

Taxonomic Reassessment of Freshwater Mussels from the Western Balkans Reveals an Overlooked but Critical Refugium and Defines Conservation Priorities
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
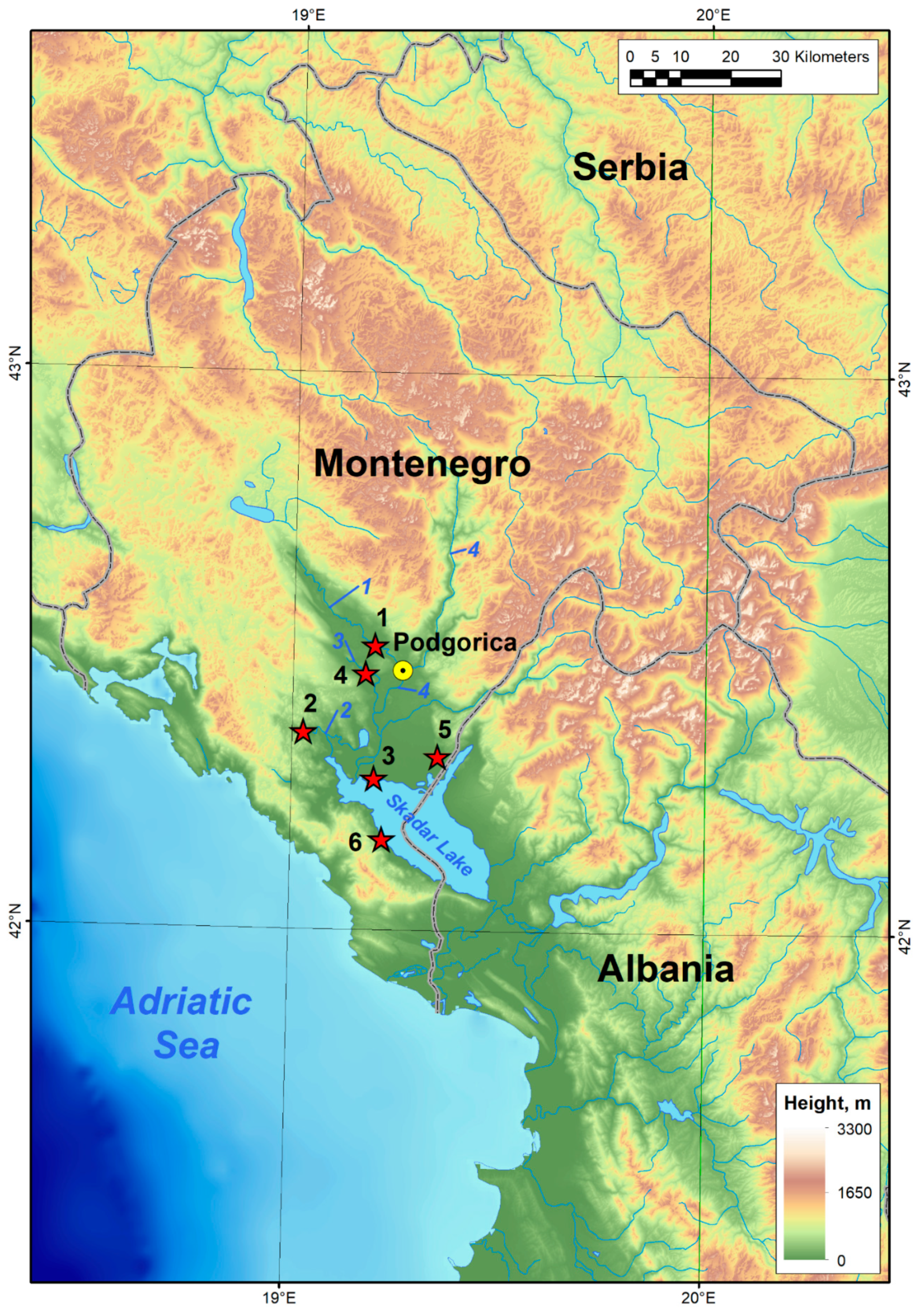
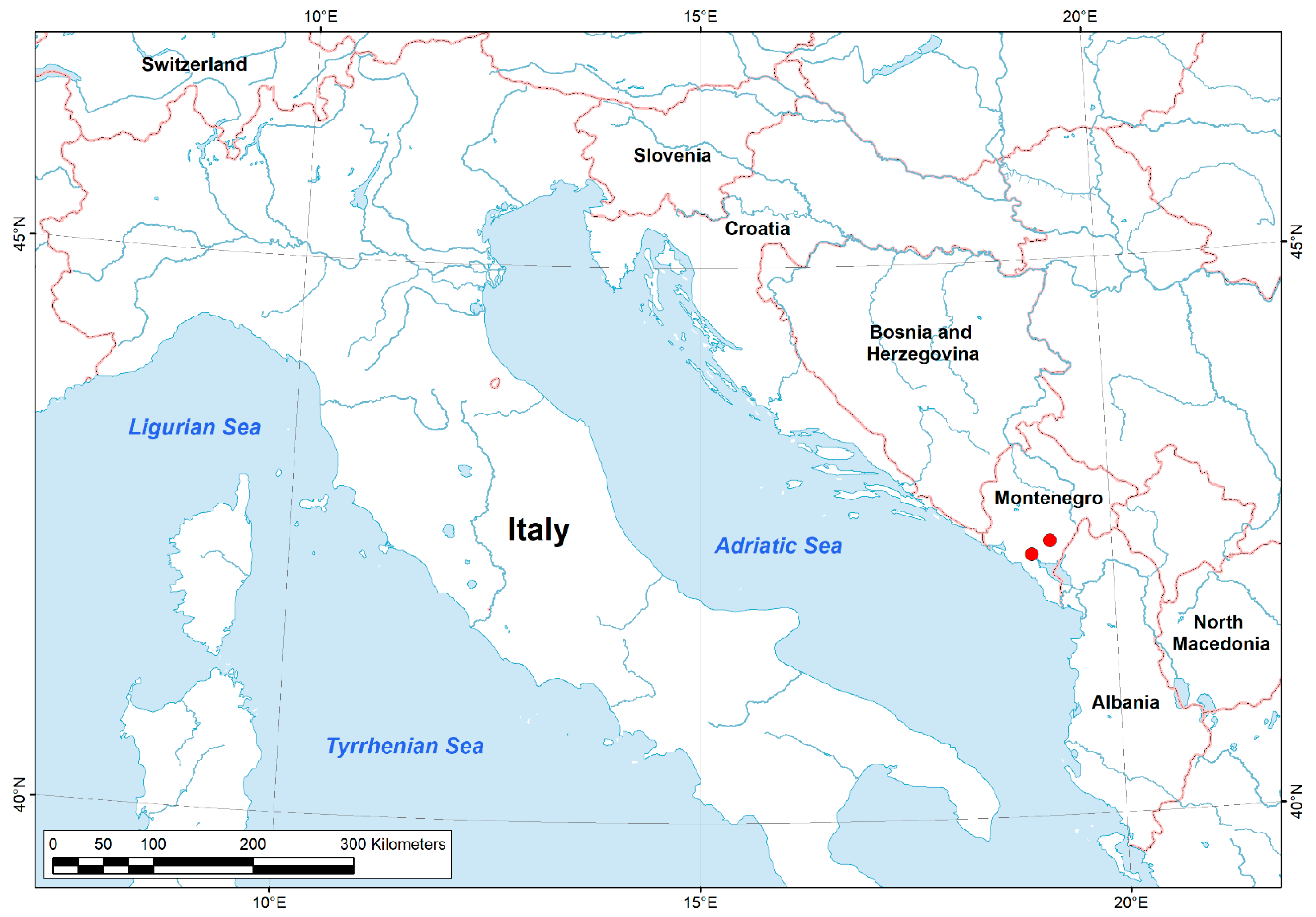
2.1. Data Sampling
2.2. DNA Extraction and Sequencing
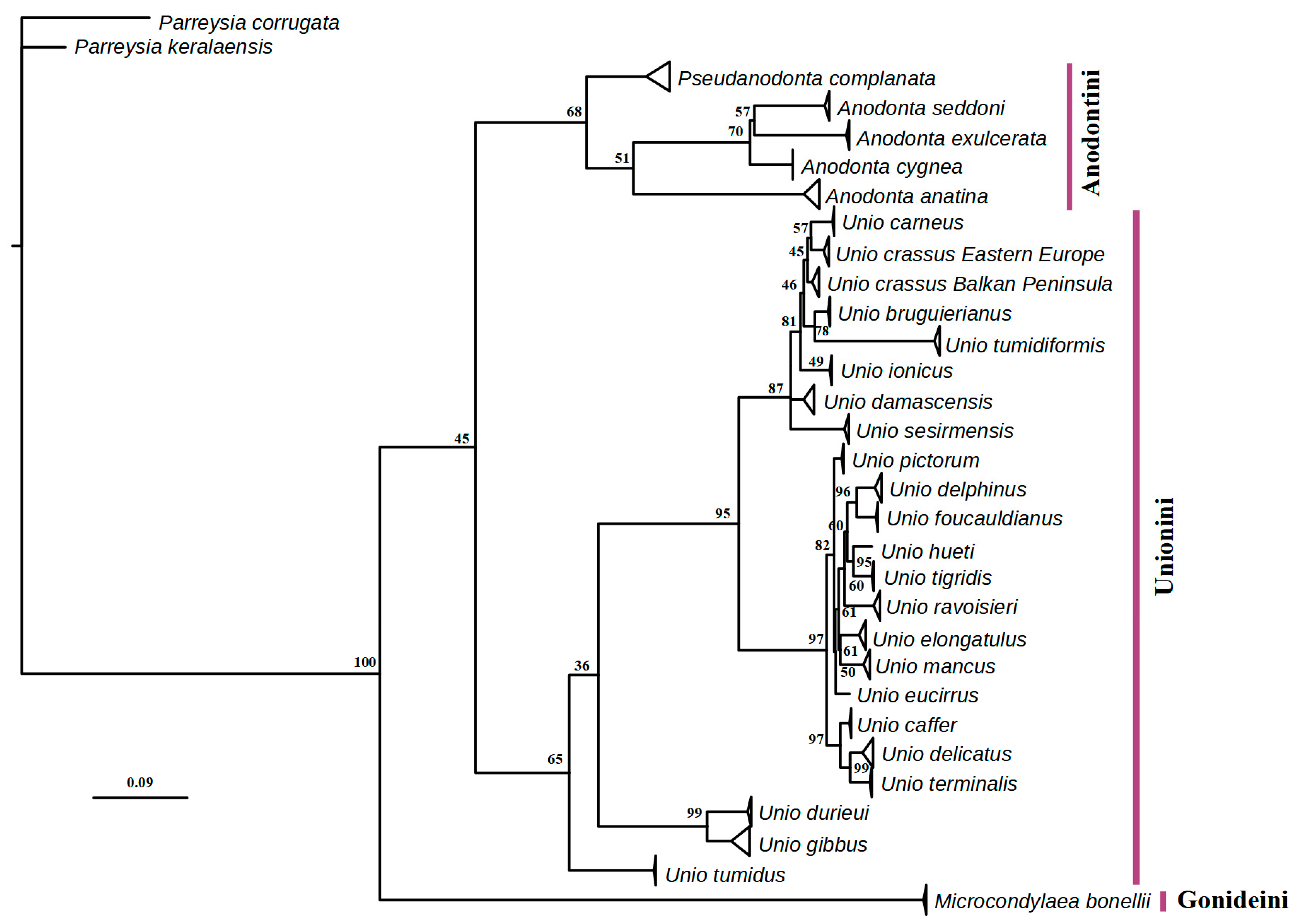
2.3. Phylogeographic and Phylogenetic Analyses
2.4. Distribution Mapping
3. Results
3.1. Species Richness of Freshwater Mussels in Montenegro
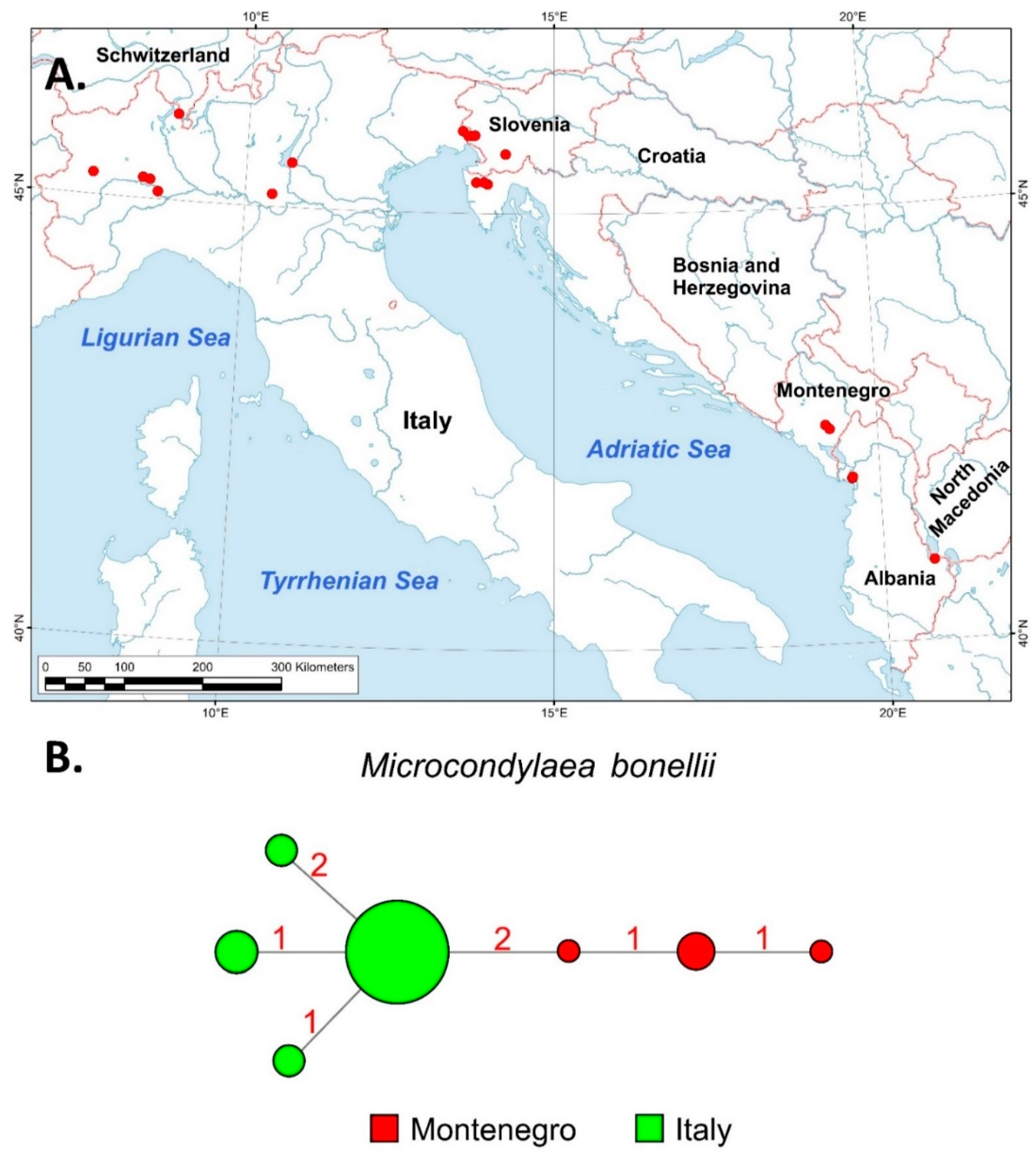
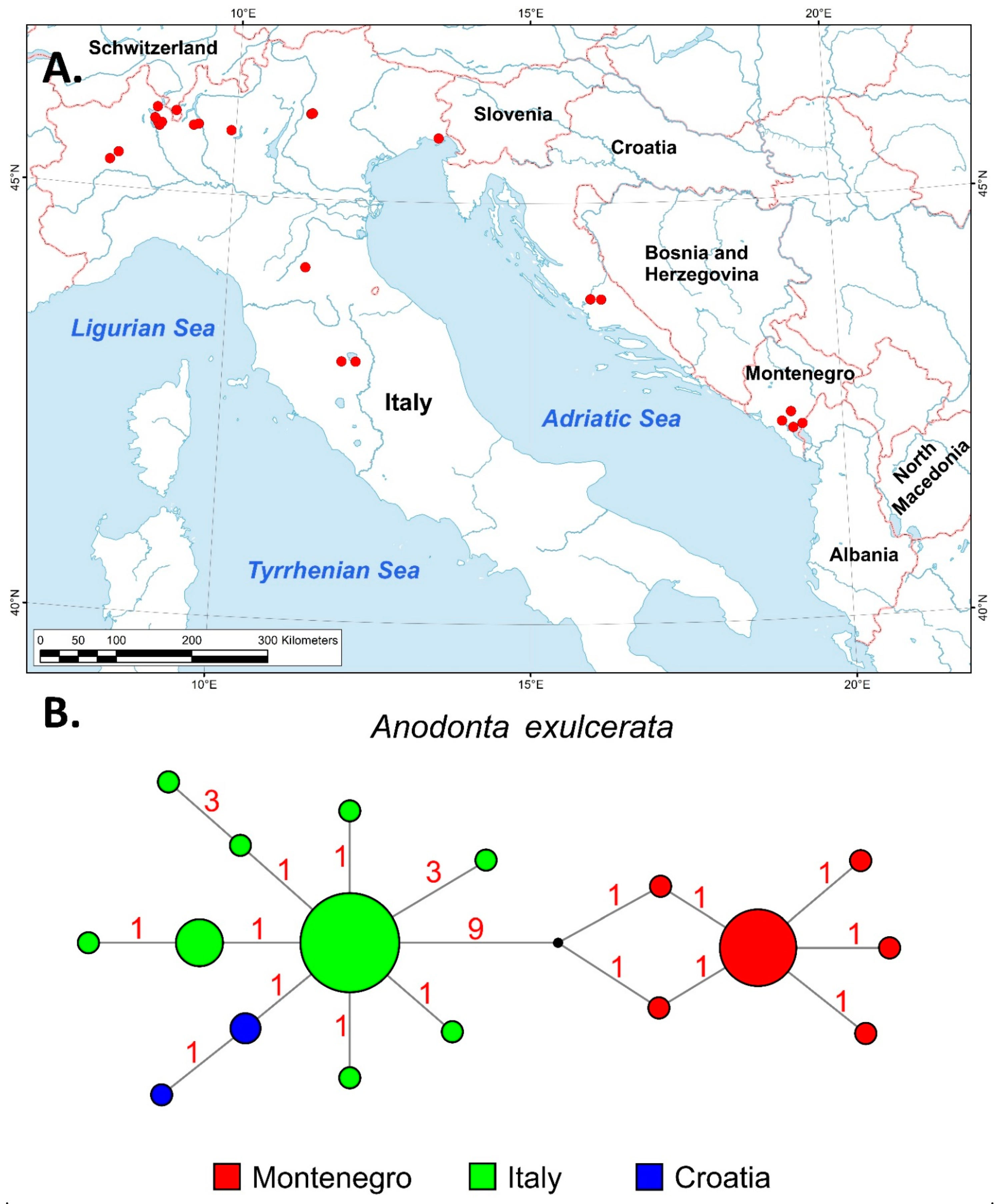
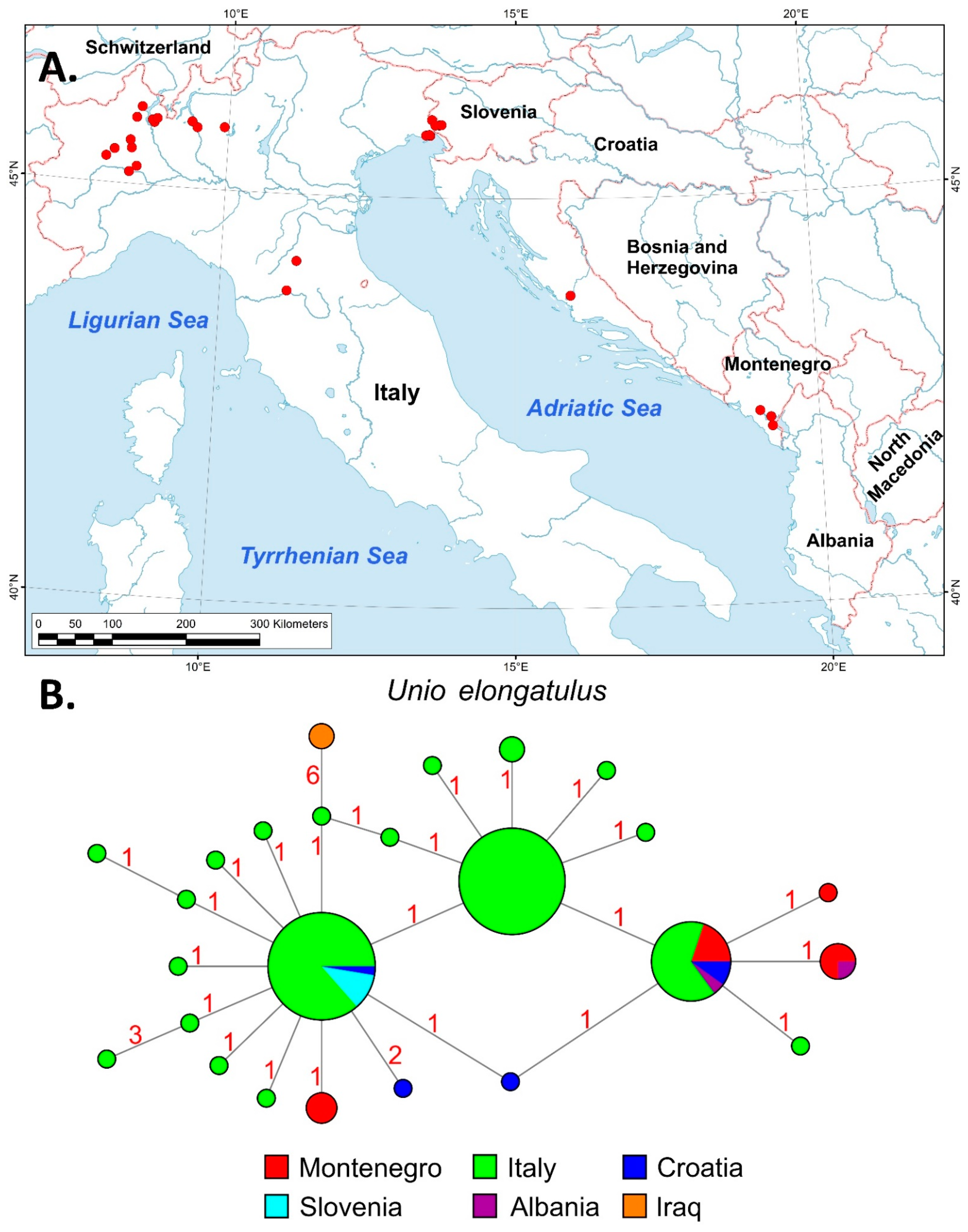
3.2. Phylogeographic and Distributional Patterns
4. Discussion
4.1. Taxonomic and Biogeographic Issues
4.2. Taxonomy
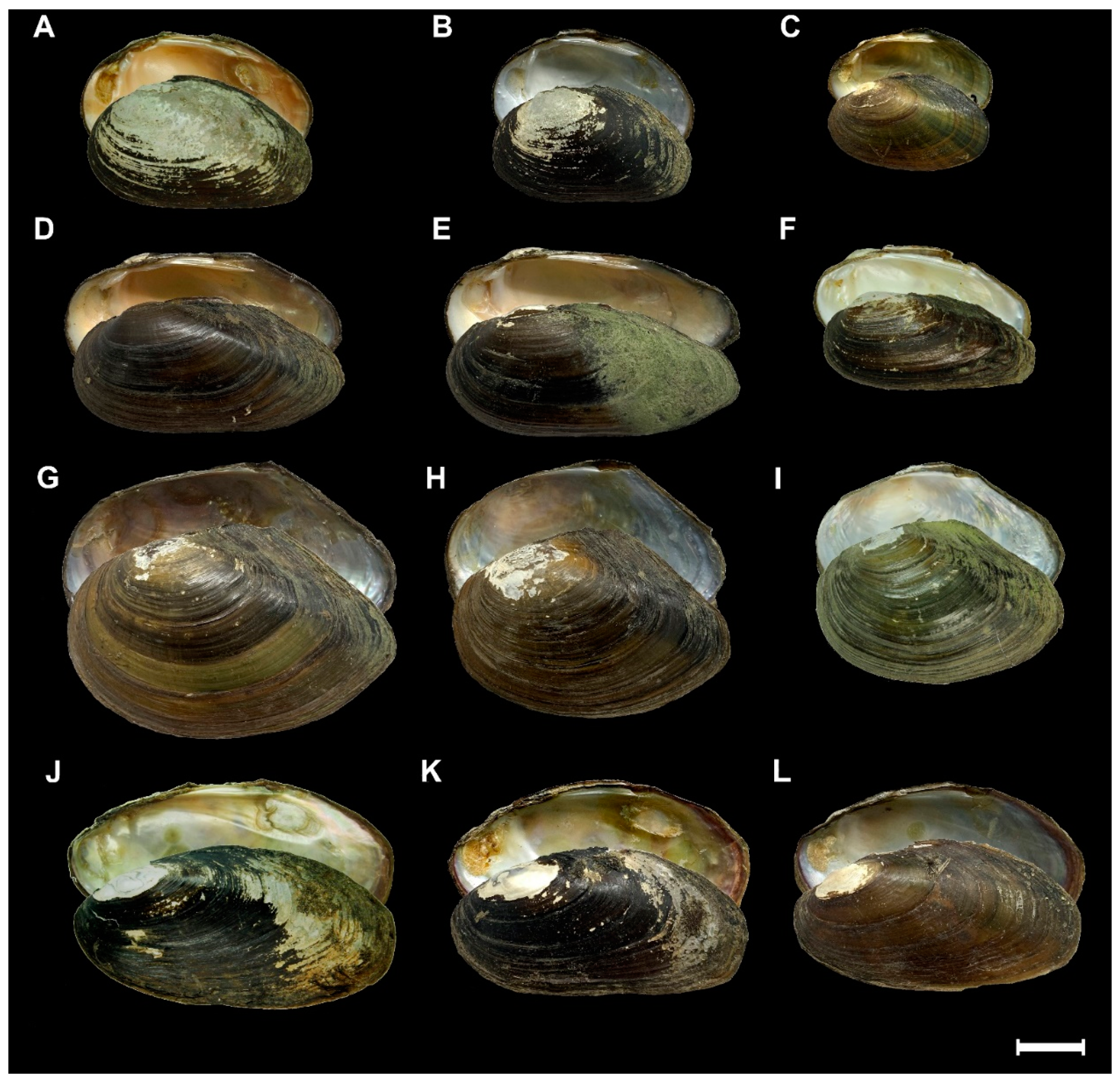
4.2.1. Unio elongatulus Pfeiffer, 1825
4.2.2. Unio carneus Küster, 1854 stat. rev.
4.2.3. Anodonta exulcerata Porro, 1838
4.2.4. Microcondylaea bonellii (Férussac, 1827)
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vaughn, C.; Hoellein, T.J. Bivalve impacts in marine and freshwater ecosystems. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 183–208. [Google Scholar] [CrossRef] [Green Version]
- Lopes-Lima, M.; Burlakova, L.E.; Karatayev, A.Y.; Mehler, K.; Seddon, M.; Sousa, R. Conservation of freshwater bivalves at the global scale: Diversity, threats and research needs. Hydrobiologia 2018, 810, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bolotov, I.N.; Makhrov, A.A.; Gofarov, M.; Aksenova, O.; Aspholm, P.E.; Bespalaya, Y.; Kabakov, M.B.; Kolosova, Y.S.; Kondakov, A.; Ofenböck, T.; et al. Climate warming as a possible trigger of keystone mussel population decline in oligotrophic rivers at the continental scale. Sci. Rep. 2018, 8, 35. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Rodríguez, N.; Akiyama, Y.B.; Aksenova, O.V.; Araujo, R.; Barnhart, M.C.; Bespalaya, Y.V.; Bogan, A.E.; Bolotov, I.N.; Budha, P.B.; Clavijo, C.; et al. Research priorities for freshwater mussel conservation assessment. Biol. Conserv. 2019, 231, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Prie, V.; Puillandre, N.; Bouchet, P. Bad taxonomy can kill: Molecular reevaluation of Unio mancus Lamarck, 1819 (Bivalvia: Unionidae) and its accepted subspecies. Knowl. Manag. Aquat. Ecosyst. 2012, 405, 8. [Google Scholar] [CrossRef] [Green Version]
