
The Foraging Window for Greater White-Fronted Geese (Anser albifrons) Is Consistent with the Growth Stage of Carex


 and
and
Abstract
:1. Introduction
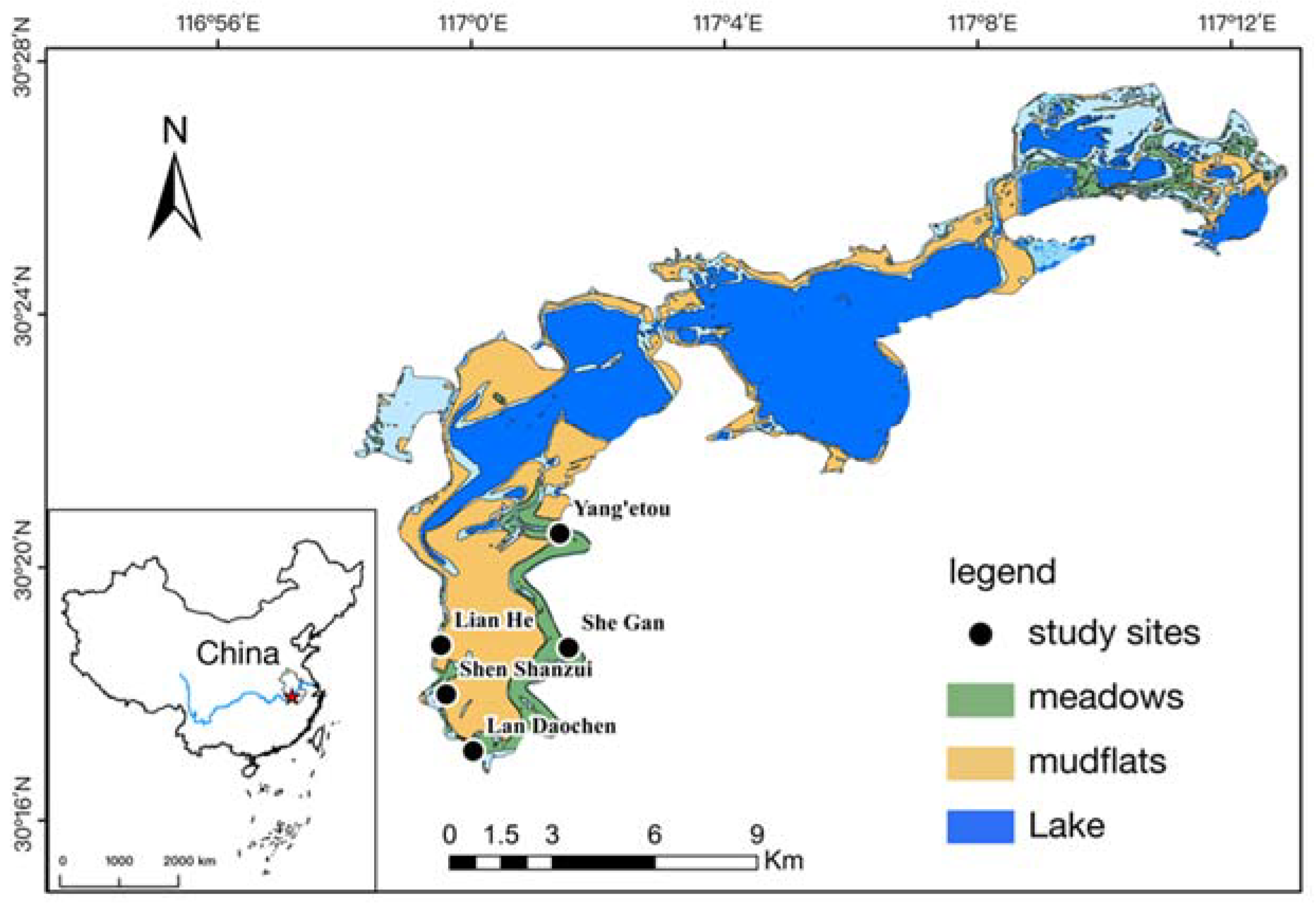
2. Materials and Methods
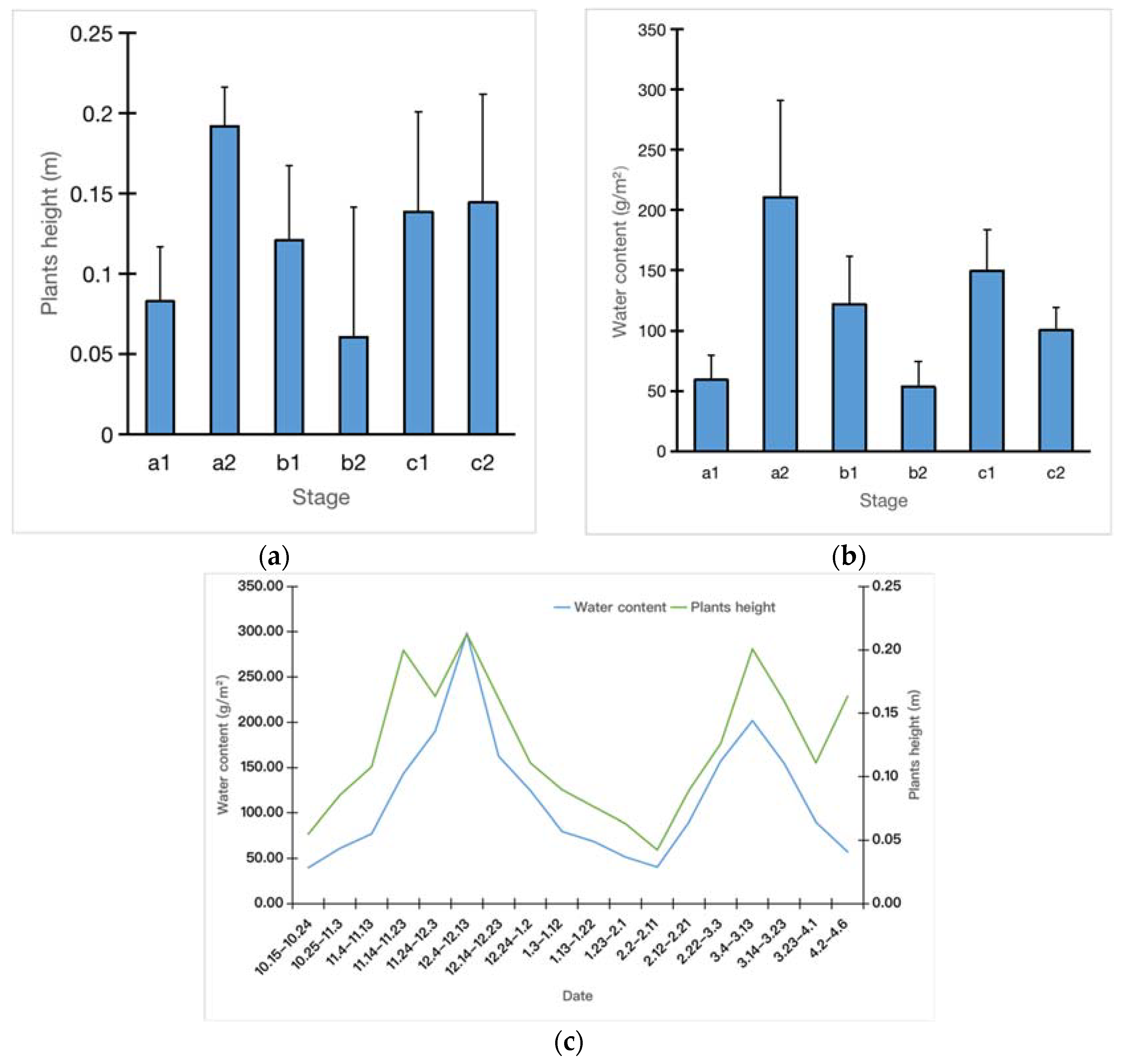
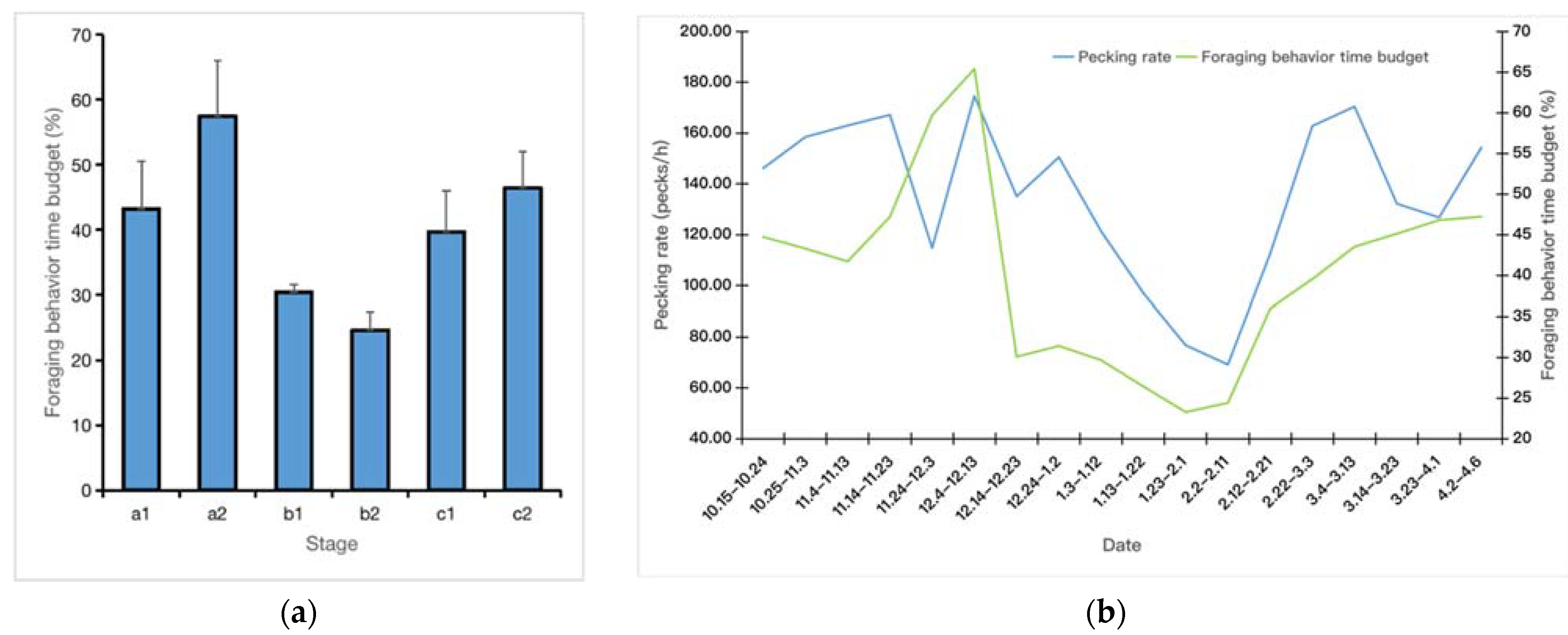
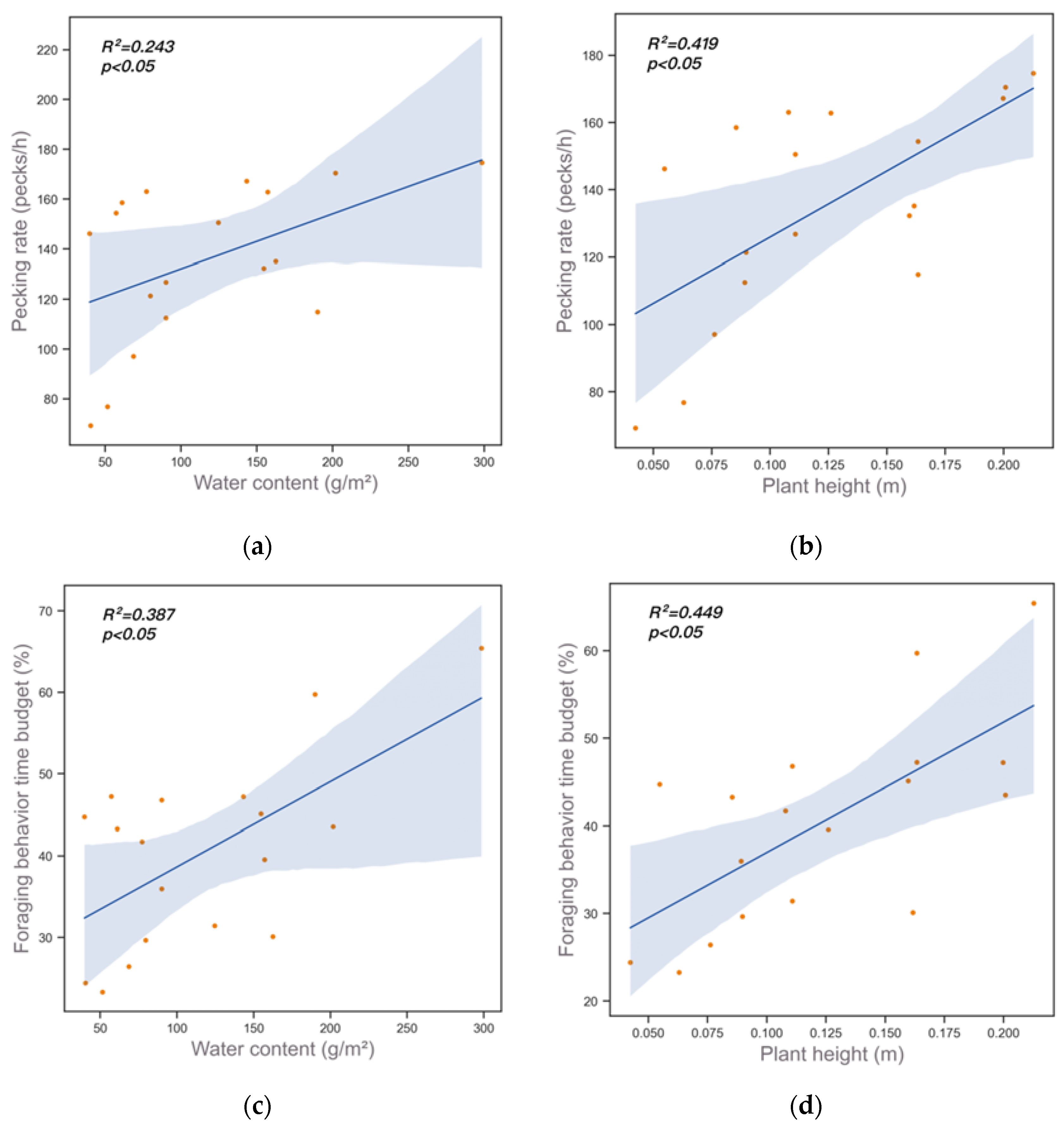
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Charnov, E.L. Optimal foraging, the marginal value theorem. Theor. Popul. Biol. 1976, 9, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Zhou, L.; Cheng, L.; Song, Y.; Xu, W. Foraging behavior of the greater white-fronted goose (Anser albifrons) wintering at Shengjin Lake: Diet shifts and habitat use. Avian Res. 2020, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Beerens, J.M.; Gawlik, D.E.; Herring, G.; Cook, M.I. Dynamic habitat selection by two wading bird species with divergent foraging strategies in a seasonally fluctuating wetland. The Auk 2011, 128, 651–662. [Google Scholar] [CrossRef]
- Říha, M.; Prchalová, M. Models of animal distributions in inland waters. In Encyclopedia of Inland Waters; Elsevier: Amsterdam, The Netherlands, 2022; pp. 292–301. ISBN 978-0-12-822041-2. [Google Scholar]
- Durant, D.; Fritz, H.; Blais, S.; Duncan, P. The functional response in three species of herbivorous Anatidae: Effects of sward height, body mass and bill size. J. Anim. Ecology 2003, 72, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Cumming, G.S.; Paxton, M.; King, J.; Beuster, H. Foraging guild membership explains variation in waterbird responses to the hydrological regime of an arid-region flood-pulse river in Namibia: Community dynamics of waterbirds in Namibia. Freshw. Biol. 2012, 57, 1202–1213. [Google Scholar] [CrossRef]
