
Mapping the Distribution and Dispersal Risks of the Alien Invasive Plant Ageratina adenophora in China

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
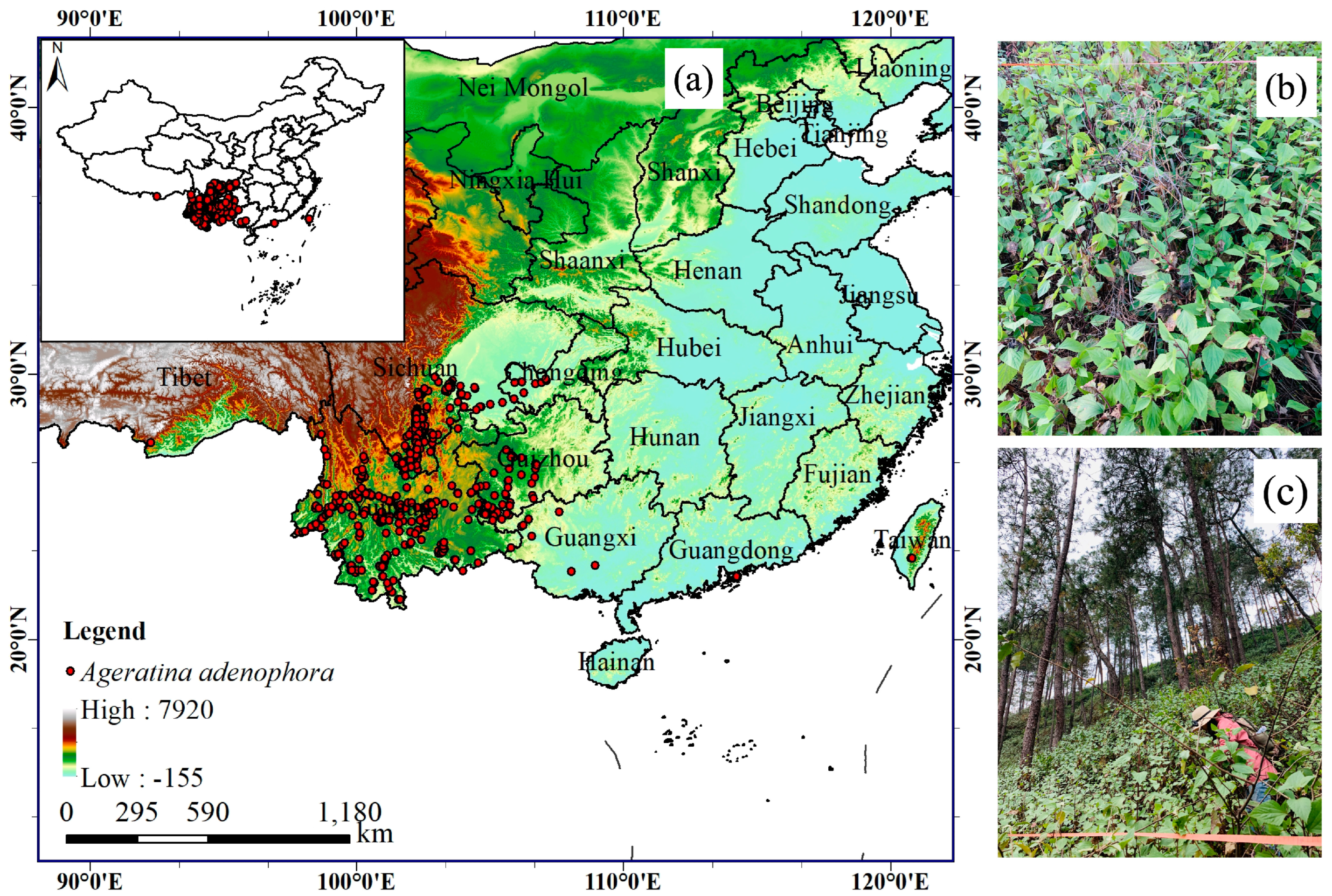
2.1. Species Occurrence Records
2.2. Environmental Data and Pre-Processing
2.3. Model Construction and Data Processing
2.4. Niche Similarity Examination
3. Results
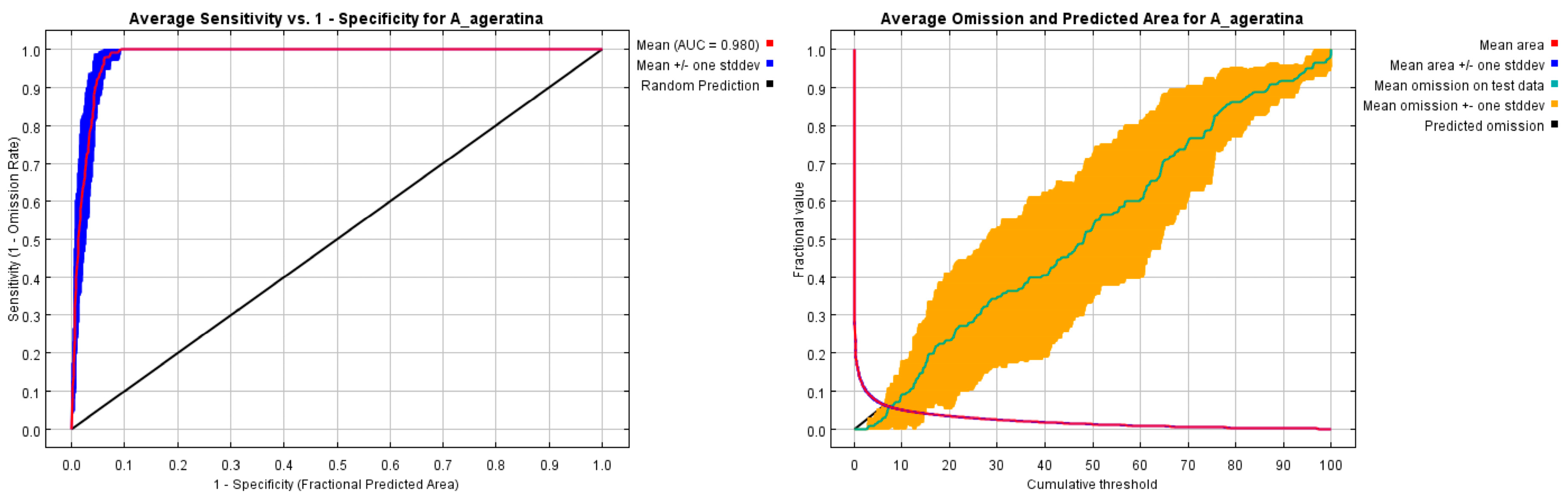
