
The Key Environmental Factors Shaping Coastal Fish Community in the Eastern Gulf of Finland, Baltic Sea


Abstract
:1. Introduction
2. Materials and Methods
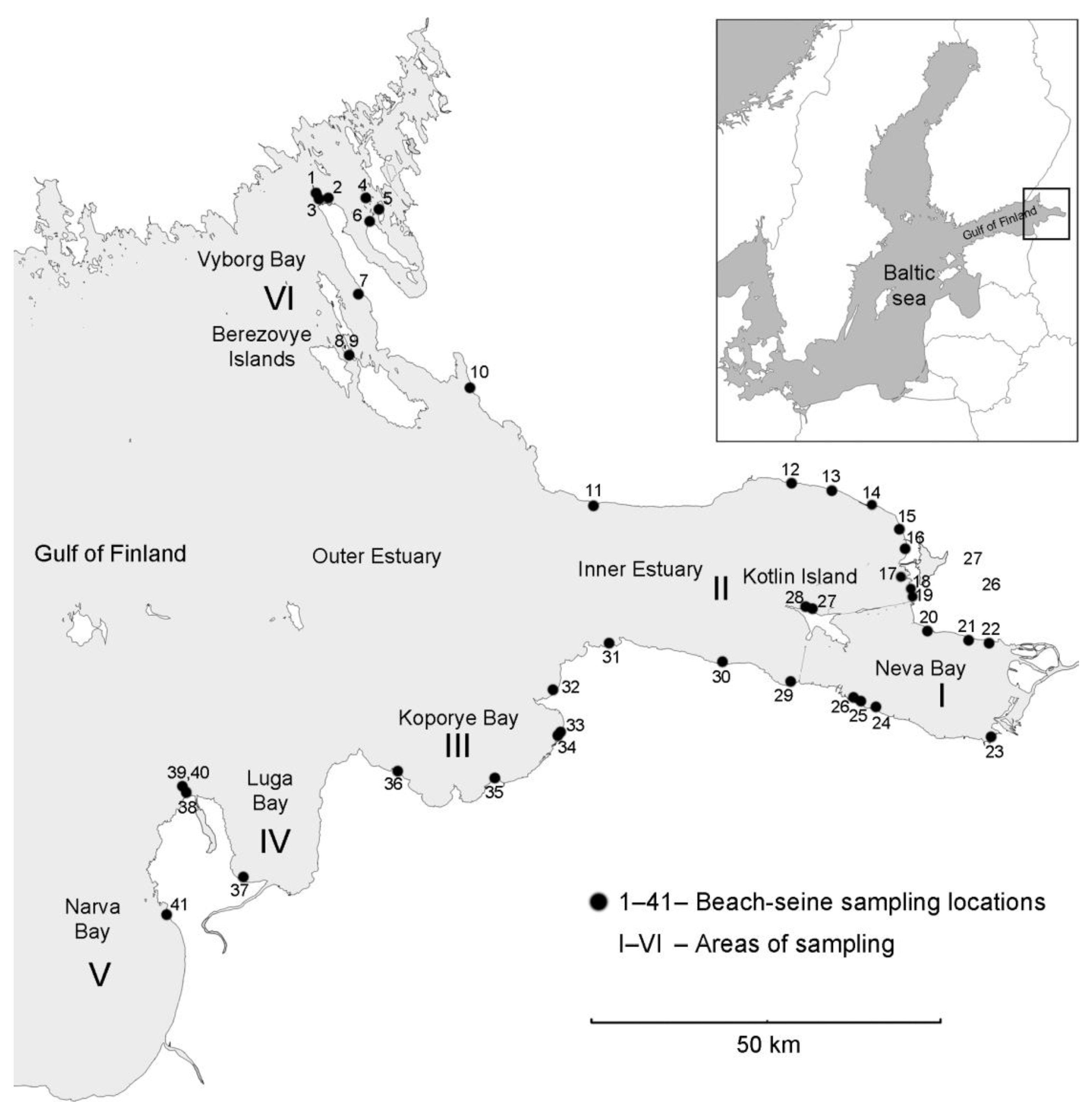
2.1. Study Site, Sampling and Laboratory Analysis
2.2. Measurement of Environmental Parameters
2.3. Statistical Analysis
3. Results
3.1. Results of Field Observations
3.1.1. Coastal Fish Species Diversity
3.1.2. Environmental Conditions
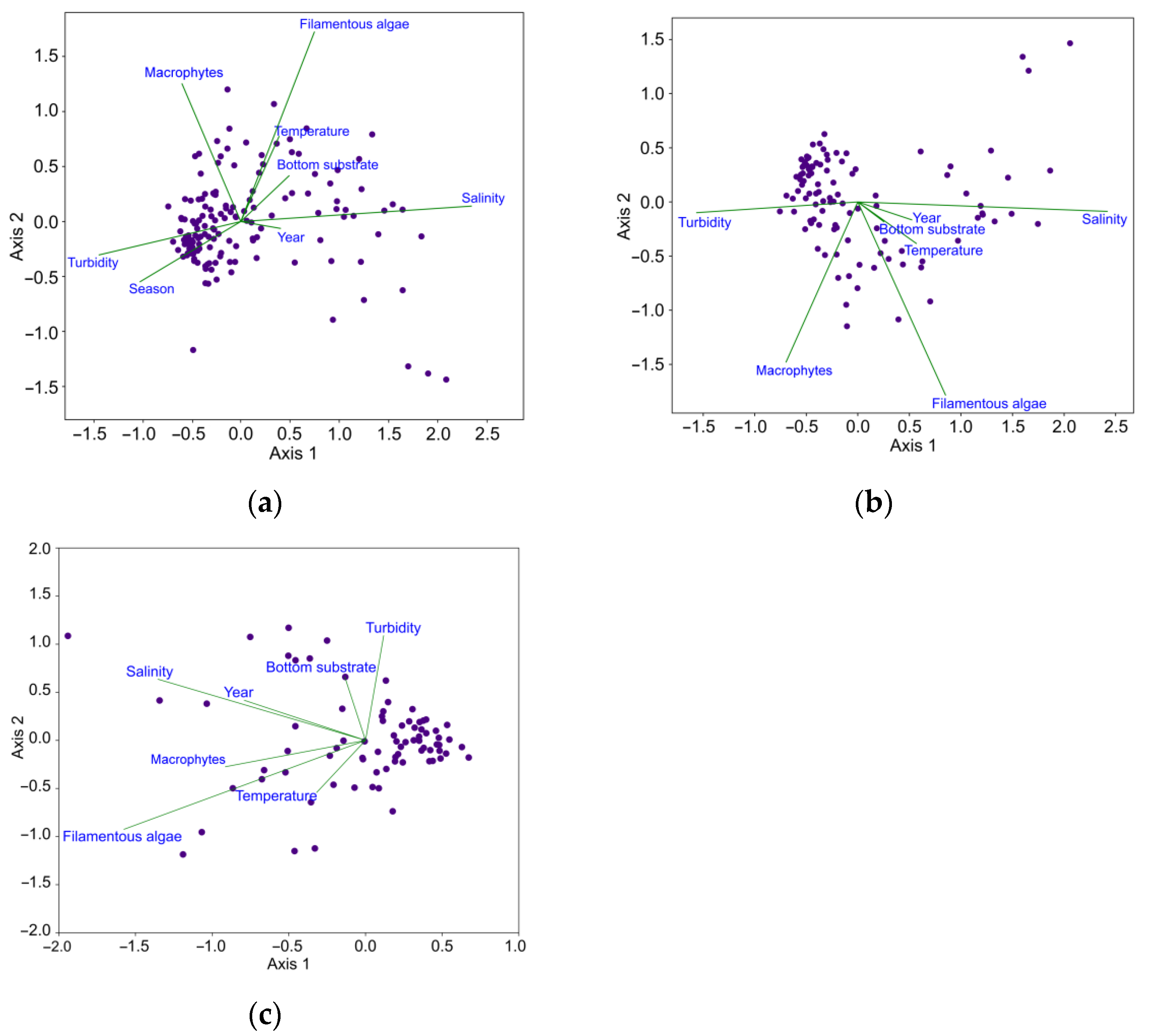
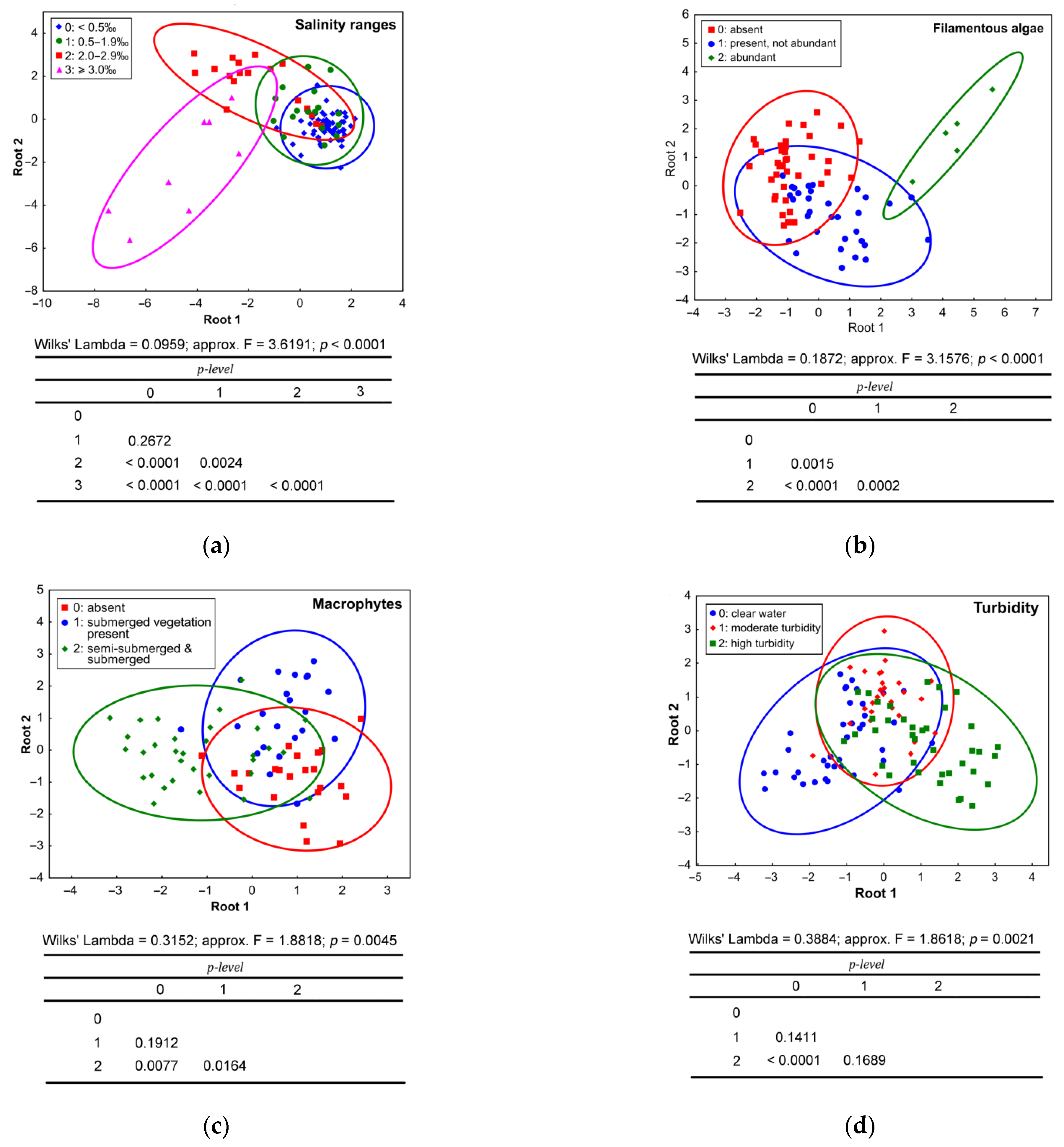
3.2. Results of Data Analysis
3.2.1. Identification of Major Environmental Factors Affecting the Fish Community
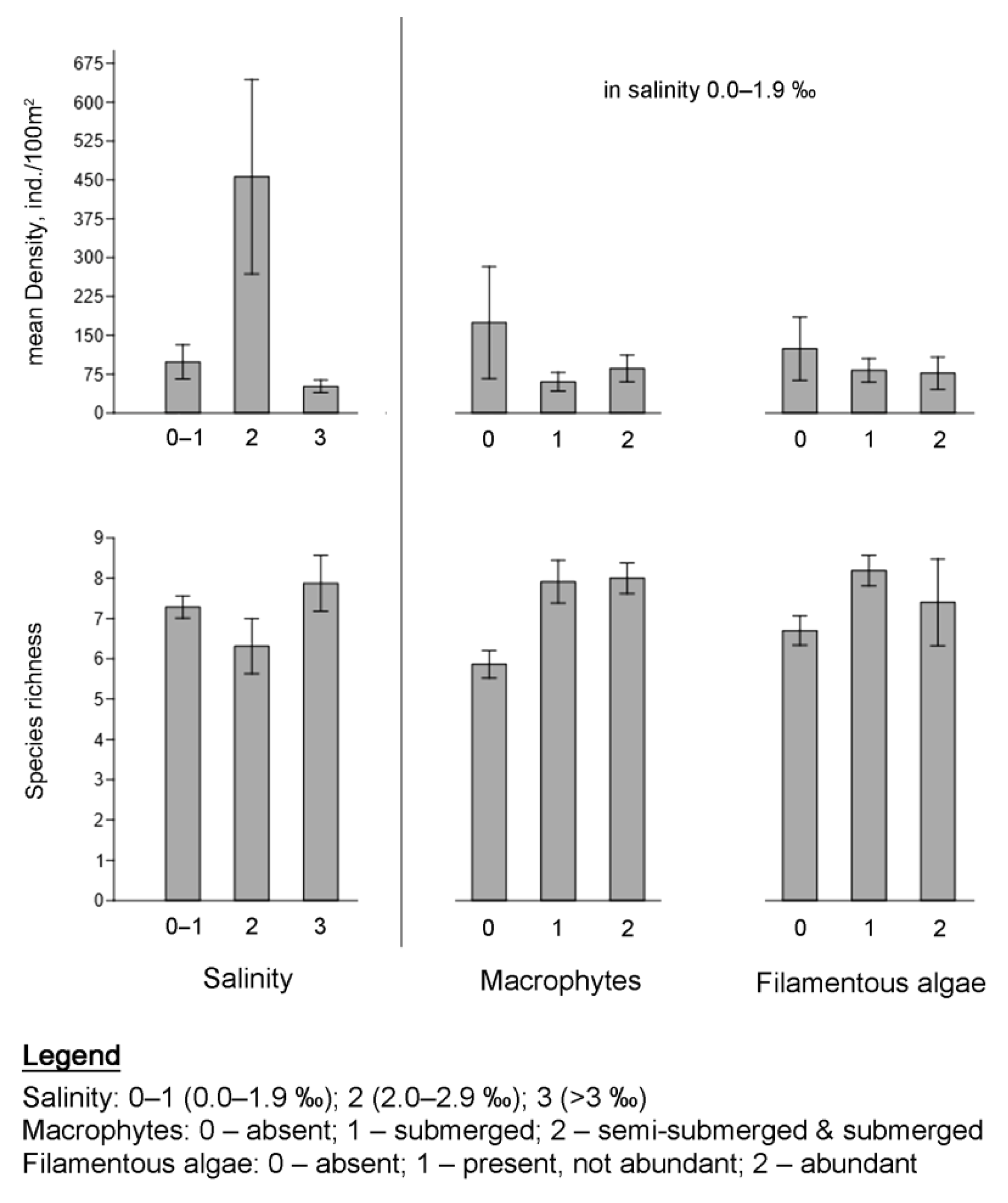
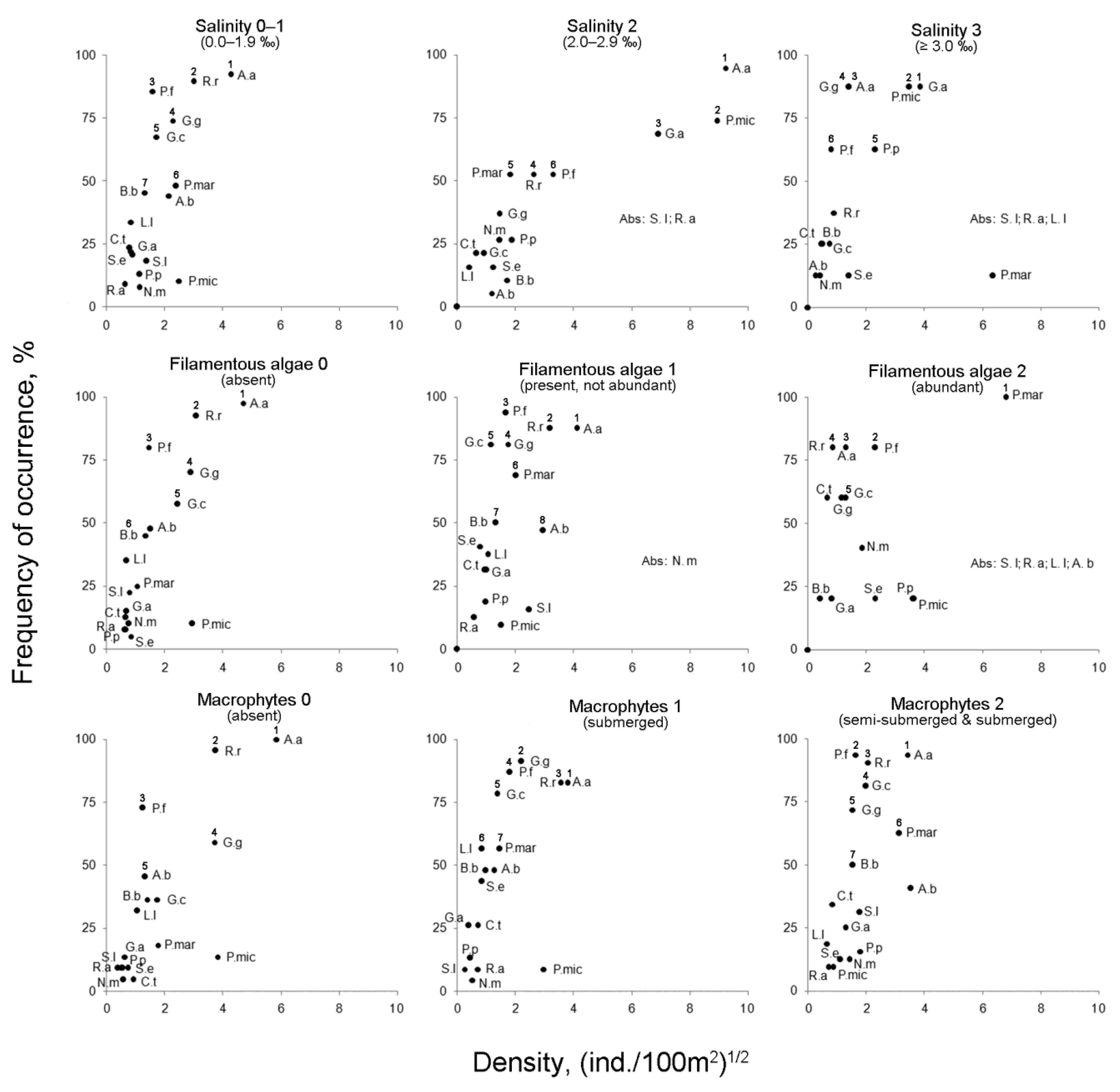
3.2.2. Influence of Environmental Factors on the Species Richness and the Fish Density
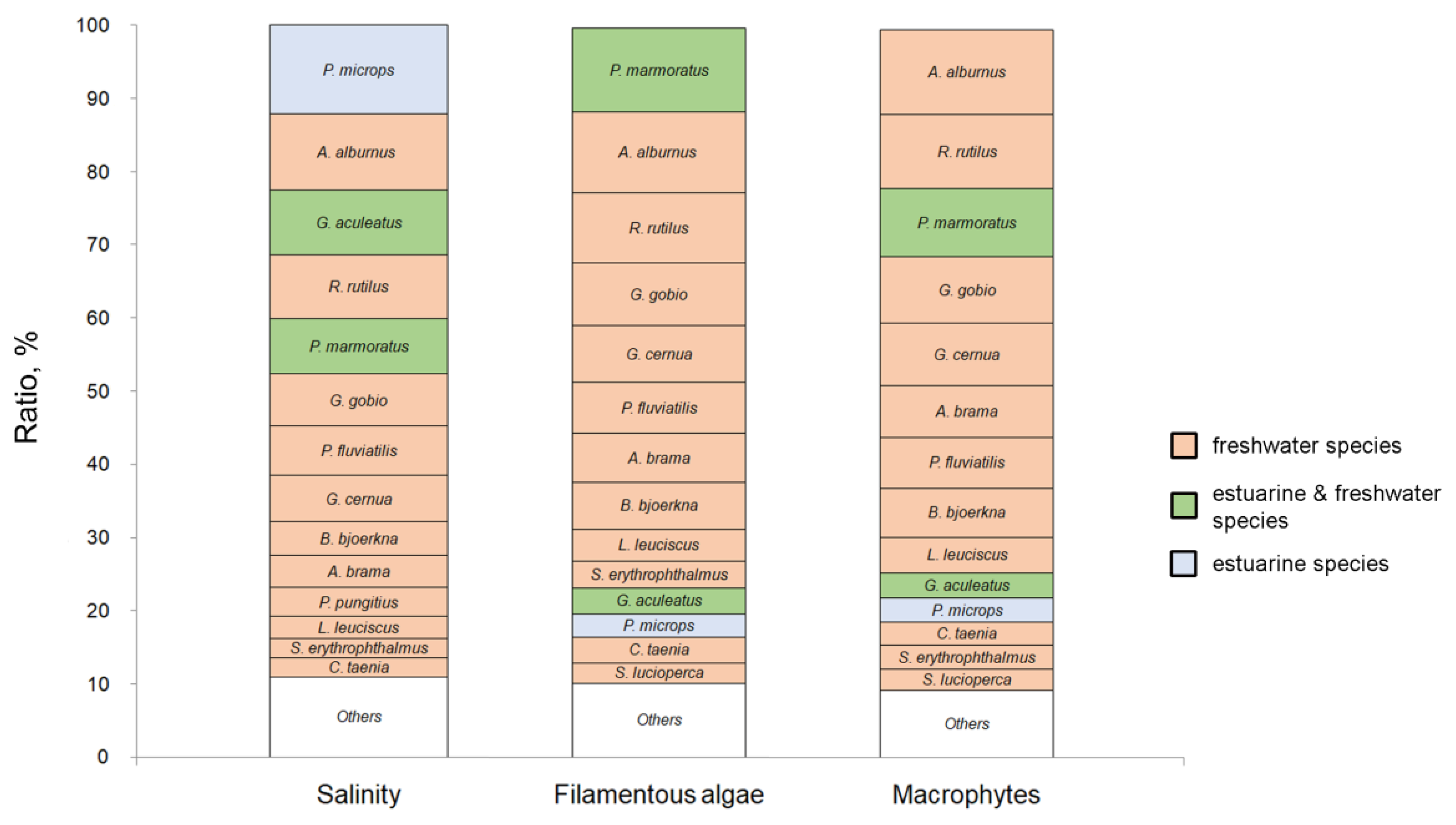
3.2.3. Differences in the Species Composition, Occurrence, and Density
4. Discussion
4.1. Salinity
4.2. Filamentous Algae
4.3. Macrophytes
4.4. Other Environmental Factors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variables | Area | |||||||
|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | |||
| A = 40 | A = 78 | A = 8 | A = 7 | A = 8 | A = 10 | |||
| Ranged variables | ||||||||
| Variables | Values | Description | Location N° | |||||
| Salinity ranges | 0 | <0.5‰ | 20–26 | 13–19; 27–30 | ||||
| 40 | 51 | |||||||
| 1 | 0.5–1.9‰ | 11–15; 27; 29–31 | 35 | |||||
| 24 | 1 | |||||||
