
Pseudocercospora rizhaoensis sp. nov. Causing Leaf Spot Disease of Ligustrum japonicum in China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Survey, Collection and Fungal Isolation
2.2. DNA Extraction, Sequencing and Phylogenetic Analyses
2.3. Morphological Identification and Characterization
2.4. Pathogenicity Testing
3. Results
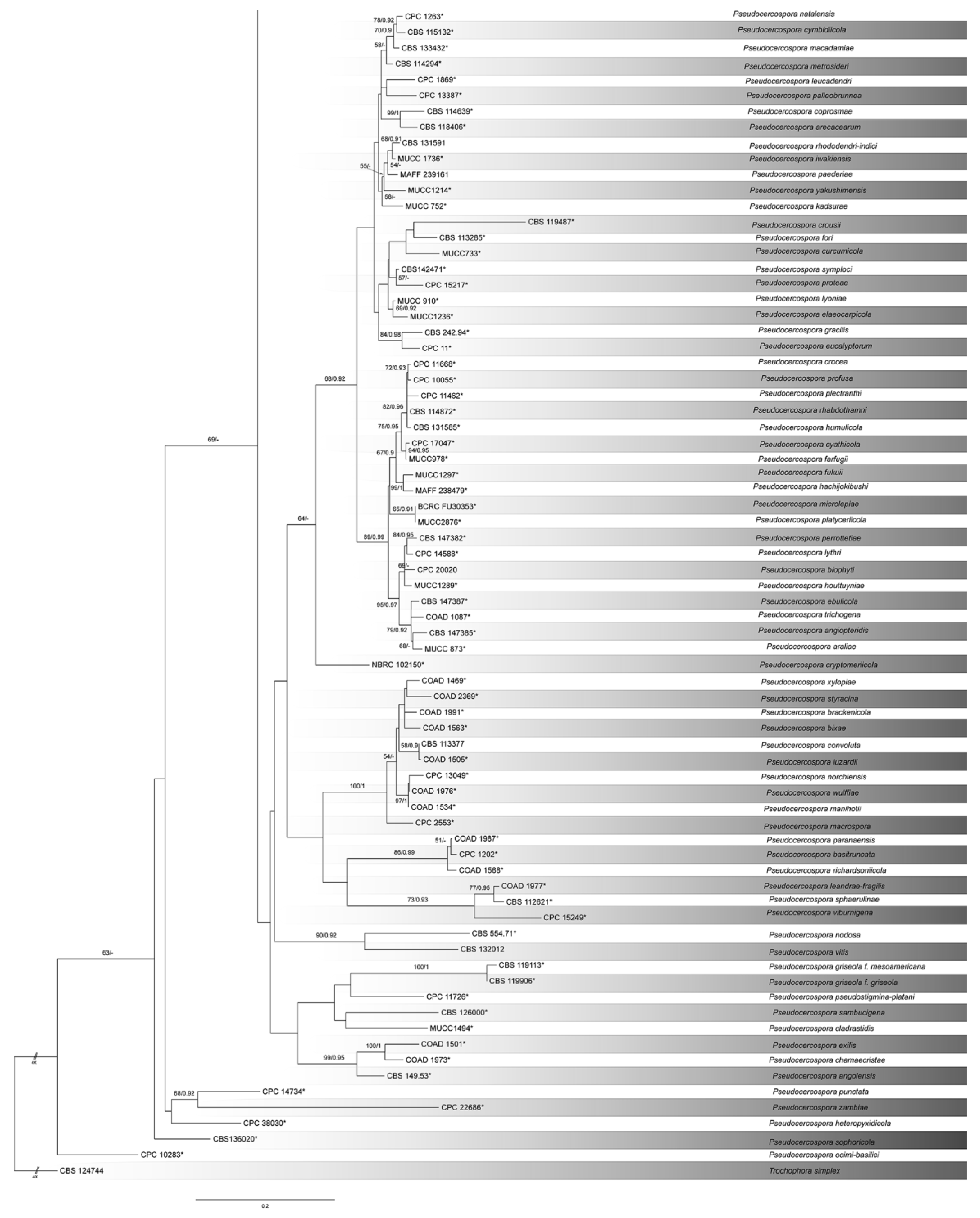
3.1. Phylogeny
3.2. Taxonomy
3.3. Pathogenicity Tests
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Crous, P.W.; Braun, U.; Hunter, G.C.; Wingfield, M.; Verkley, G.; Shin, H.-D.; Nakashima, C.; Groenewald, J. Phylogenetic lineages in Pseudocercospora. Stud. Mycol. 2013, 75, 37–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, U.; Nakashima, C.; Bakhshi, M.; Zare, R.; Shin, H.D.; Alves, R.F.; Sposito, M.B. Taxonomy and phylogeny of cercosporoid ascomycetes on Diospyros spp. with special emphasis on Pseudocercospora spp. Fungal Syst. Evol. 2020, 6, 95–127. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Schoch, C.L.; Hyde, K.D.; Wood, A.R.; Gueidan, C.; De Hoog, G.S.; Groenewald, J.Z. Phylogenetic lineages in the Capnodiales. Stud. Mycol. 2009, 64, 17–47. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, C.; Motohashi, K.; Chen, C.Y.; Groenewald, J.Z.; Crous, P.W. Species diversity of Pseudocercospora from Far East Asia. Mycol. Prog. 2016, 15, 1093–1117. [Google Scholar] [CrossRef]
- Osorio, J.A.; Wingfield, M.J.; de Beer, Z.W.; Roux, J. Pseudocercospora mapelanensis sp. nov., associated with a fruit and leaf disease of Barringtonia racemosa in South Africa. Australas. Plant Pathol. 2015, 44, 349–359. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Verma, S.K.; Singh, V.K.; Singh, R.; Singh, A.; Kumar, S. Morphology and phylogeny of a new species, Pseudocercospora haldinae (Mycosphaerellaceae) on Haldina cordifolia from India. Phytotaxa 2021, 501, 281–292. [Google Scholar] [CrossRef]
- Silva, M.; Barreto, R.W.; Pereira, O.L.; Freitas, N.; Groenewald, J.; Crous, P. Exploring fungal megadiversity: Pseudocercospora from Brazil. Persoonia 2016, 37, 142–172. [Google Scholar] [CrossRef] [Green Version]
- Shivas, R.G.; Marney, T.S.; Tan, Y.P.; McTaggart, A.R. Novel species of Cercospora and Pseudocercospora (Capnodiales, Mycosphaerellaceae) from Australia. Fungal Biol. 2015, 119, 362–369. [Google Scholar] [CrossRef] [Green Version]
- Kirschner, R.; Wang, H. New species and records of mycosphaerellaceous fungi from living fern leaves in East Asia. Mycol. Prog. 2015, 14, 65. [Google Scholar] [CrossRef]
- Kimunye, J.N.; Were, E.; Mussa, F.; Tazuba, A.; Jomanga, K.; Viljoen, A.; Swennen, R.; Muthoni, F.K.; Mahuku, G. Distribution of Pseudocercospora species causing Sigatoka leaf diseases of banana in Uganda and Tanzania. Plant Pathol. 2020, 69, 50–59. [Google Scholar] [CrossRef]
