

Spatial Variation in the Frequency of Left-Sided Morph in European Flounder Platichthys flesus (Linnaeus, 1758) from the Marginal Arctic (the White Sea)


Abstract
:1. Introduction
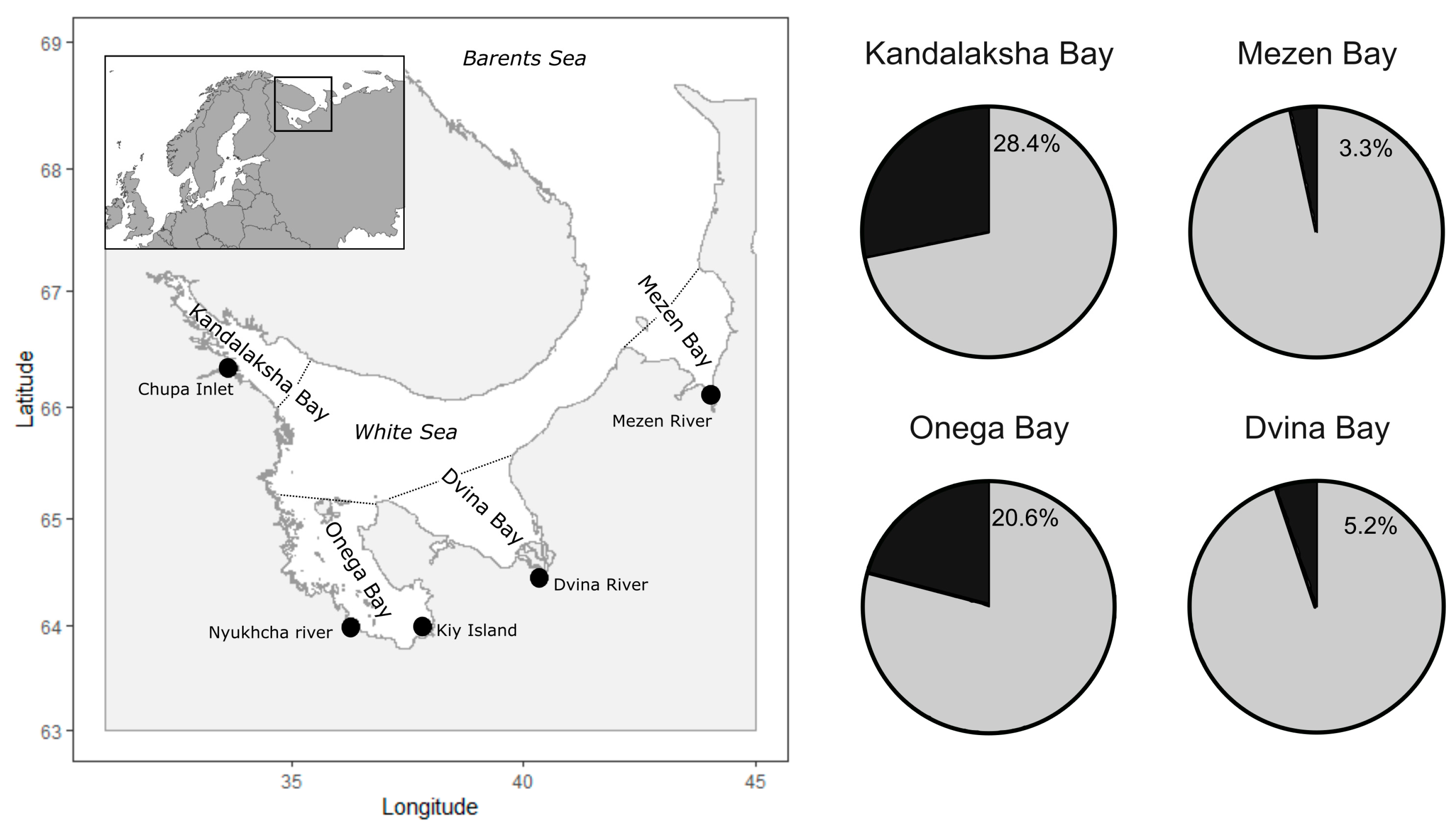
2. Materials and Methods
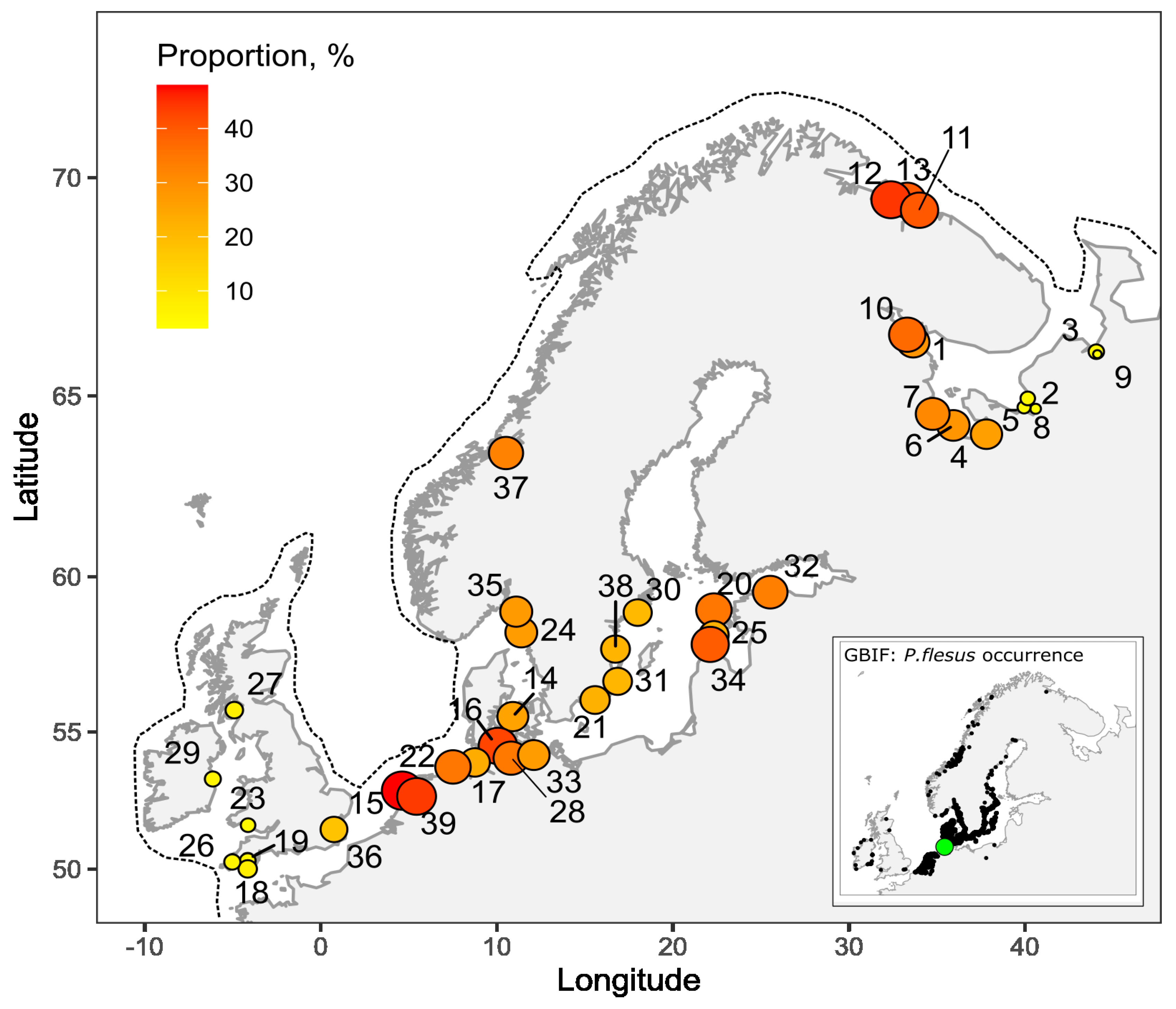
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berg, L.S. Freshwater Fishes of the USSR and Adjacent Countries; Nauka: Moscow, Russia, 1949. (In Russian) [Google Scholar]
- Andriashev, A.P. Fishes of the Northern Seas of the USSR; Izdatelstvo Akademii Nauk SSSR: Moscow, Russia, 1954. (In Russian) [Google Scholar]
- Mecklenburg, C.W.; Lynghammar, A.; Johannesen, E.; Byrkjedal, I.; Christiansen, J.S.; Dolgov, A.V.; Karamushko, O.V.; Mecklenburg, T.A.; Møller, P.R.; Steinke, D.; et al. Marine Fishes of the Arctic Region; Conservation of Arctic Flora and Fauna: Akureyri, Iceland, 2018. [Google Scholar]
- Fuks, G.V.; Goncharov, Y.V. First capture of European flounder Platichthys flesus (Pleuronectidae) in the southwestern part of the Kara Sea. J. Ichthyol. 2021, 61, 310–313. [Google Scholar] [CrossRef]
- Duncker, G. Variation und Asymmetrie bei Pleuronectes flesus L. Wiss. Meereuntersuchungen 1900, 3, 333–406. [Google Scholar]
- Mikelsaar, N. Method of equalized scales. In Hydrobiological Investigations, Issue 1; Izdatelstvo Akademii Nauk ESSR: Tartu, Russia, 1958; pp. 286–312. (In Russian) [Google Scholar]
- Fornbacke, M.; Gombrii, M.; Lundberg, A. Sidedness frequencies in the flounder Platichthys flesus (Pleuronectiformes) along a biogeographical cline. Sarsia 2002, 87, 392–395. [Google Scholar] [CrossRef]
- Galleguillos, R.A.; Ward, R.D. Genetic and morphological divergence between populations of the flatfish Platichthys flesus (L.) (Pleuronectidae). Biol. J. Linn. Soc. 1982, 17, 395–408. [Google Scholar] [CrossRef]
- Boklage, C.E. On the inheritance of directional asymmetry (sidedness) in the starry flounder, Platichthys stellatus: Additional analyses of Policansky’s data. Behav. Brain Sci. 1984, 7, 725–730. [Google Scholar] [CrossRef]
- Policansky, D. The asymmetry of flounders. Sci. Am. 1982, 246, 116–122. [Google Scholar] [CrossRef]