| 2 | 2.0–2.9‰ | 29–30 | 32–36 | 37 | 41 | 1–10 | ||
| 2 | 7 | 2 | 1 | 10 | ||||
| 3 | ≥3.0‰ | 31 | 37 | 38–41 | ||||
| 1 | 6 | 7 | ||||||
| Bottom substrate | 1 | pure sand | 23–25 | 14; 16; 27–30 | 32; 34; 36 | 37 | 9 | |
| 22 | 28 | 3 | 7 | 1 | ||||
| 2 | stones prevail | 11–13; 15; 17; 31 | 33 | 40 | 1; 10 | |||
| 13 | 1 | 1 | 3 | |||||
| 3 | mixed | 20–22; 26 | 13; 15; 18; 19; 27; 30; 31 | 35 | 38; 39; 41 | 2; 3; 5–8 | ||
| 18 | 37 | 4 | 7 | 6 | ||||
| Macrophytes | 0 | absent | 20; 22; 23 | 11; 13–19 | 32; 34; 36 | 37 | 7; 10 | |
| 7 | 30 | 3 | 7 | 3 | ||||
| 1 | submerged | 20; 22 | 12; 13; 18; 28 | 35 | 39; 40 | 1; 2; 3; 5 | ||
| 8 | 18 | 2 | 2 | 4 | ||||
| 2 | semi-submerged and submerged | 21; 24–26 | 18; 27; 29–31 | 33; 35 | 38; 41 | 6; 8; 9 | ||
| 25 | 30 | 3 | 6 | 3 | ||||
| Filamentous algae | 0 | absent | 20–25 | 11–19; 29–31 | 34–36 | 37 | 41 | 5–7 |
| 31 | 43 | 3 | 5 | 1 | 3 | |||
| 1 | present, not abundant | 20; 22; 24–26 | 11; 13; 15; 17; 18; 27–30 | 32 | 37 | 38; 39; 41 | 3; 10 | |
| 8 | 30 | 1 | 2 | 5 | 2 | |||
| 2 | abundant | 24 | 13; 27; 31 | 33; 35 | 40; 41 | 1; 2; 8–10 | ||
| 1 | 5 | 4 | 2 | 5 | ||||
| Turbidity | 0 | clear water | 20; 22–25 | 12–14; 17–19; 27; 28; 30; 31 | 32–36 | 37 | 38–41 | 1–3; 6; 8–10 |
| 7 | 22 | 7 | 4 | 7 | 8 | |||
| 1 | moderate turbidity | 20; 22; 25 | 11; 13; 15; 17–19; 27; 29–31 | 37 | 5; 7 | |||
| 10 | 26 | 2 | 2 | |||||
