
Understanding Spatiotemporal Variation in Richness and Rate of Within-Site Turnover for Vegetation Communities in Western Eurasia over the Last 4000 Years


Abstract
:1. Introduction
2. Materials and Methods
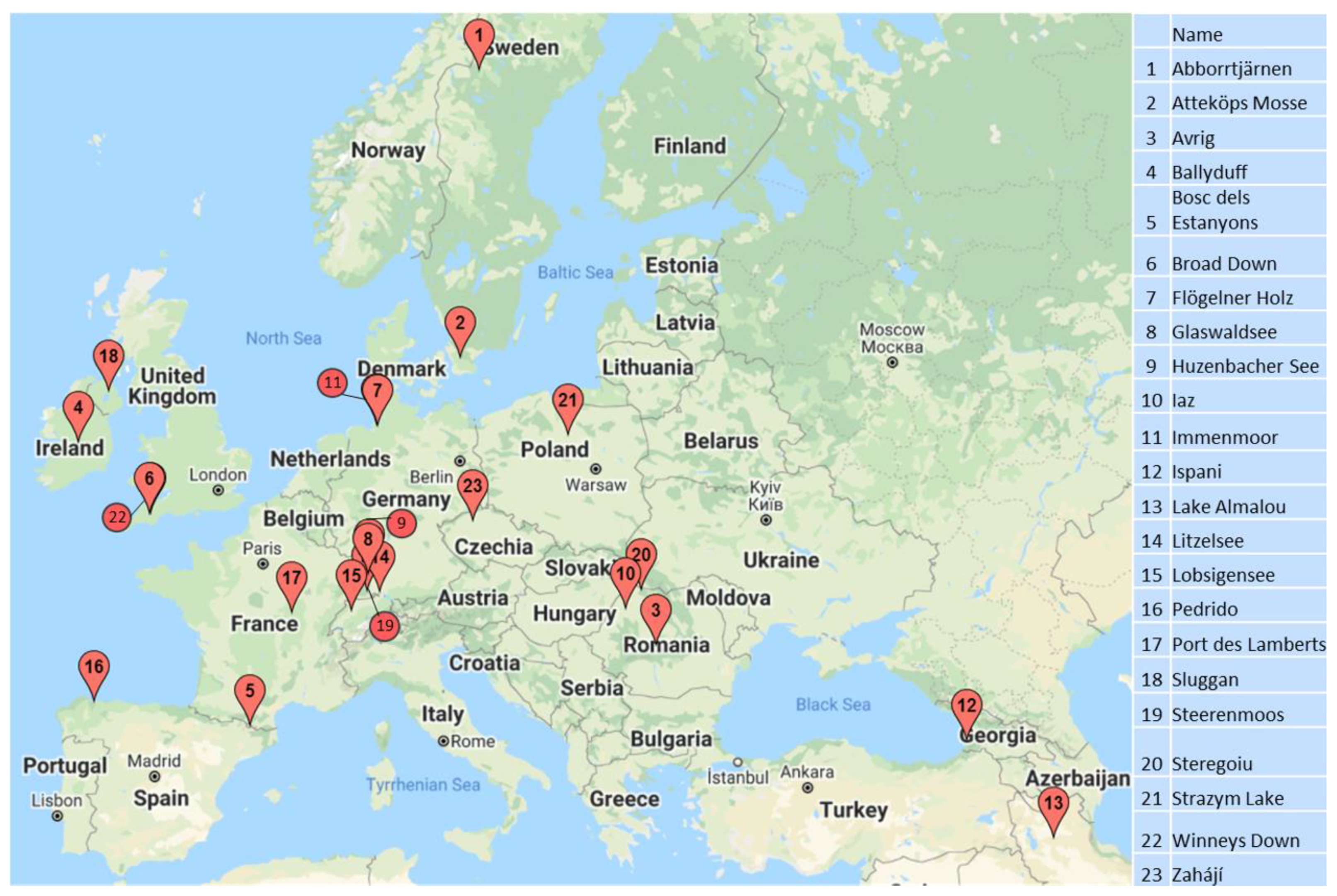
2.1. Data Acquisition
2.2. Data Manipulation
2.3. Data Analysis
3. Results
3.1. Spatiotemporal Change in Taxonomic Richness: Univariate Patterns
3.2. Spatiotemporal Change in Taxonomic Richness: Multivariate Modelling
3.3. Spatiotemporal Change in Rate of Within-Site Turnover
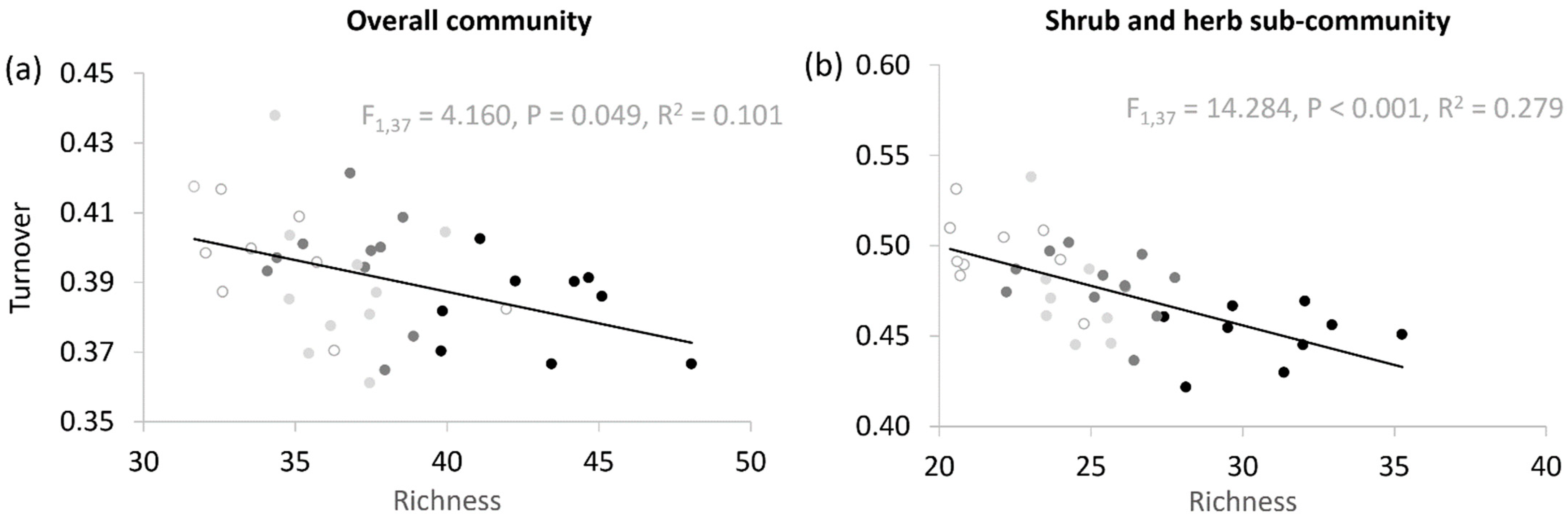
3.4. Relationship between Richness and Rate of Within-Site Turnover: Is the Diversity-Stability Hypothesis Supported?
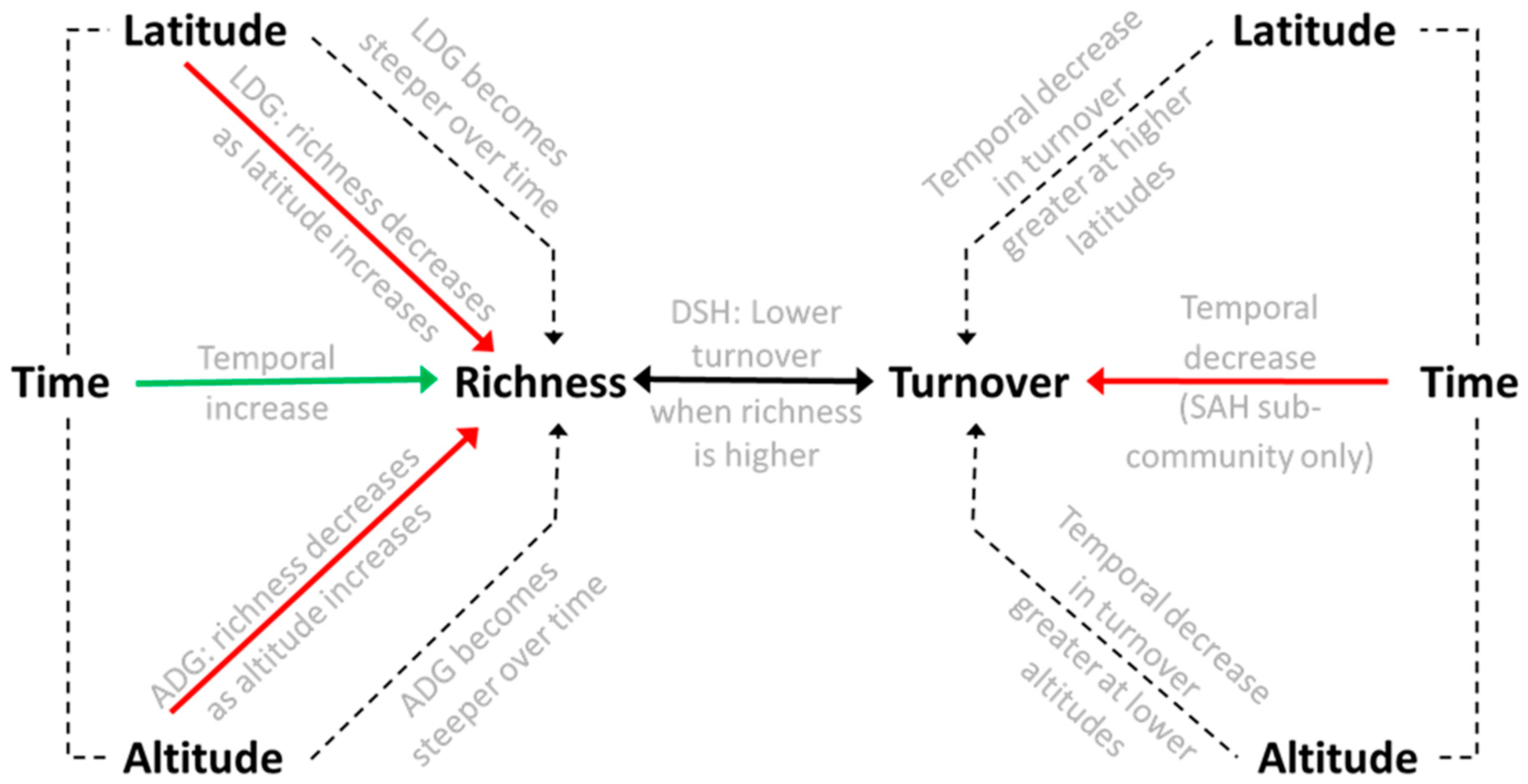
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Millien, V.; Kathleen Lyons, S.; Olson, L.; Smith, F.A.; Wilson, A.B.; Yom-Tov, Y. Ecotypic Variation in the Context of Global Climate Change: Revisiting the Rules. Ecol. Lett. 2006, 9, 853–869. [Google Scholar] [CrossRef] [PubMed]
- Mannion, P.D. A Deep-Time Perspective on the Latitudinal Diversity Gradient. Proc. Natl. Acad. Sci. USA 2020, 117, 17479–17481. [Google Scholar] [CrossRef] [PubMed]
- Ives, A.R.; Carpenter, S.R. Stability and Diversity of Ecosystems. Science 2007, 317, 58–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rull, V. Ecology and Palaeoecology: Two Approaches, One Objective. Open Ecol. J. 2010, 3, 102–116. [Google Scholar] [CrossRef] [Green Version]
- Rull, V. Community Ecology: Diversity and Dynamics over Time. Community Ecol. 2012, 13, 102–116. [Google Scholar] [CrossRef]
- Berglund, B.E.; Gaillard, M.-J.; Björkman, L.; Persson, T. Long-Term Changes in Floristic Diversity in Southern Sweden: Palynological Richness, Vegetation Dynamics and Land-Use. Veg. Hist. Archaeobot. 2008, 17, 573–583. [Google Scholar] [CrossRef]
- Salgado, J.; Sayer, C.; Carvalho, L.; Davidson, T.; Gunn, I. Assessing Aquatic Macrophyte Community Change through the Integration of Palaeolimnological and Historical Data at Loch Leven, Scotland. J. Paleolimnol. 2010, 43, 191–204. [Google Scholar] [CrossRef]
- Bishop, I.J.; Bennion, H.; Sayer, C.D.; Patmore, I.R.; Yang, H. Filling the “Data Gap”: Using Paleoecology to Investigate the Decline of Najas Flexilis (a Rare Aquatic Plant). Geo 2019, 6, e00081. [Google Scholar] [CrossRef]
