




Deep Ecology, Biodiversity and Assisted Natural Regeneration of European Hemiboreal Forests




Abstract
:1. Introduction
2. European Hemiboreal Tree Species: The Case of Lithuania
3. Hemiboreal Tree Dynamics
3.1. Tree Regeneration Strategy
3.2. Natural Regeneration of European Hemiboreal Tree Species
4. Concept of Assisted Natural Regeneration of Trees
5. Grounding Assisted Natural Regeneration on Ethical Framework
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Olff, H.; Alonso, D.; Berg, M.P.; Eriksson, B.K.; Loreau, M.; Piersma, T.; Rooney, N. Parallel Ecological Networks in Ecosystems. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1755–1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camazine, S.; Deneubourg, J.-L.; Franks, N.R.; Sneyd, J.; Theraula, G.; Bonabeau, E. Self-Organization in Biological Systems; Princeton University Press: Princeton, NJ, USA, 2020; ISBN 978-0-691-21292-0. [Google Scholar]
- Kazansky, A.B. Bootstrapping of Life through Holonomy and Self-Modification. In Proceedings of the 9th International Conference on Computing Anticipatory Systems, Liege, Belgium, 3–8 August 2009; Dubois, D.M., Ed.; American Institute of Physics: Melville, NY, USA, 2010; pp. 297–306. [Google Scholar]
- Emery, S.M. Succession: A Closer Look. Nat. Educ. 2010, 3, 10–45. [Google Scholar]
- Schwarz, C.; Gourgue, O.; van Belzen, J.; Zhu, Z.; Bouma, T.J.; van de Koppel, J.; Ruessink, G.; Claude, N.; Temmerman, S. Self-Organization of a Biogeomorphic Landscape Controlled by Plant Life-History Traits. Nat. Geosci. 2018, 11, 672–677. [Google Scholar] [CrossRef]
- Turner, M.G. Landscape Ecology: The Effect of Pattern on Process. Annu. Rev. Ecol. Syst. 1989, 20, 171–197. [Google Scholar] [CrossRef]
- Minore, D.; Laacke, R.J. Natural Regeneration. In Reforestation Practices in Southwest Oregon and Northern California: Chapter 11; Hobbs, S.D., Tesch, S.D., Owston, P.W., Stewart, R.E., Tappeiner, J.C., Wells, G.E., Eds.; Oregon State University Press: Corvallis, OR, USA, 1992; pp. 258–283. ISBN 0-87437-001-9. [Google Scholar]
- Bajc, M.; Aravanopoulos, F.; Westergren, M.; Fussi, B.; Kavaliauskas, D.; Alizoti, P.; Kiourtsis, F.; Kraigher, H. Manual for Forest Genetic Monitoring; Slovenian Forestry Institute: Ljubljana, Slovenia, 2020. [Google Scholar]
- Holbrook, M. Adventures in Complexity: An Essay on Dynamic Open Complex Adaptive Systems, Butterfly Effects, Self-Organizing Order, Coevolution, the Ecological Perspective, Fitness Landscapes, Market Spaces, Emergent Beauty at the Edge of Chaos, and All That Jazz. Acad. Mark. Sci. Rev. 2003, 2003, 1–181. [Google Scholar]
- Borman, M.M.; Pyke, D.A. Successional Theory and the Desired Plant Community Approach. Rangelands 1994, 16, 82–84. [Google Scholar]
- Chazdon, R.L. Second Growth: The Promise of Tropical Forest Regeneration in an Age of Deforestation; University of Chicago Press: Chicago, IL, USA, 2014; ISBN 978-0-226-11791-1. [Google Scholar]
- Chazdon, R.L.; Brancalion, P.H.S.; Laestadius, L.; Bennett-Curry, A.; Buckingham, K.; Kumar, C.; Moll-Rocek, J.; Vieira, I.C.G.; Wilson, S.J. When Is a Forest a Forest? Forest Concepts and Definitions in the Era of Forest and Landscape Restoration. Ambio 2016, 45, 538–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, N.; Fomin, V.; Kusbach, A. Experience of Forest Ecological Classification in Assessment of Vegetation Dynamics. Sustainability 2022, 14, 3384. [Google Scholar] [CrossRef]