| 2 | high turbidity | 20–26 | 13; 15–18; 27; 29–31 | 35 | 37 | 41 | ||
| 23 | 30 | 1 | 1 | 1 | ||||
| Measured parameters | ||||||||
| Parameter | Value | Limits (min–max) | ||||||
| Temperature | °C | 3.8–25.1 | 3.5–24.5 | 16.0–28.8 | 12.5–27.2 | 10.9–22.5 | 15.1–22.9 | |
| Salinity | ‰ | 0.0–0.4 | 0.1–3.1 | 1.3–2.7 | 2.8–3.8 | 2.9–4.5 | 1.8–2.9 | |
References
- Hupfer, P. Baltic Sea—Small Sea, Big Problems; Gidrometizdat: Leningrad, Russia, 1982; p. 136. (In Russian) [Google Scholar]
- Horackiewicz, J.; Skora, K.E. A seasonal pattern of occurrence of gobiid fish (Gobiidae) in the shallow littoral zone (0–1 m depth) of Puck bay. Oceanol. Stud. 1998, 3, 3–17. [Google Scholar]
- Sapota, M.R.; Skora, K.E. Fish abundance in shallow inshore waters of the Gulf of Gdansk. In Proceedings of the Polish—Swedish Symposium on Baltic Costal Fisheries, Sea fisheries institute, Gdynia, Poland, 2–3 April 1996; pp. 215–223. [Google Scholar]
- Kallasvuo, M.; Lappalainen, A.; Urho, L. Coastal reed belts as fish reproduction habitats. Boreal Environ. Res. 2011, 16, 1–14. [Google Scholar]
- Ustups, D.; Urtans, E.; Minde, A.; Uzars, D. The structure and dynamics of fish communities in the Latvian coastal zone (Pape—Pērkone), Baltic Sea. Acta Univ. Latv. 2003, 662, 33–44. [Google Scholar]
- Burke, L.; Kura, Y.; Kassem, K. Pilot Analysis of Global Ecosystems: Coastal Ecosystems; World Resources Institute: Washington DC, USA, 2001. [Google Scholar]
