
Amphiboreality and Distribution of Snailfishes (Cottiformes: Liparidae) in the Arctic and the North Atlantic


Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Overview of the Family Liparidae
3.2. Snailfishes of the Genus Liparis
3.2.1. Liparis of the Boreal Atlantic and the Arctic
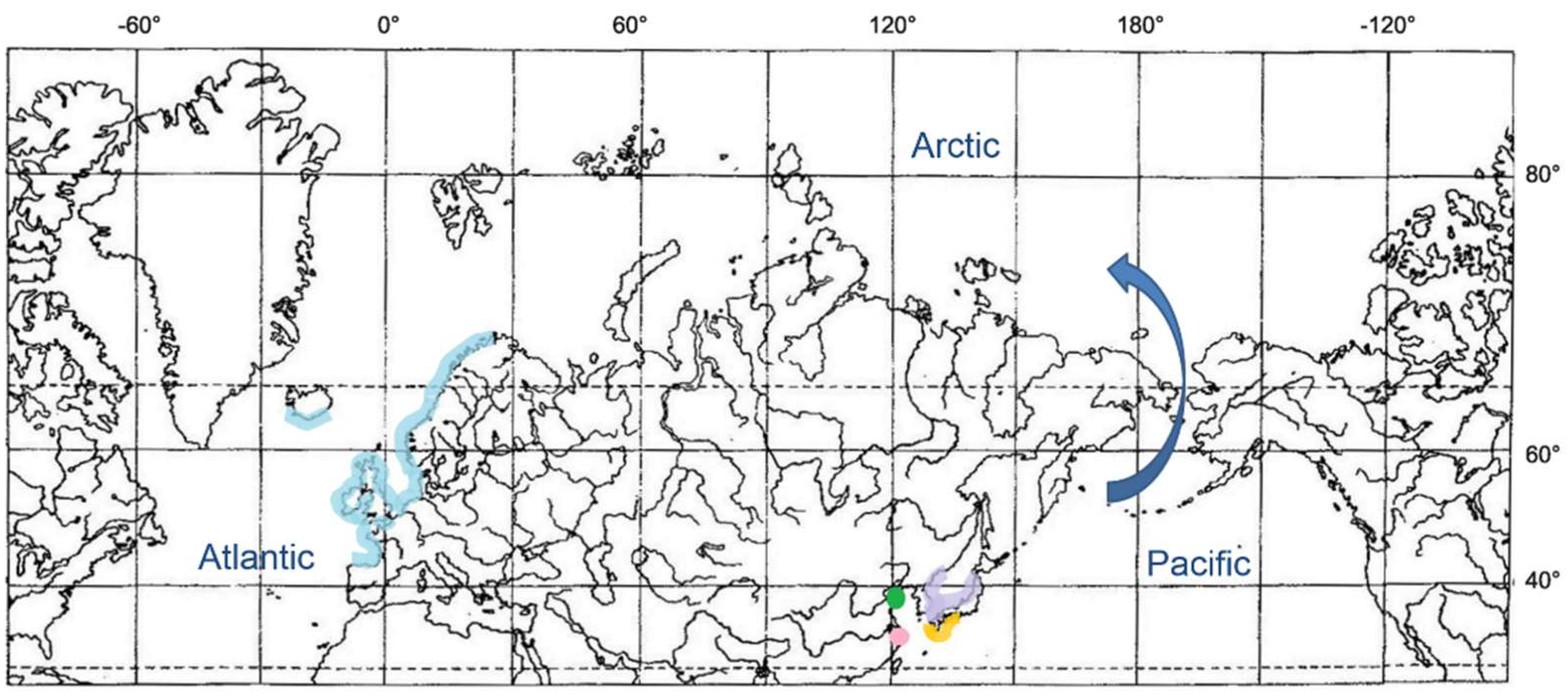
3.2.2. Groups of Liparis with Amphiboreal and Boreal–Arctic Distribution
3.2.3. Amphiboreal Concept and Speciation of Liparis in the Arctic and North Atlantic
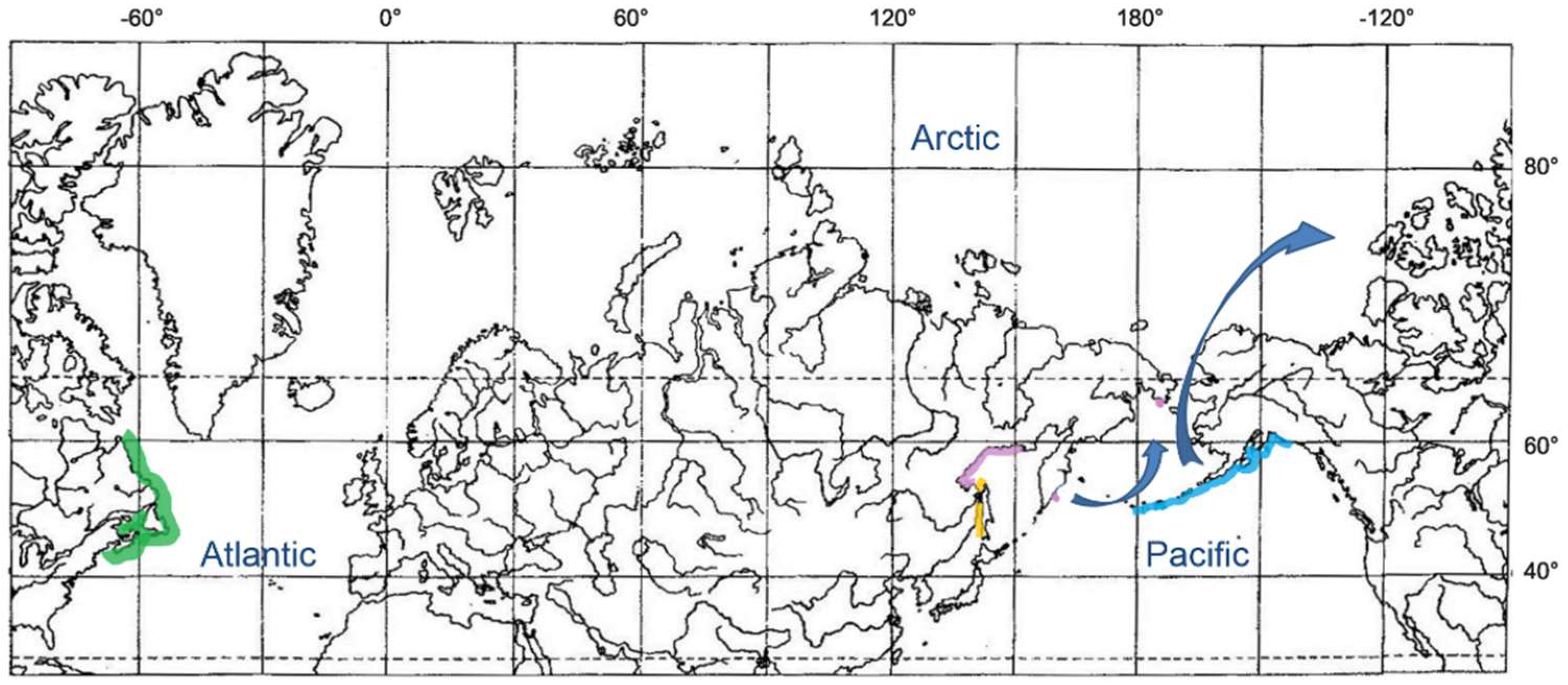
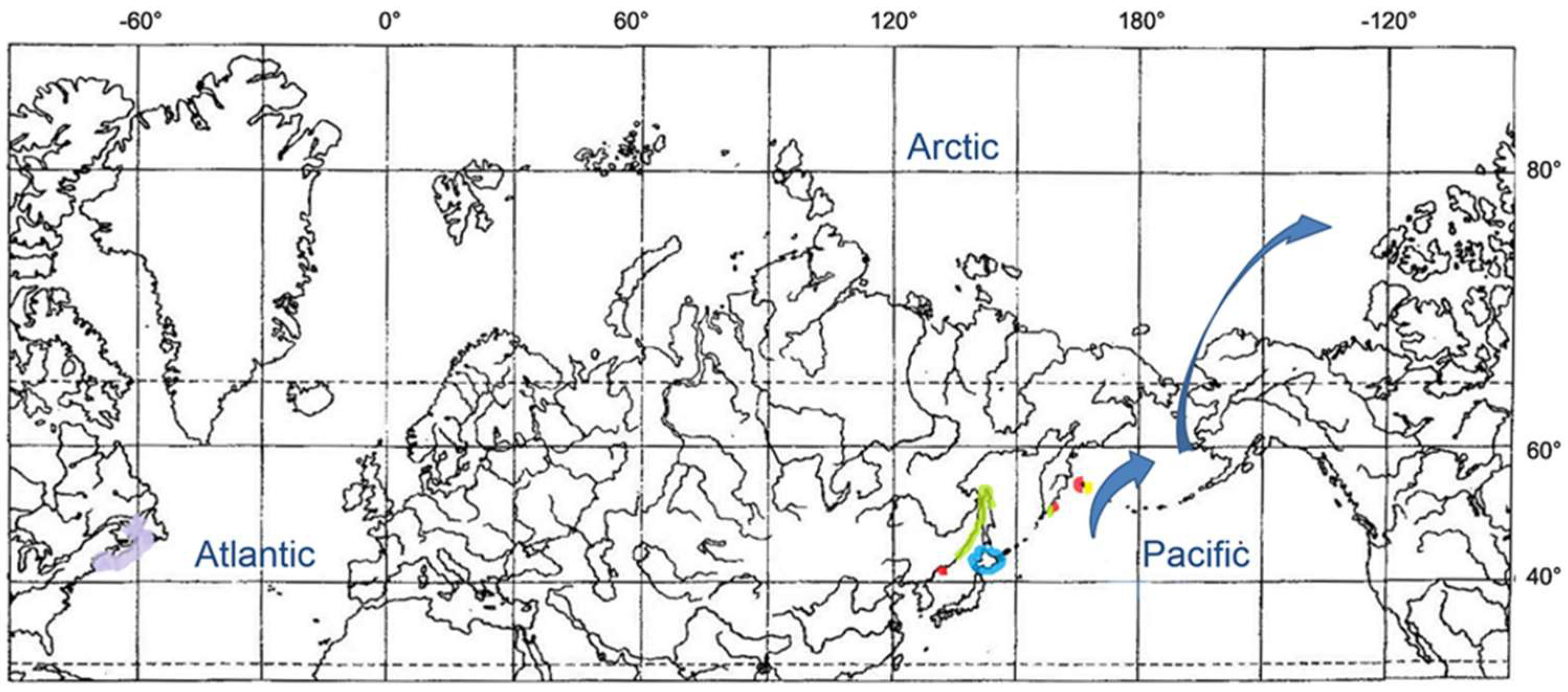
3.3. Snailfishes of the Arctic—Genus Careproctus
3.3.1. Careproctus of the Arctic and North Atlantic
3.3.2. Amphiboreal Concept and Speciation of Sea Tadpoles in the Arctic and North Atlantic
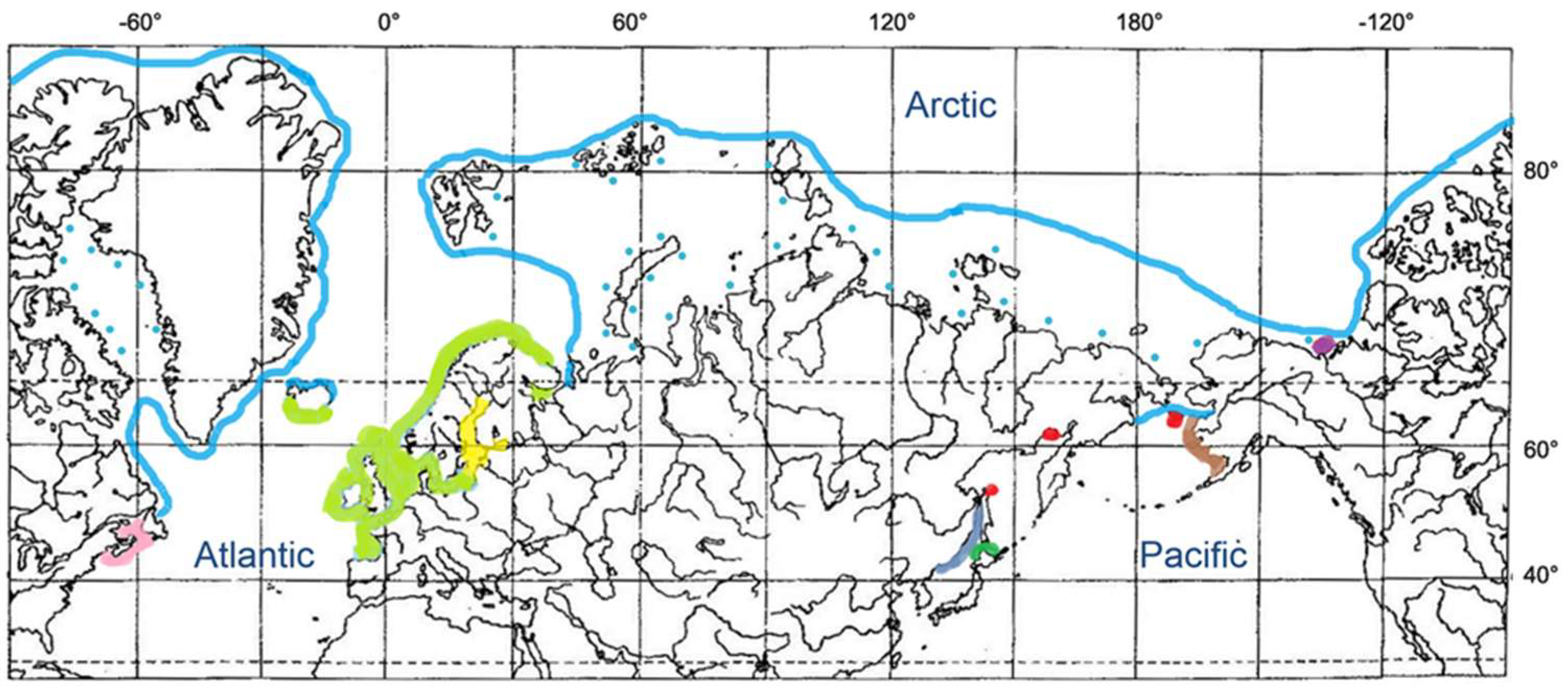
3.4. Other Liparids of the Arctic and Atlantic
4. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darwin, C.R. Chapter XI. Dispersal during the Glacial Period. In On the Origin of Species by Means of Natural Selection, or the Preservation of Favored Races in the Struggle for Life; John Murray: London, UK, 1859; pp. 365–382. Available online: https://www.biodiversitylibrary.org/bibliography/82303 (accessed on 15 August 2022).
- Berg, L.S. On the amphiboreal (discontinuous) distribution of marine fauna in the Northern Hemisphere. Izv. Geogr. Obs. (Proc. Geogr. Soc.) 1934, 16, 69–78. (In Russian) [Google Scholar]
- Andriyashev, A.P. Discontinuous distribution of marine life in the Northern Hemisphere. Prir. (Mosc.) 1944, 1, 44–52. (In Russian) [Google Scholar]
- Svetovidov, A.N. Fauna of the USSR, Fishes, Vol. II, No. 4, Gadiformes; Izdatelstvo AN USSR: Moscow, Russia; Leningrad, Russia, 1948; pp. 1–221, English transl.: Israel Program for Scientific Translations: Jerusalem, 1962. (In Russian) [Google Scholar]
- Andriashev, A.P. Some Issues of Amphiboreal Distribution with Notes on Possible Origin of Navaga and Low-Vertebrate Herring. In Materials on Complex Investigation of the White Sea; USSR Academy of Sciences Press: Moscow, Russia; Leningrad, Russia, 1957; Volume 1, pp. 117–125. (In Russian) [Google Scholar]
- Guryanova, E.F. Features of the fauna of the Arctic Ocean and their importance for understanding the history of its formation. In Arctic Ocean and Its Coast in the Cenozoic; Tolmachev, A., Ed.; Gidrometeoizdat Press: Leningrad, Russia, 1970; pp. 126–161. (In Russian) [Google Scholar]
- Andriashev, A.P. Possible pathways of Paraliparis (Pisces: Liparididae) and some other North Pacific secondarily deep-sea fishes into North Atlantic and Arctic depths. Polar Biol. 1991, 11, 213–218. [Google Scholar] [CrossRef]
- Carr, S.M.; Marshall, H.D. Phylogeographic analysis of complete mtDNA genomes from Walleye Pollock (Gadus chalcogrammus Pallas, 1811) shows an ancient origin of genetic biodiversity. DNA Seq. 2008, 19, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Makhrov, A.A.; Layus, D.L. Postglacial colonization of the North European seas by Pacific fishes and lamprey. Sib. Ekol. Zhurnal (Contemp. Probl. Ecol.) 2018, 3, 265–279, (In Russian with English summary). [Google Scholar] [CrossRef]
- Andriashev, A.P. An Outline of the Zoogeography and Origin of the Fish Fauna of the Bering Sea and Neighboring Waters; Leningrad State University Press: Leningrad, Russia, 1939; pp. 1–188, English transl.: Northwest and Alaska Fish Cent., Seattle, WA, USA. (In Russian) [Google Scholar]
- Berg, L.S. On the reasons for the similarity of the fauna of the Northern parts of the Atlantic and Pacific oceans. Izv. Akad. Nauk SSSR (Proc. USSR Acad. Sci.) 1918, 16, 1835–1842. (In Russian) [Google Scholar]
- Berg, L.S. Climate and life. Chapter 7. In On the Amphiboreal Distribution of Marine Fauna in the Northern Hemisphere, 2nd ed.; OGIZ: Moscow, Russia, 1947; pp. 116–127. [Google Scholar]
- Nesis, K.N. The routes and the periods of formation of the interrupted area of distribution of amphiboreal species of marine bottom animals. Okeanologiya (Oceanology) 1961, 1, 893–902. [Google Scholar] [CrossRef]
- Vinogradov, L.G.; Neiman, A.A. Some questions of the formation of the marine benthic fauna of the northern part of the Pacific Ocean. Tr. VNIRO 1972, 47, 11–22. (In Russian) [Google Scholar]
- Dyakonov, A.M. Interrelations between Arctic and Pacific faunas on the example of zoogeographical analysis of echinoderms. Zhurnal Obs. Biol. (J. Gen. Biol.) 1945, 6, 125–155. (In Russian) [Google Scholar]
- Briggs, J.C. Marine Zoogeography; McGraw Hill Book Company: New York, NY, USA, 1974; pp. 1–475. [Google Scholar] [CrossRef]
- Guryanova, E.F. To the zoogeographic zoning of cold and temperate waters of the northern hemisphere. In Hydrobiology and Biogeography of the Shelves of Cold and Temperate Waters of the World Ocean; Golikov, A., Ed.; USSR Academy of Sciences Press: Leningrad, Russia, 1974; pp. 11–12. (In Russian) [Google Scholar]
- Kafanov, A.I. Bivalve mollusks of the family Cardiidae of cold and temperate waters of the Northern Hemisphere, subfamily Clinocardiidae Kafanov. Ph.D. Thesis, Zoological Institute of the Russian Academy of Sciences, Leningrad, Russia, 1976; 228p. (In Russian). [Google Scholar]
- Kafanov, A.I. Cenozoic history of the malacofauna of the North Pacific shelf. In Marine Biogeography; Kusakin, O., Ed.; Nauka: Moscow, Russia; pp. 134–176. (In Russian)