- Apstein, C. Junge Butt (Schollen, Pleuronectes platessa) in der Ostsee. Wiss. Meereuntersuchungen 1905, 8, 1–26. [Google Scholar]
- Redeke, H.C. Ueber den gegenwärtigen Stand unserer Kenntnis von den Rassen der wichtigsten Nutzfische. Die Lokalformen der Pleuronektiden (Scholle und Flunder). Rapports et Procès-Verbaux des Réunions - Conseil Permanent International Pour L’exploration De La Mer 1915, 22, 1–25. [Google Scholar]
- Nikolaev, A.P. Materials on the biology of European flounder of the Kuz Inlet of the White Sea. Izvestiya. Karelo-Fin. Fil. Akad. Nauk SSSR 1949, 4, 43–51. [Google Scholar]
- Shatunovsky, M.I. Materials on systematics of European flounder Pleuronectes flesus L. from the White Sea. Vestn. Mosc. State Univ. 1964, 1, 32–38. [Google Scholar]
- Dietrich, R. Populationsökologie der Plattfische (Familie Pleuronectidae) im Küsten- und Ästuarbereich des Weißen Meeres. Ph.D. Dissertation, University of Rostock, Rostock, Germany, 2009. [Google Scholar]
- Semushin, A.V.; Fuks, G.V.; Shilova, N.A. Flatfishes of the White Sea: New data on the biology of the Arctic flounder Liopsetta glacialis, European flounder Platichthys flesus, and common dab Limanda limanda. J. Ichthyol. 2015, 55, 527–539. [Google Scholar] [CrossRef]
- Yershov, P.N.; Fuks, G.V.; Khaitov, V.M. Frequencies of lateral morphs in different age classes of the flounder Platichthys flesus (Linnaeus, 1758) from the White Sea. Proc. Zool. Inst. RAS 2022, 326. in press. [Google Scholar]
- Fuks, G.V.; Yershov, P.N.; Khaitov, V.M. Long-term dynamics of the proportion of left-sided individuals in the populations of the European flounder Platichthys flesus (Linnaeus, 1758) in the White Sea. Proc. Zool. Inst. RAS 2021, 325, 273–277. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Сambridge, UK, 2002. [Google Scholar]
- Suvorov, E.K. Biology of Murmansk Pleuronectes flesus. Tr. Inst. Izuchenia Sev. 1927, 38, 56–63. (In Russian) [Google Scholar]
- Sych, N.S. Pleuronectes flesus of the Barents and White Seas. Tr. Gos. Nauchno-Issledovatelskogo Inst. Rybn. Khozyaistva 1930, 5, 89–116. (In Russian) [Google Scholar]