- Jõgiste, K.; Frelich, L.E.; Laarmann, D.; Vodde, F.; Baders, E.; Donis, J.; Jansons, A.; Kangur, A.; Korjus, H.; Köster, K.; et al. Imprints of Management History on Hemiboreal Forest Ecosystems in the Baltic States. Ecosphere 2018, 9, e02503. [Google Scholar] [CrossRef] [Green Version]
- Angelstam, P.; Manton, M.; Pedersen, S.; Elbakidze, M. Disrupted Trophic Interactions Affect Recruitment of Boreal Deciduous and Coniferous Trees in Northern Europe. Ecol. Appl. 2017, 27, 1108–1123. [Google Scholar] [CrossRef] [Green Version]
- European Commission. New EU Forest Strategy for 2030. In Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions; European Union: Brussels, Belgium, 2021; p. 28. [Google Scholar]
- Manton, M.; Ruffner, C.; Kibirkštis, G.; Brazaitis, G.; Marozas, V.; Pukienė, R.; Makrickiene, E.; Angelstam, P. Fire Occurrence in Hemi-Boreal Forests: Exploring Natural and Cultural Scots Pine Fire Regimes Using Dendrochronology in Lithuania. Land 2022, 11, 260. [Google Scholar] [CrossRef]
- Karazija, S. Forest Types of Lithuania; Mokslas: Vilnius, Lietuva, 1988; ISBN 978-5-420-00421-0. [Google Scholar]
- Brazaitis, G.; Marozas, V.; Augutis, D.; Preikša, Ž.; Šaudytė-Manton, S. Lithuanian Forest Habitat Management Recommendations–“Guidelines for the Management of Natural Forest Habitat Types of EC Importance”; Naturalit: Vilnius, Lietuva, 2021. [Google Scholar]
- Vaičys, M.; Mažvila, J. The Influence of Soil Characteristics on Plant Productivity and Ecological Stability. Ekologija 2009, 55, 99–106. [Google Scholar] [CrossRef]
- Buivydaitė, V. Classification of Soils of Lithuania Based on FAO-UNESCO Soil Classification System and WRB. In Proceedings of the 17th World Congress of Soil Science, Bangkok, Thailand, 14–20 August 2002; The Soil and Fertilizer Society of Thailand: Bangkok, Thailand, 2002; pp. 814–826. [Google Scholar]
- Kuliešis, A.; Kulbokas, G.; Kasperavičius, A.; Kazanavičiūtė, V.; Kvalkauskienė, M. Lietuvos nacionalinė miškų inventorizacija, 1998–2017. Nuo matavimų iki sprendimų. Lithuanian national forest inventory, 1998–2017. In From Measurements to Decision Making; Lututė: Kaunas, Lietuva, 2021; ISBN 978-9955-37-234-9. [Google Scholar]
- Kulbokas, G. Experiences from Lithuania-UNECE Technical Workshop on European Forest Types. In Presented at the UNECE Technical Workshop on European Forest Types: Reporting Using the New European Forest Types, Bordeaux, France, 19–21 May 2010; Lithuanian State Forest Service: Kaunas, Lithuania, 2010; pp. 1–16. [Google Scholar]
- Angelstam, P. Landscape Analysis as a Tool for the Scientific Management of Biodiversity. Ecol. Bull. 1997, 46, 140–170. [Google Scholar]
- Fomin, V.; Mikhailovich, A.; Zalesov, S.; Popov, A.; Terekhov, G. Development of Ideas within the Framework of the Genetic Approach to the Classification of Forest Types. Balt. For. 2020, 27, 26–39. [Google Scholar] [CrossRef]
- Christensen, N.L.; Peet, R.K. Secondary Forest Succession on the North Carolina Piedmont. In Forest Succession: Concepts and Application; West, D.C., Shugart, H.H., Botkin, D.B., Eds.; Springer Advanced Texts in Life Sciences; Springer: New York, NY, USA, 1981; pp. 230–245. ISBN 978-1-4612-5950-3. [Google Scholar]
- Angelstam, P.; Kuuluvainen, T. Boreal Forest Disturbance Regimes, Successional Dynamics and Landscape Structures: A European Perspective. Ecol. Bull. 2004, 51, 117–136. [Google Scholar] [CrossRef]
- Angelstam, P.; Dönz-Breuss, M. Measuring Forest Biodiversity at the Stand Scale: An Evaluation of Indicators in European Forest History Gradients. Ecol. Bull. 2004, 51, 305–332. Available online: https://www.jstor.org/stable/20113319 (accessed on 19 September 2022).
