
Flower-Visiting Insect Assemblages on Fall-Blooming Native California Sage Scrub Shrubs

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling
2.2.1. Hand Netting
2.2.2. Photo Sampling
2.2.3. Malaise Trap Sampling
2.3. Insect Sorting and Identification
2.4. Analysis of Flower-Visiting Insect Assemblages
2.5. Pollen Analyses
2.5.1. Species Selection
2.5.2. Pollen Sample Removal
2.5.3. DNA Extraction
2.5.4. Pollen DNA Analyses
3. Results
3.1. Documentation of Flower-Visiting Insects
3.2. Comparison between Active and Passive Sampling Techniques
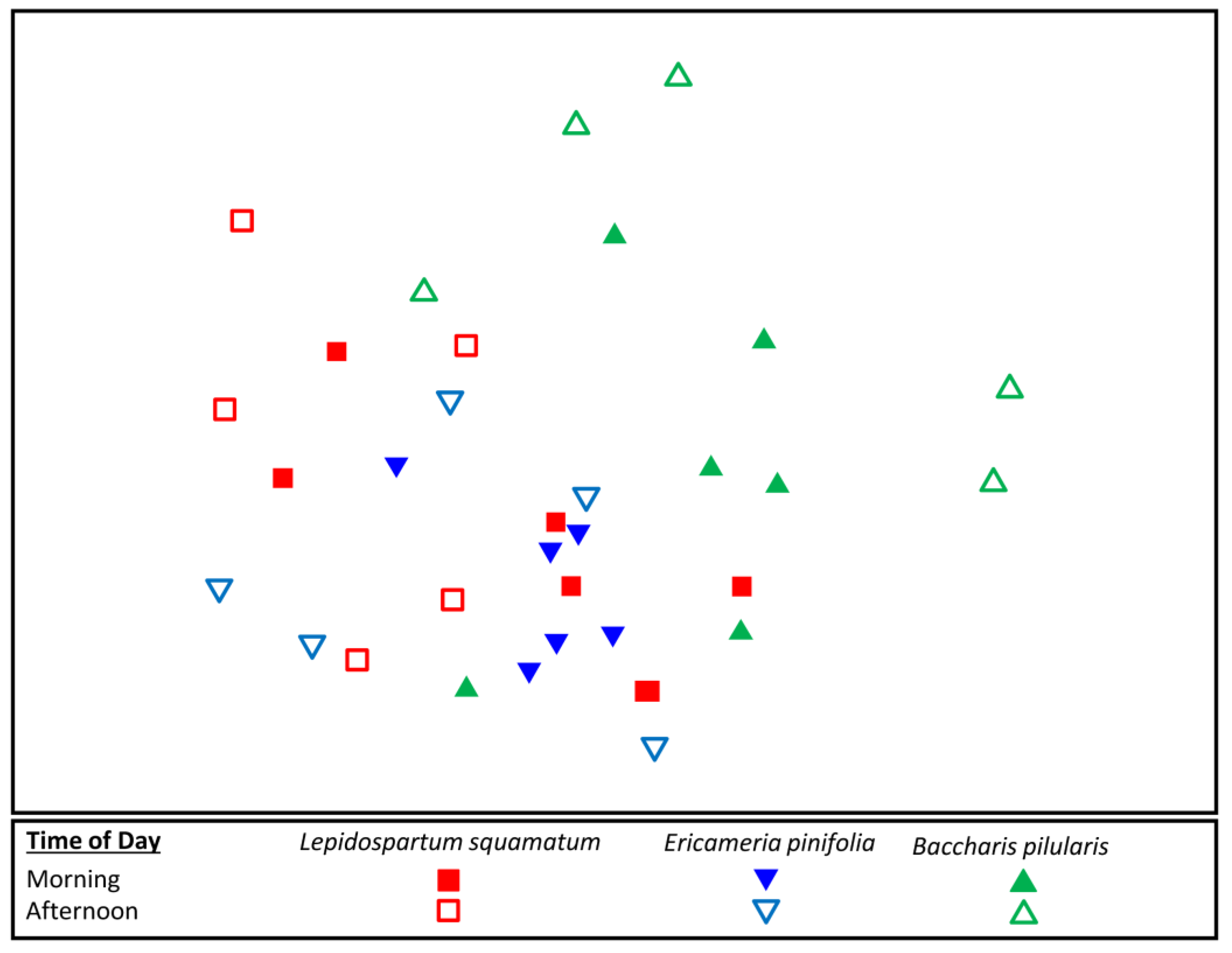
3.3. Flower-Visiting Insect Assemblage Differences on Fall Flowering Shrubs
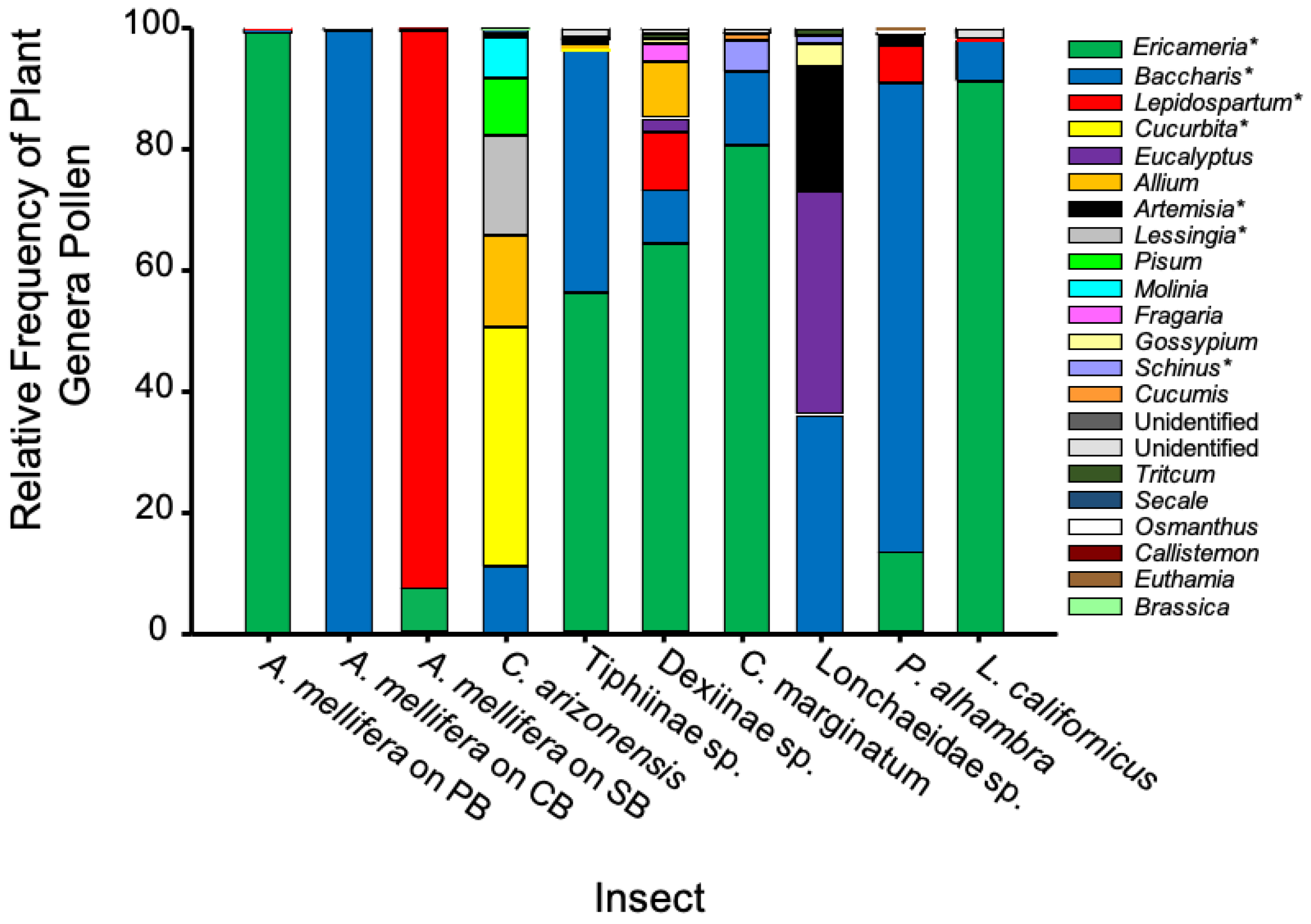
3.4. Pollen Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michener, C.D. Biogeography of the bees. Ann. Mo. Bot. Garden 1979, 66, 277–347. [Google Scholar] [CrossRef]
- Cincotta, R.P.; Wisnewski, J.; Engelman, R. Human population in the biodiversity hotspots. Nature 2000, 404, 990–992. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.-L.J.; Ascher, J.S.; Gibbs, J.; Irwin, R.E.; Bolger, D.T. Effects of fragmentation on a distinctive coastal sage scrub bee fauna revealed through incidental captures by pitfall traps. J. Insect Conserv. 2015, 19, 175–179. [Google Scholar] [CrossRef]
- Moldenke, A.R. California pollinationecology and vegetation types. Phytologia 1976, 34, 305–361. [Google Scholar]
- Hung, K.-L.J.; Ascher, J.S.; Holway, D.A. Urbanization-induced habitat fragmentation erodes multiple components of temporal diversity in a Southern California native bee assemblage. PLoS ONE 2017, 12, e0184136. [Google Scholar] [CrossRef] [Green Version]
- Munz, P.A.; Renshaw, D.L.; Faber, P.M. Introduction to California Desert Wildflowers; University of California Press: Berkeley, CA, USA, 2020. [Google Scholar]
- Westman, W.E. Diversity relations and succession in Californian coastal sage scrub. Ecology 1981, 62, 170–184. [Google Scholar] [CrossRef]
- Bolger, D.T.; Suarez, A.V.; Crooks, K.R.; Morrison, S.A.; Case, T.J. Arthropods in urban habitat fragments in southern California: Area age, and edge effects. Ecol. Appl. 2000, 10, 1230–1248. [Google Scholar] [CrossRef]
- Rubinoff, D. Evaluating the California gnatcatcher as an umbrella species for conservation of southern California coastal sage scrub. Conserv. Biol. 2001, 15, 1374–1383. [Google Scholar] [CrossRef]
- Riordan, E.C.; Rundel, P.W. Modelling the distribution of a threatened habitat: The California sage scrub. J. Biogeogr. 2009, 36, 2176–2188. [Google Scholar] [CrossRef]
- Shreeve, T.G.; Mason, C.F. The number of butterfly species in woodlands. Oecologia 1980, 45, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Bowers, M.A. Bumble bee colonization, extinction, and reproduction in subalpine meadows in northeastern Utah. Ecology 1985, 66, 914–927. [Google Scholar] [CrossRef]