- Voronina, E.P. Morphology and systematics of river flounders of the genus Platichthys. J. Ichthyol. 1999, 39, 588–599. [Google Scholar]
- Elmhirst, R. On some ambicoloured flatfish from the Clyde. Ann. Scott. Nat. Hist. 1911, 78, 77–79. [Google Scholar]
- Russo, T.; Pulcini, D.; Costantini, D.; Pedreschi, D.; Palamara, E.; Boglione, C.; Cataudella, S.; Scardi, M.; Mariani, S. “Right” or “wrong”? Insights into the ecology of sidedness in European flounder, Platichthys flesus. J. Morphol. 2012, 273, 337–346. [Google Scholar] [CrossRef]
- Momigliano, P.; Denys, G.P.J.; Jokinen, H.; Merilä, J. Platichthys solemdali sp. nov. (Actinopterygii, Pleuronectiformes): A new flounder species from the Baltic Sea. Front. Mar. Sci. 2018, 5, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Nordgaard, O. Contributions to the life history of the fishes in Trondhjem fjord and environs. Det K. Nor. Vidensk. Selsk. Skr. 1915, 9, 1–38. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 31 May 2021).
- Cribari-Neto, F.; Zeileis, A. Beta Regression in R. J. Stat. Softw. 2010, 34, 1–24. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 19 April 2021).
- Ojaveer, E.; Drevs, T. Flounder, Platichthys flesus trachurus (Duncker). In Fishes of Estonia; Ojaveer, E., Pihu, E., Saat, T.E., Eds.; Estonian Academy Publishers: Tallinn, Estonia, 2003; pp. 362–370. [Google Scholar]
- Nikolaev, A.P. Fish species of Pomor and Karelia Coasts of the White Sea. Izvestiya Karelo-Finskogo Filiala. Akademii Nauk SSSR 1951, 3, 93–99. [Google Scholar]
- Mukhomediyarov, F.B. Biology and fishery of the noncommercial fishes in the coastal waters of the Karelia. In Papers on the Comprehensive Studies of the White Sea; Palenichko, Z.G., Ed.; Izdatelstvo Akademii Naus SSSR: Moscow, Russia, 1963; pp. 131–143. (In Russian) [Google Scholar]
- Florin, A.B. Flatfishes in the Baltic Sea—A review of biology and fishery with a focus on Swedish conditions. Finfo 2005, 14, 1–56. [Google Scholar]
- Nissling, A.; Dahlman, G. Fecundity of flounder, Pleuronectes flesus, in the Baltic Sea—Reproductive strategies in two sympatric populations. J. Sea Res. 2010, 64, 190–198. [Google Scholar] [CrossRef]
