

Diversity of Palaearctic Dragonflies and Damselflies (Odonata)

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
- -
- Are there differences in diversity patterns shown by lotic and lentic species?
- -
- Are there areas with relatively high endemism?
- -
- Are there areas with a relatively high percentage of globally threatened species, and do these match with Odonate endemism and richness hotspots?
2. Materials and Methods
2.1. Definition of Palaearctic Realm
2.2. Species Occurrence Data
2.3. Functional Traits and Conservation Status
2.4. Species Distribution Modelling
2.5. Calculating Richness and Endemism
3. Results
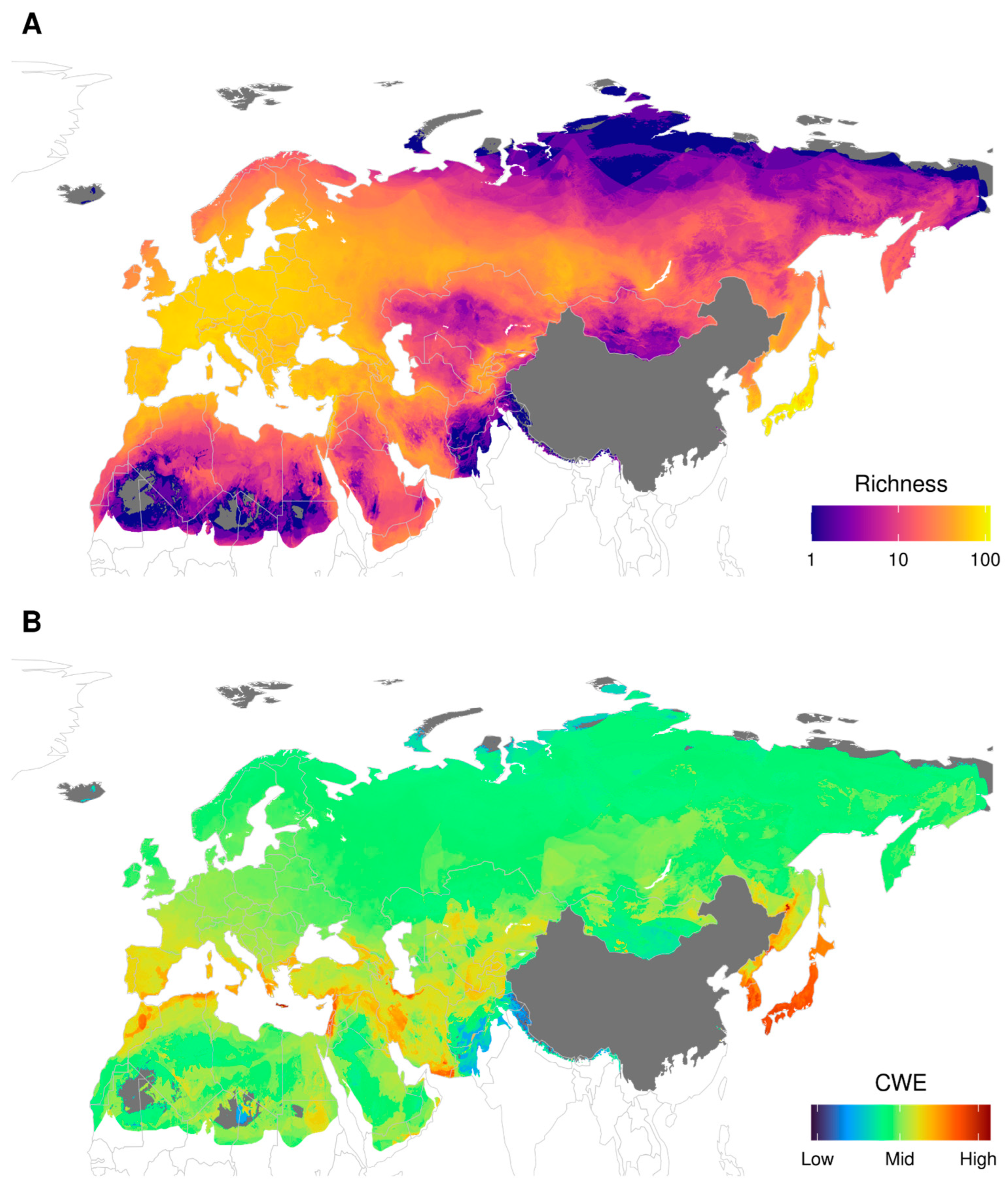
3.1. Richness and Corrected Weighted Endemism (CWE)
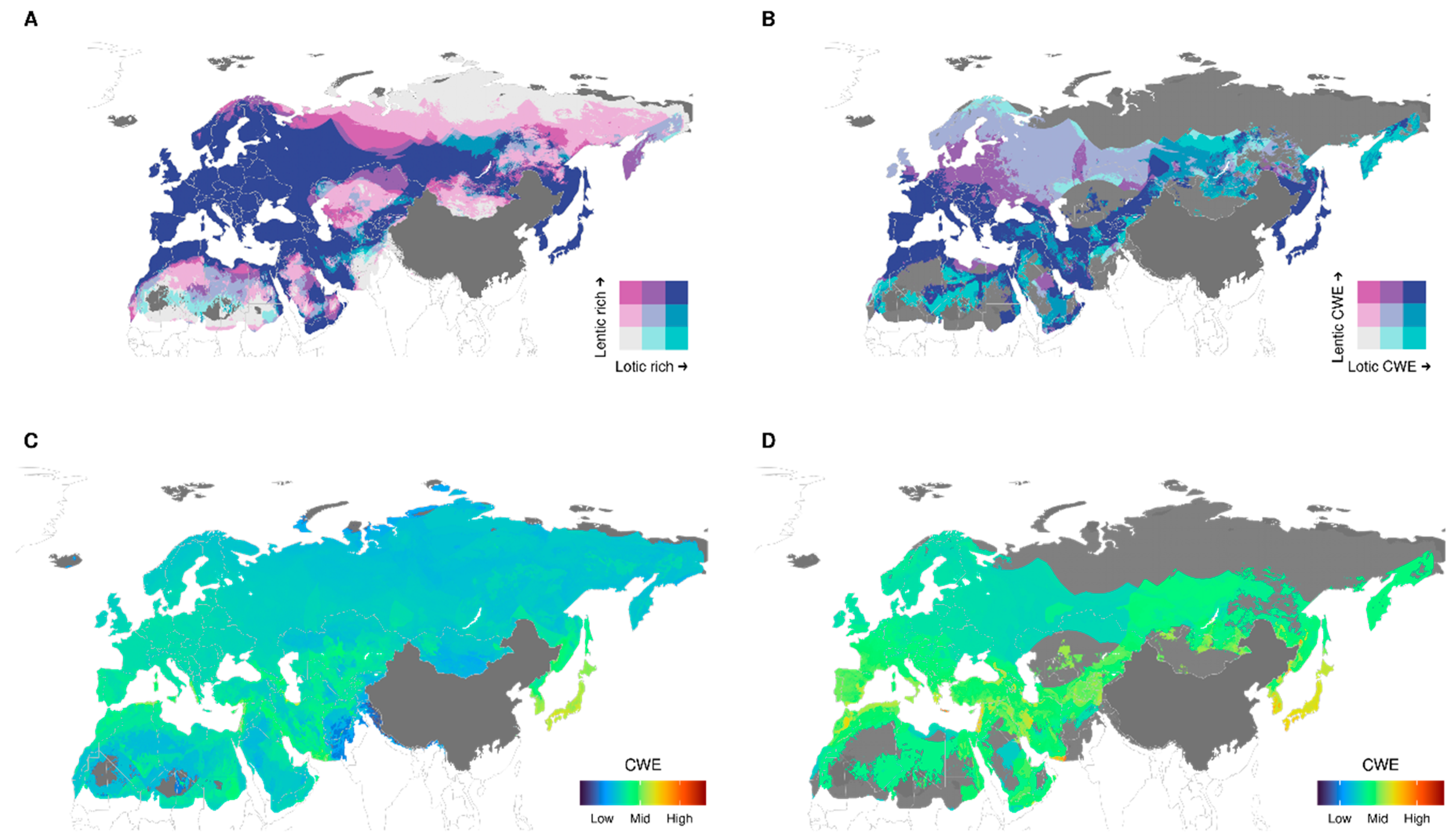
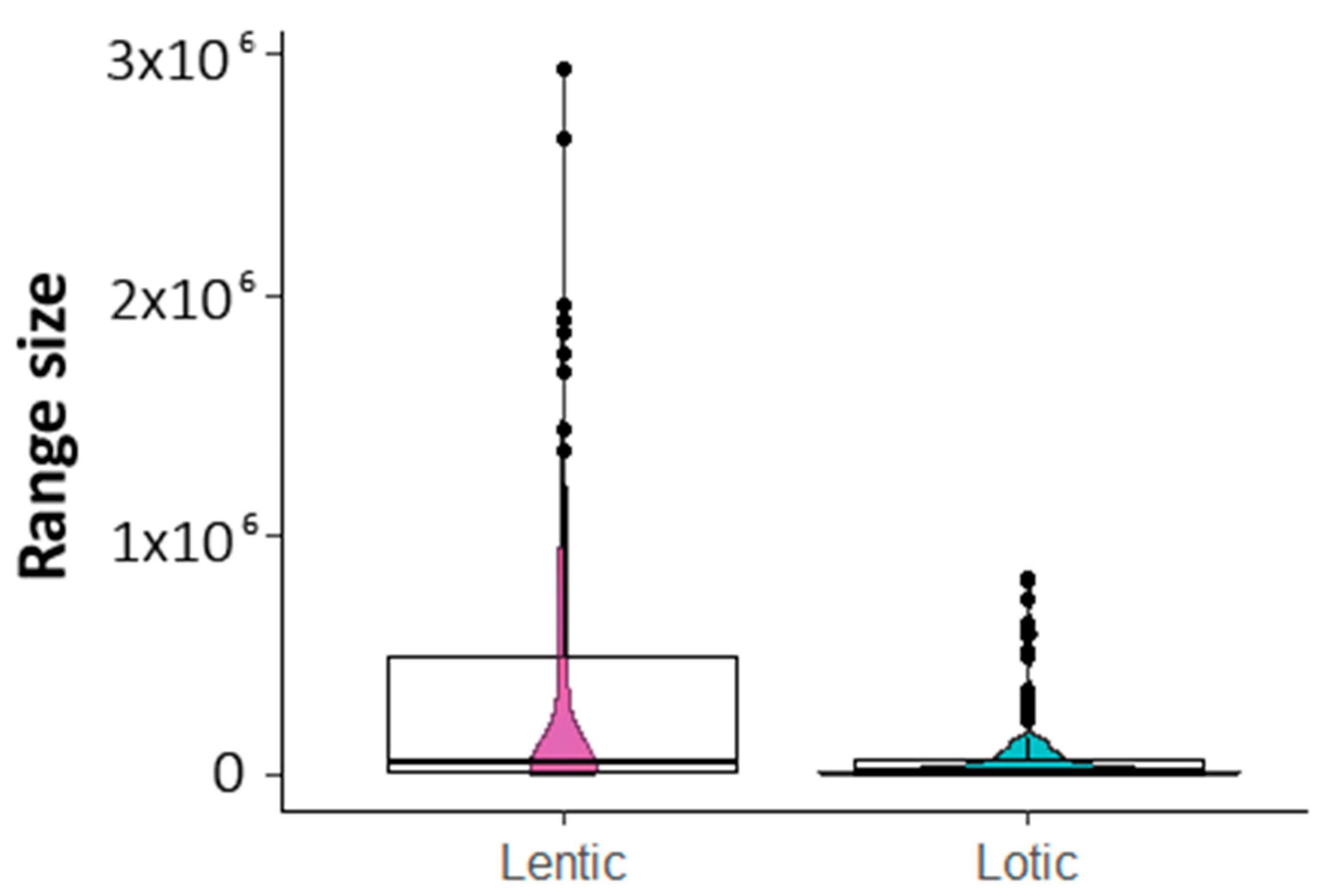
3.2. Richness and Endemism by Aquatic Habitats
3.3. Richness of Species According to IUCN Red List Category
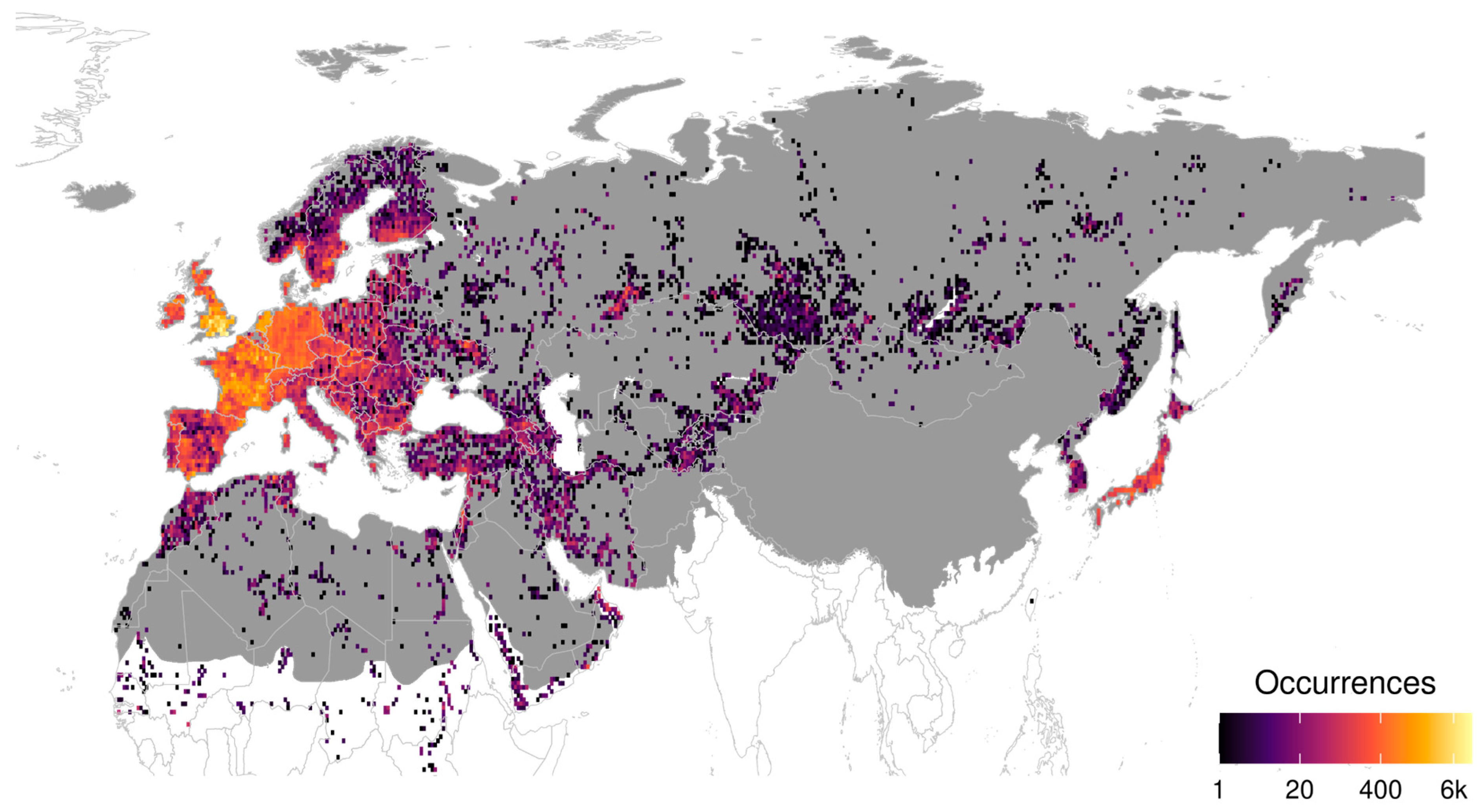
3.4. Sampling Effort
4. Discussion
4.1. General Diversity Patterns