- Jennersten, O. Pollination in Dianthus deltoides (Caryophyllaceae): Effects of habitat fragmentation on visitation and seed set. Conserv. Biol. 1988, 2, 359–366. [Google Scholar] [CrossRef]
- Cane, J.H. Habitat fragmentation and native bees: A premature verdict? Conserv. Ecol. 2001, 5, 1. [Google Scholar] [CrossRef]
- Gezon, Z.J.; Wyman, E.S.; Ascher, J.S.; Inouye, D.W.; Irwin, R.E. The effect of repeated, lethal sampling on wild bee abundance and diversity. Methods Ecol. Evol. 2015, 6, 1044–1054. [Google Scholar] [CrossRef]
- Sawyer, J.O.; Keeler-Wolf, T.; Evens, J.M. A Manual of California Vegetation; California Native Plant Society Press: Sacramento, CA, USA, 1995. [Google Scholar]
- Tabush, C.P. Temporal Bee Diversity and Abundance within the California Sage Scrub of the San Jose Hills and Chino Hills. Master’s Dissertation, California State Polytechnic University, Pomona, CA, USA, 2019. [Google Scholar]
- Montalvo, A.M.; Riordan, E.C.; Beyers, J. Plant Profile for Lepidospartum squamatum. In Riverside-Corona Resource Conservation District and U.S. Department of Agriculture, Forest Service; Pacific Southwest Research Station: Riverside, CA, USA, 2017. [Google Scholar]
- Francis, J.K. Wildland Shrubs of the United States and Its Territories: Thamnic Descriptions, Volume 1; U.S. Department of Agriculture, Forest Service, International Institute of Tropical Forestry: San Juan, PR, USA; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2004.
- Hoffman, L. Spatiotemporal Variation in Pollinator Taxa on the Santa Ana River Woolly Star Eriastrum densifolium ssp. sanctorum (Milliken) Mason (Polemoniaceae). Master’s Dissertation, California State University, Fullerton, CA, USA, 2010. [Google Scholar]
- Barrios-Garcia, M.N.; Rodriguez-Cabal, M.A.; Rudgers, J.A.; Crutsinger, G.M. Soil fertilization does not alter plant architectural effects on arthropod communities. J. Plant Ecol. 2016, 10, 800–807. [Google Scholar] [CrossRef] [Green Version]
- Rudgers, J.A.; Whitney, K.D. Interactions between insect herbivores and a plant architectural dimorphism. J. Ecol. 2006, 94, 1249–1260. [Google Scholar] [CrossRef]
- Huey, S.; Nieh, J.C. Foraging at a safe distance: Crab spider effects on pollinators: Crab spider effects on pollinators. Ecol. Entomol. 2017, 42, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Wania, A.; Kühn, I.; Klotz, S. Plant richness patterns in agricultural and urban landscapes in central Germany—Spatial gradients of species richness. Landsc. Urb. Plan. 2006, 75, 97–110. [Google Scholar] [CrossRef]