- Douda, K.; Sell, J.; Kubíková-Peláková, L.; Horký, P.; Kaczmarczyk, A.; Mioduchowska, M. Host compatibility as a critical factor in management unit recognition: Population-level differences in mussel–fish relationships. J. Appl. Ecol. 2014, 51, 1085–1095. [Google Scholar] [CrossRef]
- Froufe, E.; Lopes-Lima, M.; Riccardi, N.; Zaccara, S.; Vanetti, I.; Lajtner, J.; Teixeira, A.; Varandas, S.; Prié, V.; Zieritz, A.; et al. Lifting the curtain on the freshwater mussel diversity from the Italian Peninsula and Croatian Adriatic coast. Biodivers. Conserv. 2017, 26, 3255–3274. [Google Scholar] [CrossRef] [Green Version]
- Tomilova, A.A.; Lyubas, A.A.; Kondakov, A.V.; Vikhrev, I.V.; Gofarov, M.Y.; Kolosova, Y.S.; Vinarski, M.V.; Palatov, D.M.; Bolotov, I.N. Evidence for Plio-Pleistocene Duck Mussel Refugia in the Azov Sea River Basins. Diversity 2020, 12, 118. [Google Scholar] [CrossRef] [Green Version]
- Mrkvicka, A.C. Populations of Microcondylaea bonellii (Férussac 1827), Unionidae—An European freshwater mussel at rapid decline and Unio mancus in Istria, Croatia. Ecol. Montenegrina 2018, 17, 53–59. [Google Scholar] [CrossRef]
- Govedič, M.; Govedič, T.B. The distribution of Microcondylaea bonellii (Bivalvia: Unionidae) in Slovenia. Nat. Slov. 2019, 21, 5–20. [Google Scholar]
- Serb, J.M. Discovery of genetically distinct sympatric lineages in the freshwater mussel Cyprogenia aberti (Bivalvia: Unionidae). J. Molluscan Stud. 2006, 72, 425–434. [Google Scholar] [CrossRef]
- Grabowski, M.; Bącela-Spychalska, K.; Pešić, V. Reproductive traits and conservation needs of the endemic gammarid Laurogammarus scutarensis from the Skadar Lake system, Balkan Peninsula. Limnologica 2014, 47, 44–51. [Google Scholar] [CrossRef]
- Grabowski, M.; Mamos, T.; Bącela-Spychalska, K.; Rewicz, T.; Wattier, R.A. Neogene paleogeography provides context for understanding the origin and spatial distribution of cryptic diversity in a widespread Balkan freshwater amphipod. PeerJ 2017, 5, e3016. [Google Scholar] [CrossRef] [Green Version]