4.2. Are There Differences in Diversity Patterns Shown by Lotic and Lentic Species?
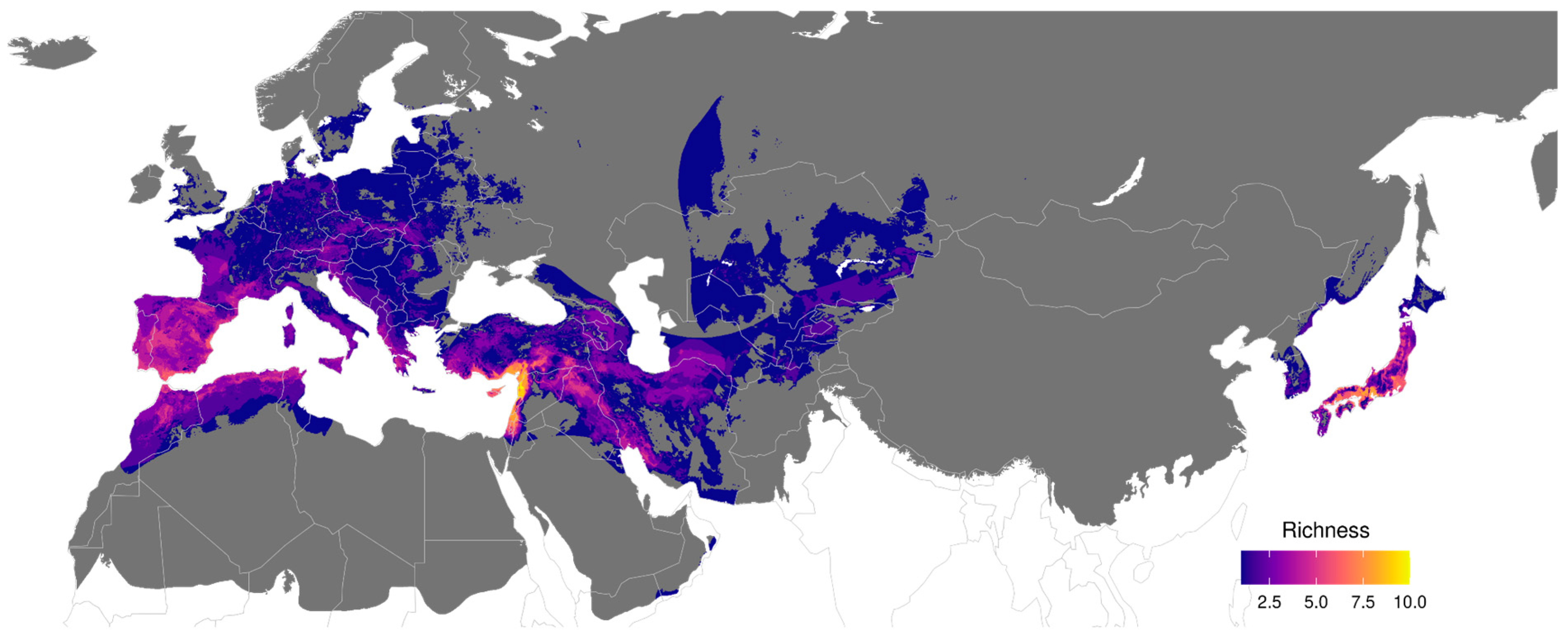
4.3. Are There Areas with Relatively High Endemism?
4.4. Are There Areas with a Relatively High Percentage of Globally Threatened Species?
5. Further Research
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wasscher, M.T.; Bos, F.G. The European dragonflies: Notes on the checklist and on species diversity. Odonatologica 2000, 29, 31–43. [Google Scholar]
- Abbott, J.C.; Bota-Sierra, C.A.; Guralnick, R.; Kalkman, V.; González-Soriano, E.; Novelo-Gutiérrez, R.; Bybee, S.; Ware, J.; Belitz, M.W. Diversity of Nearctic Dragonflies and Damselflies (Odonata). Diversity 2022, 14, 575. [Google Scholar] [CrossRef]
- Boudot, J.-P.; Kalkman, V.J. Atlas of the European Dragonflies and Damselflies; KNNV-Publishing: Utrecht, The Netherlands, 2015. [Google Scholar]
- Clausnitzer, V.; Dijkstra, K.-D.B.; Koch, R.; Boudot, J.-P.; Darwall, W.R.T.; Kipping, J.; Samraoui, B.; Samways, M.J.; Simaika, J.P.; Suhling, F. Focus on African Freshwaters: Hotspots of dragonfly diversity and conservation concern. Front. Ecol. Environ. 2012, 10, 129–134. [Google Scholar] [CrossRef]