- Neelov, A.V. Seismosensory System and Classification of the Sculpins (Cottidae: Myoxocephalinae, Artediellinae); Nauka: Leningrad, Russia, 1979; pp. 1–208. (In Russian) [Google Scholar]
- Kherman, I. Paleo-oceanology of the Arctic Basin in the Late Cenozoic and its relationship with global climates. Okeanologiya (Oceanology) 1983, 23, 112–119. (In Russian) [Google Scholar]
- Golikov, A.N. Stages and boundaries of the Cenozoic. Analysis of the course of geological time and the development of the fauna of marine gastropods. Tr. ZIN AN SSSR (Proc. Zool. Inst. Russ. Acad. Sci.) 1985, 130, 31–51. (In Russian) [Google Scholar]
- Briggs, J.C. Developments in paleonthology and stratigraphy. In Biogeoraphy and Plate Tectonics; Elsevier: Amsterdam, The Netherlands; Oxford, UK; New York, NY, USA; Tokyo, Japan, 1987; Volume 10, pp. 1–204. [Google Scholar]
- Gladenkov, A.Y.; Oleinik, A.E.; Marincovich, L., Jr.; Barinov, K.B. A recened age for the earliest opening of Bering Strait. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 183, 321–328. [Google Scholar] [CrossRef]
- Svendsen, J.I.; Alexanderson, H.; Astakhov, V.I.; Demidov, I.; Dowdeswell, J.A.; Funder, S.; Gataullin, V.; Henriksen, M.; Hjort, C.; Houmark-Nielsen, M.; et al. Late Quaternary ice sheet history of northern Eurasia. Quat. Sci. Rev. 2004, 23, 1229–1271. [Google Scholar] [CrossRef]
- Hughes, P.D.; Gibbard, P.L. Global glacier dynamics during 100 ka Pleistocene glacial cycles. Quat. Res. 2018, 90, 222–243. [Google Scholar] [CrossRef] [Green Version]
- Elias, S.A.; Short, S.K.; Nelson, C.H.; Birks, H.H. Life and times of the Bering Land Bridge. Nature 1996, 382, 60–63. [Google Scholar] [CrossRef]
- Elias, S.A.; Crocker, B. The Bering Land Bridge: A moisture barrier to the dispersal of steppe–tundra biota? Quat. Sci. Rev. 2008, 27, 2473–2483. [Google Scholar] [CrossRef]
- Jakobsson, M.; Pearce, C.; Cronin, T.M.; Backman, J.; Anderson, L.G.; Barrientos, N.; Björk, G.; Coxall, H.; de Boer, A.; Mayer, L.A.; et al. Postglacial flooding of the Bering Land Bridge dated to 11 cal ka BP based on new geophysical and sediment records. Clim. Past 2017, 13, 991–1005. [Google Scholar] [CrossRef] [Green Version]
- Lennox, B.; Spooner, I.; Jull, T.; Patterson, W.P. Postglacial climate change and its effect on a shallow dimictic lake in Nova Scotia, Canada. J. Paleolimnol. 2010, 43, 15–27. [Google Scholar] [CrossRef]
- Salonen, J.S.; Seppä, H.; Väliranta, M.; Jones, V.J.; Self, A.; Heikkilä, M.; Kultti, S.; Yang, H. The Holocene thermal maximum and late-Holocene cooling in the tundra of NE European Russia. Quat. Res. 2011, 75, 501–511. [Google Scholar] [CrossRef]
- Nazarkin, M.V. Freshwater fishes from Late Quaternary sediments of the coast of the East Siberian Sea. Vopr. Ichthyol. 1992, 32, 48–56. (In Russian) [Google Scholar]
- Andriashev, A.P. Neue Angaben über die Systematik und geographische Verbreitung der zweihörnigen pazifischen Icelus Arten. Zoologische Jahrbücher. Abteilung für Systematik. Ökologie Und Geogr. Der Tiere 1937, 69, 253–276. [Google Scholar]
- Andriashev, A.P. On the species composition and distribution of sculpins of the genus Triglops Reinh. in the northern seas. Tr. Vsesoyuznogo Geogr. Obs. 1949, 1, 194–209. (In Russian) [Google Scholar]
- Chernova, N.V. Snailfishes (Liparididae) from the Eurasian Arctic; Kola Scientific Centre of the USSR Academy of Sciences: Apatyti, Russia, 1991; pp. 1–112. (In Russian) [Google Scholar]
- Chernova, N.V. Careproctus solidus sp. n., a new species of liparid fish (Scorpaeniformes, Liparidae) from the lower bathyal of the Polar Basin. In Land-Ocean Systems in the Siberian Arctic: Dynamics and History; Kassens, H., Bauch, H.A., Dmitrenko, I.A., Eicken, H., Hubberten, H.W., Melles, M., Thiede, J., Timokhov, L.A., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA; Barcelona, Spain; Hong Kong, China; Milan, Italy; Paris, France; Singapore; Tokyo, Japan, 1999; pp. 181–188. [Google Scholar] [CrossRef]
- Chernova, N.V. Review of Careproctus (Liparidae) of the North Atlantic and adjacent Arctic, including the generic type C. reinhardti, with rehabilitation of C. gelatinosus (Pallas) from Kamchatka. J. Ichthyol. 2005, 45 (Suppl. 1), 1–22. [Google Scholar]
- Chernova, N.V. New Species of Careproctus (Liparidae) from the Barents Sea and adjacent Waters. Vopr. Ichthyol. 2005, 45, 725–736. English translation: J. Ichthyol. 2005, 45, 689–699.(In Russian) [Google Scholar]
- Chernova, N.V. Systematics and phylogeny of the genus Liparis (Liparidae, Scorpaeniformes). J. Ichthyol. (Suppl.) 2008, 48, 831–852. [Google Scholar] [CrossRef]
- Chernova, N.V. New species of the genus Careproctus (Liparidae) from the Kara Sea with notes on spongiophilia, reproduvctive commensalism between fishes and sponges (Rossellidae). Vopr. Ichthyol. 2014, 54, 508–519. English translation: J. Ichthyol. 2014, 54, 501–512.(In Russian) [Google Scholar] [CrossRef]
- Chernova, N.V. New species of the genus Careproctus (Liparidae) from the Kara Sea and identification key for congeners of the North Atlantic and Arctic. J. Ichthyol. 2014, 54, 757–780. [Google Scholar] [CrossRef]
- Chernova, N.V.; Stein, D.L.; Andriashev, A.P. Family Liparidae Scopoli 1777—Snailfishes. Calif. Acad. Sci. Annot. Checkl. Fishes 2004, 31, 1–72. Available online: https://www.researchgate.net/publication/242550096_Family_Liparidae_Scopoli_1777_-_Snailfishes (accessed on 10 September 2022).