- Jabłońska, A.; Wrzesińska, W.; Zawal, A.; Pešić, V.; Grabowski, M. Long-term within-basin isolation patterns, different conservation units, and interspecific mitochondrial DNA introgression in an amphipod endemic to the ancient Lake Skadar system, Balkan Peninsula. Freshw. Biol. 2020, 65, 209–225. [Google Scholar] [CrossRef]
- Glöer, P.; Pešić, V. The Bithynia species from Skadar Lake (Montenegro) (Gastropoda: Bithyniidae). Mollusca 2007, 25, 7–12. [Google Scholar]
- Pešić, V.; Glöer, P. A new freshwater snail genus (Hydrobiidae, Gastropoda) from Montenegro, with a discussion on gastropod diversity and endemism in Skadar Lake. ZooKeys 2013, 281, 69–90. [Google Scholar] [CrossRef]
- Sworobowicz, L.; Grabowski, M.; Mamos, T.; Burzyński, A.; Kilikowska, A.; Sell, J.; Wysocka, A. Revisiting the phylogeography of Asellus aquaticus in Europe: Insights into cryptic diversity and spatiotemporal diversification. Freshw. Biol. 2015, 60, 1824–1840. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Sousa, R.; Geist, J.; Aldridge, D.C.; Araujo, R.; Bergengren, J.; Bespalaya, Y.; Bódis, E.; Burlakova, L.; Van Damme, D.; et al. Conservation status of freshwater mussels in Europe: State of the art and future challenges. Biol. Rev. 2017, 92, 572–607. [Google Scholar] [CrossRef] [Green Version]
- Araujo, R.; Buckley, D.; Nagel, K.-O.; García-Jiménez, R.; Machordom, A. Species boundaries, geographic distribution and evolutionary history of the Western Palaearctic freshwater mussels Unio (Bivalvia: Unionidae). Zool. J. Linn. Soc. 2018, 182, 275–299. [Google Scholar] [CrossRef]
- Pešić, V.; Glöer, P. The Diversity and Conservation Status of the Molluscs of Lake Skadar/Shkodra. In The Skadar/Shkodra Lake Environment; Pešić, V., Karaman, G., Kostianoy, A., Eds.; The Handbook of Environmental Chemistry; Springer: Cham, Switzerland, 2018; Volume 80, pp. 295–310. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villesen, P. FaBox: An online toolbox for fasta sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.-T.; Von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Riccardi, N.; Froufe, E.; E Bogan, A.; Zieritz, A.; Teixeira, A.; Vanetti, I.; Varandas, S.; Zaccara, S.; Nagel, K.-O.; Lopes-Lima, M. Phylogeny of European Anodontini (Bivalvia: Unionidae) with a redescription of Anodonta exulcerata. Zool. J. Linn. Soc. 2020, 189, 745–761. [Google Scholar] [CrossRef]