3.1. Model Accuracy Assessment
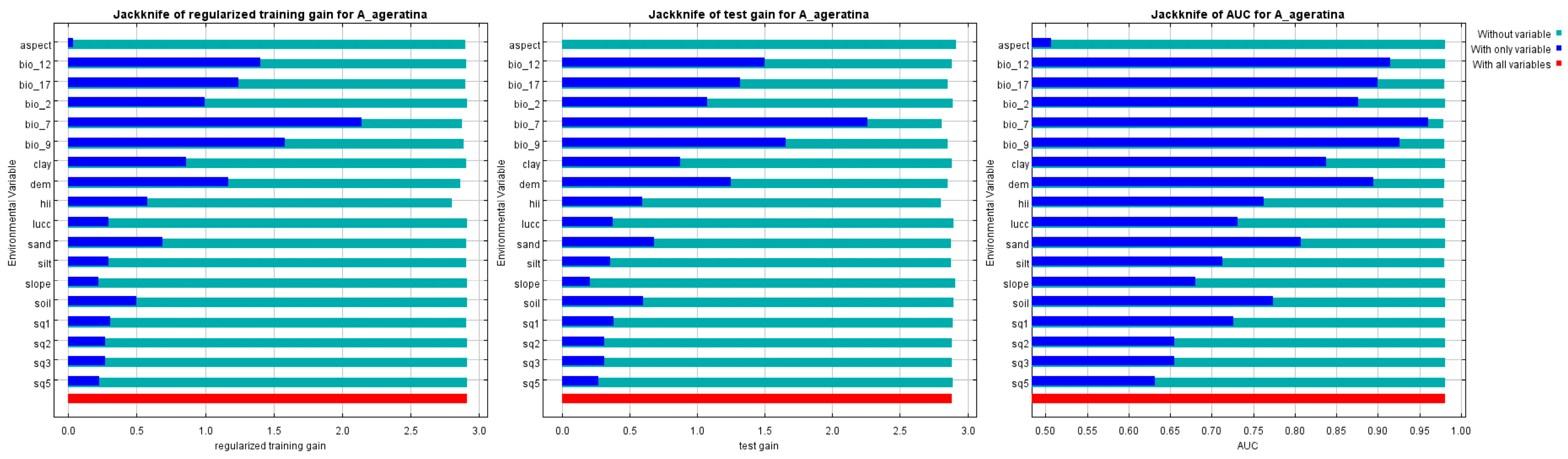
3.2. Key Drivers Affecting the Distribution of A. adenophora
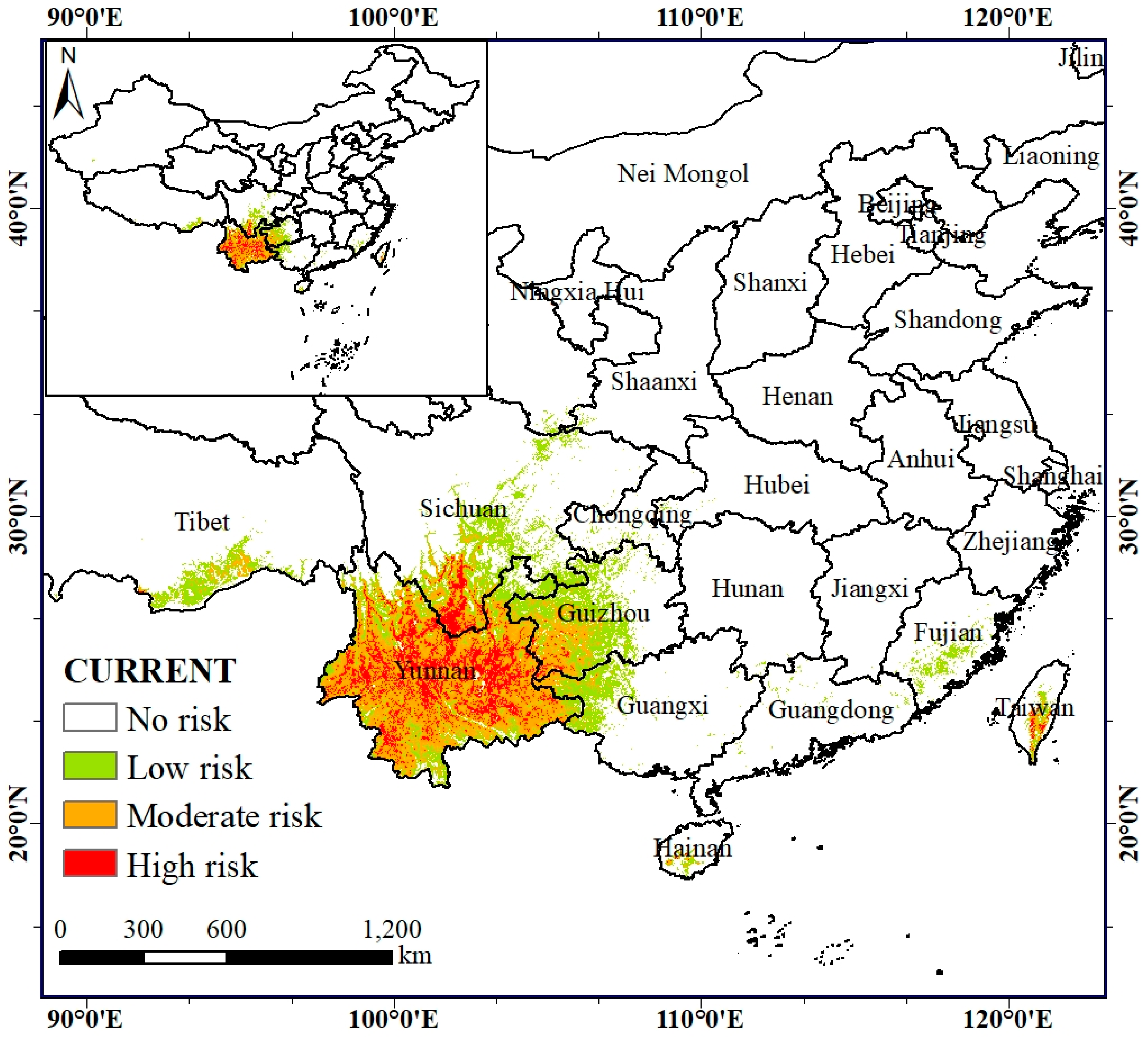
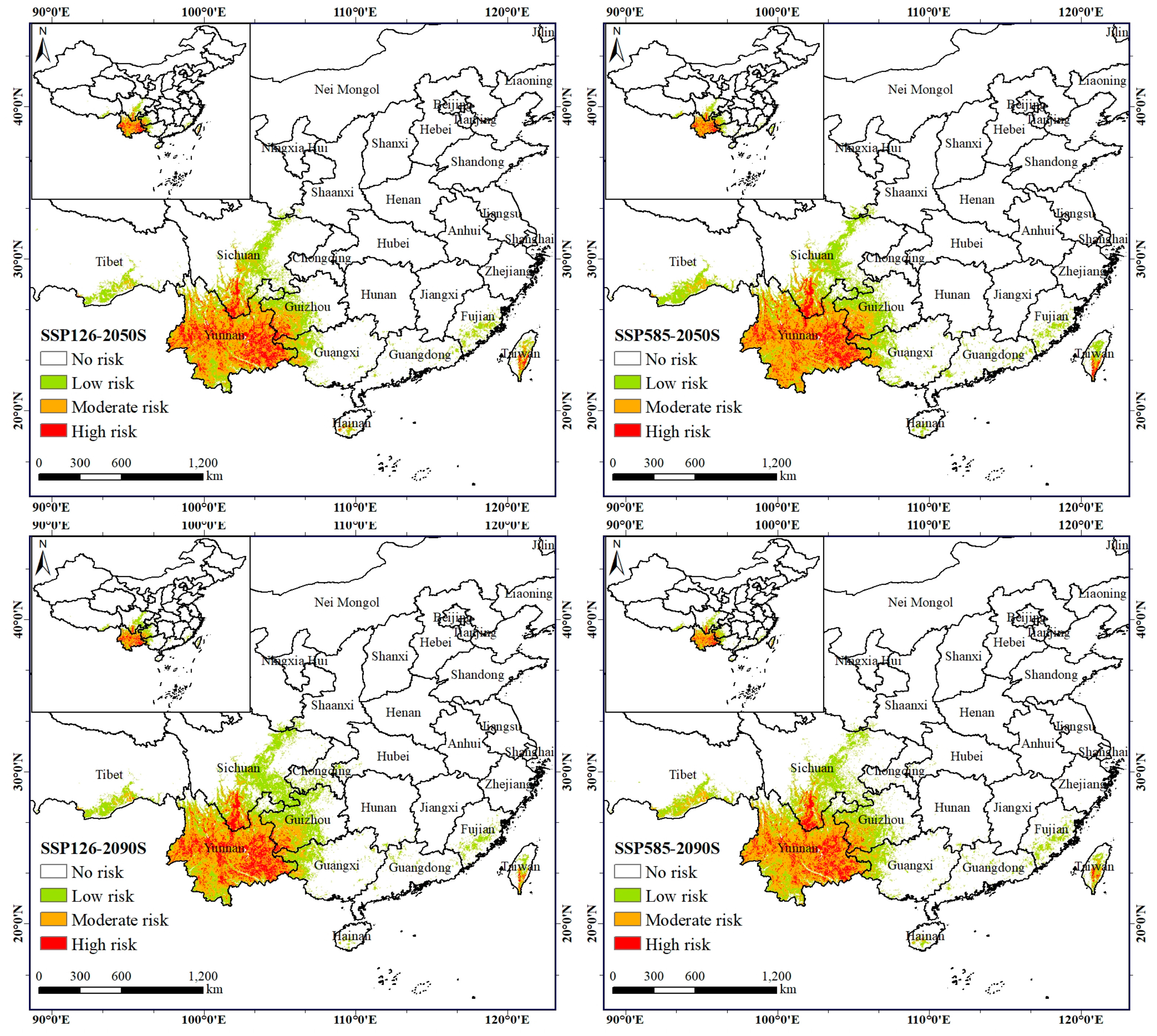
3.3. Characteristics of the Potential Risk Area of the A. Adenophora under Current Climate Conditions
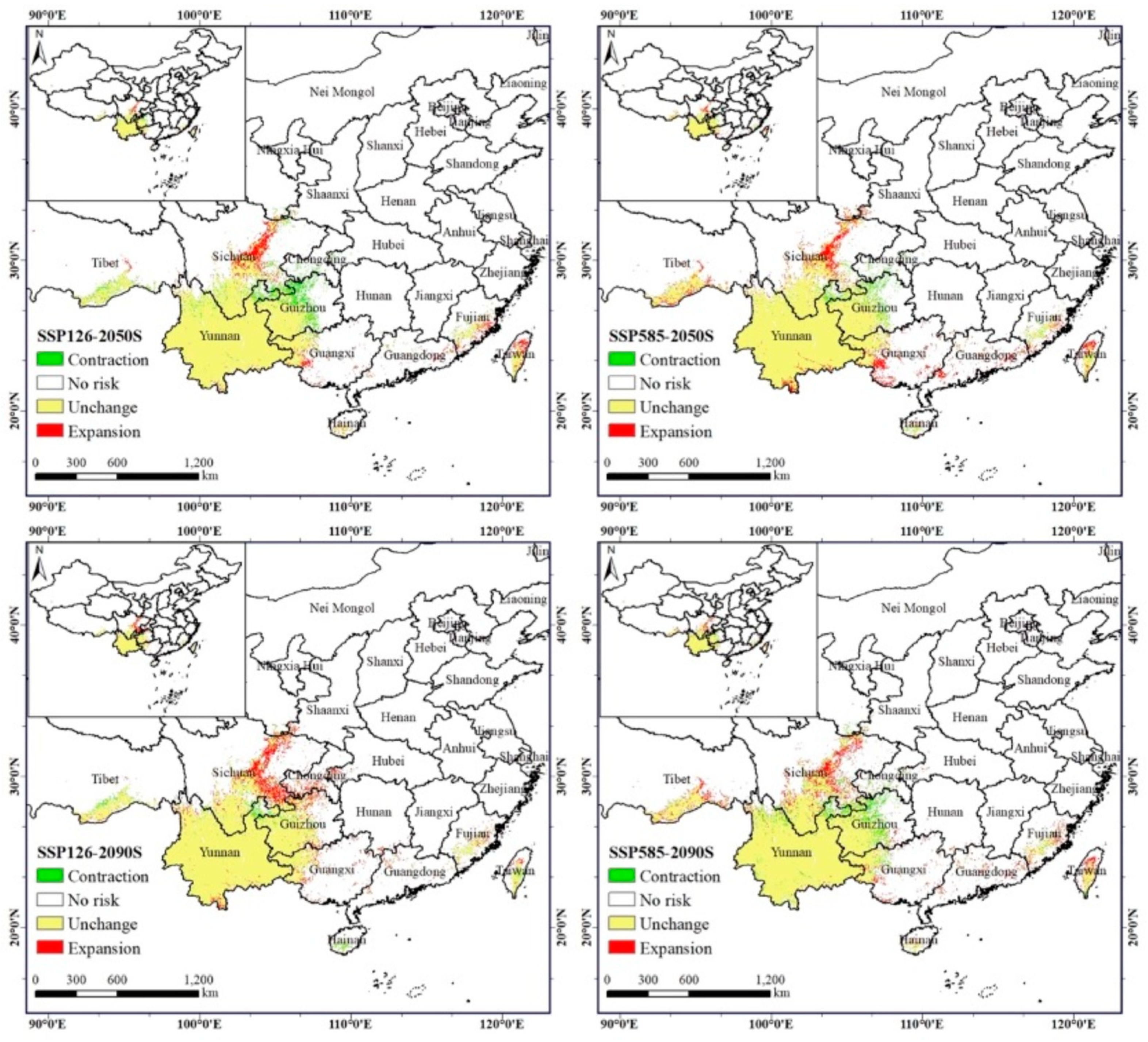
3.4. Evaluation of the Dispersal Dynamics of A. adenophora