- Chernova, N.V.; Vedischeva, E.V.; Datskii, A.V. A new species of snailfishes (Liparidae) of the genus Careproctus from the northern slope of the Aleutin Basin (Bering Sea). J. Ichthyol. 2021, 61, 487–495. Available online: https://link.springer.com/article/10.1134/S0032945221040020 (accessed on 10 September 2022). [CrossRef]
- Andriashev, A.P.; Chernova, N.V. A new species of the genus Careproctus (Liparididae) from the four-kilometer depth of the Porcupine Seabight (Northeastern Atlantic). Vopr. Ikhtiologii 1988, 28, 1023–1026. English translation: J. Ichthyol. 1989, 1, 94–98.(In Russian) [Google Scholar]
- Andriyashev, A.P.; Chernova, N.V. Two new species of liparid fish (Liparidae, Scorpaeniformes) from the abyssal depths of the Eastern North Atlantic. Vopr. Ikhtiologii 1997, 37, 437–443. English translation: J. Ichthyol. 1997, 37, 479–484.(In Russian) [Google Scholar]
- Chernova, N.V. A review of the genus Psednos (Pisces, Liparidae) with description of ten new species from the north Atlantic and southwestern Indian Ocean. Bull. Mus. Comp. Zool. 2001, 155, 477–507. [Google Scholar]
- Chernova, N.V. New finding of deep-sea snailfish Paraliparis challengeri in the Northeastern Atlantic. Vopr. Ikhtiologii 2003, 43, 279–280. English translation: J. Ichthyol. 2003, 43, 215–216.(In Russian) [Google Scholar]
- Chernova, N.V.; Møller, P.R. A new snailfish, Paraliparis nigellus sp. nov. (Scorpaeniformes: Liparidae), from the northern Mid-Atlantic Ridge with notes on occurrence of Psednos in the area. Mar. Biol. Res. 2008, 4, 369–375. [Google Scholar] [CrossRef]
- Chernova, N.V.; Geistdoerfer, P. Redescription of snailfish Careproctus hyaleius (Scorpaeniformes, Liparidae) from the area of hydrothermal vents in the Equatorial Part of the East Pacific Rise. Voprsi Ikhthyologii 2003, 43, 149–155. English translation: J. Ichthyol. 2003, 43, 139–145.(In Russian) [Google Scholar]
- Gilbert, C.H. The ichthyological collections of the steamer Albatross during the years 1890 and 1891. U.S. Comm. Fish Fish. Rep. Comm. 1896, 19, 393–476. [Google Scholar]
- Gilbert, C.H.; Burke, C.V. New cyclogasterid fishes from Japan. Proc. United States Natl. Mus. 1912, 42, 351–380. [Google Scholar] [CrossRef]
- Gilbert, C.H.; Burke, C.V. Fishes from Bering Sea and Kamchatka. Bull. Bur. Fish. 1912, 30, 31–96. [Google Scholar]
- Burke, C.V. Revision of the fishes of the family Liparidae. Bull. US Natl. Mus. 1930, 150, 1–204. [Google Scholar] [CrossRef]
- Schmidt, P.Y. Fishes of the Sea of Okhotsk; USSR Academy of Sciences Press: Moscow, Russia; Leningrad, Russia, 1950; pp. 1–383, English translation: Israel Program for Scientific Translation, 1965. (In Russian) [Google Scholar]
- Stein, D.L. A review of the deepwater Liparidae (Pisces) from the coast of Oregon and adjacent waters. Occas. Pap. Calif. Acad. Sci. 1978, 127, 1–55. [Google Scholar]
- Kido, K. Phylogeny of the family Liparididae, with the taxonomy of the species found around Japan. Mem. Fac. Fish. Hokkaido Univ. 1988, 35, 125–256. [Google Scholar]
- Lindberg, G.U.; Krasyukova, Z.V. Fishes of the Sea of Japan and the Adjacent Areas of the Okhotsk Sea and Yellow Sea. Part 5. Teleostomi, Osteichthys, Actinopterygii. XXX. Scorpaeniformes (CLXXVI. Fam. Scorpaenidae—CXCIV. Fam. Liparididae); Nauka: Leningrad, Russia, 1987; pp. 1–526. (In Russian) [Google Scholar]
- Orr, J.W.; Maslenikov, K.P. Two new variegated snailfishes of the genus Careproctus (Teleostei: Scorpaeniformes: Liparidae) from the Aleutian Islands, Alaska. Copeia 2007, 3, 699–710. [Google Scholar] [CrossRef]
- Sakurai, H.; Shinohara, G. Careproctus rotundifrons, a new snailfish (Scorpaeniformes: Liparidae) from Japan. Bull. Natl. Mus. Nat. Sci. (Ser. A) Suppl. 2008, 2, 39–45. [Google Scholar]
- Andriashev, A.P. Liparid fishes of the Southern Ocean and Adjacent Waters. Results of the Russian Antarctic Expeditions, Volume 9. Explorations of the Fauna of the Seas, Volume 53(61); Zoological Institute of the Russian Academy of Sciences: Saint Petersburg, Russia, 2003; pp. 1–478, (In Russian with English diagnoses). [Google Scholar]
- Stein, D.L.; Chernova, N.V.; Andriashev, A.P. Snailfishes (Pisces: Liparidae) of Australia, including descriptions of thirty new species. Rec. Aust. Mus. 2001, 53, 341–406. [Google Scholar] [CrossRef] [Green Version]
- Andriashev, A.P.; Chernova, N.V. Annotated list of fishlike vertebrates and fish of the Arctic seas and adjacent waters. J. Ichthyol. 1995, 35, 81–123. [Google Scholar]
- Chernova, N.V. Distribution patterns and chorological analysis of fish fauna of the Arctic Region. J. Ichthyol. 2011, 51, 825–924. [Google Scholar] [CrossRef]
- Smith, W.L.; Busby, M.S. Phylogeny and taxonomy of sculpins, sandfishes, and snailfishes (Perciformes: Cottoidei) with comments on the phylogenetic significance of their early-life-history specializations. Mol. Phylogenetics Evol. 2014, 79, 332–352. [Google Scholar] [CrossRef]
- Orr, J.W.; Spies, I.B.; Stevenson, D.E.; Longo, G.C.; Kai, Y.; Ghods, S.; Hollowed, M. Molecular phylogenetics of snailfishes (Cottiformes: Liparidae) based on MtDNA and RADseq genomic analyses, with comments on selected morphological characters. Zootaxa 2019, 4642, 1–79. [Google Scholar] [CrossRef]