- Stout, J.C.; Morales, C.L. Ecological impacts of invasive alien species on bees. Apidologie 2009, 40, 388–409. [Google Scholar] [CrossRef] [Green Version]
- Goddard, M.A.; Dougill, A.J.; Benton, T.J. Scaling up from gardens: Biodiversity conservation in urban environments. Trends Ecol. Evol. 2010, 25, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, M.A.; Sax, D.F.; Olden, J.D. The potential conservation value of non-native species: Conservation value of non-native species. Conserv. Biol. 2011, 25, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Koyama, A.; Egawa, C.; Taki, H.; Yasuda, M.; Kanzaki, N.; Ide, T.; Okabe, K. Non-native plants are a seasonal pollen source for native honeybees in suburban ecosystems. Urban Ecosyst. 2018, 21, 1113–1122. [Google Scholar] [CrossRef]
- Bjerknes, A.-L.; Totland, Ø.; Hegland, S.J.; Nielsen, A. Do alien plant invasions really affect pollination success in native plant species? Biol. Conserv. 2007, 138, 1–12. [Google Scholar] [CrossRef]
- Frankie, G.; Pawelek, J.; Chase, M.H.; Jadallah, C.C.; Feng, I.; Rizzardi, M.; Thorp, R. Native and non-native plants attract diverse bees to urban gardens in California. J. Pollination Ecol. 2019, 25, 16–23. [Google Scholar] [CrossRef]
- Alarcón, R. Congruence between visitation and pollen-transport networks in a California plant-pollinator community. Oikos 2010, 119, 35–44. [Google Scholar] [CrossRef]
- Thomson, D.M.; Meyer, W.M., III; Whitcomb, I.F. Non-native plant removal and high rainfall years promote post-fire recovery of Artemisia californica in southern California sage scrub. PLoS ONE 2021, 16, e0254398. [Google Scholar] [CrossRef]
- Dartnell, S.D.; Hamlett, N.; Meyer, W.M., III. Monitoring butterfly assemblages in southern California to assess the impacts of habitat and climate modifications. J. Insect Conserv. 2022, 26, 149–162. [Google Scholar] [CrossRef]
- Caspi, T.; Hartz, L.A.; Soto Villa, A.E.; Loesberg, J.A.; Robins, C.R.; Meyer, W.M., III. Impacts of invasive annuals on soil carbon and nitrogen storage in southern California depend on identity of the invader. Ecol. Evol. 2019, 9, 4980–4993. [Google Scholar] [CrossRef]
- Xu, X.; Ren, Z.-X.; Trunschke, J.; Kuppler, J.; Zhao, Y.-H.; Knop, E.; Wang, H. Bimodal activity of diurnal flower visitation at high elevation. Ecol. Evol. 2021, 11, 13487–13500. [Google Scholar] [CrossRef]
- Irwin, R.E.; Brody, A.K.; Waser, N.M. The impact of floral larceny on individuals, populations, and communities. Oecologia 2001, 129, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, A.L.; Harder, L.D.; Johnson, S.D. Consumptive emasculation: The ecological and evolutionary consequences of pollen theft. Biol. Rev. 2009, 84, 259–276. [Google Scholar] [CrossRef] [PubMed]