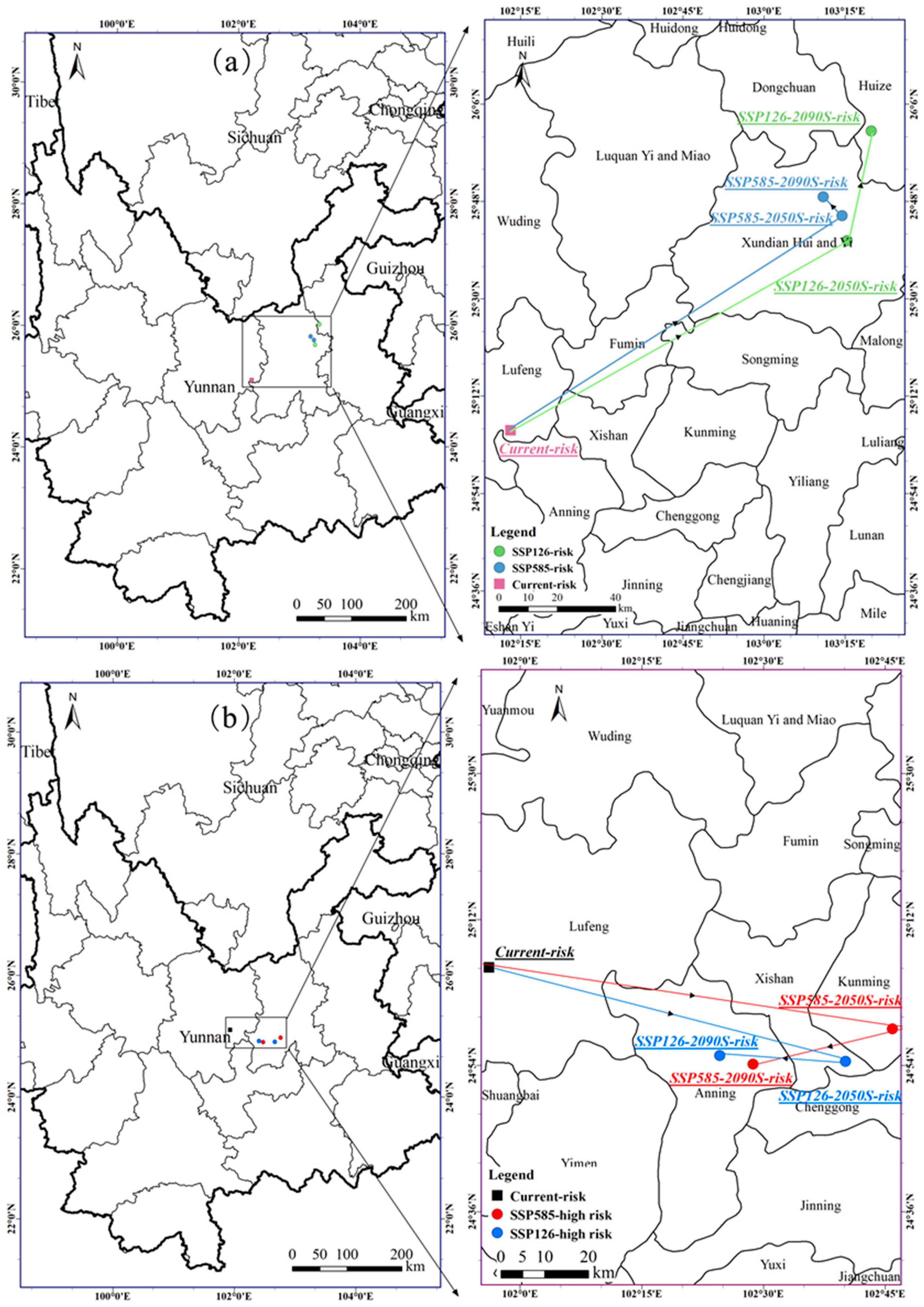
3.5. The Centroid Distribution Transfer of the A. adenophora
4. Discussion
4.1. Potential Risk of Invasive Alien Plants under the Current and Future Scenario
4.2. Key Drivers Affecting Invasive Alien Plants
4.3. Uncertainty
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Printed in Great Britain By Butler and Tanner Ltd.: London, UK, 1958. [Google Scholar]
- The Belt and Road Countries Establish International Union for Plant Protection to Jointly Prevent and Control Alien Species. Available online: https://www.sohu.com/a/205957772_100020996 (accessed on 22 November 2017).
- The 2020 China Ecological Environment Status Bulletin was released. Available online: http://www.xinhuanet.com/enterprise/2021-05/26/c_1127493313.htm (accessed on 26 May 2021).