- Andriashev, A.P. A new fish of the snailfish family (Pisces, Liparidae) found at a depth of more than 7 kilometers. Tr. Inst. Okeanol. Akad. Nauk SSSR (Proc. Inst. Oceanol. Russ. Acad. Sci.) 1955, 12, 340–344. (In Russian) [Google Scholar]
- Andriashev, A.P. A new ultra-abyssal fish, Notoliparis kurchatovi gen. et sp. n. (Liparidae) from the South-Orkney Trench (Antarctic). Tr. Inst. Okeanol. Akad. Nauk SSSR (Proc. Inst. Oceanol. Russ. Acad. Sci.) 1975, 103, 313–319. (In Russian) [Google Scholar]
- Andriashev, A.P. On the third species of the ultra-abyssal genus Notoliparis Andr. (Pisces. Liparidae), from the deepwaters of the Macquarie Trench, with some notes on zoogeographic and evolutionary significance of this discovery. Tr. Inst. Okeanol. Akad. Nauk SSSR (Proc. Inst. Oceanol. Russ. Acad. Sci.) 1978, 112, 152–161. (In Russian) [Google Scholar]
- Abe, T. New, rare or uncommon fishes from Japanese waters. I. Liparis franzi, new name. Jpn. J. Ichthyol. 1950, 1, 135–139. [Google Scholar]
- Smith, J.L.B. A new liparine fish from the Red Sea. J. Nat. Hist. 1968, 2, 105–109. [Google Scholar] [CrossRef]
- Fadeev, N.S. Manual on Biology and Fishery of Fish of the Northern Part of the Pacific Ocean; TINRO-Tsentr Press: Vladivostok, Russia, 2005; pp. 1–366. (In Russian) [Google Scholar]
- Tsynovskiy, V.D.; Mel’nikov, I.A. Records of Liparis koefoedi (Liparidae, Osteichthyes) in the Central Arctic Basin. In Biology of the Central Arctic Basin; Vinogradov, M., Mel’nikov, I., Eds.; Nauka: Moscow, Russia, 1980; pp. 211–214. (In Russian) [Google Scholar]
- Andriashev, A.P. Review of the snailfish genus Paraliparis (Scorpaeniformes: Liparididae) of the Southern Ocean. Theses Zool. 1986, 7, 1–204. [Google Scholar]
- Andriyashev, A.P. Fishes of the Northern Seas of the USSR; Zoological Institute of the USSR Academy of Sciences Publishers: Leningrad, Russia, 1954; pp. 1–567, English Transl.: Israel Program of Scientific Translations, 1964; Volume 836, pp. 1–617. (In Russian) [Google Scholar]
- Tsynovskiy, V.D. To the ichthyofauna of deep-sea basins of the Central Arctic Basin. In Biology of the Central Arctic Basin; Vinogradov, M., Mel’nikov, I., Eds.; Nauka: Moscow, Russia, 1980; pp. 214–218. (In Russian) [Google Scholar]
- Stein, D.L.; Able, K.W. Liparididae. In Fishes of the NE Atlantic and the Mediterranean; Whitehead, P., Bauchot, M., Hureau, J., Eds.; UNESCO: Paris, France, 1986; Volume 3, pp. 1275–1278. [Google Scholar]
- Møller, P.R.; Nielsen, J.G.; Knudsen, S.W.; Poulsen, J.Y.; Sünksen, K.; Jørgensen, O.A. A checklist of the fish fauna of Greenland waters. Zootaxa 2010, 2378, 1–84. [Google Scholar]
- Nozères, C. Poissons marins du Saint-Laurent: Un Photoguide/Marine Fishes of the St. Lawrence: A Photoguide, 2017. Version 2017-03-05. Available online: https://ogsl.ca/en/species-identification-guide-marine-fish (accessed on 13 September 2022).
- Krøyer, H.N. Nogle Bidrag til Nordisk ichthyologi. Nat. Tidsskr. (Kjøbenhavn) (Ser. 3) 1862, 1, 233–310. [Google Scholar]
- Burke, C.V. Note on the Cyclogasteridae. Ann. Magaz. Nat. Hist. (Ser. 8) 1912, 9, 507–513. [Google Scholar] [CrossRef]
- Orr, J.W.; Kai, Y.; Nakabo, T. Snailfishes of the Careproctus rastrinus complex (Liparidae): Redescriptions of seven species in the North Pacific Ocean region, with the description of a new species from the Beaufort Sea. Zootaxa 2015, 4018, 301–348. [Google Scholar] [CrossRef] [Green Version]
- Orr, J.W. A new snailfish of the genus Careproctus (Cottiformes: Liparidae) from the Beaufort Sea. Copeia 2020, 108, 815–819. [Google Scholar] [CrossRef]
- Knudsen, S.W.; Moller, P.R. Careproctus kidoi, a new Arctic species of snailfish (Teleostei: Liparidae) from Baffin Bay. Ichthyol. Res. 2008, 55, 175–182. [Google Scholar] [CrossRef]
- Andriyashev, A.P.; Chernova, N.V. Three new snailfishes (Scorpaeniformes: Liparidae) from bathyal depths of the Arctic. Proc. Zool. Inst. Russ. Acad. Sci. 2010, 314, 365–380, (In Russian, with English summary). [Google Scholar] [CrossRef]
- Günther, A. Report on the Deep-Sea Fishes Collected by H. M. S. Challenger during the Years 1873–76; Report on the Scientific Results of the Voyage of H.M.S. Challenger; Printed for Her Majesty’s Stationery Office: London, UK, 1887; Volume 22, pp. 1–268. [Google Scholar] [CrossRef]
- Collett, R. Fiske indsamlede under “Michael Sars”s togeter i Nordhavet 1900–1902. In Report on Norwegian Fishery and Marine-Investigations; J. Grieg: Bergen, Norway, 1905; Volume 2, pp. 1–147. [Google Scholar]
- Johnsen, S. Ichthyologiske Notiser, I. Bergens Museum Aarbook, 1918–1919. Nat. Raekke 1921, 2, 1–94. [Google Scholar]
- Joensen, J.S.; Tåning, A.V. Marine and Freshwater fishes. In The Zoology of the Faroes; Spærck, R., Tuxen, S., Eds.; Høst: Copenhagen, Danmark, 1970; Volume 3, pp. 1–241. [Google Scholar]
- Blacker, R.W. English observations on rare fish in 1975. Ann. Boil. (Cph.) 1977, 32, 184–185. [Google Scholar]
- Iglésias, S.P. Piscibus Marinis — Guide des Poisons Marins, Europe et Eaux Adjacentes (Une Classification Naturelle Basée sur des Specimens de Collection, des Barcodes ADN et des Photos Standardisées). Version provisioire 11. 12 October 2020; 421p. Available online: https://www.researchgate.net/profile/Samuel_Iglesias(accessed on 11 September 2022).