- Inouye, D.W. The terminology of floral larceny. Ecology 1980, 61, 1251–1253. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER V6: User Manual/Tutorial. Plymouth, PRIMER-E. Mar. Biol. 2006, 118, 167–176. [Google Scholar] [CrossRef]
- Bell, K.L.; Fowler, J.; Burgess, K.S.; Dobbs, E.K.; Gruenewald, D.; Lawley, B.; Morozumi, C.; Brosi, B.J. Applying pollen DNA metabarcoding to the study of plant–pollinator interactions. Appl. Plant Sci. 2017, 5, 1600124. [Google Scholar] [CrossRef]
- Sickel, W.; Ankenbrand, M.J.; Grimmer, G.; Holzschuh, A.; Härtel, S.; Lanzen, J.; Steffan-Dewenter, I.; Keller, A. Increased efficiency in identifying mixed pollen samples by meta-barcoding with a dual-indexing approach. BMC Ecol. 2015, 15, 20. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, D.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotech. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Rivers, A.R.; Weber, K.C.; Gardner, T.G.; Liu, S.; Armstrong, S.D. ITSxpress: Software to rapidly trim internally transcribed spacer sequences with quality scores for marker gene analysis. F1000Research 2018, 7, 1418. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Banchi, E.; Ametrano, C.G.; Greco, S.; Stanković, D.; Muggia, L.; Pallavicini, A. PLANiTS: A curated sequence reference dataset for plant ITS DNA metabarcoding. Database 2020, 2020, baz155. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, A.H.; Thomas, M.; Bhandari, R.; Meshram, H. Malaise trap and insect sampling: Mini review. Biol. Bull. 2016, 2, 35–40. [Google Scholar]
- Cane, J.H.; Minckley, R.L.; Kervin, L.J. Sampling bees (Hymenoptera: Apiformes) for pollinator community studies: Pitfalls of pan-trapping. J. Kans. Entomol. Soc. 2000, 73, 225–231. [Google Scholar]
- Trent, T.R.; Evans, E.W.; Tepedino, V.J. Pan-trapping for bees (Hymenoptera: Apiformes) in Utah’s West Desert: The importance of color diversity. Pan-Pac. Entomol. 2005, 81, 103–113. [Google Scholar]
- Roulston, T.H.; Smith, S.A.; Brewster, A.L. A comparison of pan trap and intensive net sampling techniques for documenting a bee (Hymenoptera: Apiformes) fauna. J. Kans. Entomol. Soc. 2007, 80, 179–181. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyôrgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different European habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Matthews, R.W.; Matthews, R.J. Malaise trap: The townes trap catches more insects. Contrib. Am. Entomol. Inst. 1983, 20, 428–432. [Google Scholar]
- Torchio, P.F.; Trostle, G.E. Biological notes on Anthophora urbana urbana and its parasite, Xeromelecta californica (Hymenoptera: Anthophoridae), including descriptions on late embryogenesis and hatching. Annal. Entomol. Soc. Am. 1986, 79, 434–447. [Google Scholar] [CrossRef]