- Altukhov, К.А.; Мikhailovskaya, А.А.; Mukhomediyarov, F.B.; Nadezhin, V.M.; Novikov, P.I.; Palenichko, Z.G. Fishes of the White Sea; Godarstvennoe Izdatelstvo Karelskoy ASSR: Petrozavodsk, Russia, 1958. (In Russian) [Google Scholar]
- White Sea. Biological Resources and Problems of their Rational Exploitations; Scarlato, O.A., Ed.; Izdatelstvo ZIN RAS: Saint-Petersburg, Russia, 1995. (In Russian) [Google Scholar]
- Shatunovsky, M.I.; Chestnova, L.G. Some biological characteristics of the flounder from the Kandalaksha Bay in the White Sea. Rep. White Sea Biol. Stn. Mosc. State Univ. 1970, 3, 166–188. (In Russian) [Google Scholar]
- Bergstrom, C.A. Morphological evidence of correlational selection and ecological segregation between dextral and sinistral forms in a polymorphic flatfish, Platichthys stellatus. J. Evol. Biol. 2007, 20, 1104–1114. [Google Scholar] [CrossRef]
- Bergstrom, C.A.; Palmer, A.R. Which way to turn? Effect of direction of body asymmetry on turning and prey strike orientation in starry flounder Platichthys stellatus (Pallas) (Pleuronectidae). J. Fish Biol. 2007, 71, 737–748. [Google Scholar] [CrossRef]
- Bergstrom, C.A.; Alba, J.; Pacheco, J.; Fritz, T.; Tamone, S.L. Polymorphism and multiple correlated characters: Do flatfish asymmetry morphs also differ in swimming performance and metabolic rate? Ecol. Evol. 2019, 9, 4772–4782. [Google Scholar] [CrossRef] [Green Version]
- Elmgren, R. Understanding human impact on the Baltic ecosystem: Changing views in recent decades. Ambio 2001, 30, 222–231. [Google Scholar] [CrossRef]
- Korpinen, S.; Meski, L.; Andersen, J.H.; Laamanen, M. Human pressures and their potential impact on the Baltic Sea ecosystem. Ecol. Indic. 2012, 15, 105–114. [Google Scholar] [CrossRef]
- Olsson, J.; Bergstrom, L.; Gardmark, A. Abiotic drivers of coastal fish community change during four decades in the Baltic Sea. ICES J. Mar. Sci. 2012, 69, 961–970. [Google Scholar] [CrossRef]
- Jokinen, H.; Wennhage, H.; Lappalainen, A.; Ådjers, K.; Rask, M.; Norkko, A. Decline of flounder (Platichthys flesus (L.)) at the margin of the species' distribution range. J. Sea Res. 2015, 105, 1–9. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No | Locality | Sample Size | Left, % | Source of Data |