- Collett, R. Fiske fra Nordhavs-Expeditionens sidste Togt, Sommeren 1878. Forh. I Vidensk.-Selsk. I Christiania 1879, 14, 1–106. [Google Scholar]
- Lütken, C.F. The Ichthyological Results. Dan. Ingolf Exped. 1898, 2, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Nizovtsev, G.P.; Ponomarenko, V.P.; Shevelev, M.S. Fish from the great depths of the Norwegian Sea. Vopr. Ichthyol. 1976, 16, 1115. [Google Scholar]
- Magnussen, E. Demersal fish assemblages of Faroe Bank: Species composition, distribution, biomass spectrum and diversity. Mar. Ecol. Prog. Ser. 2002, 238, 211–225. [Google Scholar] [CrossRef] [Green Version]
- Jensen, A.S. On some deep-sea fishes caught by the Danish Gothhab Expedition 1828, not previously known from West Greenland. Vidensk. Medd. Fra Dan. Nat. Foren. I Kbh. 1950, 112, 243–448. [Google Scholar]
- Gorbunov, G.P. The bottom animals of the Novosibirsk shallow water and the central part of the Arctic Ocean. In Proceedings of the Drifting Expedition of the Glavsevmorput on the Icebreaking Steamboat “G. Sedov” 1937–1940; Izd. Glavsevmorputi: Moscow, Russia; Leningrad, Russia, 1946; 3, pp. 30–138. (In Russian). [Google Scholar]
- Chernova, N.V.; Stein, D.L. Ten new species of Psednos (Pisces, Scorpaeniformes: Liparidae) from the Pacific and North Atlantic Oceans. Copeia 2002, 3, 755–778. [Google Scholar] [CrossRef]
- Knudsen, S.W.; Møller, P.R.; Gravlund, P. Phylogeny of the snailfishes (Teleostei: Liparidae) based on molecular and morphological data. Mol. Phylogenetics Evol. 2007, 44, 649–666. [Google Scholar] [CrossRef]
- Able, K.W.; McAllister, D.E. Revision of the snailfish genus Liparis from Arctic Canada. Can. Bull. Fish. Aquat. Sci. 1980, 208, 1–52. [Google Scholar]
- Mecklenburg, C.W.; Lynghammar, A.; Johannesen, E.; Byrkjedal, I.; Christiansen, J.S.; Dolgov, A.V.; Karamushko, O.V.; Mecklenburg, T.A.; Møller, P.R.; Steinke, D.; et al. Marine fishes of the Arctic Region; CAFF Monitoring Series Report 28; Conservation of Arctic Flora and Fauna: Akureyri, Iceland, 2018; pp. 1–454. [Google Scholar]
- Able, K.W. A new cyclopterid fish, Liparis inquilinus, associated with the sea scallop, Placopecten magellanicus, in the western North Atlantic, with notes on the Liparis liparis complex. Copeia 1973, 4, 787–794. [Google Scholar] [CrossRef]
- Able, K.W. Life history, ecology and behavior of two new liparis (Pisces: Cyclopteridae) from the western North Atlantic. Master’s Dissertation, Paper 1539616548. College of William and Mary, Virginia Institute of Marine Science, Gloucester Point, VA, USA, 1974. [Google Scholar] [CrossRef]
- Able, K.W.; Musick, J.A. Life history, ecology, and behavior of Liparis inquilinus (Pisces: Cyclopteridae) associated with the sea scallop, Placopecten magellanicus. Fish. Bull. 1976, 74, 409–421. [Google Scholar]
- Able, K.W. A new cyclopterid fish, Liparis coheni, from the western North Atlantic with notes on the life history. Copeia 1976, 3, 515–521. [Google Scholar] [CrossRef]
- Parr, F.E. A study of subspecies and racial variations in Liparis liparis Linnaeus and Liparis koefoedi sp. n. in Northern Europe and the European Arctic water. Bergen Mus. Ǻrbok Nat. Rekke 1932, 6, 1–54. [Google Scholar]
- Goode, G.B.; Bean, T.H. Description of a new species of Liparis (L. ranula) obtained by the United States Fish Commission off Halifax, Nova Scotia. Proc. U. S. Natl. Mus. 1880, 2, 46–48. [Google Scholar] [CrossRef]
- Andriashev, A.P. Paraliparis bipolaris sp. n. from the northeastern Atlantic, similar to the Antarctic P. kreffti (Liparidae, Scorpaeniformes). Vopr. Ikhtiologii 1997, 37, 261–264. English translation: J. Ichthyol. 1997, 37, 196–199.(In Russian) [Google Scholar]
- Cohen, D.M. The cyclopterid genus Paraliparis, a senior synonym of Gymnolycodes and Eutelichthys, with the description of a new species from the Gulf of Mexico. Copeia 1968, 2, 384–388. [Google Scholar] [CrossRef]
- Andriashev, A.P. The restoration of the validity of the forgotten species Amitra liparina Goode, 1881, with a description of a new species, similar to it, Paraliparis challengeri sp. n. (Scorpaeniformes, Liparidae), from Porcupine Seabight (Northeast Atlantic). Vopr. Ikhtiologii 1993, 33, 597–601. English translation: J. Ichthyol. 1994, 34, 1–7.(In Russian) [Google Scholar]
- Haedrich, R.L.; Merrett, N.R. Summary atlas of deep-living demersal fishes in the North Atlantic Basin. J. Nat. Hist. 1988, 22, 1325–1362. [Google Scholar] [CrossRef]
- Vaillant, L.L. Poissons. In Expéditions Sci. Du “Trav.” Et Du “Talisman” Pendant Les Années 1880, 1881, 1882, 1883; G. Masson: Paris, France, 1888; pp. 1–406. [Google Scholar]
- Matallanas, J. On the Mediterranean and some north-eastern Atlantic Liparidae (Pisces: Scorpaeniformes) with the restoration of Eutelichthys. J. Mar. Biol. U.K. 2000, 80, 935–939. [Google Scholar] [CrossRef]
- Merrett, N.R. A new species of the deep-sea fish genus Paraliparis Collett (Liparididae) from the eastern North Atlantic, with notes on its ecology. J. Fish Biol. 1983, 23, 429–439. [Google Scholar] [CrossRef]
- Hartel, K.E.; Kenaley, C.P.; Galbraith, J.K.; Sutton, T.T. Additional records of deep-sea fishes from off greater New England. Northeast. Nat. 2008, 15, 317–334. [Google Scholar] [CrossRef] [Green Version]
- Matallanas, J. Descripcion de una nueva especie de Liparidae: Paraliparis murieli n. sp. (Pisces, Scorpaeniformes). Invest. Pesq. 1984, 48, 563–567. [Google Scholar]
- Chernova, N.V. A new species of Snailfish (Liparidae) Paraliparis vaillanti sp.nova from the Northwestern Atlantic, with the redescription of rare species Paraliparis edwardsi Vaillant. J. Ichthyology 2004, 44, 135–139. [Google Scholar]