- Bossert, S.; Wood, T.J.; Patiny, S.; Michez, D.; Almeida, E.A.B.; Minckley, R.L.; Packer, L.; Neff, J.L.; Copeland, R.S.; Straka, J.; et al. Phylogeny, biogeography and diversification of the mining bee family Andrenidae. Syst. Entomol. 2021, 47, 283–302. [Google Scholar] [CrossRef]
- Timberlake, P.H. A Revisional Study of the Bees of the Genus Perdita F. Smith, with Special Reference to the Fauna of the Pacific Coast (Hymenoptera, Apoidea) Part III; University of California Press: Berkeley, CA, USA; Los Angeles, CA, USA, 1958. [Google Scholar]
- Riley, J.R.; Greggers, U.; Smith, A.D.; Reynolds, D.R.; Menzel, R. The flight paths of honeybees recruited by dance. Nature 2005, 425, 205–207. [Google Scholar] [CrossRef]
- Grüter, C.; Farina, W. The honeybee waggle dance: Can we follow the steps? Trends Ecol. Evol. 2009, 24, 242–247. [Google Scholar] [CrossRef]
- Nürnberger, F.; Keller, A.; Härtel, S.; Steffan-Dewenter, I. Honey bee waggle dance communication increases diversity of pollen diets in intensively managed agricultural landscapes. Mol. Ecol. 2019, 28, 3602–3611. [Google Scholar] [CrossRef] [PubMed]
- Abrol, D.P. Foraging behavior of bees as influenced by quality and quantity of rewards from flowers. J. Asia-Pac. Entomol. 2006, 9, 145–148. [Google Scholar] [CrossRef]
- Al Toufailia, H.; Couvillon, M.J.; Ratnieks, F.L.W.; Grüter, C. Honey bee waggle dance communication: Signal meaning and signal noise affect dance follower behaviour. Behav. Ecol. Sociobiol. 2013, 67, 549–556. [Google Scholar] [CrossRef]
- Saritha, R.; Vinod Chandra, S.S. Multi dimensional honey bee foraging algorithm based on optimal energy consumption. J. Inst. Eng. India Ser. B 2017, 98, 527–531. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plant Species | Sample Start Date | Sample End Date |
|---|---|---|
| L. squamatum | 16 September | 1 October |
| E. pinifolia | 12 October | 5 November |
| B. pilularis | 20 October | 5 November |
| Species | No. Indiv. | Netting Location | Extracted DNA (ng/µL) | No. Raw Reads | No. Plant Genera | % Pollen from BFS 2 |
|---|---|---|---|---|---|---|
| Apis mellifera | 44 | L. squamatum | 2.45 | 34,630 | 7 | 99.79 |
| Apis mellifera | 46 | B. pilularis | 9.0 | 32,878 | 2 | 100 |
| Apis mellifera | 62 | E. pinifolia | 4.1 | 21,249 | 6 | 99.93 |
| Ceratina arizonensis | 19 | All 3 hosts | 1.55 | 13,933 | 9 | 67.17 |
| Copestylum marginatum | 8 | All 3 hosts | 0 1 | 12,684 | 5 | 97.90 |
| Dexiinae sp. | 18 | L. squamatum & E. pinifolia | 0.05 | 23,660 | 9 | 82.84 |
| Largus californicus | 6 | All 3 hosts | 0 | 17,021 | 5 | 98.23 |
| Lonchaeidae sp.1 | 13 | B. pilularis | 0 | 11,869 | 6 | 58.16 |
| Palpada alhambra | 27 | All 3 hosts | 0.3 | 12,684 | 11 | 99.05 |