- Araujo, L.; Pinto, F.A.M.F.; de Andrade, C.C.L.; Gomes, L.B.; Mituti, T.; Duarte, V. Pseudocercospora actinidiae causes sooty spot disease on kiwifruit in Santa Catarina, Brazil. Australas. Plant Dis. Notes 2021, 16, 22. [Google Scholar] [CrossRef]
- Rezene, Y.; Tesfaye, K.; Clare, M.; Gepts, P. Pathotypes characterization and virulence diversity of Pseudocercospora griseola the causal agent of angular leaf spot disease collected from major common bean (Phaseolus vulgaris L.) growing areas of Ethiopia. J. Plant Pathol. Microbiol. 2018, 9, 1000445. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Ma, C.Y.; Xue, H.; Piao, C.G.; Li, Y. A new Arthrinium-like genus of Amphisphaeriales in China. MycoKeys 2022, 92, 27–43. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Voglmayr, H.; Bian, D.R.; Piao, C.G.; Wang, S.K.; Li, Y. Morphology and phylogeny of Gnomoniopsis (Gnomoniaceae, Diaporthales) from fagaceae leaves in China. J. Fungi 2021, 7, 792. [Google Scholar] [CrossRef]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschner, R.; Okuda, T. A new species of Pseudocercospora and new record of Bartheletia paradoxa on leaves of Ginkgo biloba. Mycol. Prog. 2013, 12, 421–426. [Google Scholar] [CrossRef]
- Chen, Q.; Bakhshi, M.; Balci, Y.; Broders, K.; Cheewangkoon, R.; Chen, S.; Fan, X.; Gramaje, D.; Halleen, F.; Jung, M.H.; et al. Genera of phytopathogenic fungi: GOPHY 4. Stud. Mycol. 2022, 101, 417–564. [Google Scholar] [CrossRef] [PubMed]
- Videira, S.I.R.; Groenewald, J.Z.; Nakashima, C.; Braun, U.; Barreto, R.W.; de Wit, P.J.; Crous, P.W. Mycosphaerellaceae—Chaos or clarity? Stud. Mycol. 2017, 87, 257–421. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Z.C.; He, W.; Zhang, Y. Pseudocercospora spp. from leaf spots of Euonymus japonicus in China. Mycosystema 2019, 38, 159–170. [Google Scholar]
- Bakhshi, M.; Arzanlou, M.; Babai-Ahari, A.; Groenewald, J.Z.; Crous, P.W. Multi-gene analysis of Pseudocercospora spp. from Iran. Phytotaxa 2014, 184, 245–264. [Google Scholar] [CrossRef]
- Deighton, F.C. Studies on Cercospora and allied genera. VI. Pseudocercospora Speg., Pantospora Cif., and Cercoseptoria Petr. Mycol. Pap. 1976, 140, 1–168. [Google Scholar]
- Wang, S.H.; Liu, S.M. First Report of leaf spot caused by Pseudocercospora ligustri on Ligustrum japonicum ‘Howardii’ in China. Plant Dis. 2019, 103, 153. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Isolates | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| LSU | ITS | act | tef1 | rpb2 | ||
| Pseudocercospora abeliae | MUCC1674 * | NA | LC599330 | LC599407 | LC599448 | LC599587 |
| P. aeschynomenicola | CPC 25227 = COAD 1972 * | KT290173 | KT290146 | KT313501 | KT290200 | NA |
| P. airliensis | BRIP 58550 * | KM055433 | KM055429 | NA | KM055436 | NA |
| P. aleuritis | MAFF237174 = MUCC1230 * | NA | LC599331 | LC599408 | LC599449 | LC599588 |
| P. amelanchieris | MAFF 237782 = MUCC885 * | NA | KX462583 | KX462550 | KX462669 | KX462616 |