- Bennion, H.; Sayer, C.D.; Clarke, S.J.; Davidson, T.A.; Rose, N.L.; Goldsmith, B.; Rawcliffe, R.; Burgess, A.; Clarke, G.; Turner, S. Sedimentary Macrofossil Records Reveal Ecological Change in English Lakes: Implications for Conservation. J. Paleolimnol. 2018, 60, 329–348. [Google Scholar] [CrossRef] [Green Version]
- Seddon, E.; Hill, M.; Greenwood, M.T.; Mainstone, C.; Mathers, K.; White, J.C.; Wood, P.J. The Use of Palaeoecological and Contemporary Macroinvertebrate Community Data to Characterize Riverine Reference Conditions. River. Res. Appl. 2019, 35, 1302–1313. [Google Scholar] [CrossRef]
- Agiadi, K.; Albano, P.G. Holocene Fish Assemblages Provide Baseline Data for the Rapidly Changing Eastern Mediterranean. Holocene 2020, 30, 1438–1450. [Google Scholar] [CrossRef]
- Buma, B.; Harvey, B.J.; Gavin, D.G.; Kelly, R.; Loboda, T.; McNeil, B.E.; Marlon, J.R.; Meddens, A.J.H.; Morris, J.L.; Raffa, K.F. The Value of Linking Paleoecological and Neoecological Perspectives to Understand Spatially-Explicit Ecosystem Resilience. Landsc. Ecol. 2019, 34, 17–33. [Google Scholar] [CrossRef]
- Goodenough, A.E.; Webb, J.C. Learning from the Past: Opportunities for Advancing Ecological Research and Practice Using Palaeoecological Data. Oecologia 2022, 199, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Willis, K.J.; Gillson, L.; Knapp, S. Biodiversity Hotspots through Time: An Introduction. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 169–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rull, V.; Vegas-Vilarrúbia, T. What Is Long-Term in Ecology? Trends Ecol. Evol. 2010, 26, 3–4. [Google Scholar] [CrossRef]
- Birks, H.J.B. Contributions of Quaternary Botany to Modern Ecology and Biogeography. Plant Ecol. Divers. 2019, 12, 189–385. [Google Scholar] [CrossRef]
- Jackson, S.T. Looking Forward from the Past: History, Ecology, and Conservation. Front. Ecol. Environ. 2007, 5, 455. [Google Scholar] [CrossRef]
- Colombaroli, D.; Tinner, W. Determining the Long-Term Changes in Biodiversity and Provisioning Services along a Transect from Central Europe to the Mediterranean. Holocene 2013, 23, 1625–1634. [Google Scholar] [CrossRef]
- Hillebrand, H. On the Generality of the Latitudinal Diversity Gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [Green Version]
- Kinlock, N.L.; Prowant, L.; Herstoff, E.M.; Foley, C.M.; Akin-Fajiye, M.; Bender, N.; Umarani, M.; Ryu, H.Y.; Şen, B.; Gurevitch, J. Explaining Global Variation in the Latitudinal Diversity Gradient: Meta-analysis Confirms Known Patterns and Uncovers New Ones. Glob. Ecol. Biogeogr. 2018, 27, 125–141. [Google Scholar] [CrossRef]