- Andriashev, A.P. Morphological evidence for the validity of the antitropical genus Psednos Barnard (Scorpaeniformes, Liparidae) with a description of a new species from the Eastern North Atlantic. U0 (Tokyo) 1992, 41, 1–18. [Google Scholar]
- Chernova, N.V.; Stein, D.L. A remarkable new species of Psednos (Teleostei: Liparidae) from the western North Atlantic Ocean. Fish. Bull. 2004, 102, 245–250. [Google Scholar]
- Tortonese, E. Un nuovo pesce Mediterraneo di profondità: Eutelichthys leptochirus, n. gen. e n. sp. (Fam. Eutelichthyidae, nov.). Ann. Mus. Civ. Stor. Nat. “Giacomo Doria” 1960, 71, 226–232. [Google Scholar]
- Iglésias, S.P.; Bariche, M.; Beau, F.; Bérenger, L.; Beucher, R.; Chabrolle, A.; Cottalorda, J.M.; Cousin, B.; Curd, A.; Danet, V.; et al. French ichthyological records for 2019. Cybium 2021, 45, 169–188. [Google Scholar]
- Tokranov, A.M. Size and age composition of liparid fishes (Liparidae) in the Pacific waters of southeastern Kamchatka and the northern Kuril Islands. Vopr. Ichthyol. 2000, 40, 347–352. (In Russian) [Google Scholar]
- Poltev, Y.N.; Nemchinova, I.A. On the feeding of cod Gadus morhua macrocephalus and its place in the trophic structure of communities in the Pacific waters of the northern Kuril Islands and the southeastern tip of Kamchatka in the late autumn of 1994. In Commercial and Biological Studies of fish in the Pacific Waters of the Kuril Islands and Adjacent Areas of Okhotsk and the Bering Seas in 1992–1998; Kotenev, B., Ed.; VNIRO Publishing House: Moscow, Russia, 2000; pp. 141–154. [Google Scholar]
- Andriashev, A.P.; Stein, D.L. Review of the snailfish genus Careproctus (Liparidae, Scorpaeniformes) in Antarctic and adjacent waters. Contrib. Sci. (Los Ang.) 1998, 470, 1–63. [Google Scholar] [CrossRef]
- Lovén, S. Till frågan om Ishafsfaunans fordna utsträckning öfver en del af Nordens fastland. Öfversigt Af Kongliga Vetensk.-Akad. Förhandlingar 1863, 8, 463–468. [Google Scholar]
- Saarnisto, M.; Grönlund, T.; Ekman, I. Lateglacial of Lake Onega—Contribution to the history of the eastern Baltic basin. Quat. Int. 1995, 27, 111–120. [Google Scholar] [CrossRef]
- Heinsalu, A.; Veski, S. The history of the Yoldia Sea in Northern Estonia: Palaeoenvironmental conditions and climatic oscillations. Geol. Q. (Warszawa) 2007, 51, 295–306. [Google Scholar]
- Rößler, D.; Moros, M.; Lemke, W. The Littorina transgression in the southwestern Baltic Sea: New insights based on proxy methods and radiocarbon dating of sediment cores. Boreas 2010, 40, 231–241. [Google Scholar] [CrossRef]
- Ekman, S. Zoogeography of the Sea; Sidgwick and Jackson Limited: London, UK, 1953; pp. 1–417. [Google Scholar] [CrossRef]
- Andriyashev, A.P. On the probability of trans-oceanic (non-Arctic) dispersal of secondary deep-water species of boreal Pacific origin to the depths of the North Atlantic and the Arctic (on the example of the family Liparidae). Zool. J. 1990, 69, 61–67. (In Russian) [Google Scholar]
- Prokofiev, A.M.; Iftime, A. A revision of the abandoned snailfish genus Menziesichthys with description of a new species (Teleostei, Scorpaeniformes, Liparidae). Trav. Du Muséum Natl. D’histoire Nat. “Grigore Antipa” 2020, 63, 93–115. [Google Scholar] [CrossRef]
- Blanc, P.-L. The opening of the Plio-Quaternary Gibraltar Strait: Assessing the size of a cataclysm. Geodin. Acta 2002, 15, 303–317. [Google Scholar] [CrossRef]
- Cornée, J.-J.; Münch, P.; Achalhi, M.; Merzeraud, G.; Azdimousa, A.; Quillévéré, F.; Melinte-Dobrinescu, M.; Chaix, C.; Moussa, A.B.; Lofi, J.; et al. The Messinian erosional surface and early Pliocene reflooding in the Alboran Sea: New insights from the Boudinar basin, Morocco. Sediment. Geol. 2016, 333, 115–129. [Google Scholar] [CrossRef]
- Laakkonen, H.; Hardman, M.; Strelkov, P.; Väinölä, R. Cycles of trans-Arctic dispersal and vicariance, and diversification of the amphi-boreal marine fauna. J. Evol. Biol. 2021, 34, 73–96. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
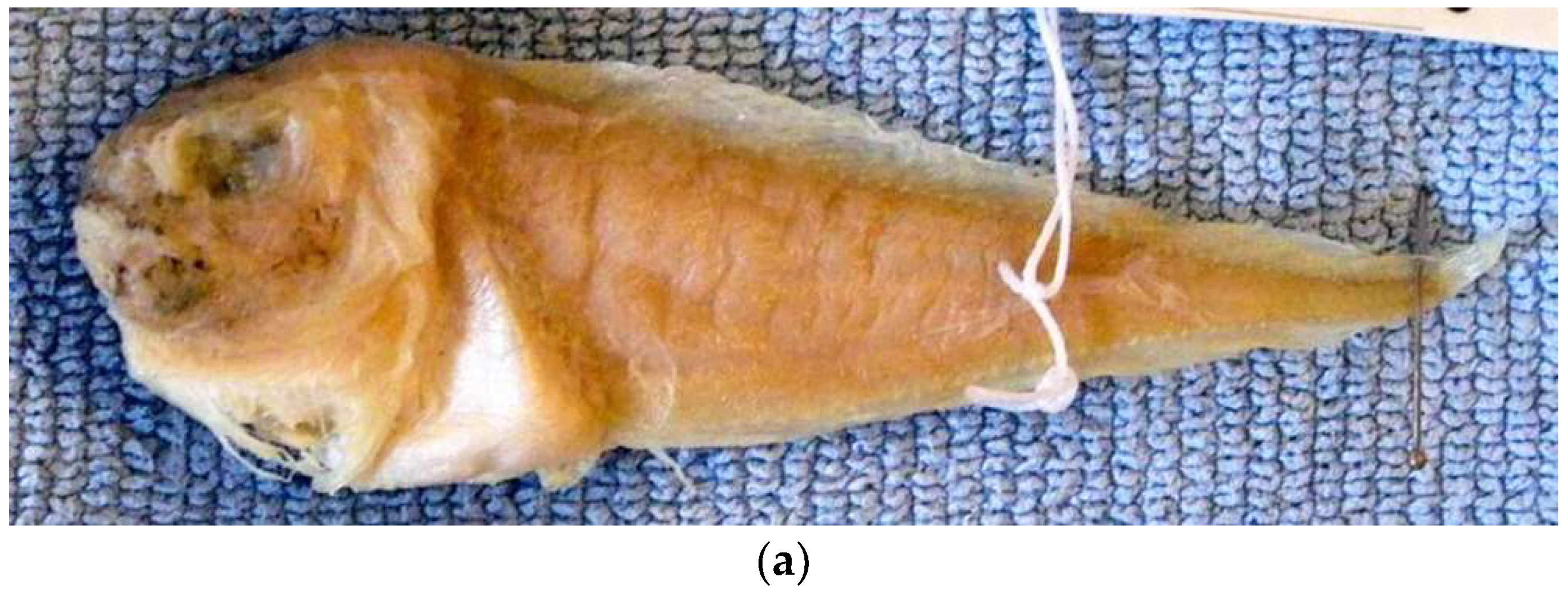{kind=link}
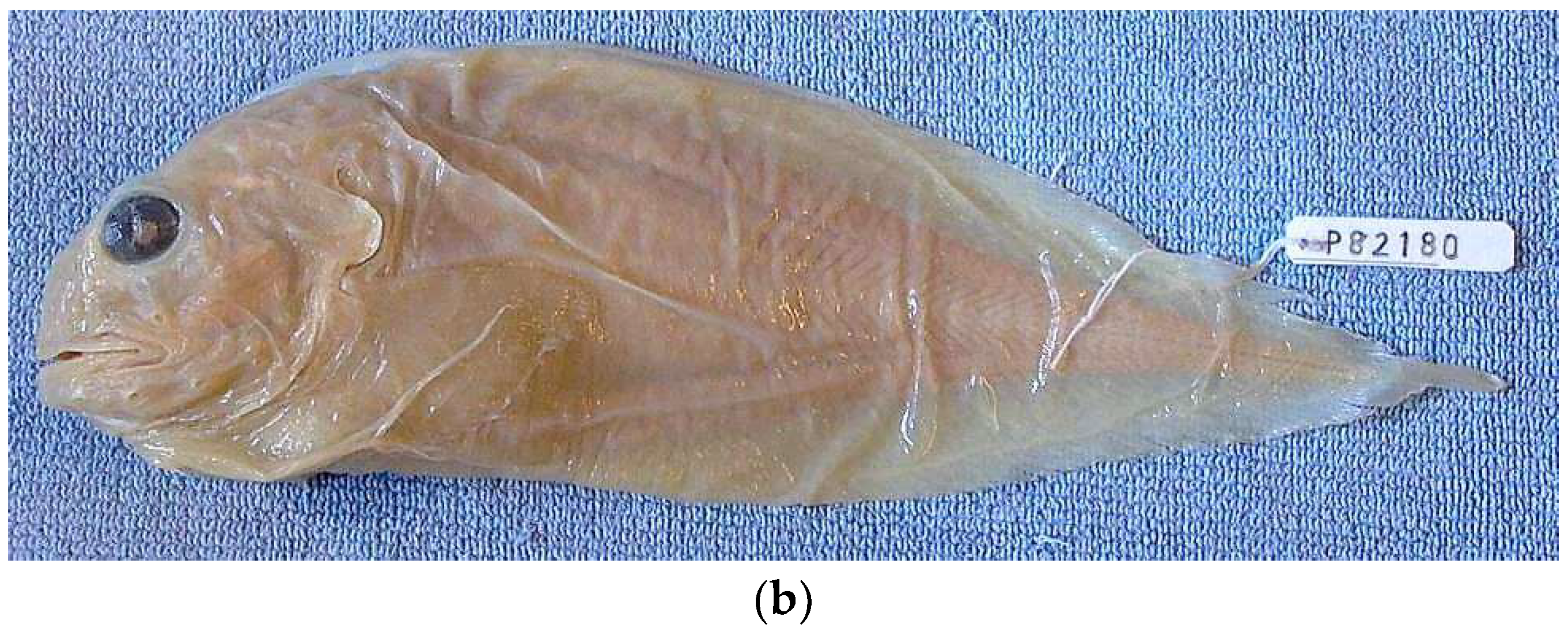{kind=link}
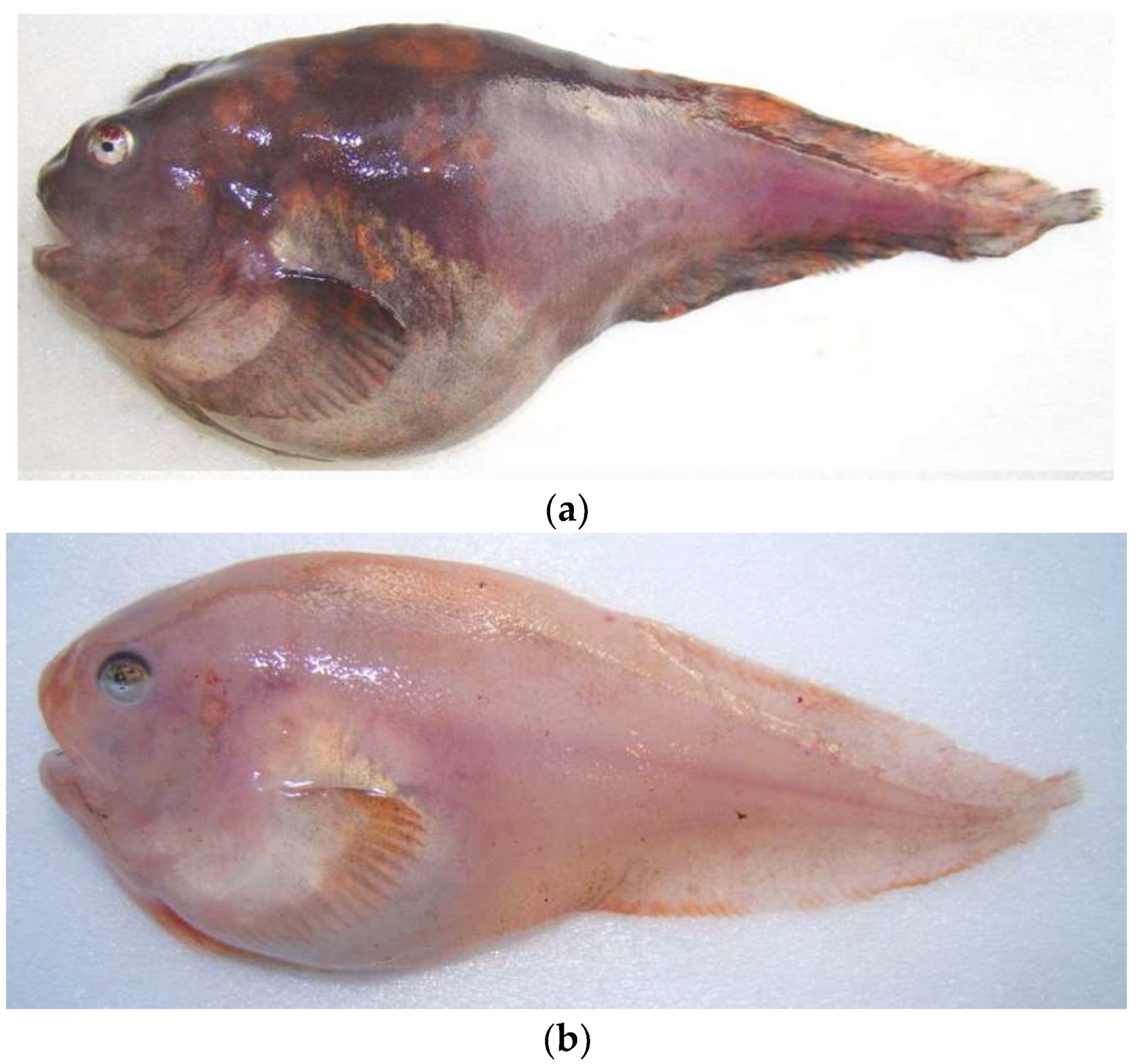{kind=link}
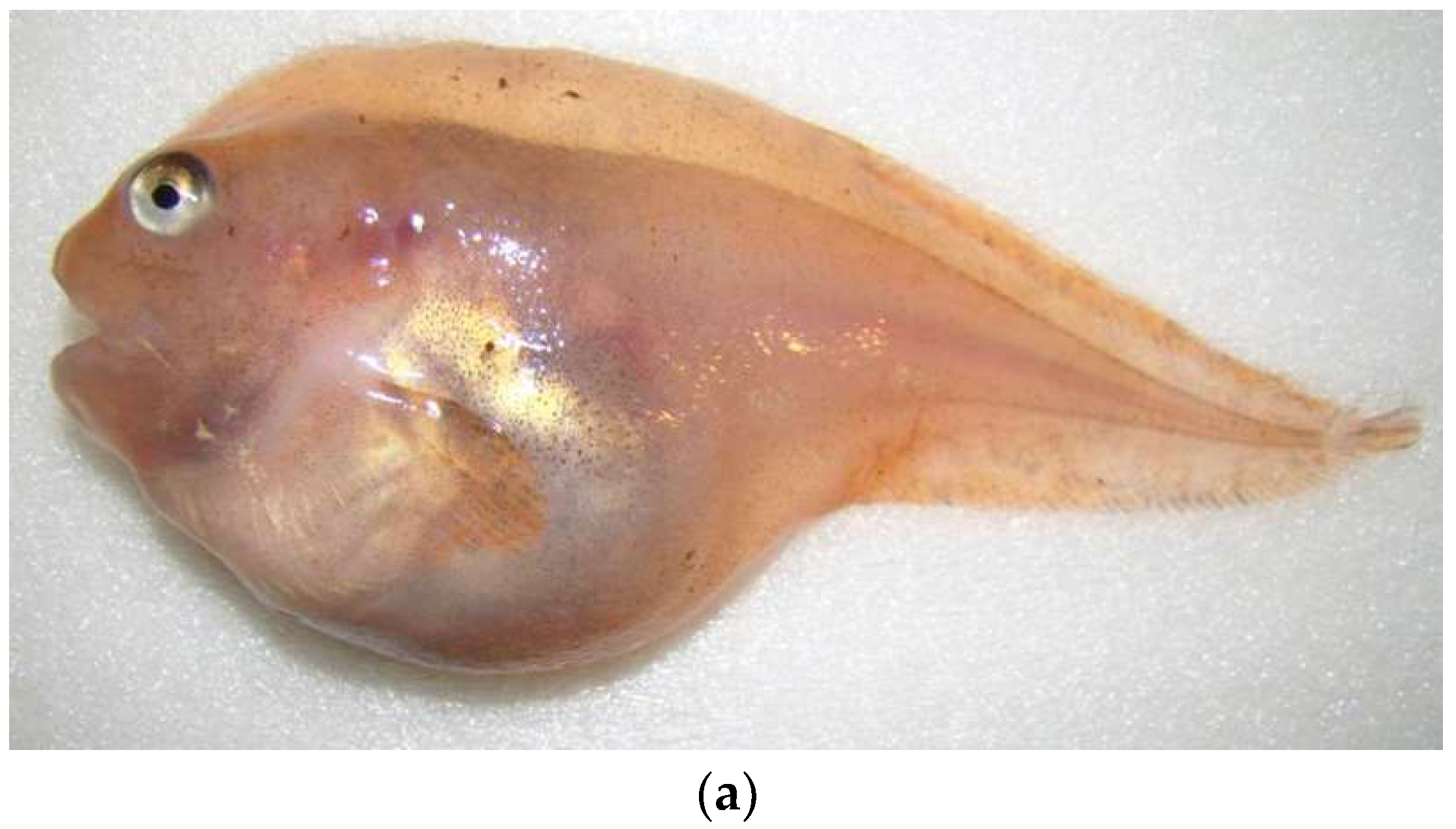{kind=link}
{kind=link}
| Genera | Species | Specimens | Stations/Lots | Radiograms |
|---|---|---|---|---|
| Liparis | 55 | 1555 | 525 | 359 |
| Careproctus | 27 | 549 | 192 | 142 |
| Paraliparis | 67 | 229 | 140 | 183 |
| Psednos | 32 | 42 | 39 | 45 |
| Rhodichthys | 2 | 7 | 5 | 5 |
| Total: | 183 | 2382 | 901 | 734 |
| Taxa | Range, Depth (m) | References |
|---|---|---|
| Genus Liparis Scopoli, 1777 | ||
| Subgenus Liparis Scopoli, 1777 | ||
| L. liparis (Linnaeus, 1766) * | SW Barents Sea (Murman), 0–78 m | [35] |
| L. montagui (Donovan, 1804) * | SW Barents Sea (Murman), 0–3 m | [35] |
| L. tunicatus Reinhardt, 1937 | Circumpolar, shallow waters [35]; SW, NW, NE Greenland [77] | [35,77] |
| Subgenus Careliparis Garman, 1892 | ||
| “L. megacephalus” group | ||
| L. bathyarcticus Parr, 1931 1 | Obviously circumpolar, depth 12–510, usually 5–350 m [35]; Gulf of St. Lawrence [78] | [35,78] |
| Subgenus Lycocara Gill, 1884 | ||
| Liparis cf. fabricii complex 2 L. fabricii Krøyer, 1847 L. koefoedi Parr, 1932 L. laptevi Popov, 1933 | Circumpolar, 12–628 (usually 40–350) m [39], near Greenland to 1460 m [77]; Gulf of St.Lawrence [78] | [39,77,78] |
| Genus Careproctus Kröyer, 1862 3 | ||
| Subgenus Careproctus Kröyer, 1862 | ||
| C. reinhardti sensu stricto (Krøyer, 1862) 3 | SW Greenland [37,79], Gulf of St. Lawrence [78] | [37,78,79] |
| C. longipinnis Burke, 1912 | Norwegian Sea, north of Faeroe Islands, 1322 m | [37,80,81] |
| C. solidus Chernova, 1999 | Laptev Sea, 2151–1934 m | [36,81] |
| C. lerikimae Orr, Kai et Nakabo, 2015 | Beaufort Sea, depth 178 m | [81] |
| “C. dubius” group | ||