| P. ampelopsis | CBS 131583 = CPC 11680 * | GU253846 | GU269830 | GU320534 | GU384542 | NA |
| P. angiopteridis | CBS 147385 * | NA | LC599332 | LC599409 | LC599450 | LC599589 |
| P. angolensis | CBS 149.53 * | JQ324941 | JQ324975 | JQ325011 | JQ324988 | NA |
| P. araliae | MUCC 873 * | GU253702 | GU269653 | GU320361 | GU384371 | KX462617 |
| P. arecacearum | CBS 118406 * | GU253704 | GU269655 | GU320363 | GU384373 | NA |
| P. assamensis | CBS 122467 * | GU253705 | GU269656 | GU320364 | GU384374 | NA |
| P. avicenniae | CBS 146479 * | NA | GU188047 | LC599410 | LC599451 | LC599590 |
| P. basiramifera | CBS 111072 = CPC 1266 * | GU253709 | GU269661 | GU320368 | DQ211677 | NA |
| P. basitruncata | CBS 114664 = CPC 1202 * | GU253710 | DQ267600 | DQ147622 | DQ211675 | NA |
| P. biophyti | CPC 20020 | NA | LC599333 | LC599411 | LC599452 | LC599591 |
| P. bixae | CPC 25244 = COAD 1563 * | KT290180 | KT290153 | KT313508 | KT290207 | NA |
| P. brackenicola | CPC 24695 = COAD 1991 * | KT037565 | KT037524 | KT037606 | KT037484 | NA |
| P. breonadiae | CBS 143489 = CPC 30153 * | MH107959 | MH107913 | MH107985 | MH108026 | MH108006 |
| P. bruceae | MUCC 2875 * | NA | LC599334 | LC599412 | LC599453 | NA |
| P. casuarinae | CBS 128218 * | HQ599604 | HQ599603 | LC599413 | LC599454 | NA |
| P. ceratoniae | CBS 147386 * | NA | LC599335 | LC599414 | LC599455 | LC599592 |
| P. cercidicola | MAFF 237791 = MUCC 896 * | GU253719 | GU269671 | GU320377 | GU384388 | KX462618 |
| P. cercidis-chinensis | CBS 132109 = CPC 14481 * | JX901884 | GU269670 | GU320376 | GU384387 | LC599593 |
| P. chamaecristae | CPC 25228 = COAD 1973 * | KT290174 | KT290147 | KT313502 | KT290201 | NA |
| P. chiangmaiensis | CBS 123244 * | NG042738 | EU882113 | KF903544 | KF903177 | NA |
| P. chibaensis | MUCC1670E * | NA | KX462584 | KX462551 | KX462670 | KX462619 |
| P. chionanthi-retusi | TUA50 = NCHUPP L1605 * | NA | KX462585 | KX462552 | KX462671 | KX462620 |
| P. cladrastidis | MUCC1494 * | NA | LC599336 | LC599415 | LC599457 | LC599594 |
| P. convoluta | CBS 113377 * | MF951226 | DQ676519 | NA | NA | MF951617 |
| P. coprosmae | CBS 114639 * | JQ324946 | GU269680 | GU320386 | GU384397 | NA |
| P. cordiana | CPC 2552 * | GU214472 | AF362054 | GU320387 | GU384398 | NA |
| P. corylopsidis | MAFF 237795 = MUCC 908 * | NG069064 | GU269684 | GU320390 | GU384401 | KX462621 |
| P. cotini | MAFF410088 = MUCC1415 * | NA | LC599337 | LC599416 | LC599458 | LC599596 |
| P. cotoneastri | MAFF 410089 = MUCC1416 * | NA | KX462586 | KX462553 | KX462672 | KX462622 |
| P. crispans | CBS 125999 = CPC 14883 * | GU253825 | GU269807 | GU320510 | GU384518 | KX462623 |
| P. crocea | CBS 126004 = CPC 11668 * | JQ324947 | GU269792 | GU320493 | GU384502 | NA |
| P. crousii | CBS 119487 | GQ852631 | GU269686 | GU320392 | GU384403 | NA |
| P. cryptomeriicola | MAFF240073 = NBRC 102150 * | NA | LC599338 | LC599418 | LC599460 | LC599598 |