- Fischer, A.; Blaschke, M.; Bässler, C. Altitudinal Gradients in Biodiversity Research: The State of the Art and Future Perspectives under Climate Change Aspects. Wald. Landsch. Und Nat. 2011, 11, 35–47. [Google Scholar]
- Scheiner, S.M.; Chiarucci, A.; Fox, G.A.; Helmus, M.R.; McGlinn, D.J.; Willig, M.R. The Underpinnings of the Relationship of Species Richness with Space and Time. Ecol. Monogr. 2011, 81, 195–213. [Google Scholar] [CrossRef] [Green Version]
- Sax, D.F.; Gaines, S.D. Species Diversity: From Global Decreases to Local Increases. Trends Ecol. Evol. 2003, 18, 561–566. [Google Scholar] [CrossRef]
- Vellend, M.; Baeten, L.; Myers-Smith, I.H.; Elmendorf, S.C.; Beauséjour, R.; Brown, C.D.; de Frenne, P.; Verheyen, K.; Wipf, S. Global Meta-Analysis Reveals No Net Change in Local-Scale Plant Biodiversity over Time. Proc. Natl. Acad. Sci. USA 2013, 110, 19456–19459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dornelas, M.; Gotelli, N.J.; McGill, B.; Shimadzu, H.; Moyes, F.; Sievers, C.; Magurran, A.E. Assemblage Time Series Reveal Biodiversity Change but Not Systematic Loss. Science 2014, 344, 296–299. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, A.; Cardinale, B.J.; Allington, G.R.H.; Byrnes, J.; Arthur Endsley, K.; Brown, D.G.; Hooper, D.U.; Isbell, F.; O’Connor, M.I.; Loreau, M. Estimating Local Biodiversity Change: A Critique of Papers Claiming No Net Loss of Local Diversity. Ecology 2016, 97, 1949–1960. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Gonzalez, A.; Allington, G.R.H.; Loreau, M. Is Local Biodiversity Declining or Not? A Summary of the Debate over Analysis of Species Richness Time Trends. Biol. Conserv. 2018, 219, 175–183. [Google Scholar] [CrossRef]
- Silvertown, J. History of a Latitudinal Diversity Gradient: Woody Plants in Europe 13,000-1000 Years BP. J. Biogeogr. 1985, 12, 519–525. [Google Scholar] [CrossRef]
- Giesecke, T.; Brewer, S.; Finsinger, W.; Leydet, M.; Bradshaw, R.H.W. Patterns and Dynamics of European Vegetation Change over the Last 15,000 Years. J. Biogeogr. 2017, 44, 1441–1456. [Google Scholar] [CrossRef] [Green Version]
- Hillebrand, H.; Soininen, J.; Snoeijs, P. Warming Leads to Higher Species Turnover in a Coastal Ecosystem. Glob. Chang. Biol. 2010, 16, 1181–1193. [Google Scholar] [CrossRef]
- Ulrich, W.; Soliveres, S.; Maestre, F.T.; Gotelli, N.J.; Quero, J.L.; Delgado-Baquerizo, M.; Bowker, M.A.; Eldridge, D.J.; Ochoa, V.; Gozalo, B. Climate and Soil Attributes Determine Plant Species Turnover in Global Drylands. J. Biogeogr. 2014, 41, 2307–2319. [Google Scholar] [CrossRef] [PubMed]