- Falk, K.; Benvenuti, S.; Dall’Antonia, L.; Kampp, K.; Ribolini, A. Time allocation and foraging behavior of chick-rearing Brünnich’s Guillemots Uria Lomvia in high-arctic greenland. Ibis 2008, 142, 82–92. [Google Scholar] [CrossRef]
- Durant, D.; Fritz, H. Variation of pecking rate with sward height in wild wigeon Anas Penelope. J. Ornithol. 2006, 147, 367–370. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, L.; Cheng, L.; Song, Y. Water level management plan based on the ecological demands of wintering waterbirds at Shengjin Lake. Glob. Ecol. Conserv. 2021, 27, e01567. [Google Scholar] [CrossRef]
- Hazra, P.; Sinha, A.; Mondal, P.; Khan, T.N. Calendar-effects and temperature-impacts in migratory waterbirds at three tropical Indian wetlands. Acta Oecologica 2012, 43, 60–71. [Google Scholar] [CrossRef]
- Saino, N.; Szep, T.; Romano, M.; Rubolini, D.; Spina, F.; Moller, A.P. Ecological conditions during winter predict arrival date at the breeding quarters in a trans-saharan migratory bird. Ecol. Lett. 2004, 7, 21–25. [Google Scholar] [CrossRef]
- Meng, Z.; Xia, S.; Yu, X.; Rao, D.; JIN, B. A study on the suitable time window of feeding vegetation fit for overwintering geese in Poyang Lake. Acta Ecol. Sin. 2018, 38, 7539–7548. [Google Scholar] [CrossRef]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S. Resource availability and plant antiherbivore defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilmshurst, J.F.; Fryxell, J.M.; Hudsonb, R.J. Forage quality and patch choice by Wapiti (Cervus elaphus). Behav. Ecol. 1995, 6, 209–217. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, J.; Xu, W.; Ye, X. Effects of waterbird herbivory on dominant perennial herb Carex thunbergii in Shengjin Lake. Diversity 2022, 14, 331. [Google Scholar] [CrossRef]
- Guan, L.; Wen, L.; Feng, D.; Zhang, H.; Lei, G. Delayed flood recession in central Yangtze floodplains can cause significant food shortages for wintering geese: Results of inundation experiment. Environ. Manag. 2014, 54, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yang, Y.; Wang, Z.; Yang, L.; Zhang, D.; Zhou, L. The relationship between seasonal water level fluctuation and habitat availability for wintering waterbirds at Shengjin Lake, China. Bird Conserv. Int. 2019, 29, 100–114. [Google Scholar] [CrossRef]
- Yu, C.; Zhou, L.; Mahtab, N.; Fan, S.; Song, Y. The influence of food density, flock size, and disturbance on the functional response of Bewick’s Swans (Cygnus columbianus bewickii) in wintering habitats. Animals 2019, 9, 946. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhou, L.Z.; Xu, W. Diversity of wintering waterbirds enhanced by restoring aquatic vegetation at Shengjin Lake, China. Sci. Total Environ. 2020, 737, 140190. [Google Scholar] [CrossRef]
- Zheng, M.; Zhou, L.; Zhao, N.; Xu, W. Effects of variation in food resources on foraging habitat use by wintering hooded cranes (Grus monacha). Avian Res. 2015, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-X.; Lu, J.-J. Feeding ecology of two wintering geese species at Poyang Lake, China. J. Freshw. Ecol. 1999, 14, 439–445. [Google Scholar] [CrossRef]