- Endersby, I. The Distribution of Australian Dragonflies; Busybird Publishing: Montmorency, VIC, Australia, 2021. [Google Scholar]
- Kalkman, V.J.; Boudot, J.P.; Bernard, R.; De Knijf, G.; Suhling, F.; Termaat, T. Diversity and conservation of European dragonflies and damselflies (Odonata). Hydrobiologia 2018, 811, 269–282. [Google Scholar] [CrossRef] [Green Version]
- Darwall, W.R.T.; Smith, K.G.; Allen, D.J.; Holland, R.A.; Harrison, I.J.; Brooks, E.G.E. (Eds.) The Diversity of Life in African Freshwaters: Under Water, under Threat. An Analysis of the Status and Distribution of Freshwater Species throughout Mainland Africa; IUCN: Cambridge, UK; Gland, Switzerland, 2011. [Google Scholar]
- Combe, F.J.; Sikes, D.S.; Tkach, V.V.; Hope, A.G. Origins and diversity of the Bering Sea Island fauna: Shifting linkages across the northern continents. Biodivers. Conserv. 2021, 30, 1205–1232. [Google Scholar] [CrossRef]
- Wen, J.; Nie, Z.-L.; Ickert-Bond, S.M. Intercontinental disjunctions between eastern Asia and western North America in vascular plants highlight the biogeographic importance of the Bering land bridge from late Cretaceous to Neogene. J. Syst. Evol. 2016, 54, 469–490. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Boudot, J.-P.; Kalkman, V.J.; Azpilicueta Amorin, M.; Bogdanović, T.; Cordero Rivera, A.; Degabriele, G.; Dommanget, J.-L.; Ferreira, S.; Garrigós, B.; Jović, M.; et al. Atlas of the Odonata of the Mediterranean and North Africa. Libellula Suppl. 2009, 9, 1–256. [Google Scholar]
- Boudot, J.P.; Borisov, S.; De Knijf, G.; Grunsven, R.H.A.; van Schröter, A.; Kalkman, V.J. Atlas of the dragonflies and damselflies of West and Central Asia. Brachytron Suppl. 2021, 22, 3–248. [Google Scholar]
- National Biodiversity Center of Japan, Ministry of the Environment, Japan, Tokyo. Data from: The National Survey on the Natural Environment Report of the Distributional Survey of Japanese Animals (Dragonflies). 2002. Available online: http://www.biodic.go.jp/reports/4-09/h000.html (accessed on 21 January 2021).