- Sousa, R.; Bogan, A.E.; Gonçalves, D.V.; Lajtner, J.; Prié, V.; Riccardi, N.; Shumka, S.; Teixeira, A.; Urbańska, M.; Varandas, S.; et al. Microcondylaea bonellii as a new host for the European bitterling Rhodeus amarus. Knowl. Manag. Aquat. Ecosyst. 2020, 421, 4. [Google Scholar] [CrossRef] [Green Version]
- Froufe, E.; Bolotov, I.; Aldridge, D.C.; Bogan, A.E.; Breton, S.; Gan, H.M.; Lopes-Lima, M. Mesozoic mitogenome rearrangements and freshwater mussel (Bivalvia: Unionoidea) macroevolution. Heredity 2020, 124, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Bolotov, I.N.; Kondakov, A.V.; Konopleva, E.S.; Vikhrev, I.V.; Aksenova, O.V.; Aksenov, A.S.; Vinarski, M.V. Integrative taxonomy, biogeography and conservation of freshwater mussels (Unionidae) in Russia. Sci. Rep. 2020, 10, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomilova, A.A.; Kondakov, A.V.; Kisil, O.Y. Usage of transcribed spacers ITS1 and ITS2 for identification of freshwater mussels of the genera Anodonta and Pseudanodonta (Bivalvia: Unionidae: Anodontinae). Zhurnal Obs. Biol. 2019, 80, 364–371. [Google Scholar] [CrossRef]
- Kilikowska, A.; Mioduchowska, M.; Wysocka, A.; Kaczmarczyk-Ziemba, A.; Rychlińska, J.; Zając, K.; Sell, J. The patterns and puzzles of genetic diversity of endangered freshwater mussel Unio crassus Philipsson, 1788 populations from Vistula and Neman drainages (Eastern Central Europe). Life 2020, 10, 119. [Google Scholar] [CrossRef] [PubMed]
- Reis, J.; Machordom, A.; Araujo, R. Morphological and molecular diversity of Unionidae (Mollusca, Bivalvia) from Portugal. Graellsia 2013, 69, 17–36. [Google Scholar] [CrossRef]
- Khalloufi, N.; Toledo, C.; Machordom, A.; Boumaïza, M.; Araujo, R. The unionids of Tunisia: Taxonomy and phylogenetic relationships, with redescription of Unio ravoisieri Deshayes, 1847 and U. durieui Deshayes, 1847. J. Molluscan Stud. 2011, 77, 103–115. [Google Scholar] [CrossRef]
- Araujo, R.; Gómez, I.; Machordom, A. The identity and biology of Unio mancus Lamarck, 1819 (= U. elongatulus) (Bivalvia: Unionidae) in the Iberian Peninsula. J. Molluscan Stud. 2005, 71, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Graf, D.L.; Foighil, D.Ó. The evolution of brooding characters among the freshwater pearly mussels (Bivalvia: Unionoidea) of North America. J. Molluscan Stud. 2000, 66, 157–170. [Google Scholar] [CrossRef] [Green Version]