- Zhao, M.; Cong, P.; Barter, M.; Fox, A.D.; Cao, L. The changing abundance and distribution of greater white-fronted geese Anser albifrons in the Yangtze River Floodplain: Impacts of recent hydrological changes. Bird Conserv. Int. 2012, 22, 135–143. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhao, Q.; Solovyeva, D.; Lameris, T.; Batbayar, N.; Bysykatova-Harmey, I.; Lee, H.; Emelyanov, V.; Rozenfeld, S.B.; Park, J.; et al. Population trends and migration routes of the east Asian bean goose Anser fabalis middendorffii and A. f. serrirostris. Wildfowl 2020, 6, 124–156. [Google Scholar]
- Lehner, P.N. Sampling methods in behavior research. Poult. Sci. 1992, 71, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jia, Y.; Guan, L.; Lu, C.; Lei, G.; Wen, L.; Liu, G. Optimising hydrological conditions to sustain wintering waterbird populations in Poyang Lake National Natural Reserve: Implications for dam operations. Freshw. Biol. 2013, 58, 2366–2379. [Google Scholar] [CrossRef]
- Bosholn, M.; Anciães, M. Focal animal sampling. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–3. ISBN 978-3-319-47829-6. [Google Scholar]
- Li, C.; Zhou, L.; Xu, L.; Zhao, N.; Beauchamp, G. Vigilance and activity time-budget adjustments of wintering hooded cranes, Grus monacha, in human-dominated foraging habitats. PLoS ONE 2015, 10, e0118928. [Google Scholar] [CrossRef]