- Falk-Petersen, J.; Bøhn, T.; Sandlund, O.T. On the numerous concepts in invasion biology. Biol. Invasions 2006, 8, 1409–1424. [Google Scholar] [CrossRef] [Green Version]
- Christian, C.E. Consequences of a biological invasion reveal the importance of mutualism for plant communities. Nature 2001, 413, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.L.; Westbrooks, R. Biological invasions as global environmental change. Am. Sci. 1996, 8, 468–478. [Google Scholar]
- Shackleton, R.T.; Foxcroft, L.C.; Pyšek, P.; Wood, L.E.; Richardson, D.M. Assessing biological invasions in protected areas after 30 years: Revisiting nature reserves targeted by the 1980s SCOPE programme. Biol. Conserv. 2020, 243, 108424. [Google Scholar] [CrossRef]
- Horvitz, N.; Wang, R.; Zhu, M.; Wan, F.H.; Nathan, R. A simple modeling approach to elucidate the main transport processes and predict invasive spread: River-mediated invasion of A geratina adenophora in China. Water Resour. Res. 2014, 50, 9738–9747. [Google Scholar] [CrossRef]
- Cook, D.C.; Thomas, M.B.; Cunningham, S.A.; Anderson, D.L.; De Barro, P.J. Predicting the economic impact of an invasive species on an ecosystem service. Ecol. Appl. 2007, 17, 1832–1840. [Google Scholar] [CrossRef]
- Mitchell, C.E.; Agrawal, A.A.; Bever, J.D.; Gilbert, G.S.; Hufbauer, R.A.; Klironomos, J.N.; Maron, J.L.; Morris, W.F.; Parker, I.M.; Power, A.G. Biotic interactions and plant invasions. Ecol. Lett. 2006, 9, 726–740. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.M.; Vila, M.; Antonio, C.M.D.; Dukes, J.S.; Grigulis, K.; Lavorel, S. Mechanisms underlying the impacts of exotic plant invasions. Proc. R. Soc. London. Ser. B Biol. Sci. 2003, 270, 775–781. [Google Scholar] [CrossRef]
- Traveset, A.; Richardson, D.M. Biological invasions as disruptors of plant reproductive mutualisms. Trends Ecol. Evol. 2006, 21, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.; De Siqueira, M.F.; Grainger, A.; Hannah, L. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-y.; Asner, G.P. Applications of remote sensing to alien invasive plant studies. Sensors 2009, 9, 4869–4889. [Google Scholar] [CrossRef]
- Dostál, P.; Müllerová, J.; Pyšek, P.; Pergl, J.; Klinerová, T. The impact of an invasive plant changes over time. Ecol. Lett. 2013, 16, 1277–1284. [Google Scholar] [CrossRef]
- Hess, M.C.; Mesléard, F.; Buisson, E. Priority effects: Emerging principles for invasive plant species management. Ecol. Eng. 2019, 127, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Weidlich, E.W.; Flórido, F.G.; Sorrini, T.B.; Brancalion, P.H. Controlling invasive plant species in ecological restoration: A global review. J. Appl. Ecol. 2020, 57, 1806–1817. [Google Scholar] [CrossRef]
- Tang, S.; Pan, Y.; Wei, C.; Li, X.; Lü, S. Testing of an integrated regime for effective and sustainable control of invasive Crofton weed (Ageratina adenophora) comprising the use of natural inhibitor species, activated charcoal, and fungicide. Weed Biol. Manage. 2019, 19, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Wang, J.F.; Qiu, Z.J.; Meng, B.; Wan, F.H.; Wang, Y.Z. Multiple mechanisms underlie rapid expansion of an invasive alien plant. New Phytol. 2011, 191, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Huang, W.; Jiang, Z.; Wang, W.; Liu, W.; Guo, J.; Qiang, S.; Li, B.; Wang, J.; Yang, G. Invasive mechanism and control strategy of Ageratina adenophora (Sprengel). Sci. China Life Sci. 2010, 53, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, Y.Z. Invasion dynamics and potential spread of the invasive alien plant species Ageratina adenophora (Asteraceae) in China. Divers. Distrib. 2006, 12, 397–408. [Google Scholar] [CrossRef]
- Kumar, M.; Garkoti, S.C. Allelopathy effects of invasive alien Ageratina adenophora on native shrub species of chir pine forest in the central Himalaya, India. J. For. Res. 2022, 27, 53–62. [Google Scholar] [CrossRef]
- Heger, T.; Trepl, L. Predicting biological invasions. Biol. Invasions 2003, 5, 313–321. [Google Scholar] [CrossRef]
- Holmes, P.M.; Esler, K.J.; Gaertner, M.; Geerts, S.; Hall, S.A.; Nsikani, M.M.; Richardson, D.M.; Ruwanza, S. Biological invasions and ecological restoration in South Africa. In Biological Invasions in South Africa; Springer International Publishing: Cham, Switzerland, 2020; pp. 665–700. [Google Scholar]
- Petrovskii, S.V.; Malchow, H.; Hilker, F.M.; Venturino, E. Patterns of patchy spread in deterministic and stochastic models of biological invasion and biological control. Biol. Invasions 2005, 7, 771–793. [Google Scholar] [CrossRef]