- Jandl, R.; Spathelf, P.; Bolte, A.; Prescott, C.E. Forest Adaptation to Climate Change—Is Non-Management an Option? Ann. For. Sci. 2019, 76, 48. [Google Scholar] [CrossRef] [Green Version]
- Russell, G.J. Turnover Dynamics Across Ecological and Geological Scales. In Turnover Dynamics Across Ecological and Geological Scales; Columbia University Press: New York, NY, USA, 2001; pp. 377–404. ISBN 978-0-231-50580-2. [Google Scholar]
- Neophytou, C.; Heer, K.; Milesi, P.; Peter, M.; Pyhäjärvi, T.; Westergren, M.; Rellstab, C.; Gugerli, F. Genomics and Adaptation in Forest Ecosystems. Tree Genet. Genomes 2022, 18, 12. [Google Scholar] [CrossRef]
- Petrokas, R.; Baliuckas, V.; Manton, M. Successional Categorization of European Hemi-Boreal Forest Tree Species. Plants 2020, 9, 1381. [Google Scholar] [CrossRef]
- Yamamoto, S. Gap Regeneration of Major Tree Species in Different Forest Types of Japan. Vegetatio 1996, 127, 203–213. [Google Scholar] [CrossRef]
- Ulft, L. Regeneration in Natural and Logged Tropical Rain Forest-Modelling Seed Dispersal and Regeneration of Tropical Trees in Guyana; Tropenbos-Guyana Series 12; Tropenbos International: Georgetown, Guyana, 2004; ISBN 90-5113-076-7. [Google Scholar]
- Franklin, J. Regeneration and Growth of Pioneer and Shade-tolerant Rain Forest Trees in Tonga. New Zealand J. Bot. 2003, 41, 669–684. [Google Scholar] [CrossRef]
- Ning, Z.; Hong, J.; Yong-Yan, J. A Phenology Study on the Common Tree Species of Natural Secondary Forests in Northeast China. Chin. J. Plant Ecol. 1990, 14, 336. [Google Scholar]
- Clark, D.A.; Clark, D.B. Life History Diversity of Canopy and Emergent Trees in a Neotropical Rain Forest. Ecol. Monogr. 1992, 62, 315–344. [Google Scholar] [CrossRef]
- Whitmore, T.C. Canopy Gaps and the Two Major Groups of Forest Trees. Ecology 1989, 70, 536–538. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Hill, M.O.; Roy, D.B.; Thompson, K. Hemeroby, Urbanity and Ruderality: Bioindicators of Disturbance and Human Impact. J. Appl. Ecol. 2002, 39, 708–720. [Google Scholar] [CrossRef]
- Shorohova, E.; Kuuluvainen, T.; Kangur, A.; Jõgiste, K. Natural Stand Structures, Disturbance Regimes and Successional Dynamics in the Eurasian Boreal Forests: A Review with Special Reference to Russian Studies. Ann. For. Sci. 2009, 66, 201. [Google Scholar] [CrossRef] [Green Version]
- Angelstam, P.K. Maintaining and Restoring Biodiversity in European Boreal Forests by Developing Natural Disturbance Regimes. J. Veg. Sci. 1998, 9, 593–602. [Google Scholar] [CrossRef]
- Rull, V. Quaternary Palaeoecology and Ecological Theory. Orsis 1990, 5, 91–111. [Google Scholar]
- Hunter, M.L.; Schmiegelow, F.K.A. Wildlife, Forests, and Forestry: Principles of Managing Forests for Biological Diversity, 2nd ed.; Prentice Hall: Boston, MA, USA, 2011; ISBN 978-0-13-501432-5. [Google Scholar]
- Birks, H. Late-Quaternary Biotic Changes in Terrestrial and Lacustrine Environments, with Particular Reference to North-West Europe. In Handbook of Holocene Palaeoecology and Palaeohydrology; Berglund, B.E., Ed.; Wiley-Interscience; John Wiley & Sons Ltd.: Chichester, UK, 1986. [Google Scholar]
- Birks, H. Contributions of Quaternary Botany to Modern Ecology and Biogeography. Plant Ecol. Divers. 2019, 12, 189–385. [Google Scholar] [CrossRef]