| P. curcumicola | MUCC733 * | NA | LC599339 | LC599419 | LC599461 | LC599599 |
| P. cyathicola | CBS 129520 = CPC 17047 * | JF951159 | JF951139 | KX462554 | KX462673 | KX462624 |
| P. cymbidiicola | CBS 115132 * | GU253733 | GU269692 | GU320397 | GU384408 | NA |
| P. dalbergiae | TUA55 * | NA | LC599340 | LC599420 | LC599462 | LC599600 |
| P. daphniphylli | MAFF 410009 = MUCC1399 * | NA | KX462587 | KX462555 | KX462674 | KX462625 |
| P. davidiicola | MAFF 240281 = MUCC296 * | GU253734 | GU269693 | GU320398 | GU384409 | KX462626 |
| P. delonicicola | MUCC2869 * | NA | LC599341 | LC599421 | LC599463 | LC599601 |
| P. dingleyae | CBS 114645 * | KX286997 | KX287299 | NA | NA | KX288454 |
| P. diplusodonii | CPC 25179 = COAD 1476 * | KT290162 | KT290135 | KT313490 | KT290189 | NA |
| P. dodonaeae | CBS 114647 * | JQ324948 | GU269697 | JQ325013 | GU384413 | NA |
| P. dovyalidis | CBS 126002 = CPC 13771 * | GU253818 | GU269800 | GU320503 | GU384513 | NA |
| P. ebulicola | CBS 147387 * | NA | LC599342 | LC599422 | NA | NA |
| P. elaeocarpicola | MAFF 237189 = MUCC1236 * | NA | KX462588 | KX462556 | KX462675 | KX462627 |
| P. emmoticola | CPC 25187 = COAD 1491 * | KT290163 | KT290136 | KT313491 | KT290190 | NA |
| P. eriobotryae | MUCC 1007 * | NA | KX462589 | KX462557 | KX462676 | KX462628 |
| P. eriobotryicola | TUA12 = NCHUPPL1601 * | NA | KX462590 | KX462558 | KX462677 | KX462629 |
| P. ershadii | CBS 136114 = CCTU 1206 * | KP717032 | KM452867 | KM452844 | KM452889 | MN786459 |
| P. eucalyptorum | CBS 114866 = CPC 11 * | JQ739817 | KF901720 | KF903474 | KF903195 | MF951618 |
| P. eumusae | CBS 114824 * | NA | EU514238 | NA | NA | NA |
| P. euonymi-japonici | CGMCC 3.18576 * | NA | MH255812 | NA | NA | MH392531 |
| P. eupatoriella | CBS 113372 * | GU253743 | GU269704 | GU320408 | GU384420 | NA |
| P. eupatorii- formosani | TUA59 = NCHUPP L1606 * | NA | KX462591 | KX462559 | KX462678 | KX462630 |
| P. euphorbiacearum | COAD 1537 * | KT290172 | KT290145 | KT313500 | KT290199 | NA |
| P. exilis | CPC 25193 = COAD 1501 * | KT290166 | KT290139 | KT313494 | KT290193 | NA |
| P. farfugii | MUCC978 * | NA | LC599343 | LC599423 | LC599464 | LC599603 |
| P. fijiensis | CBS 120258 = CIRAD 86 * | JQ324952 | EU514248 | NA | NA | NA |
| P. flavomarginata | CBS 126001 * | NA | GU269804 | GU320507 | GU384515 | LC599604 |
| P. fori | CBS 113285 * | NA | AF468869 | KF903462 | NA | KT356874 |
| P. formosana | MUCC2612 * | NA | LC599344 | LC599424 | LC599466 | LC599605 |
| P. forsythiae | MAFF 410087 = MUCC1414 * | NA | LC599345 | LC599425 | LC599467 | NA |
| P. fukuii | MAFF238121 = MUCC1297 * | NA | LC599347 | LC599427 | LC599469 | LC599607 |
| P. fukuokaensis | MAFF 237768 = MUCC 887 * | GU253751 | GU269714 | GU320418 | GU384430 | KX462632 |
| P. ginkgoana | R. Kirschner 3563 (TNM) * | NA | JX134048 | NA | NA | NA |