| Tiphiinae sp.1 | 22 | B. pilularis | 1.8 | 15,634 | 7 | 97.63 |
| Species | Pine Bush | Coyote Bush | Scale Broom | Malaise | |||
|---|---|---|---|---|---|---|---|
| Net | Photo | Net | Photo | Net | Photo | ||
| Agapostemon texanus | X | X | |||||
| Agapostemon sp. | X | ||||||
| Anthophora urbana | X | X | X | ||||
| Anthophora sp. | X | ||||||
| Apis mellifera | X | X | X | X | X | ||
| Ashmeadiella sp. | X | ||||||
| Ceratina arizonensis | X | X | X | X | |||
| Ceratina (Zadontomerus) sp. | X | ||||||
| Ceratina sp. | X | ||||||
| Colletes sp.1 | X | ||||||
| Colletes sp.2 | X | ||||||
| Dialictus sp.1 | X | ||||||
| Dialictus sp.2 | X | ||||||
| Dialictus sp.3 | X | ||||||
| Hylaeus sp. | X | ||||||
| Halictus farinosus | X | ||||||
| Halictus ligatus | X | ||||||
| Perdita ericameriae | X | X | |||||
| Rophitinae sp. | X | ||||||
| Xeromelecta californica | X | ||||||
| Xylocopa sonarina | X | ||||||
| Total: 21 bee species | 8 | 4 | 8 | 0 | 6 | 6 | 0 |
| Species | Pine Bush | Coyote Bush | Scale-Broom | Malaise | |||
|---|---|---|---|---|---|---|---|
| Net | Photo | Net | Photo | Net | Photo | ||
| Atlides halesus | X | ||||||
| Brephidium exilis | X | ||||||
| Danaus gilippus | X | ||||||
| Danaus plexippus | X | ||||||
| Erynnis tristis | X | X | |||||
| Euptoieta claudia | X | ||||||
| Heliopetes ericetorum | X | ||||||
| Hemiargus ceraunus | X | ||||||
| Hylephila phyleus | X | X | X | ||||
| Junonia coenia | X | X | |||||
| Plebejus acmon | X | ||||||
| Pyrgus albescens | X | ||||||
| Strymon melinus | X | X | X | ||||
| Vanessa cardui | X | X | X | ||||
| Total: 14 butterfly species | 1 | 4 | 0 | 1 | 2 | 14 | 0 |
| Species | Pine Bush | Coyote Bush | Scale Broom | Malaise | |||
|---|---|---|---|---|---|---|---|
| Net | Photo | Net | Photo | Net | Photo | ||
| Ancistrocerus sp. | X | ||||||
| Anomalon sp. | X | ||||||
| Bembix sp. | X | ||||||
| Braconidae sp.1 | X | ||||||
| Braconidae sp.2 | X | ||||||
| Brasema sp. | X | X | |||||
| Cerceris convergens | X | ||||||
| Cerceris sp. | X | ||||||
| Chalcidoidea sp. | X | ||||||
| Eulophidae sp. | X | ||||||
| Eurytomidae sp.1 | X | X | |||||
| Eurytomidae sp.2 | X | ||||||
| Gasteruptiidae sp.1 | X | ||||||
| Gasteruptiidae sp.2 | X | ||||||
| Leptochilus sp. | X | ||||||
| Oxybelus sp. | X | ||||||
| Paratiphia sp.1 | X | ||||||
| Paratiphia sp. 2 | X | ||||||
| Philanthus gibbosus | X | ||||||
| Philanthus sp.1 | X | ||||||
| Philanthus sp. 2 | X | ||||||
| Polistes aurifer | X | ||||||
| Tiphiinae sp. 1 | X | X | |||||
| Tiphiinae sp. 2 | X | ||||||
| Tiphiinae sp. 3 | X | ||||||
| Vespula pensylvanica | X | ||||||
| Total: 26 wasp species | 4 | 0 | 8 | 0 | 8 | 9 | 0 |
| Species | Pine Bush | Coyote Bush | Scale-Broom | Malaise | |||
|---|---|---|---|---|---|---|---|