- Godvod, K.; Brazaitis, G.; Bačkaitis, J.; Kulbokas, G. The Development and Growth of Larch Stands in Lithuania. J. For. Sci. 2018, 64, 199–206. [Google Scholar] [CrossRef]
- Petrokas, R.; Kavaliauskas, D. Concept for Genetic Monitoring of Hemiboreal Tree Dynamics in Lithuania. Land 2022, 11, 1249. [Google Scholar] [CrossRef]
- Odum, E.P.; Barrett, G.W. Fundamentals of Ecology, 5th ed.; Thomson Brooks/Cole: Belmont, CA, USA, 2005; ISBN 978-0-534-42066-6. [Google Scholar]
- Navasaitis, M.; Ozolinčius, R.; Smaliukas, D.; Balevičienė, J.M. Lietuvos Dendroflora: Monografija; Lutute: Kaunas, Lietuva, 2003; ISBN 978-9955-575-35-1. [Google Scholar]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest Disturbances under Climate Change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochard, H. The Fate of Forest Ecosystems in the Anthropocene. Sigmoidal J. 2020; e008, Preprint. [Google Scholar] [CrossRef]
- Shono, K.; Chazdon, R.; Bodin, B.; Wilson, S.; Durst, P. Assisted Natural Regeneration: Harnessing Nature for Restoration. Unasylva 2020, 252, 71–81. [Google Scholar]
- Chai, Y.; Yue, M.; Wang, M.; Xu, J.; Liu, X.; Zhang, R.; Wan, P. Plant Functional Traits Suggest a Change in Novel Ecological Strategies for Dominant Species in the Stages of Forest Succession. Oecologia 2016, 180, 771–783. [Google Scholar] [CrossRef]
- Yu, R.; Huang, J.; Xu, Y.; Ding, Y.; Zang, R. Plant Functional Niches in Forests Across Four Climatic Zones: Exploring the Periodic Table of Niches Based on Plant Functional Traits. Front. Plant Sci. 2020, 11, 841. [Google Scholar] [CrossRef]
- Ehlers, A.; Worm, B.; Reusch, T.B.H. Importance of Genetic Diversity in Eelgrass Zostera Marina for Its Resilience to Global Warming. Mar. Ecol. Prog. Ser. 2008, 355, 1–7. [Google Scholar] [CrossRef]
- Wright, D.; Bishop, J.M.; Matthee, C.A.; von der Heyden, S. Genetic Isolation by Distance Reveals Restricted Dispersal across a Range of Life Histories: Implications for Biodiversity Conservation Planning across Highly Variable Marine Environments. Divers. Distrib. 2015, 21, 698–710. [Google Scholar] [CrossRef]
- Orr, H.A. Fitness and Its Role in Evolutionary Genetics. Nat. Rev. Genet. 2009, 10, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Chazdon, R.; Calixto, B.; Oliveira, M.; Messinger, J.; Alves, J.; Calmon, M.; Anderson, W. The Benefits and Power of Assisted Natural Regeneration. Available online: https://www.wri.org/insights/what-assisted-natural-regeneration-benefits-definition (accessed on 19 September 2022).
- Yang, Y.; Wang, L.; Yang, Z.; Xu, C.; Xie, J.; Chen, G.; Lin, C.; Guo, J.; Liu, X.; Xiong, D.; et al. Large Ecosystem Service Benefits of Assisted Natural Regeneration. J. Geophys. Res. Biogeosciences 2018, 123, 676–687. [Google Scholar] [CrossRef]
- Kpolita, A.; Dubiez, E.; Yongo, O.; Peltier, R. First Evaluation of the Use of Assisted Natural Regeneration by Central African Farmers to Restore Their Landscapes. Trees For. People 2022, 7, 100165. [Google Scholar] [CrossRef]
- A European Commission Initiative Providing Environmental Research Findings in an Easy-to-Understand Way. Available online: https://environment.ec.europa.eu/research-and-innovation/science-environment-policy_en (accessed on 19 September 2022).