- Gibson-Reinemer, D.K.; Sheldon, K.S.; Rahel, F.J. Climate Change Creates Rapid Species Turnover in Montane Communities. Ecol. Evol. 2015, 5, 2340–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, C.S.; Reside, A.E.; VanDerWal, J.; Pearson, R.G.; Burrows, D.; Capon, S.J.; Harwood, T.D.; Hodgson, L.; Waltham, N.J. Sink or Swim? Potential for High Faunal Turnover in Australian Rivers under Climate Change. J. Biogeogr. 2017, 44, 489–501. [Google Scholar] [CrossRef]
- Webb, J.C.; Goodenough, A.E. Vegetation Community Changes in European Woodlands amid a Changing Climate: A Palaeoecological Modelling Perspective. Community Ecol. 2021, 22, 319–330. [Google Scholar] [CrossRef]
- Xu, Z.; Wan, S.; Ren, H.; Han, X.; Li, M.-H.; Cheng, W.; Jiang, Y. Effects of Water and Nitrogen Addition on Species Turnover in Temperate Grasslands in Northern China. PLoS ONE 2012, 7, e39762. [Google Scholar] [CrossRef] [PubMed]
- Rahel, F.J. Homogenization of Fish Faunas across the United States. Science 2000, 288, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, J.J.; Soininen, J.; Hillebrand, H. A Quantitative Analysis of Temporal Turnover in Aquatic Species Assemblages across Ecosystems. Ecology 2010, 91, 508–517. [Google Scholar] [CrossRef]
- Mottl, O.; Grytnes, J.-A.; Seddon, A.W.R.; Steinbauer, M.J.; Bhatta, K.P.; Felde, V.A.; Flantua, S.G.A.; Birks, H.J.B. Rate-of-Change Analysis in Paleoecology Revisited: A New Approach. Rev. Palaeobot. Palynol. 2021, 293, 104483. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species; John Murray: London, UK, 1859. [Google Scholar]
- MacArthur, R. Fluctuations of Animal Populations and a Measure of Community Stability. Ecology 1955, 36, 533–536. [Google Scholar] [CrossRef]
- McNaughton, S.J. Diversity and Stability of Ecological Communities: A Comment on the Role of Empiricism in Ecology. Am. Nat. 1977, 111, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.A.; McNaughton, S.J. Stability Increases with Diversity in Plant Communities: Empirical Evidence from the 1988 Yellowstone Drought. Oikos 1991, 62, 360–362. [Google Scholar] [CrossRef]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and Sustainability Influenced by Biodiversity in Grassland Ecosystems. Nature 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Fyfe, R.M.; de Beaulieu, J.-L.; Binney, H.; Bradshaw, R.H.W.; Brewer, S.; le Flao, A.; Finsinger, W.; Gaillard, M.-J.; Giesecke, T.; Gil-Romera, G.; et al. The European Pollen Database: Past Efforts and Current Activities. Veg. Hist. Archaeobot. 2009, 18, 417–424. [Google Scholar] [CrossRef] [Green Version]