| C. dubius Zugmayer, 1911 | Spitsbergen, Havre Green, 150 m | [37,81] |
| C. derjugini Chernova, 2005 | northeast of Spitsbergen, 344–363 m | [38] |
| C. knipowitschi Chernova, 2005 | Barents Sea, 298–293 m | [38,82] |
| C. macrophthalmus Chernova, 2005 | southwest of the Barents Sea, 260–275 m | [38,82] |
| C. tapirus Chernova, 2005 | southwest of the Barents Sea, 170–320 m | [38] |
| C. telescopus Chernova, 2005 | Barents Sea, 260–307 m | [38] |
| C. fulvus Chernova, 2014 | Kara Sea, Novaya Zemlya Trench, 190–414 m | [40] |
| C. rosa Chernova, 2014 | southwest of the Kara Sea, 140 m | [41] |
| C. karaensis Chernova, 2014 | Kara Sea, east of Novaya Zemlya, pelagic at 0−30 m | [41,81] |
| C. uter Chernova, 2014 | Kara Sea, Novaya Zemlya Trench, 206 m | [41] |
| C. carinatus Chernova, 2014 | southwest of the Kara Sea, pelagic at 0−30 m | [41] |
| Subgenus Careproctula Andriashev, 2003 | ||
| C. micropus (Günther, 1887) | Faeroe Channel, depth 540 and 608 fathoms (987–1112 m) | [37] |
| C. kidoi Knudsen et Møller, 2008 | Baffin Bay; SW Greenland, NW Greenland, 952–1487 m | [77,83] |
| C. moskalevi Andriashev et Chernova, 2010 | Norwegian Sea, southwest of the Bear Island, 1478–1691 m | [82,84] |
| C. latiosus Andriashev et Chernova, 2010 | Norwegian Sea, southwest of the Bear Island, 1478–1695 m | [82,84] |
| C. mica Chernova, 2014 | Kara Sea, Novaya Zemlya Trench, 204 m | [41,82] |
| C. canusocius Orr, 2020 | Beaufort Sea, 488–599 m | [82] |
| Genus Paraliparis Collett, 1879 | ||
| P. bathybius (Collett, 1879) | Norwegian Basin, 1000–1847 m and pelagic at 20–1000 m; Faroe Trench, 1170 m; Central Polar Basin: northeast of Spitsbergen [35], north of the Laptev Sea, 2824–2775 m [75]; Greenland, benthopelagic, 545–1600 m [77] | [35,75,77,85,86,87,88,89,90] |
| P. copei copei Goode et Bean, 1896 * | SW, SE Greenland, benthopelagic, (360) 710–1460 (1902) m [77]; Gulf of St.Lawrence [78]. Elsewhere found in the North Atlantic (see Table 2) | [77,78] |
| P. garmani Burke, 1912 * | SW, SE Greenland, 550–987. Elsewhere found in WN Atlantic (see Table 2). | [77] |
| P. violaceus Chernova, 1991 | Central Polar Basin, north of Severnaya Zemlya Archipelago, 2365 m | [35] |
| Genus Rhodichthys Collett, 1879 | ||
| Rh. regina Collett, 1879 | Norwegian Basin, 1394–2341 m, Faroe Bank, 400–500 m; Baffin Bay, 1200–1800 m, NW Greenland, NE Greenland, benthopelagic at 1180–1480 m; Polar Basin: northeast of Spitsbergen, 1080–1090 m, north of Severnaya Zemlya, 1445 m | [35,77,91,92,93,94,95,96] |
| Rh. melanocephalus Andriashev et Chernova, 2010 | Norwegian Basin, southwest of the Bear Island, 1470–1695 m | [84] |
| Genus Psednos Barnard 1927 | ||
| P. christinae Andriashev, l992 (*) | SE Greenland, 843–854 m. Elsewhere found in the Mid-Atlantic Ridge (see Table 2) | [77] |
| P. gelatinosus Chernova, 2001 | SE Greenland (63°05′54″ N), mesopelagic at 650–0 m | [46,77] |
| P. groenlandicus Chernova, 2001 (*) | SW Greenland (63–65° N) and SE Greenland (61°53′ N), mesopelagic at 786–1460 m. Elsewhere found in the Mid-Atlantic Ridge (see Table 2) | [46,77] |
| P. melanocephalus Chernova et Stein, 2002 | SW Greenland, 58°15′ N, 0–3172 m and 64°03′ N, 926 m; 949–962 m | [77,97,98] |
| P. micruroides Chernova, 2001 | SW Greenland (63°45′ N), 0–900 m and SE Greenland (63°50′18″ N), mesopelagic at 0–1333 m | [46,77,98] |
| Taxa | Range, Depth (m) | References |
|---|---|---|
| Genus Liparis Scopoli, 1777 | ||
| Subgenus Neoliparis Steindachner, 1876 | ||
| “L. montagui” group | ||
| L. montagui (Donovan, 1804) * | Europe: from Portugal to Murman, intertidal and shallow waters | [35,39,90] |
| “L. atlanticus” group | ||
| L. atlanticus (Jordan et Evermann, 1898) | WN Atlantic: Quebec, Newfoundland, Nova Scotia [99], Gulf of St. Lawrence [78]; intertidal to 90 m | [78,99] |
| “L. grebnitzkii” group | ||
| L. inquilinus Able, 1973 | WN Atlantic: from the Gulf of St. Lawrence to Cape Hatteras, 5–97 m | [101,102,103] |
| Subgenus Liparis Linnaeus, 1766 | ||
| L. coheni Able, 1976 | WN Atlantic: Gulf of Maine, Nova Scotia, Gulf of St. Lawrence, 2–210 m | [104] |
| L. liparis (Linnaeus, 1766) * | Europe: from the North Sea to Murman; intertidal to 78 m | [35,39,90] |
| L. barbatus Ekström, 1832 | Baltic Sea: Gulf of Finland, Gulf of Bothnia, shallow water, enters rivers | [35,105] (as L.liparis barbatus); [39] (as L. barbatus) |
| Genus Careproctus Kröyer, 1862 | ||
| Subgenus Careproctula Andriashev, 2003 | ||
| C. ranula (Goode et Bean, 1879) | WN Atlantic: Halifax Harbor, Nova Scotia, 95 m | [37,106] |
| “C. merretti” group | ||
| C. merretti Andriashev et Chernova, 1988 | EN Atlantic: Porcupine Seabight (49°37′ N, 13°49′ W), 3990–3920 m | [44] |
| C. aciculipunctatus Andriashev et Chernova, 1997 | EN Atlantic: south of the Porcupine bank (50°13.8′ N, 14°36.1′ W), 4100 m | [45] |
| Genus Paraliparis Collett, 1879 | ||
| P. abyssorum Andriashev et Chernova, 1997 | EN Atlantic: near the Porcupine Bank (49°54′ N, 13°56′ W), 3640–3715 m | [45] |
| P. bipolaris Andriashev, 1997 | WN Atlantic: southwest of Ireland (50°12′ N, 13°40′ W), 3000–3040 m | [107] |
| P. calidus Cohen, 1968 | WN Atlantic: Gulf of Mexico (27°25′ N, 93°40′ W), 730 m; Gulf of St. Lawrence | [78,108] |
| P. challengeri Andriashev, 1993 | EN Atlantic: Rockall Trough (57°01′ N, 10°05′ W), Porcupine Seabight (49°46′ N, 12°31′ W), 2000–2100 m | [109] |
| P. copei copei Goode et Bean, 1896 * | WN Atlantic: 430–1980 m; described from the Long Island (39°12′17″ N, 72°09′30″ W), 951 m. EN Atlantic: 1125–1400 m. Elsewhere found near South Greenland (see Table 1). | [48,110], our data |
| P. edwardsi (Vaillant, 1888) | EN Atlantic: coast of Marocco, between Cape Spartel and Cape Blanko (33°46′ N, 9°02′ E), 1319 m | [108,111,112] |