- Ibanga, D.-A. Is Deep Ecology Inapplicable in African Context: A Conversation with Fainos Mangena. Filos. Theor. J. Afr. Philos. Cult. Relig. 2017, 6, 101–119. [Google Scholar] [CrossRef]
- Akamani, K. Integrating Deep Ecology and Adaptive Governance for Sustainable Development: Implications for Protected Areas Management. Sustainability 2020, 12, 5757. [Google Scholar] [CrossRef]
- Sumarmi, S.; Bachri, S.; Mutia, T.; Yustesia, A.; Fathoni, M.N.; Muthi, M.A.; Nuraini, S.G. The Deep Ecology Perspective of Awig-Awig: Local Tribal Forest Preservation Laws in Tenganan Cultural Village, Indonesia. J. Sustain. Sci. Manag. 2020, 15, 102–113. [Google Scholar] [CrossRef]
- Roser, D.; Seidel, C. Climate Justice: An Introduction; Routledge: London, UK, 2016; ISBN 978-1-315-61796-1. [Google Scholar]
- Ezemonye, L.I.; Ogbe, M.G. Environmental Issues and Implications of Biofuels as Alternative Sources of Energy. In Critical Issues on Nigeria’s Development: Environment, Economy and Social Justice; Okafor, F.C., Ed.; Spectrum Books Ltd.: Ibadan, Nigeria, 2011; pp. 347–376. [Google Scholar]


{kind=link}
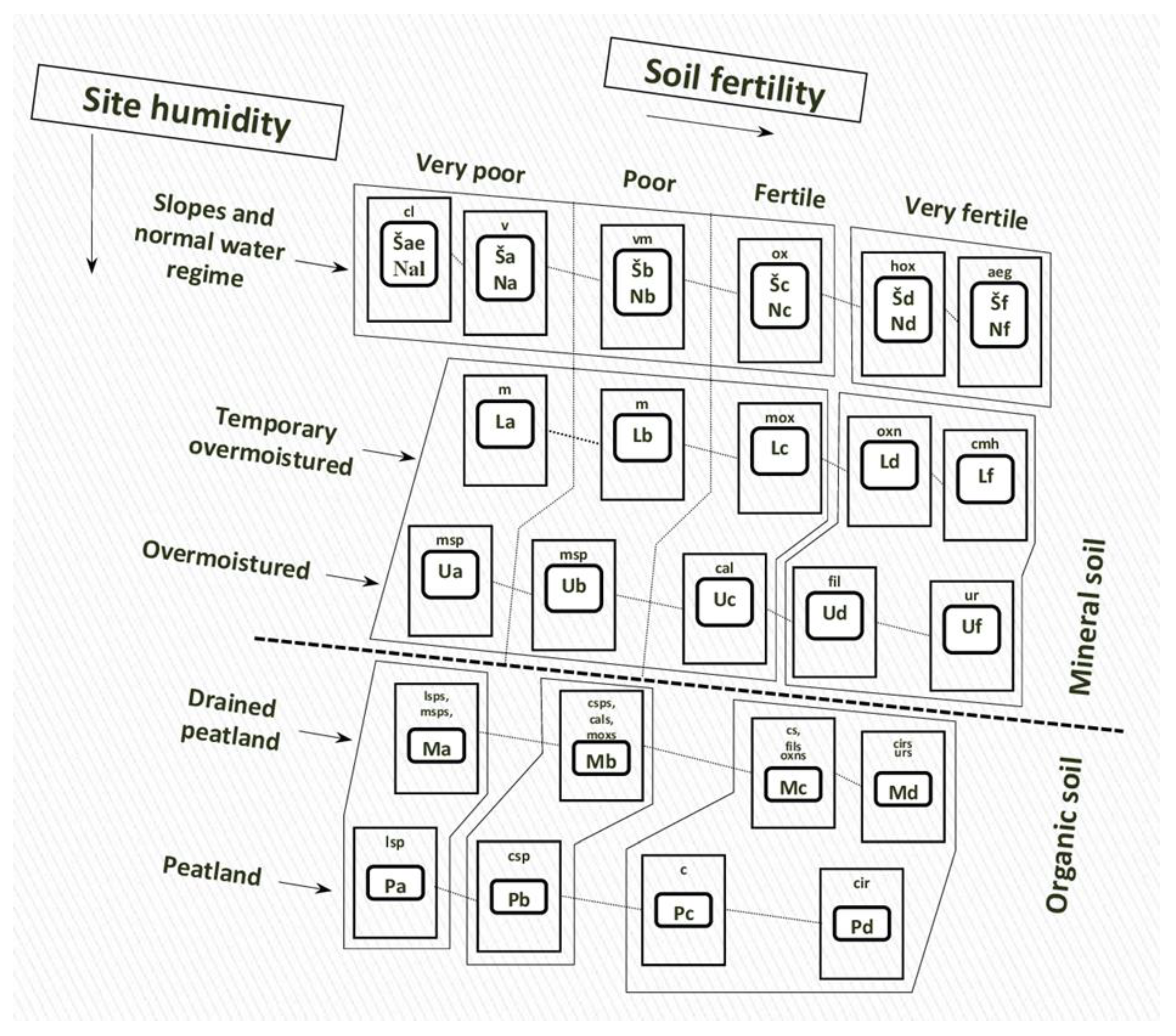
| Regeneration Strategy: C—Colonization O—Occupation I—Invasion E—Expansion | Forest Habitat Types (NATURA 2000 Codes)/Codes of the Lithuanian Forest Type Series * and Forest Site Types ** | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mixed Broadleaved Forests | Norway Spruce Mixed Forests | Scots Pine Forests | |||||||||||||||||
| (9020 9080 91F0 91E0) | (9050 9160 9180 9190 9070) | (9010 9060 91D0 91T0) | |||||||||||||||||
| aeg * Nf ** | cmh Lf | cal Uc | Fil Ud | ur Uf | cir Pd | c Pc | ox Nc | mox Lc | hox Nd | oxn Ld | cl Nal | v Na | vm Nb | m Lb | msp Ub | csp Pb | lsp Pa | ||
| C | Alnus glutinosa | G | G | G | G | G, S | G, S | G | |||||||||||