| Net | Photo | Net | Photo | Net | Photo | ||
| Bagrada hilaris | X | ||||||
| Largus californicus | X | X | X | X | X | ||
| Largus sp. | X | ||||||
| Murgantia histrionica | X | ||||||
| Neacoryphus bicrucis | X | ||||||
| Total: 5 hemipterans | 1 | 1 | 1 | 0 | 2 | 4 | 0 |
| Species | Pine Bush | Coyote Bush | Scale-Broom | Malaise | |||
|---|---|---|---|---|---|---|---|
| Net | Photo | Net | Photo | Net | Photo | ||
| Acreophthiria sp. | X | X | |||||
| Allograpta obliqua | X | ||||||
| Aphoebantus sp.1 | X | X | X | ||||
| Aphoebantus sp.2 | X | ||||||
| Bombyliidae sp.1 | X | X | |||||
| Bombyliidae sp.2 | X | ||||||
| Bombyliidae sp.3 | X | ||||||
| Calliphoridae sp.1 | X | ||||||
| Calliphoridae sp.2 | X | ||||||
| Ceratitis capitata | X | ||||||
| Coenosia sp. | X | ||||||
| Conopidae sp. | X | ||||||
| Copestylum marginatum | X | X | X | ||||
| Copestylum mexicanum | X | X | X | X | X | ||
| Copestylum satur | X | X | X | X | |||
| Copestylum violaceum | X | ||||||
| Desmometopa sp. | X | ||||||
| Dexiinae sp. | X | X | X | X | |||
| Dilophus sp. | X | ||||||
| Dioprosopa clavata | X | X | X | X | |||
| Diptera sp.1 | X | ||||||
| Diptera sp.2 | X | X | |||||
| Diptera sp.3 | X | X | |||||
| Diptera sp.4 | X | ||||||
| Diptera sp.5 | X | ||||||
| Diptera sp.6 | X | ||||||
| Diptera sp.7 | X | ||||||
| Diptera sp.8 | X | ||||||
| Diptera sp.9 | X | N/A * | |||||
| Diptera sp.10 | X | N/A * | |||||
| Eristalinus taeniops | X | X | |||||
| Eristalis hirta | X | X | |||||
| Eupeodes fumipennis | X | ||||||
| Eupeodes volucris | X | X | |||||
| Geron sp. | X | X | |||||
| Lonchaeidae sp.1 | X | ||||||
| Lonchaeidae sp.2 | X | ||||||
| Lucilia sp. | X | ||||||
| Musciodea sp. | X | X | |||||
| Oscinellinae sp. | X | ||||||
| Palpada alhambra | X | X | X | X | X | ||
| Palpada mexicana | X | X | X | ||||
| Paragus haemorrhous | X | ||||||
| Paragus sp. | X | ||||||
| Phthiriinae sp.1 | X | X | |||||
| Spilomyia interrupta | X | ||||||
| Tachinidae sp.1 | X | ||||||
| Tachinidae sp.2 | X | ||||||
| Tachinidae sp.3 | X | ||||||
| Tephritini sp. | X | ||||||
| Thaumatomyia sp. | X | ||||||
| Toxomerus marginatus | X | X | X | X | |||
| Trichopoda pennipes | X | X | |||||
| Villa lateralis | X | ||||||
| Villa sp. | X | X | |||||
| Zodion sp. | X | ||||||
| Total: 56 fly species | 13 | 8 | 30 | 5 | 16 | 15 | 7 |
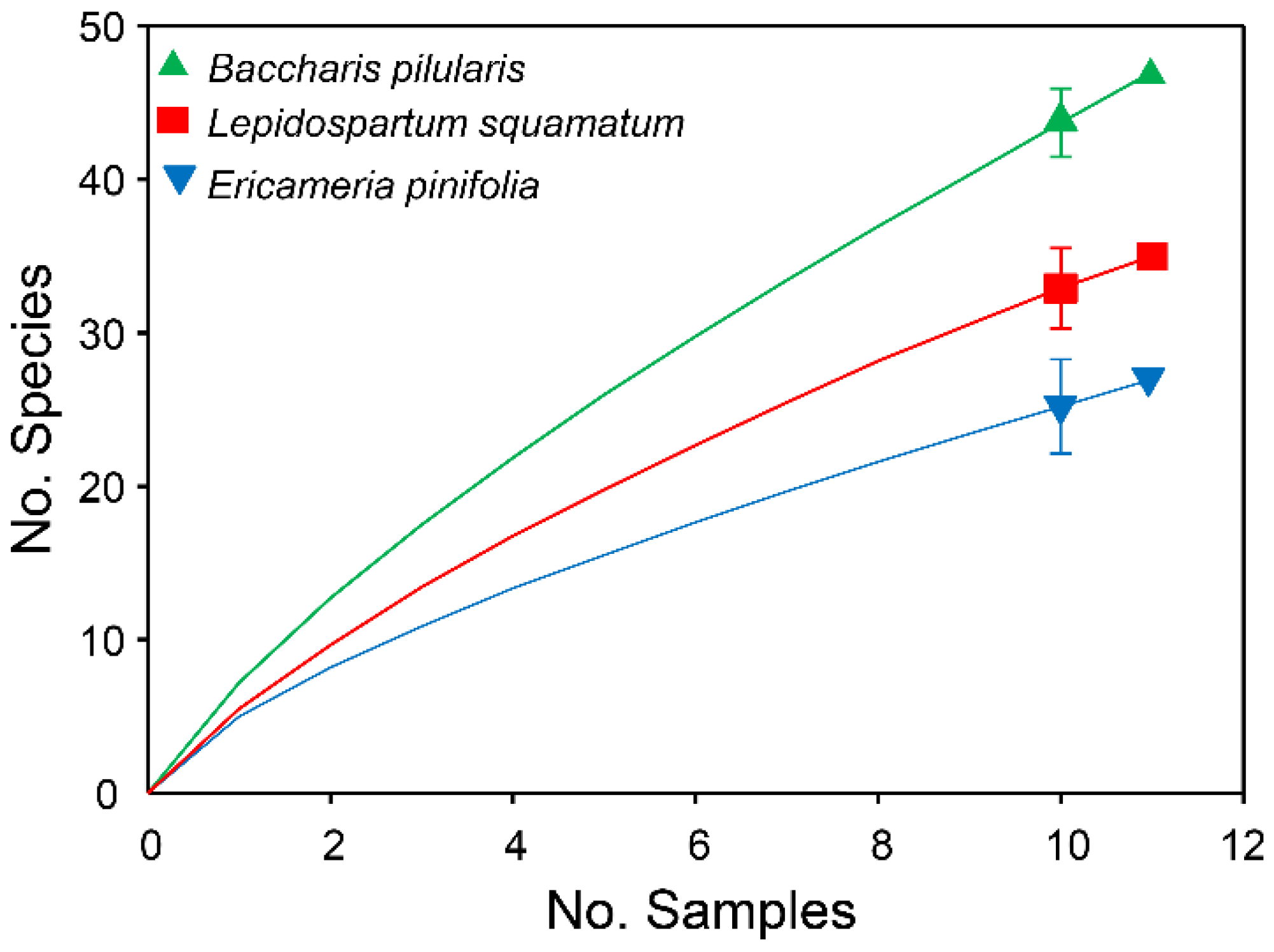
| Plant Species | No. Species Collected | No.Unique Species | No. Singleton Specimens | Predicted Species Richness (SD) | % Inventory Completeness |
|---|---|---|---|---|---|
| L. squamatum | 35 | 22 | 14 | 76.1 (23.8) | 45.99 |
| E. pinifolia | 27 | 12 | 8 | 117.3 (76.7) | 23.02 |
| B. pilularis | 47 | 36 | 26 | 209 (98.2) | 22.49 |