|---|---|---|---|---|
| 1 | Chupa Inlet, Kandalaksha Bay, White Sea, Russia | 584 | 28.4 | Present study |
| 2 | Delta of the Northern Dvina River, Dvina Bay, White Sea, Russia | 5007 | 5.2 | Present study |
| 3 | Mezen river, Mezen Bay, White Sea, Russia | 2272 | 3.3 | Present study |
| 4 | Nyukhcha river and Kiy island area, Onega Bay, White Sea, Russia | 4527 | 20.6 | Present study |
| 5 | Delta of the Northern Dvina River, White Sea, Russia | 897 | 4 | [15] |
| 6 | Kolezhma river, Onega Bay, White Sea, Russia | 358 | 28.5 | [6] |
| 7 | Kuz Inlet, Onega Bay, White Sea, Russia | 187 | 31.3 | [13] |
| 8 | Delta of the Northern Dvina River, White Sea, Russia | 2394 | 4.7 | [16] |
| 9 | Mezen Bay, White Sea, Russia | 1367 | 3.1 | [16] |
| 10 | Velikaya Salma Strait, Kandalaksha Bay, White Sea, Russia | 957 | 37.1 | [14,15] |
| 11 | Murman coast, Barents Sea, Russia | no data | 39.6 | [20] |
| 12 | Murman coast, Barents Sea, Russia | 475 | 44.5 | [21] |
| 13 | Murman coast, Barents Sea, Russia | 25 | 40 | [22] |
| 14 | Danish Belt Sea | 49 | 25.4 | [8] |
| 15 | Den Helder, North Sea, Netherlands | 75 | 48 | [12] |
| 16 | Eckernforde Bay; Laboe (Kiel), Baltic Sea, Germany | 3331 | 42.7 | [11] |
| 17 | Elbe river mouth, North Sea, Germany | 225 | 23.6 | [5] |
| 18 | English Channel, Plymouth, UK | 1120 | 5.4 | [5] |
| 19 | English Channel, Plymouth, UK | 40 | 7.5 | [8] |
| 20 | Hiiumaa, Baltic Sea, Estonia | 800 | 35 | [6] |
| 21 | Karlskrona, Baltic Sea, Sweden | 631 | 22.4 | [7] |
| 22 | Langeoog, Wadden Sea, North Sea, Germany | 26 | 35 | [22] |
| 23 | Loughor estuary, South Wales, UK | 64 | 4.7 | [8] |
| 24 | Lysekil, Skagerrak, Sweden | 653 | 27 | [7] |
| 25 | Mandjala, Saaremaa, Baltic Sea, Estonia | 200 | 22.5 | [6] |
| 26 | Mevagissey harbour, UK | 192 | 5.7 | [5] |
| 27 | Millport, Cumbrae, Scottish coast, UK | no data | 6.7 | [23] |
| 28 | Neustadt Bay, Baltic Sea, Germany | 90 | 34.4 | [5] |
| 29 | North Bull Island, Dublin Bay, Irish Sea | 590 | 5.6 | [24] |
| 30 | Nynashamn, Baltic Sea, Sweden | 186 | 20.1 | [7] |
| 31 | Oland, Baltic Sea, Sweden | 1673 | 21.1 | [7] |