- Beyst, B.; Hostens, K.; Mees, J. Factors influencing fish and macrocrustacean communities in the surf zone of sandy beaches in Belgium: Temporal variation. J. Sea Res. 2001, 46, 281–294. [Google Scholar] [CrossRef]
- Vinberg, G.G.; Gutelmacher, B.L. (Eds.) Neva Bay: Hydrobiological Research; Nauka: Leningrad, Russia, 1987; p. 216. (In Russian) [Google Scholar]
- Kraufvelin, P.; Pekcan-Hekim, Z.; Bergström, U.; Florin, A.-B.; Lehikoinen, A.; Mattila, J.; Arula, T.; Briekmane, L.; Brown, E.J.; Celmer, Z.; et al. Essential coastal habitats for fish in the Baltic Sea. Estuar. Coast. Shelf Sci. 2018, 204, 14–30. [Google Scholar] [CrossRef] [Green Version]
- Gogoberidze, G.G.; Ryabchuk, D.V.; Zhamoyda, V.A.; Chubarenko, B.V.; Bobykina, V.P.; Babakov, A.N.; Bass, O.V.; Gushchin, A.V.; Ezhova, E.E.; Stont, J.I.; et al. The Baltic Sea. In Scientific Support for Balanced Business Planning on Unique Marine Coastal Landscapes and Suggestions for Its Use on the Example of the Azov-Black Sea Coast; Kosyan, R.D., Ed.; IO RAN: Gelendzhik, Russia, 2013; pp. 238–487. (In Russian) [Google Scholar]
- Thorman, S. Physical factors affecting the abundance and species richness of fishes in the shallow waters of the southern Bothnian Sea (Sweden). Estuar. Coast. Shelf Sci. 1986, 22, 357–369. [Google Scholar] [CrossRef]
- Lappalainen, A.; Shurukhin, A.; Alekseev, G.; Rinne, J. Coastal-fish communities along the Northern coast of the Gulf of Finland, Baltic sea: Responses to salinity and Eutrophication. Internat. Rev. Hydrobiol. 2000, 85, 687–696. [Google Scholar] [CrossRef]