- Ozono, A.; Kawashima, I.; Futahashi, R. Dragonflies of Japan, Revised ed.; Bun-ichi Sogo Shuppan Co.: Tokyo, Japan, 2021. [Google Scholar]
- Zizka, A.; Silvestro, D.; Andermann, T.; Azevedo, J.; Duarte Ritter, C.; Edler, D.; Farooq, H.; Herdean, A.; Ariza, M.; Scharn, R.; et al. CoordinateCleaner: Standardized Cleaning of Occurrence Records from Biological Collection Databases. Methods Ecol. Evol. 2019, 10, 744–751. [Google Scholar] [CrossRef]
- Hof, C.; Brändle, M.; Brandl, R. Lentic Odonates Have Larger and More Northern Ranges than Lotic Species. J. Biogeogr. 2006, 33, 63–70. [Google Scholar] [CrossRef]
- Olsen, K.; Svenning, J.-C.; Balslev, H. Niche Breadth Predicts Geographical Range Size and Northern Range Shift in European Dragonfly Species (Odonata). Diversity 2022, 14, 719. [Google Scholar] [CrossRef]
- Davis Rabosky, A.R.; Cox, C.L.; Rabosky, D.L.; Title, P.O.; Holmes, I.A.; Feldman, A.; McGuire, J.A. Coral Snakes Predict the Evolution of Mimicry across New World Snakes. Nat. Commun. 2016, 7, 11484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barve, N.; Barve, V.; Jiménez-Valverde, A.; Lira-Noriega, A.; Maher, S.P.; Peterson, A.T.; Soberón, J.; Villalobos, F. The Crucial Role of the Accessible Area in Ecological Niche Modeling and Species Distribution Modeling. Ecol. Model. 2011, 222, 1810–1819. [Google Scholar] [CrossRef]
- Steen, V.A.; Elphick, C.S.; Tingley, M.W. An Evaluation of Stringent Filtering to Improve Species Distribution Models from Citizen Science Data. Divers. Distrib. 2019, 25, 1857–1869. [Google Scholar] [CrossRef]
- Hijmans, R.J. Package ‘Raster’—Geographic Data Analysis and Modeling; CRAN Repos. 2019. Available online: https://rspatial.org/raster (accessed on 10 May 2022).
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the Black Box: An Open-Source Release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling; R Package Version 1.1-4; Cran. 2017. Available online: https://cran.r-project.org/web/packages/dismo/dismo.pdf (accessed on 10 May 2022).