- Wan, W.; Zhou, L.; Song, Y. Shifts in foraging behavior of wintering hooded cranes (Grus monacha) in three different habitats at Shengjin Lake, China. Avian Res. 2016, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Meng, Q.; Li, X.; Yuan, Y.; Peng, Y.; Li, X.; Li, S.; Zhu, Z.; Yan, M. The influence of hydrological variables, climatic variables and food availability on Anatidae in interconnected river-lake systems, the middle and lower reaches of the Yangtze River Floodplain. Sci. Total Environ. 2021, 768, 144534. [Google Scholar] [CrossRef]
- Li, Y.; Zhong, Y.; Shao, R.; Yan, C.; Jin, J.; Shan, J.; Li, F.; Ji, W.; Bin, L.; Zhang, X.; et al. Modified hydrological regime from the three gorges dam increases the risk of food shortages for wintering waterbirds in Poyang Lake. Glob. Ecol. Conserv. 2020, 24, e01286. [Google Scholar] [CrossRef]
- Yang, M.; Xia, S.; Liu, G.; Wang, M.; Ding, Z.; Yu, P.; Tang, X. Effect of hydrological variation on vegetation dynamics for wintering waterfowl in China’s Poyang Lake wetland. Glob. Ecol. Conserv. 2020, 22, e01020. [Google Scholar] [CrossRef]
- Durant, D.; Fritz, H.; Duncan, P. Feeding patch selection by herbivorous Anatidae: The influence of body size, and of plant quantity and quality. J. Avian Biol. 2004, 35, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Alerstam, T.; Hedenström, A.; Åkesson, S. Long-distance migration: Evolution and determinants. Oikos 2003, 103, 247–260. [Google Scholar] [CrossRef]
- Bayly, N.J.; Rosenberg, K.V.; Easton, W.E.; Gomez, C.; Carlisle, J.; Ewert, D.N.; Drake, A.; Goodrich, L. Major stopover regions and migratory bottlenecks for Nearctic-Neotropical land birds within the Neotropics: A review. Bird Conserv. Int. 2018, 28, 1–26. [Google Scholar] [CrossRef] [Green Version]