- Shurukhin, A.S.; Lukin, A.A.; Pedchenko, A.P.; Titov, S.F. Modern condition of fishery and effectiveness using of fish supply in the Finnish Bay of the Baltic Sea. Proc. VNIRO 2016, 160, 60–69. [Google Scholar]
- HELCOM. Changing Communities of Baltic Coastal Fish Executive summary: Assessment of coastal fi sh in the Baltic Sea. Baltic Sea Environment Proceedings; Erweko Painotuote Oy: Helsinki, Finland, 2006; Volume 103B, p. 11. [Google Scholar]
- HELCOM. State of the Baltic Sea—Second HELCOM holistic assessment 2011–2016. Baltic Sea Environment Proceedings; HELCOM–Helsinki Commission: Helsinki, Finland, 2018; Volume 155, p. 155.
- Adjers, K.; Appelberg, M.; Eschbaum, R.; Lappalainen, A.; Minde, A.; Repečka, R.; Thoresson, G. Trends in coastal fish stocks of the Baltic Sea. Boreal Environ. Res. 2006, 11, 13–25. [Google Scholar]
- HELCOM. Biodiversity in the Baltic Sea—An Integrated Thematic Assessment on Biodiversity and Nature Conservation in the Baltic Sea. Baltic Sea Environment Proceedings; Erweko Painotuote Oy: Helsinki, Finland, 2009; Volume 116B, p. 188. [Google Scholar]
- HELCOM. Status of Coastal Fish Communities in the Baltic Sea during 2011–2016—The Third Thematic Assessment. Baltic Sea Environment Proceedings; HELCOM–Helsinki Commission: Helsinki, Finland, 2018; Volume 161, p. 50.
