 | 0.066 | 0.061 | 0.040 | ||
| 9 | 0.027 | 0.051 | 0.042 | 0.070 | 0.072 | 0.066 | 0.044 | 0.026 | |
| 10 | 0.036 | 0.027 | 0.019 | 0.069 | 0.069 | 0.075 | 0.046 | 0.036 | 0.042 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ualiyeva, D.; Ermakov, O.A.; Litvinchuk, S.N.; Guo, X.; Ivanov, A.Y.; Xu, R.; Li, J.; Xu, F.; Arifulova, I.I.; Kaptyonkina, A.G.; et al. Diversity, Phylogenetic Relationships and Distribution of Marsh Frogs (the Pelophylax ridibundus complex) from Kazakhstan and Northwest China. Diversity 2022, 14, 869. https://doi.org/10.3390/d14100869
Ualiyeva D, Ermakov OA, Litvinchuk SN, Guo X, Ivanov AY, Xu R, Li J, Xu F, Arifulova II, Kaptyonkina AG, et al. Diversity, Phylogenetic Relationships and Distribution of Marsh Frogs (the Pelophylax ridibundus complex) from Kazakhstan and Northwest China. Diversity. 2022; 14(10):869. https://doi.org/10.3390/d14100869
Chicago/Turabian StyleUaliyeva, Daniya, Oleg A. Ermakov, Spartak N. Litvinchuk, Xianguang Guo, Alexander Yu. Ivanov, Rui Xu, Jun Li, Feng Xu, Irina I. Arifulova, Alyona G. Kaptyonkina, and et al. 2022. "Diversity, Phylogenetic Relationships and Distribution of Marsh Frogs (the Pelophylax ridibundus complex) from Kazakhstan and Northwest China" Diversity 14, no. 10: 869. https://doi.org/10.3390/d14100869
APA StyleUaliyeva, D., Ermakov, O. A., Litvinchuk, S. N., Guo, X., Ivanov, A. Y., Xu, R., Li, J., Xu, F., Arifulova, I. I., Kaptyonkina, A. G., Khromov, V. A., Krainyuk, V. N., Sarzhanov, F., & Dujsebayeva, T. N. (2022). Diversity, Phylogenetic Relationships and Distribution of Marsh Frogs (the Pelophylax ridibundus complex) from Kazakhstan and Northwest China. Diversity, 14(10), 869. https://doi.org/10.3390/d14100869