- Jarnevich, C.S.; Holcombe, T.R.; Barnett, D.T.; Stohlgren, T.J.; Kartesz, J.T. Forecasting weed distributions using climate data: A GIS early warning tool. Invasive Plant Sci. Manage. 2010, 3, 365–375. [Google Scholar] [CrossRef]
- Tu, W.; Xiong, Q.; Qiu, X.; Zhang, Y. Dynamics of invasive alien plant species in China under climate change scenarios. Ecol. Indic. 2021, 129, 107919. [Google Scholar] [CrossRef]
- Srivastava, V.; Lafond, V.; Griess, V.C. Species distribution models (SDM): Applications, benefits and challenges in invasive species management. CAB Rev. 2019, 14, 1–13. [Google Scholar] [CrossRef]
- El-Barougy, R.F.; Dakhil, M.A.; Halmy, M.W.; Gray, S.M.; Abdelaal, M.; Khedr, A.-H.A.; Bersier, L.-F. Invasion risk assessment using trait-environment and species distribution modelling techniques in an arid protected area: Towards conservation prioritization. Ecol. Indic. 2021, 129, 107951. [Google Scholar] [CrossRef]
- Soberon, J.; Nakamura, M. Niches and distributional areas: Concepts, methods, and assumptions. Proc. Natl. Acad. Sci. USA 2009, 106 (Suppl. 2), 19644–19650. [Google Scholar] [CrossRef] [Green Version]
- Wiens, J.J.; Ackerly, D.D.; Allen, A.P.; Anacker, B.L.; Buckley, L.B.; Cornell, H.V.; Damschen, E.I.; Jonathan Davies, T.; Grytnes, J.A.; Harrison, S.P.; et al. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 2010, 13, 1310–1324. [Google Scholar] [CrossRef]
- Liu, C.; Wolter, C.; Courchamp, F.; Roura-Pascual, N.; Jeschke, J.M. Biological invasions reveal how niche change affects the transferability of species distribution models. Ecology 2022, 103, e3719. [Google Scholar] [CrossRef]
- Santini, L.; Benítez-López, A.; Maiorano, L.; Čengić, M.; Huijbregts, M.A. Assessing the reliability of species distribution projections in climate change research. Divers. Distrib. 2021, 27, 1035–1050. [Google Scholar] [CrossRef]
- Melo-Merino, S.M.; Reyes-Bonilla, H.; Lira-Noriega, A. Ecological niche models and species distribution models in marine environments: A literature review and spatial analysis of evidence. Ecol. Modell. 2020, 415, 108837. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping species distributions with MAXENT using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Wu, D.; Zhu, C. Prediction of the spatial distribution of Alternanthera philoxeroides in China based on ArcGIS and MaxEnt. Global Ecol. Conserv. 2020, 21, e00856. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander Jr, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Ma, B.; Sun, J. Predicting the distribution of Stipa purpurea across the Tibetan Plateau via the MaxEnt model. BMC Ecol. 2018, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Global Change Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Lamsal, P.; Kumar, L.; Aryal, A.; Atreya, K. Invasive alien plant species dynamics in the Himalayan region under climate change. AMBIO 2018, 47, 697–710. [Google Scholar] [CrossRef]
- Zhan, P.; Wang, F.; Xia, P.; Zhao, G.; Wei, M.; Wei, F.; Han, R. Assessment of suitable cultivation region for Panax notoginseng under different climatic conditions using MaxEnt model and high-performance liquid chromatography in China. Ind. Crops Prod. 2022, 176, 114416. [Google Scholar] [CrossRef]
- Verlinden, M.; De Boeck, H.; Nijs, I. Climate warming alters competition between two highly invasive alien plant species and dominant native competitors. Weed Res. 2014, 54, 234–244. [Google Scholar] [CrossRef]
- Zhang, P.; Li, B.; Wu, J.; Hu, S. Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecol. Lett. 2019, 22, 200–210. [Google Scholar] [CrossRef]
- Hobbs, R.J. Land-use changes and invasions. Invasive Species A Chang. World 2000, 55–64. [Google Scholar]
- Pauchard, A.; Alaback, P.B. Influence of elevation, land use, and landscape context on patterns of alien plant invasions along roadsides in protected areas of South-Central Chile. Conserv. Biol. 2004, 18, 238–248. [Google Scholar]
- Chytrý, M.; Jarošík, V.; Pyšek, P.; Hájek, O.; Knollová, I.; Tichý, L.; Danihelka, J. Separating habitat invasibility by alien plants from the actual level of invasion. Ecology 2008, 89, 1541–1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzoor, S.A.; Griffiths, G.; Lukac, M. Land use and climate change interaction triggers contrasting trajectories of biological invasion. Ecol. Indic. 2021, 120, 106936. [Google Scholar] [CrossRef]
- Bai, H.; Xiao, D.; Wang, B.; Liu, D.L.; Feng, P.; Tang, J. Multi-model ensemble of CMIP6 projections for future extreme climate stress on wheat in the North China Plain. Int. J. Climatol. 2021, 41, E171–E186. [Google Scholar] [CrossRef]