- Sundblad, G.; Bergström, U. Shoreline development and degradation of coastal fish reproduction habitats. Ambio 2014, 43, 1020–1028. [Google Scholar] [CrossRef] [Green Version]
- Shiewer, U. Ecology of Baltic Coastal Waters; Springer: Berlin/Heidelberg, Germany, 2008; p. 430. [Google Scholar]
- Zhigulsky, V.A.; Shuisky, V.F.; Chebykina, E.Y.; Fedorov, V.A.; Panichev, V.V.; Uspenskiy, A.A.; Zhigulskaya, D.V.; Bylina, T.S.; Bulysheva, M.M.; Bulysheva, A.M. Macrophyte Thicket Ecosystems of the Neva Bay; Scientific research programme results of the 1st stage/Eco-Express-Service LLC; Renome: St. Petersburg, Russia, 2020; p. 304, (In Russian). [Google Scholar] [CrossRef]
- Thiel, R.; Potter, I.C. The ichthyofaunal composition of the Elbe Estuary: An analysis in space and time. Mar. Biol. 2001, 138, 603–616. [Google Scholar] [CrossRef]
- Thiel, R.; Cabral, H.; Costa, M.J. Composition, temporal changes and ecological guild classification of the ichthyofaunas of large European estuaries—A comparison between the Tagus (Portugal) and the Elbe (Germany). J. App. Ichthyol. 2003, 19, 330–342. [Google Scholar] [CrossRef]
- Ioganzen, B.G.; Fajzova, L.V. On the determination of indicators of occurrence, abundance, biomass and their ratio in some aquatic organisms. Proc. All-Union Hydrobiol. Soc. Acad. Sci. USSR 1978, 22, 215–225. (In Russian) [Google Scholar]
- Žiliukas, V.; Žiliukienė, V.; Repečka, R. Temporal variation in juvenile fish communities of Kaunas reservoir littoral zone, Lithuania. Cent. Eur. J. Biol. 2012, 7, 858–866. [Google Scholar] [CrossRef] [Green Version]
- Khlebovich, V.V. Critical Salinity of Biological Processes; Nauka: Leningrad, Russia, 1974; p. 236. (In Russian) [Google Scholar]
- Andreeva, S.I.; Andreev, N.I. Evolutionary Transformations of Bivalve Molluscs of the Aral Sea in Conditions of Ecological Crisis; Publishing House of the Omsk State Pedagogical University: Omsk, Russia, 2003; p. 382. (In Russian) [Google Scholar]
- Aladin, N.V.; Plotnikov, I.S. The concept of relativity and multiplicity of barrier salinity zones and forms of existence of the hydrosphere. Proc. Zool. Inst. Russ. Acad. Sci. 2013, 3, 7–21. (In Russian) [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1998; p. 853. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; Plymouth Marine Laboratory: Plymouth, UK, 2001; p. 176. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E LTD: Plymouth, UK, 2006; p. 190. [Google Scholar]
- Pihl, L.; Wennhage, H.; Nilsson, S. Fish assemblage structure in relation to macrophytes and filamentous epiphytes in shallow non-tidal rocky- and soft-bottom habitats. Environ. Biol. Fishes 1994, 39, 271–288. [Google Scholar] [CrossRef]
- Konstantinov, A.S. General Hydrobiology, 4th ed.; Vishaya schkola: Moskow, Russia, 1986; p. 472. (In Russian) [Google Scholar]
- Alimov, A.F.; Golubkov, S.M. Ecosystem of the Neva River Estuary: Biodiversity and Ecological Problems; Association of Scientific Publications KMK: St. Petersburg/Moskow, Russia, 2008; p. 477. (In Russian) [Google Scholar]
- Ostov, I.M. Characteristic features of the hydrological and hydrochemical regime of the Gulf of Finland as the basis for its fishery development. Izv. GosNIORKh 1971, 76, 18–45. (In Russian) [Google Scholar]
- Alenius, P.; Myrberg, K.; Nekrasov, A. The physical oceanography of the Gulf of Finland: A review. Boreal Environ. Res. 1998, 3, 97–125. [Google Scholar]
- Ojaveer, E.; Pihy, E.; Saat, T. Fishes of Estonia; Estonian Academy Publishers: Tallin, Estonia, 2003; p. 416. [Google Scholar]
- Kudersky, L.A.; Shurukhin, A.S.; Popov, A.N.; Bogdanov, D.V.; Yakovlev, A.S. Fish population of the Neva river estuary. In Ecosystem of the Neva River Estuary: Biological Diversity and Ecological Problems; Alimov, A.F., Golubkov, S.M., Eds.; Association of Scientific Publications KMK: St. Petersburg/Moskow, Russia, 2008; pp. 223–240. (In Russian) [Google Scholar]
- Lappalainen, A.; Urho, L. Young-of-the-year fish species composition in small coastal bays in the northern Baltic Sea, surveyed with beach seine and small underwater detonations. Boreal Environ. Res. 2006, 11, 431–440. [Google Scholar]
- Remane, A. Die Brackwasserfauna. Zool. Anz. 1934, 7, 34–74. [Google Scholar]
- Khlebovich, V.V. Critical salinity and horohalinicum: Modern analysis of concepts. Tr. Zool. Inst. Acad. Sci. USSR 1989, 196, 5–11. (In Russian) [Google Scholar]
- Arroyo, N.L.; Bonsdorff, E. The Role of Drifting Algae for Marine Biodiversity. In Marine Macrophytes as Foundation Species; Olafsson, E., Ed.; Science Publisher/CRC Press: Boca Raton, FL, USA, 2016; pp. 100–129. [Google Scholar]
- Engström-Öst, J.; Immonen, E.; Candolin, U.; Mattila, J. The indirect effects of eutrophication on habitat choice and survival of fish larvae in the Baltic Sea. Mar. Biol. 2007, 151, 393–400. [Google Scholar] [CrossRef]
- Salovius, S.; Bonsdorff, E. Effects of depth, sediment and grazers on the degradation of drifting filamentous algae (Cladophora glomerata and Pilayella littoralis). J. Exp. Mar. Biol. Ecol. 2004, 298, 93–109. [Google Scholar] [CrossRef]
- Gubelit, Y.I. Biomass and Primary Production of Cladophora glomerata (L.) Kütz. in the Neva Estuary. Inland Water Biol. 2009, 2, 300–304. [Google Scholar] [CrossRef]