- Mottl, O.; Flantua, S.G.A.; Bhatta, K.P.; Felde, V.A.; Giesecke, T.; Goring, S.; Grimm, E.C.; Haberle, S.; Hooghiemstra, H.; Ivory, S.; et al. Global Acceleration in Rates of Vegetation Change over the Past 18,000 Years. Science 2021, 372, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Jaccard, P. The Distribution of the Flora in the Alpine Zone. 1. New Phytol. 1912, 11, 37–50. [Google Scholar] [CrossRef]
- Hart, A.G.; Dawson, M.; Fourie, R.; MacTavish, L.; Goodenough, A.E. Comparing the Effectiveness of Camera Trapping, Driven Transects and Ad Hoc Records for Surveying Nocturnal Mammals against a Known Species Assemblage. Community Ecol. 2022, 23, 27–39. [Google Scholar] [CrossRef]
- Birks, H.J.B. Estimating the Amount of Compositional Change in Late-Quaternary Pollen-Stratigraphical Data. Veg. Hist. Archaeobot. 2007, 16, 197–202. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Line, J.M. The Use of Rarefaction Analysis for Estimating Palynological Richness from Quaternary Pollen-Analytical Data. Holocene 1992, 2, 1–10. [Google Scholar] [CrossRef]
- Reille, M.; Pons, A. Leçons de Palynologie et d’analyse Pollinique; Editions du Centre National de la Recherche Scientifique: Paris, France, 1990. [Google Scholar]
- Moore, P.; Webb, J.; Collinson, M. Pollen Analysis; Blackwell Scientific: Oxford, UK, 1991. [Google Scholar]
- Djamali, M.; Cilleros, K. Statistically Significant Minimum Pollen Count in Quaternary Pollen Analysis; the Case of Pollen-Rich Lake Sediments. Rev. Palaeobot. Palynol. 2020, 275, 104156. [Google Scholar] [CrossRef]
- Akaike, H. Information Theory and an Extension of the Maximum Likelihood Principle. In Second International Symposium on Information Theory; Petrov, B.V., Csaki, B.F., Eds.; Academia Kiado: Budapest, Hungary, 1973; pp. 267–281. [Google Scholar]
- Hu, M.-C.; Pavlicova, M.; Nunes, E.V. Zero-Inflated and Hurdle Models of Count Data with Extra Zeros: Examples from an HIV-Risk Reduction Intervention Trial. Am. J. Drug Alcohol. Abuse 2011, 37, 367–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reitalu, T.; Bjune, A.E.; Blaus, A.; Giesecke, T.; Helm, A.; Matthias, I.; Peglar, S.M.; Salonen, J.S.; Seppä, H.; Väli, V. Patterns of Modern Pollen and Plant Richness across Northern Europe. J. Ecol. 2019, 107, 1662–1677. [Google Scholar] [CrossRef]