| Pairwise Comparison | R Value | p Value |
|---|---|---|
| L. squamatum, E. pinifolia | −0.025 | 0.614 |
| L. squamatum, B. pilularis | 0.319 | 0.002 * |
| E. pinifolia, B. pilularis | 0.283 | 0.005 * |
| Pairwise Comparison Insect Species | Proportion of Samples Present | Avg. Dissimilarity ± 1 SD | Contributed % Dissimilarity | |
|---|---|---|---|---|
| B. pilularis vs. E. pinifolia | B. pilularis | E. pinifolia | ||
| Tiphiinae sp.1 | 0.55 | 0.00 | 4.66 ± 1.01 | 5.94 |
| Palpada alhambra | 0.55 | 0.64 | 4.40 ± 0.91 | 5.62 |
| Largus californicus | 0.09 | 0.36 | 3.12 ± 0.76 | 3.97 |
| Copestylum marginatum | 0.09 | 0.36 | 3.10 ± 0.73 | 3.96 |
| Lonchaeidae sp.2 | 0.36 | 0.00 | 2.78 ± 0.70 | 3.54 |
| Braconidae sp.1 | 0.36 | 0.00 | 2.68 ± 0.71 | 3.42 |
| Dexiinae sp. | 0.00 | 0.27 | 2.33 ± 0.58 | 2.97 |
| Desmometopa sp. | 0.27 | 0.00 | 2.13 ± 0.58 | 2.71 |
| Ceratina arizonensis | 0.18 | 0.09 | 2.05 ± 0.51 | 2.61 |
| Braconidae sp.2 | 0.00 | 0.27 | 2.05 ± 0.60 | 2.61 |
| B. pilularis vs. L. squamatum | B. pilularis | L. squamatum | ||
| Tiphiinae sp.1 | 0.55 | 0.00 | 4.39 ± 1.03 | 5.34 |
| Palpada alhambra | 0.55 | 0.36 | 4.24 ± 0.96 | 5.17 |
| Dexiinae sp. | 0.00 | 0.45 | 4.02 ± 0.87 | 4.90 |
| Copestylum marginatum | 0.09 | 0.27 | 2.74 ± 0.65 | 3.34 |
| Lonchaeidae sp.2 | 0.36 | 0.00 | 2.63 ± 0.71 | 3.20 |
| Braconidae sp.1 | 0.36 | 0.00 | 2.55 ± 0.72 | 3.10 |
| Ceratina arizonensis | 0.18 | 0.18 | 2.22 ± 0.62 | 2.71 |
| Desmometopa sp. | 0.27 | 0.00 | 2.01 ± 0.58 | 2.45 |
| Lonchaeidae sp.1 | 0.27 | 0.00 | 1.89 ± 0.59 | 2.30 |
| Apis mellifera | 0.82 | 1.00 | 1.69 ± 0.46 | 2.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dartnell, S.; Cavalcanti, A.R.O.; Misaki Bluebell, A.; Hamlett, N.V.; Crane, E.J., III; Meyer, W.M., III. Flower-Visiting Insect Assemblages on Fall-Blooming Native California Sage Scrub Shrubs. Diversity 2022, 14, 958. https://doi.org/10.3390/d14110958
Dartnell S, Cavalcanti ARO, Misaki Bluebell A, Hamlett NV, Crane EJ III, Meyer WM III. Flower-Visiting Insect Assemblages on Fall-Blooming Native California Sage Scrub Shrubs. Diversity. 2022; 14(11):958. https://doi.org/10.3390/d14110958
Chicago/Turabian StyleDartnell, Sofia, Andre R. O. Cavalcanti, Ayame Misaki Bluebell, Nancy V. Hamlett, Edward J. Crane, III, and Wallace M. Meyer, III. 2022. "Flower-Visiting Insect Assemblages on Fall-Blooming Native California Sage Scrub Shrubs" Diversity 14, no. 11: 958. https://doi.org/10.3390/d14110958
APA StyleDartnell, S., Cavalcanti, A. R. O., Misaki Bluebell, A., Hamlett, N. V., Crane, E. J., III, & Meyer, W. M., III. (2022). Flower-Visiting Insect Assemblages on Fall-Blooming Native California Sage Scrub Shrubs. Diversity, 14(11), 958. https://doi.org/10.3390/d14110958