| P. glochidionis | MAFF 237000; MUCC1211 * | NA | LC599348 | LC599428 | LC599470 | LC599608 |
| P. gracilis | CBS 242.94 * | NA | DQ267582 | NA | DQ211666 | NA |
| P. griseola f. griseola | CBS 119906 * | NA | DQ289812. | NA | NA | NA |
| P. griseola f. mesoamericana | CBS 119113 * | NA | DQ289824 | NA | NA | NA |
| P. hachijokibushii | MAFF 238479 * | NA | KX462593 | KX462561 | KX462680 | KX462633 |
| P. haiweiensis | CBS 131584 = CPC 14084 * | GU253821 | GU269803 | GU320506 | GU384514 | KX462634 |
| P. hardenbergiae | CBS 147381 * | NA | LC599349 | LC599429 | LC599471 | LC599609 |
| P. heteropyxidicola | CBS 146082 = CPC 38030 * | NA | MN562151 | MN556791 | NA | NA |
| P. hiratsukana | MAFF 238300 = MUCC1105 * | NA | KX462594 | KX462562 | KX462681 | KX462635 |
| P. houttuyniae | MAFF 238071 = MUCC1289 * | NA | KX462595 | KX462563 | KX462682 | KX462636 |
| P. humuli | MUCC 742 * | GU253758 | GU269725 | GU320428 | GU384439 | KX462637 |
| P. humulicola | CBS 131585 * | JQ324956 | GU269723 | GU320427 | GU384438 | NA |
| P. imazekii | MUCC 1668 * | NA | KX462596 | KX462564 | KX462683 | KX462638 |
| P. indonesiana | CBS 122473 * | NA | GU269735 | GU320437 | GU384448 | NA |
| P. iwakiensis | MUCC 1736 * | NA | KX462607 | KX462574 | KX462693 | KX462657 |
| P. ixoriana | MUCC2608 * | NA | LC599350 | LC599430 | LC599472 | LC599610 |
| P. izuohshimense | MAFF 238478 = MUCC1336 * | NA | KX462597 | KX462565 | KX462684 | KX462639 |
| P. jagerae | BRIP 58549 * | NA | KM055431 | NA | KM055438 | NA |
| P. kadsurae | MUCC 752 * | NA | KX462598 | KX462566 | KX462685 | KX462640 |
| P. kaki | MAFF 238214 * | GU253761 | LC512001 | LC512007 | LC515783 | LC515794 |
| P. kakiicola | MAFF 238238 = MUCC 900 * | NA | GU269729 | GU320431 | GU384442 | NA |
| P. kenyirana | MUCC 2873 * | NA | LC599351 | LC599431 | LC599473 | NA |
| P. kiggelariae | CBS 132016 = CPC 11853 * | GU253762 | GU269730 | GU320432 | GU384443 | NA |
| P. kobayashiana | MAFF 236999 * | NA | LC511998 | LC512004 | LC515780 | LC515791 |
| P. leandrae-fragilis | COAD 1977 * | NA | KY574288 | NA | NA | NA |
| P. leucadendri | CPC 1869 * | GU214480 | GU269842 | GU320545 | GU384555 | NA |
| P. liquidambaricola | MAFF410455 * | NA | LC599352 | LC599432 | LC599474 | LC599611 |
| P. longispora | CBS 122470 * | NA | GU269734 | GU320436 | GU384447 | NA |
| P. lonicericola | MUCC 889 = MAFF 237785 * | GU253766 | GU269736 | GU320438 | JQ324999 | KX462641 |
| P. luzardii | CPC 25196 = COAD 1505 * | KT290167 | KT290140 | KT313495 | KT290194 | NA |
| P. lyoniae | MAFF 237775 = MUCC 910 * | GU253768 | GU269739 | GU320441 | GU384451 | KX462642 |
| P. lythri | CBS 132115 = CPC 14588 * | NA | GU269742 | GU320444 | GU384454 | LC599612 |
| P. macadamiae | CBS 133432 * | KX286998 | KX287300 | KU878551 | KU878504 | KX288455 |
| P. macrospora | CBS 114696 = CPC 25538 | GU214478 | AF362055 | GU320447 | GU384457 | NA |