- Mannion, P.D.; Upchurch, P.; Benson, R.B.J.; Goswami, A. The Latitudinal Biodiversity Gradient through Deep Time. Trends Ecol. Evol. 2014, 29, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, P.J.; Fox, D.L.; Marcot, J.; Badgley, C. Flat Latitudinal Gradient in Paleocene Mammal Richness Suggests Decoupling of Climate and Biodiversity. Geology 2011, 39, 163–166. [Google Scholar] [CrossRef]
- Song, H.; Huang, S.; Jia, E.; Dai, X.; Wignall, P.B.; Dunhill, A.M. Flat Latitudinal Diversity Gradient Caused by the Permian–Triassic Mass Extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 17578–17583. [Google Scholar] [CrossRef] [PubMed]
- Colombaroli, D.; Beckmann, M.; van der Knaap, W.O.; Curdy, P.; Tinner, W. Changes in Biodiversity and Vegetation Composition in the Central S Wiss A Lps during the Transition from Pristine Forest to First Farming. Divers. Distrib. 2013, 19, 157–170. [Google Scholar] [CrossRef]
- Schwörer, C.; Colombaroli, D.; Kaltenrieder, P.; Rey, F.; Tinner, W. Early Human Impact (5000–3000 BC) Affects Mountain Forest Dynamics in the A Lps. J. Ecol. 2015, 103, 281–295. [Google Scholar] [CrossRef]
- Hillebrand, H.; Blasius, B.; Borer, E.T.; Chase, J.M.; Downing, J.A.; Eriksson, B.K.; Filstrup, C.T.; Harpole, W.S.; Hodapp, D.; Larsen, S. Biodiversity Change Is Uncoupled from Species Richness Trends: Consequences for Conservation and Monitoring. J. Appl. Ecol. 2018, 55, 169–184. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.T.; Sax, D.F. Balancing Biodiversity in a Changing Environment: Extinction Debt, Immigration Credit and Species Turnover. Trends Ecol. Evol. 2010, 25, 153–160. [Google Scholar] [CrossRef]
- Clavel, J.; Julliard, R.; Devictor, V. Worldwide Decline of Specialist Species: Toward a Global Functional Homogenization? Front. Ecol. Environ. 2011, 9, 222–228. [Google Scholar] [CrossRef] [Green Version]
- le Viol, I.; Jiguet, F.; Brotons, L.; Herrando, S.; Lindström, Å.; Pearce-Higgins, J.W.; Reif, J.; van Turnhout, C.; Devictor, V. More and More Generalists: Two Decades of Changes in the European Avifauna. Biol. Lett. 2012, 8, 780–782. [Google Scholar] [CrossRef]
- Tweddle, J.C.; Edwards, K.J. Pollen Preservation Zones as an Interpretative Tool in Holocene Palynology. Rev. Palaeobot. Palynol. 2010, 161, 59–76. [Google Scholar] [CrossRef]