- Bolotov, I.N.; Pasupuleti, R.; Subba Rao, N.V.; Unnikrishnan, S.K.; Chan, N.; Lunn, Z.; Nagel, K.O. Oriental freshwater mussels arose in East Gondwana and arrived to Asia on the Indian Plate and Burma Terrane. Sci. Rep. 2022, 12, 1–26. [Google Scholar] [CrossRef]
- The North Carolina Museum of Natural Sciences’ Mollusks Collection (NCSM Mollusk Collection). Available online: https://collections.naturalsciences.org/search/mollusks (accessed on 10 June 2022).
- The Senckenberg Research Institute and Natural History Museum (SMF Collection). Available online: https://www.senckenberg.de/en/institutes/senckenberg-research-institute-natural-history-museum-frankfurt/ (accessed on 10 June 2022).
- Musée d’histoire Naturelle, Neuchâtel, Switzerland (MHNN Collection). Available online: https://www.museum-neuchatel.ch/ (accessed on 10 June 2022).
- Biologiezentrum Linz Oberoesterreich. Available online: https://www.ooekultur.at/location-detail/biologiezentrum (accessed on 10 June 2022).
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; Dewaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. London Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes-Lima, M.; Gürlek, M.E.; Kebapçı, Ü.; Şereflişan, H.; Yanık, T.; Mirzajani, A.; Neubert, E.; Prié, V.; Teixeira, A.; Gomes-Dos-Santos, A.; et al. Diversity, biogeography, evolutionary relationships, and conservation of Eastern Mediterranean freshwater mussels (Bivalvia: Unionidae). Mol. Phylogenet. Evol. 2021, 163, 107261. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, M.; Jabłońska, A.; Wysocka, A.; Pešić, V. The Obscure History of the Lake Skadar and Its Biota: A Perspective for Future Research. In The Skadar/Shkodra Lake Environment; Pešić, V., Karaman, G., Kostianoy, A., Eds.; The Handbook of Environmental Chemistry; Springer International Publishing AG: Cham, Switzerland, 2018; Volume 80, pp. 47–61. [Google Scholar] [CrossRef]