| P. madagascariensis | CBS 124155 * | NA | GQ852767 | KF253625 | KF253265 | KX462643 |
| P. maetaengensis | MFLUCC 14-0411 * | NA | MN648323 | NA | NA | NA |
| P. mangifericola | BRIP 52776b * | NA | GU188048 | NA | NA | NA |
| P. manihotii | CPC 25219 = COAD 1534 * | KT290171 | KT290144 | KT313499 | KT290198 | NA |
| P. mapelanensis | CMW40581 * | KM203121 | KM203118 | KM203127 | KM203124 | NA |
| P. marginalis | CBS 131582 = CPC 12497 * | GU253812 | GU269794 | GU320495 | GU384504 | NA |
| P. mazandaranensis | CCTU 1102 = CBS 136115 * | KP717020 | KM452854 | KM452831 | KM452876 | LC599613 |
| P. metrosideri | CBS 114294 * | KX286999 | KX287301 | NA | NA | KX288456 |
| P. microlepiae | BCRC FU30353 * | NA | KR348740 | NA | NA | NA |
| P. musae | CBS 116634 * | GU253775 | GU269747 | GU320449 | GU384459 | NA |
| P. naitoi | MAFF 237906 = MUCC1072 * | NA | KX462599 | KX462567 | KX462686 | KX462644 |
| P. nandinae | MAFF 237633 = MUCC1260 * | NA | KX462600 | KX462568 | KX462687 | KX462645 |
| P. natalensis | CBS 111069 = CPC 1263 * | DQ267576 | DQ303077 | DQ147620 | JQ325000 | NA |
| P. nelumbonicola | BCRC FU30367 * | NA | KY304492 | NA | NA | NA |
| P. neriicola | CBS 138010 = CPC 23765 * | KJ869222 | KJ869165 | KJ869231 | KJ869240 | KX462647 |
| P. nodosa | CBS 554.71 * | MF951227 | MF951367 | NA | NA | MF951620 |
| P. norchiensis | CBS 120738 = CPC 13049 * | GU253780 | EF394859 | GU320455 | GU384464 | KX462648 |
| P. ocimi-basilici | CPC 10283 * | NA | GU269754 | GU320456 | GU384465 | NA |
| P. paederiae | MAFF 239161 | NA | KX462603 | KX462570 | KX462689 | KX462651 |
| P. palleobrunnea | CBS 124771 = CPC 13387 * | GQ303319 | GQ303288 | GU320500 | GU384509 | KX462652 |
| P. pancratii | CBS 137.94 * | GU253784 | GU269759 | GU320460 | GU384470 | NA |
| P. paranaensis | CPC 24680 = COAD 1987T | KT037563 | KT037522 | KT037604 | KT037482 | NA |
| P. parapseudarthriae | CBS 137996 = CPC 23449 * | KJ869208 | KJ869151 | KJ869229 | KJ869238 | NA |
| P. perae | CPC 25171 = COAD 1465 * | KT290159 | KT290132 | KT313487 | KT290186 | NA |
| P. perrottetiae | CBS 147382 * | NA | LC599353 | LC599433 | LC599477 | LC599614 |
| P. photiniae | MUCC 1661 * | NA | KX462604 | KX462571 | KX462690 | KX462653 |
| P. pini-densiflorae | MUCC 1714 * | NA | LC599354 | LC599434 | LC599478 | LC599615 |
| P. planaltinensis | CPC 25189 = COAD 1495 * | KT290164 | KT290137 | KT313492 | KT290191 | NA |
| P. platyceriicola | MUCC2876 * | NA | LC599355 | LC599435 | LC599479 | LC599616 |
| P. plectranthi | CBS 131586 = CPC 11462 * | NG070621 | GU269791 | GU320492 | GU384501 | NA |
| P. plumeriifolii | CPC 25191 = COAD 1498 * | KT290165 | KT290138 | KT313493 | KT290192 | NA |
| P. pothomorphes | CPC 25166 = COAD 1450 * | KT290158 | KT290131 | KT313486 | KT290185 | NA |
| P. profusa | CBS 132306 = CPC 10055 * | GU253787 | GU269762 | GU320463 | GU384473 | NA |
| P. proiphydis | BRIP 58545 * | KM055434 | KM055430 | NA | KM055437 | NA |