- Kamruzzaman, M.; Shahid, S.; Islam, A.; Hwang, S.; Cho, J.; Zaman, M.; Uz, A.; Ahmed, M.; Rahman, M.; Hossain, M. Comparison of CMIP6 and CMIP5 model performance in simulating historical precipitation and temperature in Bangladesh: A preliminary study. Theor. Appl. Climatol. 2021, 145, 1385–1406. [Google Scholar] [CrossRef]
- Chen, H.; Sun, J.; Lin, W.; Xu, H. Comparison of CMIP6 and CMIP5 models in simulating climate extremes. Sci. Bull. 2020, 65, 1415–1418. [Google Scholar] [CrossRef]
- Gao, T.; Xu, Q.; Liu, Y.; Zhao, J.; Shi, J. Predicting the potential geographic distribution of Sirex nitobei in China under climate change using maximum entropy model. Forests 2021, 12, 151. [Google Scholar] [CrossRef]
- Ramasamy, M.; Das, B.; Ramesh, R. Predicting climate change impacts on potential worldwide distribution of fall armyworm based on cmip6 projections. J. Pest. Sci. 2022, 95, 841–854. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Xu, X.; Liu, J.; Zhang, S.; Li, R.; Yan, C.; Wu, S. China Multi-period Land Use land Cover Remote sensing Monitoring Data Set (CNLUCC) Data Registration and Publishing System of Resource and Environmental Science Data Center. Chin. Acad. Sci. 2018, 35, 315–325. [Google Scholar]
- Nachtergaele, F.; Velthuizen, H.v.; Verelst, L.; Wilberg, D. Harmonized World Soil Database (version 1.2); FAO: Rome, Italy; IIASA: Laxenburg, Austria, 2012. [Google Scholar]
- Hengl, T.; Mendes de Jesus, J.; Heuvelink, G.B.; Ruiperez Gonzalez, M.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef] [PubMed]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Osorio-Olvera, L.; Jiménez-García, D. An exhaustive analysis of heuristic methods for variable selection in ecological niche modeling and species distribution modeling. Ecol. Inf. 2019, 53, 100983. [Google Scholar] [CrossRef]
- Peterson, B.G.; Carl, P.; Boudt, K.; Bennett, R.; Ulrich, J.; Zivot, E.; Lestel, M.; Balkissoon, K.; Wuertz, D. PerformanceAnalytics: Econometric tools for performance and risk analysis. R Package Version 2014, 1, 3541. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing, Version 3.6.1; R Foundation for Statistical Computin: Vienna, Austria, 2019. [Google Scholar]
- Petrie, R.; Denvil, S.; Ames, S.; Levavasseur, G.; Fiore, S.; Allen, C.; Antonio, F.; Berger, K.; Bretonnière, P.-A.; Cinquini, L. Coordinating an operational data distribution network for CMIP6 data. Geosci. Model Dev. 2021, 14, 629–644. [Google Scholar] [CrossRef]
- Hamed, M.M.; Nashwan, M.S.; Shahid, S.; bin Ismail, T.; Wang, X.-J.; Dewan, A.; Asaduzzaman, M. Inconsistency in historical simulations and future projections of temperature and rainfall: A comparison of CMIP5 and CMIP6 models over Southeast Asia. Atmos. Res. 2022, 265, 105927. [Google Scholar] [CrossRef]
- Wu, T.; Yu, R.; Lu, Y.; Jie, W.; Fang, Y.; Zhang, J.; Zhang, L.; Xin, X.; Li, L.; Wang, Z. BCC-CSM2-HR: A high-resolution version of the Beijing Climate Center Climate System Model. Geosci. Model Dev. 2021, 14, 2977–3006. [Google Scholar] [CrossRef]
- Meinshausen, M.; Nicholls, Z.R.; Lewis, J.; Gidden, M.J.; Vogel, E.; Freund, M.; Beyerle, U.; Gessner, C.; Nauels, A.; Bauer, N. The shared socio-economic pathway (SSP) greenhouse gas concentrations and their extensions to 2500. Geosci. Model Dev. 2020, 13, 3571–3605. [Google Scholar] [CrossRef]
- Wei, B.; Liu, L.; Gu, C.; Yu, H.; Zhang, Y.; Zhang, B.; Cui, B.; Gong, D.; Tu, Y. The climate niche is stable and the distribution area of Ageratina adenophora is predicted to expand in China. Biodivers. Sci. 2022, 30, 21443. [Google Scholar] [CrossRef]
- Muniappan, R.; Marutani, M. Ecology and distribution of Chromolaena odorata in Asia and the Pacific. In Proceedings of the 1st International Workshop on Biological Control of Chromolaena Odorata, Mangilao, Guam, 29 February–4 March 1988; pp. 21–24. [Google Scholar]
- Fang, Y.; Zhang, X.; Wei, H.; Wang, D.; Chen, R.; Wang, L.; Gu, W. Predicting the invasive trend of exotic plants in China based on the ensemble model under climate change: A case for three invasive plants of Asteraceae. Sci. Total Environ. 2021, 756, 143841. [Google Scholar] [CrossRef]
- Lu, Z.; Ma, K. Effects of topographical factors on the invasive species Ageratina adenophora. Chin. J. Plant Ecol. 2004, 28, 761. [Google Scholar]
- Changjun, G.; Yanli, T.; Linshan, L.; Bo, W.; Yili, Z.; Haibin, Y.; Xilong, W.; Zhuoga, Y.; Binghua, Z.; Bohao, C. Predicting the potential global distribution of Ageratina adenophora under current and future climate change scenarios. Ecol. Evol. 2021, 11, 12092–12113. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Sun, O.J.; Sang, W.; Li, Z.; Ma, K. Predicting the spatial distribution of an invasive plant species (Eupatorium adenophorum) in China. Landsc. Ecol. 2007, 22, 1143–1154. [Google Scholar] [CrossRef]