- Merow, C.; Smith, M.J.; Silander, J.A. A Practical Guide to MaxEnt for Modeling Species’ Distributions: What It Does, and Why Inputs and Settings Matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Naimi, B.; Hamm, N.A.S.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where Is Positional Uncertainty a Problem for Species Distribution Modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Kass, J.M.; Muscarella, R.; Galante, P.J.; Bohl, C.L.; Pinilla-Buitrago, G.E.; Boria, R.A.; Soley-Guardia, M.; Anderson, R.P. ENMeval 2.0: Redesigned for Customizable and Reproducible Modeling of Species’ Niches and Distributions. Methods Ecol. Evol. 2021, 12, 1602–1608. [Google Scholar] [CrossRef]
- Calabrese, J.M.; Certain, G.; Kraan, C.; Dormann, C.F. Stacking Species Distribution Models and Adjusting Bias by Linking Them to Macroecological Models. Glob. Ecol. Biogeogr. 2014, 23, 99–112. [Google Scholar] [CrossRef]
- González-Orozco, C.E.; Laffan, S.W.; Miller, J.T. Spatial Distribution of Species Richness and Endemism of the Genus Acacia in Australia. Aust. J. Bot. 2011, 59, 601–609. [Google Scholar] [CrossRef]
- Earl, C.; Belitz, M.W.; Laffan, S.W.; Barve, V.; Barve, N.; Soltis, D.E.; Allen, J.M.; Soltis, P.S.; Mishler, B.D.; Kawahara, A.Y.; et al. Spatial phylogenetics of butterflies in relation to environmental drivers and angiosperm diversity across North America. iScience 2021, 24, 102239. [Google Scholar] [CrossRef] [PubMed]
- Sandall, E.L.; Pinkert, S.; Jetz, W. Country-level checklists and occurrences for the world’s Odonata (dragonflies and damselflies). J. Biogeogr. 2022, 49, 1586–1598. [Google Scholar] [CrossRef]
- Yanybaeva, V.A.; Dumont, H.J.; Haritonov, A.Y.; Popova, O.N. The Odonata of the South Ural, Russia, with special reference to Ischnura aralensis Haritonov, 1979. Odonatologica 2006, 35, 167–185. [Google Scholar]
- Haritonov, A.Y.; Eremina, E.E. The dragonflies (Odonata) of South Ural: The value of regional faunistic research. Euroasian Entomol. J. 2010, 9, 263–273. [Google Scholar]
- Kalkman, V.J.; Boudot, J.-P.; Bernard, R.; Conze, K.-J.; De Knijf, G.; Dyatlova, E.; Ferreira, S.; Jović, M.; Ott, J.; Riservato, E.; et al. European Red List of Dragonflies; IUCN: Gland, Switzerland; Cambridge, UK; Office for Official Publications of the European Communities: Luxembourg, 2010. [Google Scholar]
- Riservato, E.; Boudot, J.-P.; Ferreira, S.; Jović, M.; Kalkman, V.J.; Schneider, W.; Samraoui, B.; Cuttelod, A. The Status and Distribution of Dragonflies of the Mediterranean Basin; IUCN Red List of Threatened Species, Regional Assessments Series; IUCN: Gland, Switzerland; Malaga, Spain, 2009. [Google Scholar]
- Samraoui, B.; Boudot, J.-P.; Ferreira, S.; Riservato, E.; Jović, M.; Kalkman, V.J.; Schneider, W. The Status and Distribution of Dragonflies (Odonata). In The Status and Distribution of Freshwater Biodiversity in Northern Africa; IUCN Red List of Threatened Species, Regional Assessments Series; Garcia, N., Cuttelod, A., Abdul Malak, D., Eds.; IUCN: Gland, Switzerland; Malaga, Spain, 2010; Chapter 5; pp. 51–70. [Google Scholar]
- Kosterin, O.; Malikova, E.I.; Haritonov, A.Y. Critical species of Odonata in the Asian part of the former USSR and the Republic of Mongolia. Int. J. Odonatol. 2004, 7, 341–370. [Google Scholar] [CrossRef]
- Karube, H. Conservation of endangered dragonflies. Nat. Insects 2020, 55, 2–3. [Google Scholar]
- Huang, C.; Ebach, M.; Ahyong, S.T. Bioregionalisation of the freshwater zoogeographical areas of mainland China. Zootaxa 2020, 4742, 271–298. [Google Scholar] [CrossRef]
- Kohli, M.; Djernæs, M.; Sanchez Herrera, M.; Sahlen, G.; Pilgrim, E.; Simonsen, T.J.; Olsen, K.; Ware, J. Comparative phylogeography uncovers evolutionary past of Holarctic dragonflies. PeerJ 2021, 9, e11338. [Google Scholar] [CrossRef]