| 32 | Pudisoo, Baltic Sea, Estonia | 1271 | 33.1 | [6] |
| 33 | Rostock, Baltic Sea, Germany | 15 | 26.7 | [25] |
| 34 | Sorve peninsula, Saaremaa, Baltic Sea, Estonia | 200 | 39.5 | [6] |
| 35 | Strömstad, Skagerrak, Sweden | 455 | 27.5 | [7] |
| 36 | Thames estuary, London, England | 50 | 18 | [8] |
| 37 | Trondheimsfjord, Norway | 269 | 32.3 | [26] |
| 38 | Vastervik, Baltic Sea, Sweden | 193 | 21.2 | [7] |
| 39 | Zuiderzee, North Sea, Netherlands | 50 | 44 | [12] |
| Term | Parameter | SE | z-Value | p-Value |
|---|---|---|---|---|
| (Intercept) | −1.043 | 0.1957 | −5.33 | <0.0001 |
| Distance | −0.001 | 0.0003 | −3.86 | <0.0001 |
| Longitude | 0.049 | 0.0109 | 4.55 | <0.0001 |
| Precision coefficient (phi) | 25.049 | 6.8835 | 3.64 | <0.0001 |
| Term | Parameter | SE | z-Value | p-Value |
|---|---|---|---|---|
| (Intercept) | 9.166 | 1.7262 | 5.31 | <0.0001 |
| Distance | −0.00003 | 0.0009 | −0.03 | 0.975 |
| Longitude | −0.286 | 0.0348 | −8.22 | <0.0001 |
| Precision coefficient (phi) | 54.5867 | 21.4711 | 2.54 | 0.01101 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yershov, P.N.; Fuks, G.V.; Khaitov, V.M. Spatial Variation in the Frequency of Left-Sided Morph in European Flounder Platichthys flesus (Linnaeus, 1758) from the Marginal Arctic (the White Sea). Diversity 2022, 14, 1004. https://doi.org/10.3390/d14111004
Yershov PN, Fuks GV, Khaitov VM. Spatial Variation in the Frequency of Left-Sided Morph in European Flounder Platichthys flesus (Linnaeus, 1758) from the Marginal Arctic (the White Sea). Diversity. 2022; 14(11):1004. https://doi.org/10.3390/d14111004
Chicago/Turabian StyleYershov, Peter N., Gennadiy V. Fuks, and Vadim M. Khaitov. 2022. "Spatial Variation in the Frequency of Left-Sided Morph in European Flounder Platichthys flesus (Linnaeus, 1758) from the Marginal Arctic (the White Sea)" Diversity 14, no. 11: 1004. https://doi.org/10.3390/d14111004
APA StyleYershov, P. N., Fuks, G. V., & Khaitov, V. M. (2022). Spatial Variation in the Frequency of Left-Sided Morph in European Flounder Platichthys flesus (Linnaeus, 1758) from the Marginal Arctic (the White Sea). Diversity, 14(11), 1004. https://doi.org/10.3390/d14111004