- Froufe, E.; Gonçalves, D.V.; Teixeira, A.; Sousa, R.; Varandas, S.; Ghamizi, M.; Zieritz, A.; Lopes-Lima, M. Who lives where? Molecular and morphometric analyses clarify which Unio species (Unionida, Mollusca) inhabit the southwestern Palearctic region. Org. Divers. Evol. 2016, 16, 597–611. [Google Scholar] [CrossRef] [Green Version]
- Tomilova, A.A. Morphological Variability and Phylogeography of the Duck Mussel Anodonta anatina in Russia and Adjacent Territories. Ph.D. Thesis, Papanin Institute for Biology of Inland Waters RAS, Borok, Russia, 2021. [Google Scholar]
- Marrone, F.; Nardi, G.; Cianfanelli, S.; Govedič, M.; Barra, S.A.; Arculeo, M.; Bodon, M. Diversity and taxonomy of the genus Unio Philipsson in Italy, with the designation of a neotype for Unio elongatulus C. Pfeiffer (Mollusca, Bivalvia, Unionidae). Zootaxa 2019, 4545, 339–374. [Google Scholar] [CrossRef]
- Mladineo, I.; Poljak, V. Ecology and genetic structure of zoonotic Anisakis spp. from Adriatic commercial fish species. Appl. Environ. Microbiol. 2014, 80, 1281–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.K.; Joshi, P.C.; Joshi, B.D. Molecular data suggest population expansion and high level of gene flow in the Plain Tiger (Danaus chrysippus; Nymphalidae: Danainae). Mitochondrial DNA B 2018, 3, 707–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa, C.E.; Acosta, D.B.; Mac Allister, M.E.; Merele, M.; Fernández, G.P.; Carpinetti, B.N.; Winter, M.; Abate, S.; Barandiaran, S.; Merino, M.L. Patterns of genetic variation on wild pig (Sus scrofa) populations over a complete range of the species in Argentina. Mammalia 2022, 86, 359–372. [Google Scholar] [CrossRef]
- Jenkins, T.L.; Castilho, R.; Stevens, J.R. Meta-analysis of northeast Atlantic marine taxa shows contrasting phylogeographic patterns following post-LGM expansions. PeerJ 2018, 6, e5684. [Google Scholar] [CrossRef] [Green Version]
- Wohlberedt, O. Zur Fauna Montenegros und Nordalbaniens (Mollusken, Käfer, Isopoden, Chilopoden, Diplopoden); Wissenschaftliche Mitteilungen aus Bosnien und der Herzegowina; Adolf Holzhausen: Vienna, Austria, 1909; Volume 11, pp. 119–124. [Google Scholar]
- Inoue, K.; Berg, D.J. Predicting the effects of climate change on population connectivity and genetic diversity of an imperiled freshwater mussel, Cumberlandia monodonta (Bivalvia: Margaritiferidae), in riverine systems. Glob. Change Biol. 2017, 23, 94–107. [Google Scholar] [CrossRef]