- Kaufmann, R.K.; Stern, D.I. Evidence for human influence on climate from hemispheric temperature relations. Nature 1997, 388, 39–44. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. BIOCLIM: The first species distribution modelling package, its early applications and relevance to most current MAXENT studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Weterings, R.; Vetter, K.C. Invasive house geckos (Hemidactylus spp.): Their current, potential and future distribution. Curr. Zool. 2018, 64, 559–573. [Google Scholar] [CrossRef]
- Sun, X.y.; Lu, Z.h.; Sang, W.g. Review on studies of Eupatorium adenophorum--an important invasive species in China. Chin. J. For. Res. 2004, 15, 319–322. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environment Variables | Description | Abbreviation |
|---|---|---|
| Climate | Mean diurnal range (mean of monthly (max temp–min temp)) (0.1 °C) | bio_2 |
| Temperature annual range (0.1 °C) | bio_7 | |
| Mean temperature of driest quarter (0.1 °C) | bio_9 | |
| Annual precipitation (mm) | bio_12 | |
| Precipitation of driest quarter (mm) | bio_17 | |
| Soil | Soil type | soil |
| Soil texture | Sand | sand |
| Silt | silt | |
| Soil | clay | |
| Soil quality | Nutrient availability | sq1 |
| Nutrient retention capacity | sq2 | |
| Rooting conditions | sq3 | |
| Excess salts | sq5 | |
| Workability (constraining field management) | sq7 | |
| Land use type | Land use/cover change | lucc |
| Human activity | Human footprint dataset | hii |
| Terrain | DEM | dem |
| Slope | slope | |
| Aspect | aspect |
| Variable | Percent Contribution (%) | Permutation Importance (%) |
|---|---|---|
| dem | 32.8 | 15.4 |
| bio_7 | 27.5 | 28 |
| bio_12 | 12.6 | 26.3 |
| hii | 9.1 | 5.2 |
| bio_9 | 4.2 | 5.1 |
| bio_3 | 4.2 | 2.3 |
| bio_19 | 2.7 | 7.3 |
| slope | 2.1 | 1.3 |
| soil | 1.4 | 0.4 |
| aspect | 1.3 | 1.1 |
| sq3 | 0.5 | 1.6 |
| sq4 | 0.4 | 1.4 |
| sq1 | 0.4 | 0.3 |
| bio_17 | 0.4 | 2.5 |
| sq5 | 0.3 | 1.7 |
| Variable | Percent Contribution (%) | Permutation Importance (%) |
|---|---|---|
| bio_7 | 27.3 | 33.3 |
| dem | 22.5 | 14.8 |
| bio_12 | 20.4 | 0.2 |
| bio_9 | 16.4 | 37.9 |
| hii | 7.8 | 5.7 |
| bio_2 | 1.7 | 0.1 |
| sand | 0.7 | 0.4 |
| clay | 0.6 | 0.6 |
| bio_17 | 0.5 | 3 |
| aspect | 0.4 | 0.5 |
| sq2 | 0.4 | 0.6 |
| soil | 0.3 | 0 |
| silt | 0.3 | 1.4 |
| sq1 | 0.2 | 0.3 |
| sq3 | 0.1 | 0.6 |
| lucc | 0.1 | 0.2 |
| slope | 0.1 | 0.2 |
| sq5 | 0.1 | 0.3 |
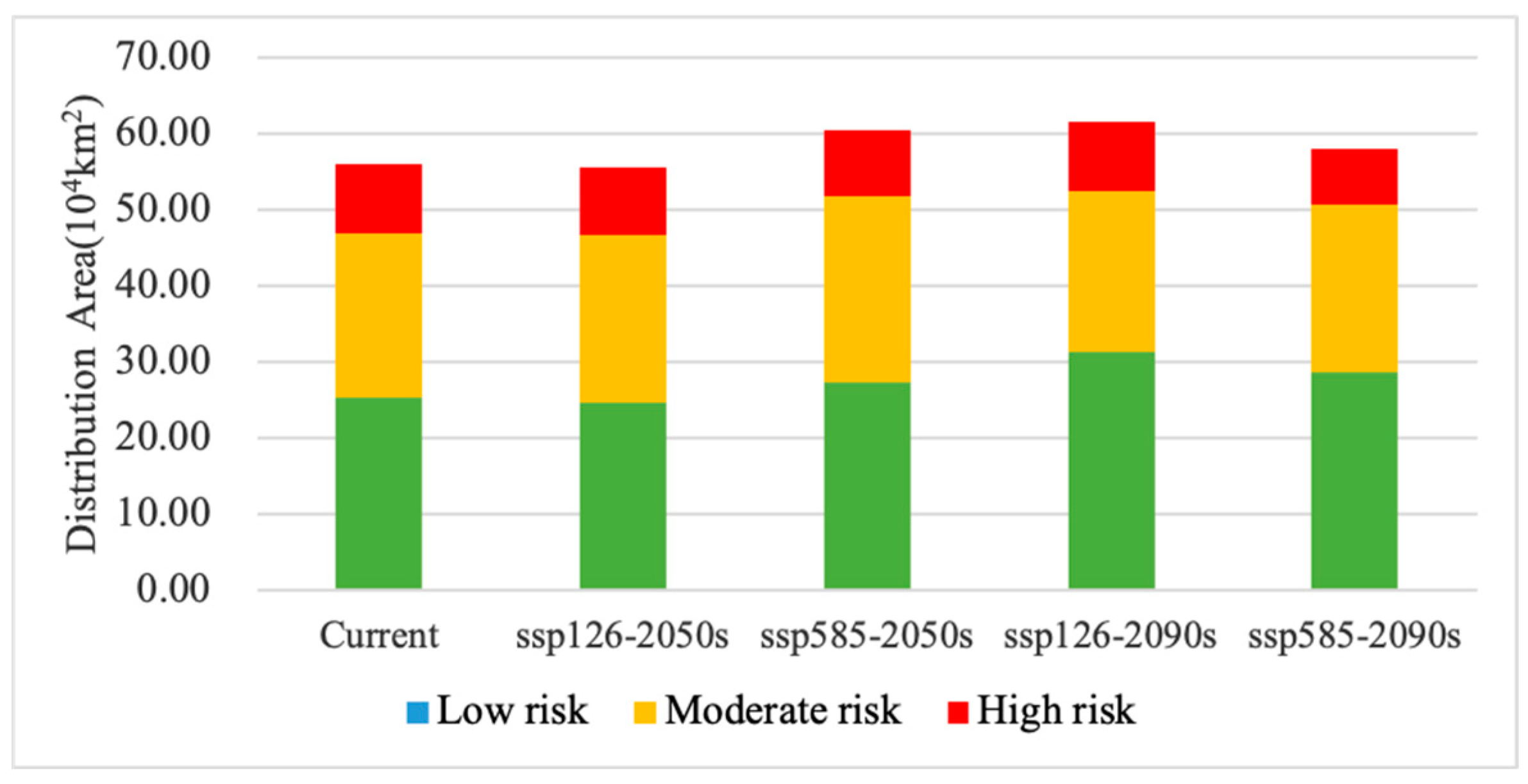
| Climate Scenarios | Area (×104 km2) | Area Ratio (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Low Risk | Moderate Risk | High Risk | Total Area | Low Risk | Moderate Risk | High Risk | ||
| Current | - | 25.41 | 21.40 | 9.30 | 56.11 | 45.29 | 38.13 | 16.58 |
| 2050s | ssp126 | 24.66 | 22.05 | 8.83 | 55.53 | 44.40 | 39.70 | 15.90 |
| ssp585 | 27.36 | 24.38 | 8.76 | 60.50 | 45.22 | 40.31 | 14.48 | |
| 2090s | ssp126 | 31.33 | 21.09 | 9.27 | 61.69 | 50.79 | 34.19 | 15.02 |
| ssp585 | 28.77 | 21.90 | 7.43 | 58.10 | 49.52 | 37.70 | 12.78 | |
| Climate Scenarios | Area (×104 km2) | Area Ratio (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Contraction | Unchanged | Expansion | Total Change | Contraction | Unchanged | Expansion | Total Change | ||
| 2050s | ssp126 | 5.32 | 50.79 | 4.75 | −0.57 | 9.49% | 90.51% | 8.46% | −1.02% |
| ssp585 | 2.61 | 53.50 | 7.00 | 4.39 | 4.65% | 95.35% | 12.47% | 7.83% | |
| 2090s | ssp126 | 1.93 | 54.17 | 7.52 | 5.59 | 3.45% | 96.55% | 13.40% | 9.96% |
| ssp585 | 4.61 | 51.50 | 6.60 | 1.99 | 8.22% | 91.78% | 11.77% | 3.55% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Wang, Y.; Peng, P.; Wang, G.; Zhao, G.; Zhou, Y.; Tang, Z. Mapping the Distribution and Dispersal Risks of the Alien Invasive Plant Ageratina adenophora in China. Diversity 2022, 14, 915. https://doi.org/10.3390/d14110915
Zhang X, Wang Y, Peng P, Wang G, Zhao G, Zhou Y, Tang Z. Mapping the Distribution and Dispersal Risks of the Alien Invasive Plant Ageratina adenophora in China. Diversity. 2022; 14(11):915. https://doi.org/10.3390/d14110915
Chicago/Turabian StyleZhang, Xiaojuan, Yanru Wang, Peihao Peng, Guoyan Wang, Guanyue Zhao, Yongxiu Zhou, and Zihao Tang. 2022. "Mapping the Distribution and Dispersal Risks of the Alien Invasive Plant Ageratina adenophora in China" Diversity 14, no. 11: 915. https://doi.org/10.3390/d14110915
APA StyleZhang, X., Wang, Y., Peng, P., Wang, G., Zhao, G., Zhou, Y., & Tang, Z. (2022). Mapping the Distribution and Dispersal Risks of the Alien Invasive Plant Ageratina adenophora in China. Diversity, 14(11), 915. https://doi.org/10.3390/d14110915