| C | Alnus incana | G | G | g | G | s, g | s, g | G | G | ||||||||||
| C | Betula pendula | G | G | g | G | g | S | S | S | S | s | s | S | ||||||
| C | Betula pubescens | G | G | S | S | g | G | S | s | S | |||||||||
| C | Larix decidua | S | s | ||||||||||||||||
| C | Pinus sylvestris | s, m | M | M | M | M | M | M | M | M | M | ||||||||
| O | Fraxinus excelsior | G | G | G | g | G | |||||||||||||
| O | Populus tremula | G | G | G | S | S | s | S | S | ||||||||||
| O | Quercus robur | G | G | G | G | g | |||||||||||||
| O | Ulmus laevis | G | G | g | |||||||||||||||
| I | Acer platanoides | G | g | ||||||||||||||||
| I | Carpinus betulus | G | G | ||||||||||||||||
| I | Picea abies | g | G | S | S | S | S | s | s | s | |||||||||
| I | Ulmus glabra | G | |||||||||||||||||
| E | Fagus sylvatica | G | G | ||||||||||||||||
| E | Tilia cordata | G | g | G | g | ||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrokas, R.; Ibanga, D.-A.; Manton, M. Deep Ecology, Biodiversity and Assisted Natural Regeneration of European Hemiboreal Forests. Diversity 2022, 14, 892. https://doi.org/10.3390/d14100892
Petrokas R, Ibanga D-A, Manton M. Deep Ecology, Biodiversity and Assisted Natural Regeneration of European Hemiboreal Forests. Diversity. 2022; 14(10):892. https://doi.org/10.3390/d14100892
Chicago/Turabian StylePetrokas, Raimundas, Diana-Abasi Ibanga, and Michael Manton. 2022. "Deep Ecology, Biodiversity and Assisted Natural Regeneration of European Hemiboreal Forests" Diversity 14, no. 10: 892. https://doi.org/10.3390/d14100892
APA StylePetrokas, R., Ibanga, D. -A., & Manton, M. (2022). Deep Ecology, Biodiversity and Assisted Natural Regeneration of European Hemiboreal Forests. Diversity, 14(10), 892. https://doi.org/10.3390/d14100892