- Berezina, N.A.; Tsiplenkina, I.G.; Pankova, E.S.; Gubelit, J.I. Dynamics of invertebrate communities on the stony littoral of the Neva Estuary (Baltic Sea) under macroalgal blooms and bioinvasions. Transit. Waters Bull. 2007, 1, 65–76. [Google Scholar] [CrossRef]
- Johnson, D.A.; Welsh, B.L. Detrimental effects of Ulva lactuca (L.) exudates and low oxygen on estuarine crab larvae. J. Exp. Mar. Biol. Ecol. 1985, 86, 73–83. [Google Scholar] [CrossRef]
- Aneer, G. High natural mortality of Baltic herring (Clupea harengus) eggs caused by algal exudates? Mar. Biol. 1987, 94, 163–169. [Google Scholar] [CrossRef]
- Berezina, N.A.; Golubkov, S.M.; Gubelit, Y.I. Structure of Littoral Zoocenoses in the Macroalgae Zones of the Neva River Estuary. Inland Water Biol. 2009, 2, 340–347. [Google Scholar] [CrossRef]
- Gubelit, Y.I. Climatic impact on community of filamentous macroalgae in the Neva estuary (eastern Baltic Sea). Mar. Pollut. Bull. 2015, 91, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Alimov, A.F. Communities of the Freswater invertebrates among macrophytes associations. Proc. Zool. Inst. 1988, 186, 1–198. (In Russian) [Google Scholar]
- Demchuk, A.S.; Uspenskiy, A.A.; Golubkov, S.M. Abundance and feeding of fish in the coastal zone of the Neva Estuary, eastern Gulf of Finland. Boreal Environ. Res. 2021, 26, 1–16. [Google Scholar]
- Uspenskiy, A.A. Distribution and population characteristics of the invasive tubenose goby Proterorhinus marmoratus (Pallas, 1814) in the eastern Gulf of Finland. Proc. Zool. Inst. RAS 2020, 324, 459–475. [Google Scholar] [CrossRef]
- Pihl, L.; Isaksson, I.; Wennhage, H.; Moksnes, P.-O. Recent Increase of Filamentous Algae in Shallow Swedish Bays: Effects on the Community Structure of Epibenthic Fauna and Fish. Aquat. Ecol. 1995, 29, 349–358. [Google Scholar] [CrossRef]
- Von Nordheim, L.; Kotterba, P.; Moll, D.; Polte, P. Lethal effect of filamentous algal blooms on Atlantic herring (Clupea harengus) eggs in the Baltic Sea. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 1362–1372. [Google Scholar] [CrossRef]
- Rajasilta, M.; Mankki, J.; Ranta-Aho, K.; Vuorinen, I. Littoral fish communities in the Archipelago Sea, SW Finland: A preliminary study of changes over 20 years. Hydrobiologia 1999, 393, 253–260. [Google Scholar] [CrossRef]
- Rozas, L.R.; Odum, W.E. Occupation of submerged aquatic vegetation by fishes: Testing the roles of food and refuge. Oecologia 1988, 77, 101–106. [Google Scholar] [CrossRef]
- Lenanton, R.C.J.; Caputi, N. The roles of food supply and shelter in the relationship between fishes, in particular Cnidoglanis macrocephalus (Valenciennes), and detached macrophytes in the surf zone of sandy beaches. J. Exp. Mar. Biol. Ecol. 1989, 128, 165–176. [Google Scholar] [CrossRef]
- Korelyakova, I.L. Higher Aquatic Vegetation in the Eastern Part of the Gulf of Finland; GosNIORKh: St. Petersburg, Russia, 1997; p. 159. (In Russian) [Google Scholar]
- Zhakova, L.V. Macrophytes: Higher aquatic plants and macroalgae. In Ecosystem of the Neva River Estuary: Biodiversity and Ecological Problems; Alimov, A.F., Golubkov, S.M., Eds.; Association of Scientific Publications KMK: St. Petersburg/Moskow, Russia, 2008; pp. 105–125. (In Russian) [Google Scholar]
- Zhakova, L.V.; Drozdov, V.V.; Golubev, D.A. The impact of hydraulic engineering construction and soil storage in underwater marine dumps on coastal macrophyte thickets (on the example of the Neva Bay). In Basic Concepts of Modern Coastal Management. Vol. III. Assessment of the Effects of Natural and Anthropogenic Impacts on Coastal Ecosystems; RSHU: St. Petersburg, Russia, 2011; pp. 138–167. (In Russian) [Google Scholar]
- Sherstneva, O.A. Effect of water turbidity on the abundance and productivity of submerged macrophytes in the eastern coast of the Gulf of Finland. Collect. Sci. Pap. GosNIORKh 2006, 331, 12–36. (In Russian) [Google Scholar]
- Pekcan-Hekim, Z. Effects of Turbidity on Feeding and Distribution of Fish. Academic Ph.D. Dissertation, Department of Biological and Environmental Sciences, University of Helsinki, Helsinki, Finland, 8 June 2007. [Google Scholar]
- Kemp, P.; Sear, D.; Collins, A.; Naden, P.; Jones, I. The impacts of fine sediment on riverine fish. Hydrol. Process 2011, 25, 1800–1821. [Google Scholar] [CrossRef]
- Kjelland, M.E.; Woodley, C.M.; Swannack, T.M.; Smith, D.L. A review of the potential effects of suspended sediment on fishes: Potential dredging-related physiological, behavioral, and transgenerational implications. Environ. Syst. Decis. 2015, 35, 334–350. [Google Scholar] [CrossRef] [Green Version]
- Sukhacheva, L.L.; Orlova, M.I. On the application of the results of satellite observations of the eastern part of the Gulf of Finland to assess the impact of natural and anthropogenic factors on the state of the water area and biotic components of the ecosystem. Reg. Ecol. 2014, 1–2, 62–76. (In Russian) [Google Scholar]
- Almesjö, L.; Limén, H. Fish Populations in Swedish Waters. How Are They Influenced by Fishing, Eutrophication and Contaminants? The Riksdag Printing Office: Stockholm, Sweden, 2009; p. 80. [Google Scholar]
- Korpinen, S.; Meski, L.; Andersen, J.H.; Laamanen, M. Human pressures and their potential impact on the Baltic Sea ecosystem. Ecol. Indic. 2012, 15, 105–114. [Google Scholar] [CrossRef]
- Veneranta, L.; Urho, L. Assemblage Structure of Small Sized Fishes and Reflections from Differences in Abiotic Conditions. In Proceedings of the ICES CM 2007/G:15, ICES Annual Science Conference 2007, Helsinki, Finland, 17–21 September 2007; pp. 1–7. [Google Scholar]