- Pešić, V.; Paunović, M.; Kostianoy, A.G. (Eds.) The Rivers of Montenegro; The Handbook of Environmental Chemistry; Springer: Cham, Switzerland, 2019; Volume 93, pp. 1–12. [Google Scholar] [CrossRef]
- Riccardi, N.; Bo, T.; Bogan, A.E.; Douda, K.; Froufe, E.; Gonçalves, D.V.; Lajtner, J.; Modesto, V.; Moro, G.A.; Prié, V.; et al. Microcondylaea bonellii, a testimonial for neglected endangered species. In Imperiled: The Encyclopedia of Conservation; DellaSala, D.A., Goldstein, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1–11. [Google Scholar] [CrossRef]
- Prié, V.; Puillandre, N. Molecular phylogeny, taxonomy, and distribution of French Unio species (Bivalvia, Unionidae). Hydrobiologia 2014, 735, 95–110. [Google Scholar] [CrossRef]
- Riccardi, N.R.; Froufe, E.; Lopes-Lima, M.; Mazzoli, C. When and how? Freshwater mussel recolonisation in Lake Orta. J. Limnol. 2016, 75, 120–130. [Google Scholar] [CrossRef]
- Küster, H.C. Die Flussperlmuscheln (Unio et Hyria). Systematisches Conchylien-Cabinet von Martini und Chemnitz Fortgesetzt von Hofrath Dr. G. v. Schubert und Professor Dr. J.A. Wagner 1854, 9, 89–120. [Google Scholar]
- Clessin, S. Genus Anodonta Cuv. Systematisches Conchylien-Cabinet von Martini und Chemnitz Fortgesetzt von Hofrath Dr. G. v. Schubert und Professor Dr. J.A. Wagner 1873, 9, 65–287. [Google Scholar]
- Albrecht, C.; Bodon, M.; Cianfanelli, S.; Giusti, F.; Manganelli, G. Microcondylaea bonellii; The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland; Cambridge, UK, 2011; p. e.T155595A4805631. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Description of Sampling Sites | Coordinates | Number of Collected Specimens | |
|---|---|---|---|---|
| Latitude | Longitude | |||
| 1 | Lower part of the Zeta River at Vranjske Njive (Podgorica) | 42°28′6” N | 19°15′29” E | 4 |
| 2 | Zeta River near Spuž town (Danilovgrad) | 42°30′41” N | 19°11′55” E | 12 |
| 3 | Crnojevića River near River Crnojevića village | 42°21′16” N | 19°1′40” E | 23 |
| 4 | Lower part of the Plavnica River near its mouth into Skadar Lake | 42°16′20” N | 19°12′3” E | 13 |
| 5 | Skadar Lake near Donji Murići village | 42°9′48” N | 19°13′18” E | 3 |