- Gámez-Virués, S.; Perović, D.J.; Gossner, M.M.; Börschig, C.; Blüthgen, N.; de Jong, H.; Simons, N.K.; Klein, A.-M.; Krauss, J.; Maier, G. Landscape Simplification Filters Species Traits and Drives Biotic Homogenization. Nat. Commun. 2015, 6, 8568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devictor, V.; Julliard, R.; Clavel, J.; Jiguet, F.; Lee, A.; Couvet, D. Functional Biotic Homogenization of Bird Communities in Disturbed Landscapes. Glob. Ecol. Biogeogr. 2008, 17, 252–261. [Google Scholar] [CrossRef]
- Goodenough, A.E.; Hart, A.G. Correlates of Vulnerability to Climate-Induced Distribution Changes in European Avifauna: Habitat, Migration and Endemism. Clim. Chang. 2013, 118, 659–669. [Google Scholar] [CrossRef]
- Walther, G.; Beißner, S.; Burga, C.A. Trends in the Upward Shift of Alpine Plants. J. Veg. Sci. 2005, 16, 541–548. [Google Scholar] [CrossRef]
- Rohde, K. Latitudinal Gradients in Species Diversity: The Search for the Primary Cause. Oikos 1992, 65, 514–527. [Google Scholar] [CrossRef] [Green Version]
- Erwin, D.H. Climate as a Driver of Evolutionary Change. Curr. Biol. 2009, 19, R575–R583. [Google Scholar] [CrossRef] [Green Version]
- Vogel, A.; Scherer-Lorenzen, M.; Weigelt, A. Grassland Resistance and Resilience after Drought Depends on Management Intensity and Species Richness. PLoS ONE 2012, 7, e36992. [Google Scholar] [CrossRef]
- Gosling, W.D.; Julier, A.; Adu-Bredu, S.; Djagbletey, G.D.; Fraser, W.T.; Jardine, P.E.; Lomax, B.H.; Malhi, Y.; Manu, E.A.; Mayle, F.E. Pollen-Vegetation Richness and Diversity Relationships in the Tropics. Veg. Hist. Archaeobot. 2018, 27, 411–418. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overall Vegetation Community | Shrub and Herb Sub-Community | Relationship Direction and Interpretation | |||
|---|---|---|---|---|---|
| x2 | p | x2 | p | ||
| Overall Model (df = 5) | 1145 | <0.001 | 1293 | <0.001 | N/A |
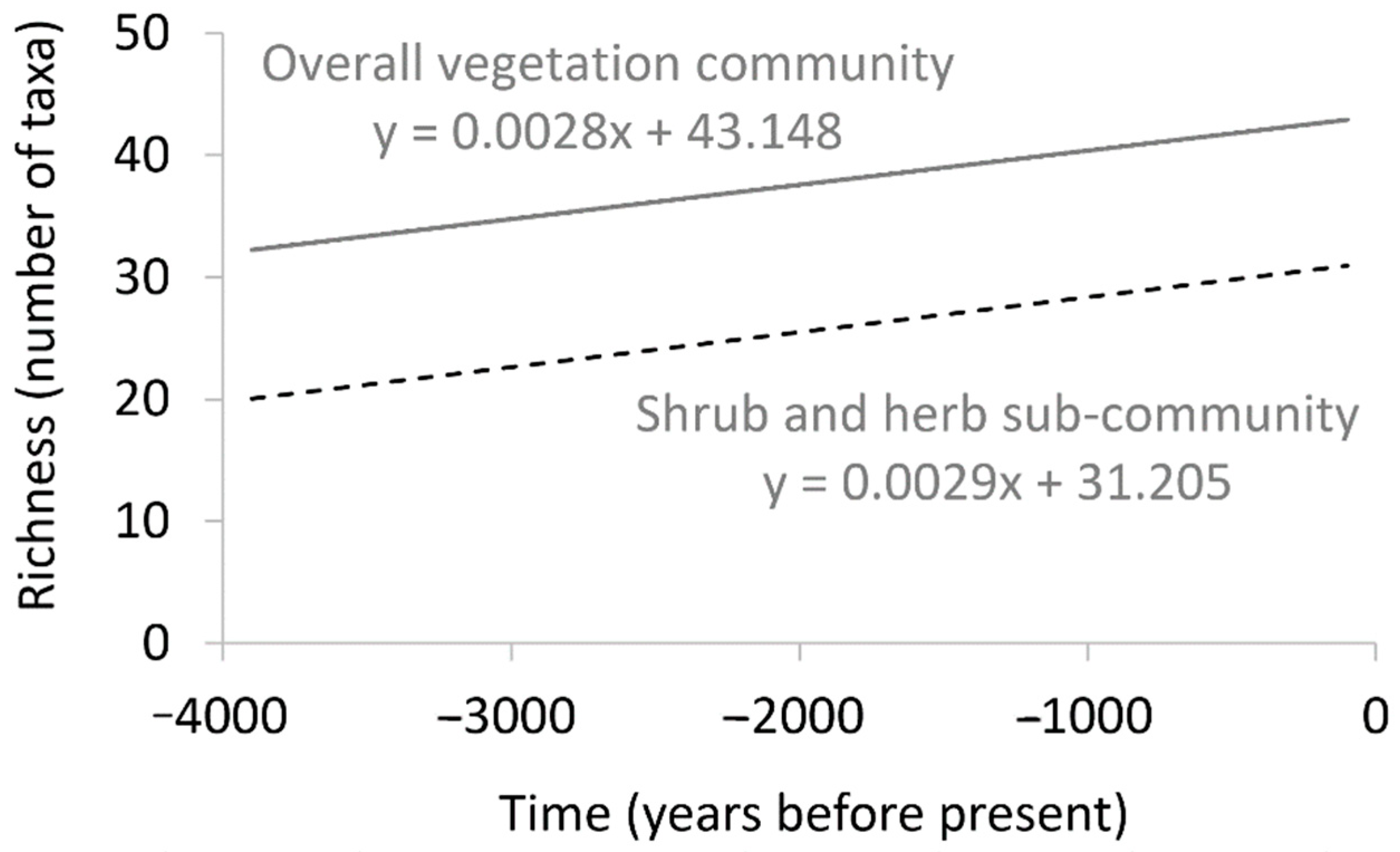
| Time (df = 1) | 35 | <0.001 | 22 | <0.001 | Positive in both models: richness higher in more recent times compared to historically |
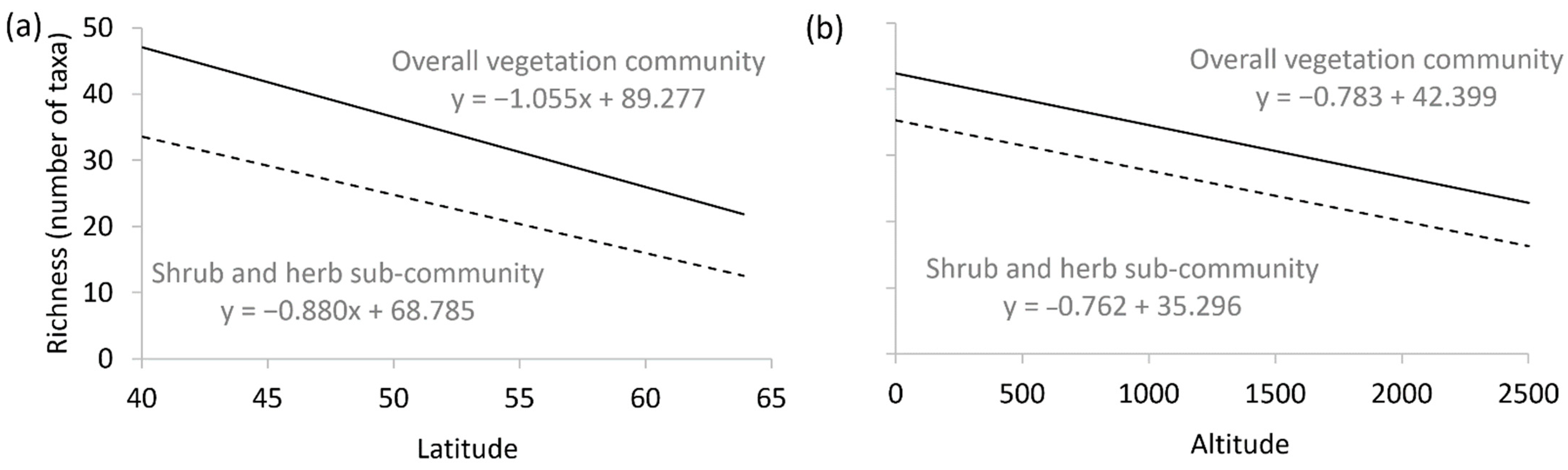
| Latitude (df = 1) | 162 | <0.001 | 102 | <0.001 | Negative in both models: richness higher at lower latitudes as predicted by Latitudinal Diversity Gradient (LDG) |
| Altitude (df = 1) | 16 | <0.001 | 2 | 0.169 | Negative for overall vegetation community: richness higher at lower altitudes as predicted by Altitudinal Diversity Gradient (ADG). No pattern for sub-community |