| P. garmani Burke, 1912 * | WN Atlantic: from Labrador to Cape Hatteras; benthopelagic, 550–987 m. Elsewhere found near South Greenland (see Table 1). | [77,80] |
| P. hystrix Merrett, 1983 | EN Atlantic: to the west of Britain, (49–59° N, 07–18° W), 255–1140 m [113]. WN Atlantic: (36–38° N, 70–74° W), 0–1008 m [90] | [90,113,114] |
| P. liparinus (Goode, 1881) | WN Atlantic: southeast of Long Island, 891 m | [109] |
| P. murielae Matallanas, 1984 | West of the Mediterranean Sea, 500–600 m | [115] |
| P. nigellus Chernova et Moller, 2008 | North Atlantic: Mid-Atlantic Ridge, between the Azores and Charlie–Gibbs Fracture Zone, 1950–2107 m | [48] |
| P. vailanti Chernova, 2004 | WN Atlantic: Laurentian Channel (46°39′ N, 58°41′ W), mesopelagic, 423 m over a depth of 1150 m | [116] |
| Genus Psednos Barnard, 1927 | ||
| P. andriashevi Chernova, 2001 | EN Atlantic: west of Ireland (54°21′ N, 17°59′ W), mesopelagic at 800 m | [46] |
| P. barnardi Chernova, 2001 | WN Atlantic: slope of New England (39°49′ N, 70°39′ W), mesopelagic at 1042–1368 m, juv at 750–1001 m | [46] |
| P. christinae Andriashev, l992 (*),1 | North Atlantic: Mid-Atlantic Ridge (49°48′ N, 25°55′ W), 1000–1500 m; Greenland (see Table 1) | [117] |
| P. delawarei Chernova et Stein, 2002 | WN Atlantic: south of Cape Cod (39°48′05″ N, 70°43′28″ W), 0–1000 m | [97] |
| P. islandicus Chernova et Stein, 2002 | EN Atlantic: 59°59.7′ N, 19°42.2′ W, 1250–1500 m | [97] |
| P. groenlandicus Chernova, 2001 (*) | North Atlantic: north of the Mid-Atlantic Ridge, mesopelagic, 981–2015 m over greater depth. Ellsewhere found near South Greenland (see Table 1) | [48] |
| P. harteli Chernova, 2001 | WN Atlantic: 40°45′ N, 65°03′ W, 1008–0 m | [46] |
| P. mirabilis Chernova, 2001 | WN Atlantic: slope of New England (39° N, 70°39′ W), 0–700 m over 1370–1700 m | [46] |
| P. rossi Chernova et Stein, 2004 2 | WN Atlantic: Cape Hatteras (35°30.036″ N, 74°46.497″ W), 500–674 m over 900 m | [118] |
| P. sargassicus Chernova, 2001 | Sargasso Sea (35°30′ N, 67°14′30″ W, 0–1050 m | [46] |
| P. spirohira Chernova et Stein, 2002 | EN Atlantic: west of northern Spain (41°56.2′ N, 16°50.1′ W), 985–1010 m | [92] |
| Genus Eutelichthys Tortonese, 1959 | ||
| E. leptochirus Tortonese, 1959 | EN Atlantic: southwestern part of the Mediterranean Sea and Bay of Lyon, 500–917 m | [90,112,119,120] |
| North Atlantic | Arctic | North Pacific |
|---|---|---|
| Subgenus Neoliparis Steindachner, 1876 | ||
| “L. montagui” group | ||
| * L. montagui Eur | – | L. burkei (Jordan et Thompson, 1914) Jap |
| L. chefuensis Wu et Wang, 1933 Yell | ||
| L. petschiliensis (Rendahl, 1926) Yell | ||
| L. punctulatus (Tanaka, 1916) Jap | ||
| “L. atlanticus” group | ||
| L. atlanticus WNA | – | L. rutteri (Gilbert et Snyder, 1898) Amer Ale |
| L. schantarensis (Lindberg et Dulkeit, 1929) Okh | ||
| L. schmidti Lindberg et Krasyukova, 1987 Jap | ||
| “L. grebnitzkii” group | ||
| L. inquilinus WNA | – | L. miostomus Matsubara et Iwai, 1954 Hokk, Kam |
| L. kusnetzovi Taranetz, 1935 Jap | ||
| L. mednius (Soldatov, 1930) Com | ||
| L. grebnitzkii (Schmidt, 1904) Com | ||
| Subgenus Liparis Scopoli, 1777 | ||
| L. coheni WNA | L. brashnikovi Soldatov, 1930 Jap, Kam | |
| * L. liparis Eur | L. tunicatus circ | L. frenatus (Gilbert et Burke, 1912) Jap, Hokk, Okh |
| L. barbatus Balt | L. marmoratus Schmidt, 1950 Okh, Ber | |
| L. bristolensis (Burke, 1912) Ber, Al | ||
| Subgenus Careliparis Garman, 1892 | ||
| “L. megacephalus” group | ||
| – | L. bathyarcticus circ | L. meridionalis Schmidt, 1950 Jap |
| L. punctatus Schmidt, 1950 Okh | ||
| L. eos Krasyukova, 1894 Sakh | ||
| L. rotundirostris Krasyukova, 1894 Sakh | ||
| L. megacephalus (Burke, 1912) eBer | ||
| L. brevicaudus Mori, 1956 Jap | ||
| L. quasimodo Krasyukova, 1894 Sakh | ||
| L. lindbergi Krasyukova, 1894 Sakh | ||
| Subgenus Lycocara Gill, 1884 | ||
| – | L. cf. fabricii complex circ (L. fabricii s. str. Spits L. koefoedi Spits L. laptevi Lap) | – |
| Subgenus Careproctus (pectoral radials notched, 2–3 interradial fenestra present; pleural ribs saber-like) | ||||
| Pectoral radials | 4 (3 + 1) | 3 (2 + 0 + 1) | 3 (2 + 0 + 1) | 2 (1 + 0 + 0 + 1) |
| Vertebrae | 59–64 | 60 | – | – |
| Pectoral fin rays | 29–36 | 31 | – | – |
| Pores 2–5–7–2 | C. reinhardti | C. solidus | – | – |
| C. longipinnis | ||||
| C. lerikimae | ||||
| “C. dubius”(11 species) 1 | ||||
| Subgenus Careproctula (pectoral radials unnotched; interradial fenestra absent; pleural ribs absent or rudimentary) | ||||
| Pectoral radials | 4 (3 + 1) | 3 (2 + 0 + 1) | 3 (2 + 0 + 1) | 2 (1 + 0 + 0 + 1) |
| Vertebrae | 60–62 | 57–62 | 64–65 | 69 |
| Pectoral fin rays | 26–28 | 22–29 | 27–28 | 22–25 |
| Pores 2–6–7–1 | C. ranula * C. kidoi | C. micropus | C. canusocius | C. merretti * C. aciculipunctatus * |
| C. moskalevi | ||||
| C. latiosus2 | ||||
| C. mica | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chernova, N.V. Amphiboreality and Distribution of Snailfishes (Cottiformes: Liparidae) in the Arctic and the North Atlantic. Diversity 2022, 14, 1097. https://doi.org/10.3390/d14121097
Chernova NV. Amphiboreality and Distribution of Snailfishes (Cottiformes: Liparidae) in the Arctic and the North Atlantic. Diversity. 2022; 14(12):1097. https://doi.org/10.3390/d14121097
Chicago/Turabian StyleChernova, Natalia V. 2022. "Amphiboreality and Distribution of Snailfishes (Cottiformes: Liparidae) in the Arctic and the North Atlantic" Diversity 14, no. 12: 1097. https://doi.org/10.3390/d14121097
APA StyleChernova, N. V. (2022). Amphiboreality and Distribution of Snailfishes (Cottiformes: Liparidae) in the Arctic and the North Atlantic. Diversity, 14(12), 1097. https://doi.org/10.3390/d14121097