| 6 | Skadar Lake near sublacustrine spring at Podhum | 42°18′48” N | 19°21′13” E | 6 |
| 7 | Podgorica, Sitnica River, 800 m northeast of Ćafa village | 42°27′39” N | 19°10′37” E | 10 |
| No | Species | Sampling Sites | ||||||
|---|---|---|---|---|---|---|---|---|
| Zeta River Near Vranjske Njive | Zeta River at Spuž | Crnojevića River | Skadar Lake (Donji Murići Village) | Plavnica River | Skadar Lake (Podhum) | Sitnica River | ||
| 1 | Unio carneus | 4 | 6 | 4 | - | - | - | - |
| 2 | Unio elongatulus | - | - | 10 | 3 | 3 | - | - |
| 3 | Microcondylaea bonellii | - | 6 | - | - | - | - | - |
| 4 | Anodonta exulcerata | - | - | 9 | - | 10 | 6 | 10 |
| Number of species | 1 | 2 | 3 | 1 | 2 | 1 | 1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyubas, A.A.; Kondakov, A.V.; Tomilova, A.A.; Gofarov, M.Y.; Eliseeva, T.A.; Konopleva, E.S.; Vikhrev, I.V.; Yunitsyna, O.A.; Pešić, V.; Bolotov, I.N. Taxonomic Reassessment of Freshwater Mussels from the Western Balkans Reveals an Overlooked but Critical Refugium and Defines Conservation Priorities. Diversity 2022, 14, 935. https://doi.org/10.3390/d14110935
Lyubas AA, Kondakov AV, Tomilova AA, Gofarov MY, Eliseeva TA, Konopleva ES, Vikhrev IV, Yunitsyna OA, Pešić V, Bolotov IN. Taxonomic Reassessment of Freshwater Mussels from the Western Balkans Reveals an Overlooked but Critical Refugium and Defines Conservation Priorities. Diversity. 2022; 14(11):935. https://doi.org/10.3390/d14110935
Chicago/Turabian StyleLyubas, Artem A., Alexander V. Kondakov, Alena A. Tomilova, Mikhail Yu. Gofarov, Tatyana A. Eliseeva, Ekaterina S. Konopleva, Ilya V. Vikhrev, Olesya A. Yunitsyna, Vladimir Pešić, and Ivan N. Bolotov. 2022. "Taxonomic Reassessment of Freshwater Mussels from the Western Balkans Reveals an Overlooked but Critical Refugium and Defines Conservation Priorities" Diversity 14, no. 11: 935. https://doi.org/10.3390/d14110935
APA StyleLyubas, A. A., Kondakov, A. V., Tomilova, A. A., Gofarov, M. Y., Eliseeva, T. A., Konopleva, E. S., Vikhrev, I. V., Yunitsyna, O. A., Pešić, V., & Bolotov, I. N. (2022). Taxonomic Reassessment of Freshwater Mussels from the Western Balkans Reveals an Overlooked but Critical Refugium and Defines Conservation Priorities. Diversity, 14(11), 935. https://doi.org/10.3390/d14110935