- Waltari, E.; Hijmans, R.J.; Peterson, A.T.; Nyári, A.S.; Perkins, S.L.; Guralnick, R.P. Locating Pleistocene Refugia: Comparing Phylogeographic and Ecological Niche Model Predictions. PLoS ONE 2007, e563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husemann, M.; Schmitt, T.; Zachos, F.E.; Ulrich, W.; Habel, J.C. Palaearctic biogeography revisited: Evidence for the existence of a North African refugium for Western Palaearctic biota. J Biogeogr. 2014, 41, 81–94. [Google Scholar] [CrossRef]
- Harrison, S.; Noss, R. Endemism hotspots are linked to stable climatic refugia. Ann Bot. 2017, 119, 207–214. [Google Scholar] [CrossRef]
- De Knijf, G.; Flenker, U.; Vanappelghem, C.; Manci, C.O.; Kalkman, V.J.; Demolder, H. The status of two boreo-alpine species, Somatochlora alpestris and S. arctica, in Romania and their vulnerability to the impact of climate change (Odonata: Corduliidae). Int. J. Odonatol. 2011, 14, 111–126. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Kalkman, V.J. Changing temperature regimes have advanced the phenology of Odonata in the Netherlands. Ecol. Entomol. 2008, 33, 394–402. [Google Scholar] [CrossRef]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Thomas, C.D. A northward shift of range margins in British Odonata. Glob. Change Biol. 2005, 11, 502–506. [Google Scholar] [CrossRef]
- Hassall, C.; Thompson, D.J.; French, G.C.; Harvey, I.F. Historical changes in the phenology of British Odonata are related to climate. Glob. Change Biol. 2007, 13, 933–941. [Google Scholar] [CrossRef]
- Doi, H. Delayed phenological timing of dragonfly emergence in Japan over five decades. Biol. Lett. 2008, 4, 388–391. [Google Scholar] [CrossRef]
- Aoki, T. Northward expansion of Ictinogomphus pertinax (Selys, 1854) in Japan. Nat. Insects 2021, 56, 12–17. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioclimatic Variable | Description | Mean Permutation Contribution |
|---|---|---|
| Bio 2 | Mean diurnal range | 15.8 |
| Bio 4 | Temperature seasonality | 12.0 |
| Bio 1 | Annual mean temperature | 11.6 |
| Bio 5 | Max. temperature of warmest month | 10.7 |
| Bio 15 | Precipitation seasonality | 9.2 |
| Bio 8 | Mean temperature of wettest quarter | 7.2 |
| Bio 9 | Mean temperature of driest quarter | 6.9 |
| Bio 13 | Precipitation of wettest month | 5.7 |
| Bio 12 | Annual precipitation | 5.3 |
| Bio 6 | Min. temperature of coldest month | 5.1 |
| Bio 14 | Precipitation of driest month | 4.5 |
| Bio 17 | Precipitation of driest quarter | 3.2 |
| Bio 16 | Precipitation of wettest quarter | 2.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalkman, V.J.; Boudot, J.-P.; Futahashi, R.; Abbott, J.C.; Bota-Sierra, C.A.; Guralnick, R.; Bybee, S.M.; Ware, J.; Belitz, M.W. Diversity of Palaearctic Dragonflies and Damselflies (Odonata). Diversity 2022, 14, 966. https://doi.org/10.3390/d14110966
Kalkman VJ, Boudot J-P, Futahashi R, Abbott JC, Bota-Sierra CA, Guralnick R, Bybee SM, Ware J, Belitz MW. Diversity of Palaearctic Dragonflies and Damselflies (Odonata). Diversity. 2022; 14(11):966. https://doi.org/10.3390/d14110966
Chicago/Turabian StyleKalkman, Vincent J., Jean-Pierre Boudot, Ryo Futahashi, John C. Abbott, Cornelio A. Bota-Sierra, Robert Guralnick, Seth M. Bybee, Jessica Ware, and Michael W. Belitz. 2022. "Diversity of Palaearctic Dragonflies and Damselflies (Odonata)" Diversity 14, no. 11: 966. https://doi.org/10.3390/d14110966
APA StyleKalkman, V. J., Boudot, J. -P., Futahashi, R., Abbott, J. C., Bota-Sierra, C. A., Guralnick, R., Bybee, S. M., Ware, J., & Belitz, M. W. (2022). Diversity of Palaearctic Dragonflies and Damselflies (Odonata). Diversity, 14(11), 966. https://doi.org/10.3390/d14110966