| P. proteae | CBS 131587 = CPC 15217 * | MH877381 | GU269808 | GU320511 | GU384519/ | NA |
| P. pruni-grayanae | MUCC 1715 * | NA | LC599356 | NA | LC599481 | LC599618 |
| P. pseudomusae | CBS 147147 * | NA | MW063423 | MW070772 | MW071091 | MW070919 |
| P. pseudomyrticola | CBS 145554 = CPC 35448 * | MK876446 | MK876405 | MK876461 | MK876499 | MK876490 |
| P. pseudostigminaplatani | CBS 131588 = CPC 11726 * | JQ324963 | GU269857 | GU320560 | GU384568 | NA |
| P. punctata | CBS 132116 = CPC 14734 * | GU253791 | GU269765 | GU320468 | GU384477 | MF951622 |
| P. punicae | MAFF236998 = MUCC 1209 | NA | KX462606 | KX462573 | KX462692 | KX462655 |
| P. pyracanthae | MAFF237140 = MUCC 1226 * | GU253792 | GU269767 | NA | GU384479 | LC599619 |
| P. pyracanthigena | CBS 131589 = CPC 10808 * | NA | GU269766 | GU320469 | GU384478 | NA |
| P. ravenalicola | CBS 122468 * | GU253828 | GU269810 | GU320513 | GU384521 | NA |
| P. rhabdothamni | CBS 114872 * | JQ324964 | GU269768 | GU320471 | GU384480 | NA |
| P. rhamnellae | CBS 131590 = CPC 12500 * | GU253813 | GU269795 | GU320496 | GU384505 | NA |
| P. rhapisicola | MAFF305042 = MUCC1484 * | NA | LC599357 | LC599436 | LC599483 | LC599620 |
| P. rhododendri-indici | CBS 131591 = CPC 10822 * | JQ324965 | GU269722 | GU320426 | NA | NA |
| P. riachueli var. horiana | MUCC2141 * | NA | LC599358 | LC599437 | LC599484 | LC599621 |
| P. richardsoniicola | CPC 25248 = COAD 1568 * | KT290181 | KT290154 | KT313509 | KT290208 | NA |
| P. rigidae | CPC 25175 = COAD 1472 * | KT290161 | KT290134 | KT313489 | KT290188 | NA |
| P. rosae | MFLUCC 14-0408 * | MG829063 | MG828952 | NA | NA | NA |
| P. rizhaoensis | CFCC 57581 * | NA | OP661350 | OP651770 | OP651772 | OP651774 |
| P. rizhaoensis | CFCC 57582 | NA | OP661351 | OP651771 | OP651773 | OP651775 |
| P. sambucigena | CBS 126000 * | GU253809 | GU269788 | GU320508 | GU384498 | NA |
| P. sawadae | MAFF 239714 | NA | LC599359 | LC599438 | LC599485 | LC599622 |
| P. schizolobii | CBS 120029 = CPC 12962 * | KF251826 | KF251322 | KF253628 | KF253269 | NA |
| P. sennae-multijugae | CPC 25206 = COAD 1519 * | KT290169 | KT290142 | KT313497 | KT290196 | NA |
| P. serpocaulonicola | CPC 25077 = COAD 1866 * | KT037566 | KT037525 | KT037607 | KT037485 | NA |
| P. solani-pseudocapsicicola | CPC 25229 = COAD 1974 * | KT290175 | KT290148 | KT313503 | KT290202 | NA |
| P. sophoricola | CCTU 1037 = CBS 136020 * | KP717027 | KM452861 | KM452838 | KM452883 | MW272931 |
| P. sphaerulinae | CBS 112621 * | KF901958 | KF901625 | NA | KF903215 | NA |
| P. stemonicola | MUCC2874 * | NA | LC599360 | LC599439 | LC599487 | NA |
| P. stephanandrae | MAFF237799 = MUCC914 * | GU253831 | GU269814 | GU320516 | GU384526 | KX462658 |
| P. stranvaesiae | MAFF410090 = MUCC1417 * | NA | LC599361 | LC599440 | LC599488 | LC599623 |
| P. struthanthi | CPC 25199 = COAD 1512 * | KT290168 | KT290141 | KT313496 | KT290195 | NA |