| Latitude*Time (df = 1) | 18 | <0.001 | 8 | 0.005 | Negative in both models: LDG stronger in more recent times compared to historically |
| Altitude*Time (df = 1) | 54 | <0.001 | 47 | <0.001 | Negative in both models: ADG stronger in more recent times compared to historically |
| Overall Vegetation Community | Shrub and Herb Sub-Community | Relationship Direction and Interpretation | |||
|---|---|---|---|---|---|
| x2 | p | x2 | p | ||
| Overall Model (df = 3) | 108 | <0.001 | 78 | <0.001 | N/A |
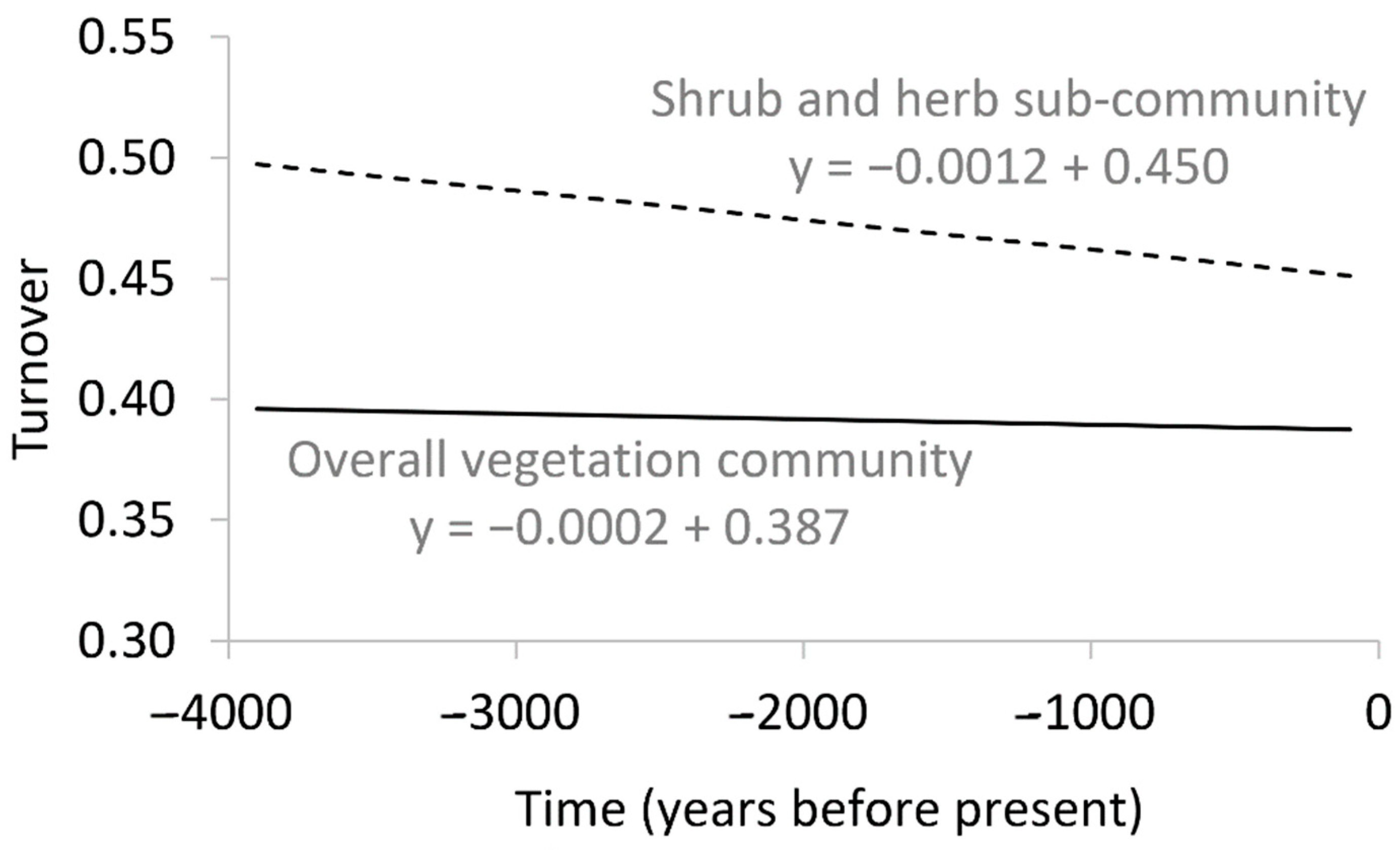
| Time (df = 1) | 1 | 0.313 | 21 | <0.001 | Not significant for overall vegetation community. Negative in SAH sub-community: rate of within-site turnover lower in more recent times compared to historically. |
| Time*Latitude (df = 1) | 14 | <0.001 | 21 | <0.001 | Positive in both models: general negative effect of time on rate of turnover is greater for sites at higher latitudes (steeper negative relationship) and weaker at lower latitudes (shallower negative relationship). |
| Time*Altitude (df = 1) | 30 | <0.001 | 10 | 0.002 | Negative in both models: general negative effect of time on rate of turnover was greater for sites at lower altitudes (steeper negative relationship) and weaker for sites at higher altitudes (shallower negative relationship). At very high altitudes, the relationship became positive in some cases. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goodenough, A.E.; Webb, J.C. Understanding Spatiotemporal Variation in Richness and Rate of Within-Site Turnover for Vegetation Communities in Western Eurasia over the Last 4000 Years. Diversity 2022, 14, 1096. https://doi.org/10.3390/d14121096
Goodenough AE, Webb JC. Understanding Spatiotemporal Variation in Richness and Rate of Within-Site Turnover for Vegetation Communities in Western Eurasia over the Last 4000 Years. Diversity. 2022; 14(12):1096. https://doi.org/10.3390/d14121096
Chicago/Turabian StyleGoodenough, Anne E., and Julia C. Webb. 2022. "Understanding Spatiotemporal Variation in Richness and Rate of Within-Site Turnover for Vegetation Communities in Western Eurasia over the Last 4000 Years" Diversity 14, no. 12: 1096. https://doi.org/10.3390/d14121096
APA StyleGoodenough, A. E., & Webb, J. C. (2022). Understanding Spatiotemporal Variation in Richness and Rate of Within-Site Turnover for Vegetation Communities in Western Eurasia over the Last 4000 Years. Diversity, 14(12), 1096. https://doi.org/10.3390/d14121096