- Petrov, O.V. (Ed.) Atlas of Geological and Ecological-Geological Maps of the Russian Sector of the Baltic Sea; VSEGEI: St. Petersburg, Russia, 2010; p. 78. (In Russian) [Google Scholar]






| Variable | Value | Description |
|---|---|---|
| Salinity ranges | 0 | <0.5‰; fresh water |
| 1 | 0.5–1.9‰; oligohaline barrier “δ-horogalinicum” | |
| 2 | 2.0–2.9‰; oligohaline water | |
| 3 | ≥3.0‰; oligohaline water | |
| Bottom substrate | 1 | pure sand |
| 2 | stones prevail | |
| 3 | mixed sand and stones | |
| Macrophytes | 0 | absent |
| 1 | submerged vegetation present | |
| 2 | semi-submerged and submerged vegetation are present and abundant | |
| Filamentous algae | 0 | absent |
| 1 | present, not abundant | |
| 2 | abundant | |
| Turbidity | 0 | clear water; bottom is visible in the deepest point of hauling, lack of suspended sediments |
| 1 | moderate turbidity; bottom is visible in the depth 0.5–1.0 m at least, suspended sediments are visible | |
| 2 | high turbidity; bottom is invisible in the depth around 0.5 m, abundant suspended sediments |
| Species | V | D | EG | LC | Occurrence in the Areas | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | ||||||
| Core species (V > 50 %) | |||||||||||
| 1 | Alburnus alburnus (Linnaeus, 1758) | 93.4 | 60.0 ± 16.5 | f | jv; ad | + | + | + | + | + | + |
| 2 | Rutilus rutilus (Linnaeus, 1758) | 79.5 | 21.4 ± 6.6 | f | jv; ad | + | + | + | + | + | + |
| 3 | Perca fluviatilis Linnaeus, 1758 | 70.9 | 18.4 ± 10.7 | f | jv; ad | + | + | + | + | + | + |
| 4 | Gobio gobio (Linnaeus, 1758) | 68.9 | 11.2 ± 6.8 | f | jv; ad | + | + | + | + | + | + |
| 5 | Gymnocephalus cernua (Linnaeus, 1758) | 55.0 | 4.9 ± 1.3 | f | jv; ad | + | + | + | + | + | |
| Secondary species (V = 25–49%) | |||||||||||
| 6 | Gasterosteus aculeatus Linnaeus, 1758 | 45.7 | 38.8 ± 22.1 | f;e | jv; ad | + | + | + | + | + | + |
| 7 | Blicca bjoerkna (Linnaeus, 1758) | 39.7 | 2.2 ± 0.4 | f | jv; ad | + | + | + | + | + | |
| 8 | Proterorhinus marmoratus (Pallas, 1814) * | 38.4 | 9.7 ± 3.0 | f;e | jv; ad | + | + | + | + | ||
| 9 | Abramis brama (Linnaeus, 1758) | 33.1 | 14.1 ± 10.3 | f | jv | + | + | + | + | ||
| 10 | Pungitius pungitius (Linnaeus, 1758) | 29.1 | 5.6 ± 1.9 | f | jv; ad | + | + | + | + | + | + |
| 11 | Leuciscus leuciscus (Linnaeus, 1758) | 27.2 | 1.2 ± 0.5 | f | jv; ad | + | + | + | + | ||
| 12 | Pomatoschistus microps (Kroyer, 1838) | 25.8 | 59.2 ± 22.4 | e | jv; ad | + | + | + | + | + | |
| Rare species (V = 8–24%) | |||||||||||
| 13 | Cobitis taenia Linnaeus, 1758 | 21.9 | 1.0 ± 0.3 | f | jv; ad | + | + | + | + | + | + |
| 14 | Scardinius erythrophthalmus (Linnaeus, 1758) | 17.9 | 2.2 ± 0.9 | f | jv; ad | + | + | + | + | ||
| 15 | Sander lucioperca (Linnaeus, 1758) | 12.6 | 5.2 ± 2.8 | f | jv | + | + | + | |||
| 16 | Romanogobio albipinnatus (Lukasch, 1933) * | 11.3 | < 1 | f | jv; ad | + | + | ||||
| 17 | Neogobius melanostomus (Pallas, 1814) * | 8.6 | 2.3 ± 1.0 | f;e | jv; ad | + | + | + | |||
| Sporadic species (V < 7%) | |||||||||||
| 18 | Osmerus eperlanus (Linnaeus, 1758) | 7.3 | 4.4 ± 2.9 | a | jv; ad | + | + | ||||
| 19 | Phoxinus phoxinus (Linnaeus, 1758) | 5.3 | 1.2 ± 0.3 | f | jv; ad | + | + | + | + | + | |
| 20 | Leucaspius delineatus (Heckel, 1843) | 5.3 | <1 | f | jv | + | + | + | |||
| 21 | Ammodytes tobianus Linnaeus, 1758 | 4.6 | 6.1 ± 1.7 | m | jv | + | + | ||||
| 22 | Coregonus albula (Linnaeus, 1758) | 4.6 | 1.0 ± 0.5 | a | jv | + | + | + | |||
| 23 | Leuciscus idus (Linnaeus, 1758) | 4.6 | <1 | f | jv | + | + | + | |||
| 24 | Vimba vimba (Linnaeus, 1758) | 4.6 | <1 | a | jv | + | + | + | |||
| 25 | Perccottus glenii Dybowski, 1877 * | 3.3 | 35.9 ± 14.9 | f | jv; ad | + | + | ||||
| 26 | Squalius cephalus (Linnaeus, 1758) | 3.3 | <1 | f | jv | + | + | + | |||
| 27 | Pomatoschistus minutus (Pallas, 1770) | 2.6 | <1 | o | jv; ad | + | + | ||||
| 28 | Clupea harengus membras Linnaeus, 1761 | 2.0 | 1.7 ± 1.2 | o | jv | + | + | + | |||
| 29 | Carassius gibelio (Bloch, 1782) * | 2.0 | <1 | f | jv | + | + | ||||
| 30 | Esox lucius Linnaeus, 1758 | 1.3 | <1 | f | jv | + | |||||
| 31 | Tinca tinca (Linnaeus, 1758) | 0.7 | 1.1 | f | jv | + | |||||
| 32 | Barbatula barbatula (Linnaeus, 1758) | 0.7 | <1 | f | jv | + | |||||
| 33 | Nerophis ophidion (Linnaeus, 1758) | 0.7 | <1 | m | ad | + | |||||
| 34 | Pelecus cultratus (Linnaeus, 1758) | 0.7 | <1 | f | jv | + | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uspenskiy, A.; Zhidkov, Z.; Levin, B. The Key Environmental Factors Shaping Coastal Fish Community in the Eastern Gulf of Finland, Baltic Sea. Diversity 2022, 14, 930. https://doi.org/10.3390/d14110930
Uspenskiy A, Zhidkov Z, Levin B. The Key Environmental Factors Shaping Coastal Fish Community in the Eastern Gulf of Finland, Baltic Sea. Diversity. 2022; 14(11):930. https://doi.org/10.3390/d14110930
Chicago/Turabian StyleUspenskiy, Anton, Zakhar Zhidkov, and Boris Levin. 2022. "The Key Environmental Factors Shaping Coastal Fish Community in the Eastern Gulf of Finland, Baltic Sea" Diversity 14, no. 11: 930. https://doi.org/10.3390/d14110930
APA StyleUspenskiy, A., Zhidkov, Z., & Levin, B. (2022). The Key Environmental Factors Shaping Coastal Fish Community in the Eastern Gulf of Finland, Baltic Sea. Diversity, 14(11), 930. https://doi.org/10.3390/d14110930