| P. styracina | COAD 2369 * | MH480643 | MH397664 | MH480641 | MH480642 | NA |
| P. symploci | NCHUPP L1685 = CBS142471 * | NA | LC599362 | LC599441 | LC599489 | LC599624 |
| P. tabernaemontanae | CPC 19198 * | NA | LC599363 | LC599442 | NA | LC599625 |
| P. tereticornis | CBS 125214 = CPC 13299 * | NA | GQ852770 | GU320499 | GU384508 | KX462659 |
| P. terengganuensis | MUCC2871 * | NA | LC599364 | LC599443 | LC599490 | NA |
| P. tinea | TUA40 = NCHUPP L1603 * | NA | KX462608 | KX462577 | KX462696 | KX462660 |
| P. togashiana | MAFF410006 * | NA | LC599365 | LC599444 | LC599491 | LC599626 |
| P. trichogena | CPC 24664 = COAD 1087 * | KT037560 | KT037519 | KT037601 | KT037479 | NA |
| P. trinidadensis | COAD 1756 * | NA | KT290157 | NA | KT290210 | NA |
| P. tumulosa | CBS 121158 * | NA | DQ530217 | NA | NA | NA |
| P. vassobiae | CPC 25251 = COAD 1572 * | KT290182 | KT290155 | KT313510 | NA | NA |
| P. viburnigena | CBS 125998 = CPC 15249 * | GU253827 | GU269809 | GU320512 | GU320512 | NA |
| P. violamaculans | MUCC 1660 * | NA | KX462610 | KX462579 | KX462698 | KX462662 |
| P. vitis | CBS 132012 =CPC 11595 | GU214483 | GU269829 | GU320533 | GU384541 | KX462663 |
| P. wulffiae | CPC 25232 = COAD 1976 * | KT290177 | KT290150 | KT313505 | KT290204 | NA |
| P. xanthocercidis | CBS 131593 = CPC 11665Iso * | JQ324971 | JQ324983 | JQ325026 | JQ325005 | NA |
| P. xenopunicae | CBS 147384 * | NA | LC599367 | LC599446 | LC599493 | LC599628 |
| P. xenosyzygiicola | MAFF237986 = MUCC1481 * | NA | KX462611 | KX462580 | KX462699 | KX462664 |
| P. xylopiae | CPC 25173 = COAD 1469 * | KT290160 | KT290133 | KT313488 | KT290187 | NA |
| P. yakushimensis | MAFF237025 = MUCC1214 * | NA | LC599368 | LC599447 | LC599494 | LC599629 |
| P. zambiae | CBS 136423 = CPC 22686 * | NA | KF777175 | NA | NA | MF951630 |
| P. zelkovae | MAFF 238237 = MUCC872 * | NA | GU269835 | GU320537 | GU384547 | KX462665 |
| Trochophora simplex | CBS 124744 | NA | NA | GU320568 | GU384580 | KX462666 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Guo, S.; Liu, J.; Yang, X. Pseudocercospora rizhaoensis sp. nov. Causing Leaf Spot Disease of Ligustrum japonicum in China. Diversity 2022, 14, 990. https://doi.org/10.3390/d14110990
Liu Y, Guo S, Liu J, Yang X. Pseudocercospora rizhaoensis sp. nov. Causing Leaf Spot Disease of Ligustrum japonicum in China. Diversity. 2022; 14(11):990. https://doi.org/10.3390/d14110990
Chicago/Turabian StyleLiu, Yun, Shumei Guo, Jin Liu, and Xiangli Yang. 2022. "Pseudocercospora rizhaoensis sp. nov. Causing Leaf Spot Disease of Ligustrum japonicum in China" Diversity 14, no. 11: 990. https://doi.org/10.3390/d14110990
APA StyleLiu, Y., Guo, S., Liu, J., & Yang, X. (2022). Pseudocercospora rizhaoensis sp. nov. Causing Leaf Spot Disease of Ligustrum japonicum in China. Diversity, 14(11), 990. https://doi.org/10.3390/d14110990