- La Sorte, F.A.; Fink, D. Migration distance, ecological barriers and en-route variation in the migratory behavior of terrestrial bird populations: Migratory behavior of terrestrial bird populations. Glob. Ecol. Biogeogr. 2017, 26, 216–227. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Foraging Behavior | Pearson Correlation between Plant Characteristics (r) | |
|---|---|---|
| Water Content | Plant Height | |
| Pecking rate | 0.492 | 0.626 |
| Foraging behavior time budget | 0.622 | 0.670 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, Y.; Cheng, L.; Fan, Y.; Zhou, L.; Song, Y. The Foraging Window for Greater White-Fronted Geese (Anser albifrons) Is Consistent with the Growth Stage of Carex. Diversity 2022, 14, 943. https://doi.org/10.3390/d14110943
Zhong Y, Cheng L, Fan Y, Zhou L, Song Y. The Foraging Window for Greater White-Fronted Geese (Anser albifrons) Is Consistent with the Growth Stage of Carex. Diversity. 2022; 14(11):943. https://doi.org/10.3390/d14110943
Chicago/Turabian StyleZhong, Yundong, Lei Cheng, Yanguang Fan, Lizhi Zhou, and Yunwei Song. 2022. "The Foraging Window for Greater White-Fronted Geese (Anser albifrons) Is Consistent with the Growth Stage of Carex" Diversity 14, no. 11: 943. https://doi.org/10.3390/d14110943
APA StyleZhong, Y., Cheng, L., Fan, Y., Zhou, L., & Song, Y. (2022). The Foraging Window for Greater White-Fronted Geese (Anser albifrons) Is Consistent with the Growth Stage of Carex. Diversity, 14(11), 943. https://doi.org/10.3390/d14110943