
The Evolution and Fossil Record of Palaeognathous Birds (Neornithes: Palaeognathae)


Abstract
:1. Introduction
2. Overview of the Palaeognath Fossil Record
2.1. Lithornithidae
2.1.1. North American Lithornithids
2.1.2. European Lithornithids
2.1.3. Systematics of Lithornithidae
2.2. African and Eurasian Palaeognaths: Struthioniformes
2.2.1. Eurasian Stem Struthionids
2.2.2. African and Eurasian Crown Struthionids
2.3. South American Palaeognaths: Rheiformes and Tinamiformes
2.3.1. Rheid Fossil Record
2.3.2. Tinamid Fossil Record
2.4. Australian Ratites: Casuariiformes
2.5. New Zealand Ratites: Apterygiformes and Dinornithiformes
2.5.1. Apterygid Fossil Record
2.5.2. Dinornithid Fossil Record
2.6. Malagasy Ratites: Aepyornithiformes
2.7. Antarctic Ratites
3. Molecular Phylogenetic Hypotheses of Palaeognath Interrelationships
Molecular Divergence Time Estimates
4. Key Gaps in the Palaeognath Fossil Record
4.1. Cretaceous Stem Palaeognaths
4.2. Stem Group Representatives of Extant Palaeognath Subclades
5. Reconstructing the Most Recent Common Ancestor of Palaeognaths
5.1. The Flight Apparatus of the Crown Palaeognath MRCA
5.2. Inferred Ecology of the Palaeognath MRCA and K-Pg Survivorship
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Billerman, S.; Keeney, B.; Rodewald, P.; Schulenberg, T. Birds of the World. Available online: https://birdsoftheworld.org/bow/home (accessed on 30 March 2021).
- Pycraft, W.P. On the Morphology and Phylogeny of the Palæognathæ (Ratitæ and Crypturi) and Neognathæ (Carinatæ). Trans. Zool. Soc. Lond. 1900, 15, 149–290. [Google Scholar] [CrossRef]
- Bock, W. The cranial evidence for ratite affinities. In Proceedings of the XIII International Ornithological Congress, Ithaca, NY, USA, 17–24 June 1962; pp. 39–54. [Google Scholar]
- Parkes, K.C.; Clark, G.A., Jr. An Additional Character Linking Ratites and Tinamous, and an Interpretation of their Monophyly. Condor 1966, 68, 459–471. [Google Scholar] [CrossRef]
- Cracraft, J. Phylogeny and evolution of the ratite birds. Ibis 1974, 116, 494–521. [Google Scholar] [CrossRef]
- Mayr, G.; Zelenkov, N. Extinct crane-like birds (Eogruidae and Ergilornithidae) from the Cenozoic of Central Asia are indeed ostrich precursors. Ornithology 2021, 138, 1–15. [Google Scholar] [CrossRef]
- Huxley, T.H. On the Classification of Birds and on the Taxonomic Value of the Modifications of Certain of the Cranial Bones Observed in that Class. In Proceedings of the Zoological Society of London; Zoological Society of London: London, UK, 1867. [Google Scholar]
- Torres, C.R.; Norell, M.A.; Clarke, J.A. Bird neurocranial and body mass evolution across the end-Cretaceous mass extinction: The avian brain shape left other dinosaurs behind. Sci. Adv. 2021, 7, eabg7099. [Google Scholar] [CrossRef]
- Field, D.J.; Benito, J.; Kuo, P.; Jagt, J. Mesozoic Fossil Insight into the Palaeognath-Neognath Anatomical Dichotomy. In Proceedings of the The Society of Vertebrate Paleontology Annual Meeting, Vurtual Meeting, 1–5 November 2021; p. 278. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Kiwis (Apterygidae), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Bunce, M.; Worthy, T.H.; Phillips, M.J.; Holdaway, R.N.; Willerslev, E.; Haile, J.; Shapiro, B.; Scofield, R.P.; Drummond, A.; Kamp, P.J.J.; et al. The evolutionary history of the extinct ratite moa and New Zealand Neogene paleogeography. Proc. Natl. Acad. Sci. USA 2009, 106, 20646–20651. [Google Scholar] [CrossRef] [Green Version]
- Hansford, J.P.; Turvey, S.T. Unexpected diversity within the extinct elephant birds (Aves: Aepyornithidae) and a new identity for the world’s largest bird. R. Soc. Open Sci. 2018, 5, 181295. [Google Scholar] [CrossRef] [Green Version]
- Fromm, A.; Meiri, S. Big, flightless, insular and dead: Characterising the extinct birds of the Quaternary. J. Biogeogr. 2021, 1–10. [Google Scholar] [CrossRef]
- Winkler, D.; Billerman, S.; Lovette, I. Tinamous (Tinamidae). Birds World 2020. [Google Scholar] [CrossRef]
- Cabot, J.; Christie, D.A.; Jutglar, F.; Sharpe, C.J. Dwarf Tinamou (Taoniscus nanus), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Cabot, J.; Jutglar, F.; Garcia, E.F.J.; Boesman, P.F.D.; Sharpe, C.J. Gray Tinamou (Tinamus tao), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Folch, A.; Christie, D.A.; Jutglar, F.; Garcia, E.F.J. Common Ostrich (Struthio camelus), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Davies, S.J.J.F. Moas. In Grzimek’s Animal Life Encyclopedia. Birds I: Tinamous and Ratites to Hoatzins, 2nd ed.; Hutchins, M., Ed.; Gale Group: Farmington Hills, MI, USA, 2003; Volume 8, pp. 95–98. [Google Scholar]
- Szabo, M.J. South Island Giant Moa. Available online: http://nzbirdsonline.org.nz/species/south-island-giant-moa (accessed on 18 April 2021).
- McDowell, S. The Bony Palate of Birds. Part I The Palaeognathae. Auk 1948, 65, 520–549. [Google Scholar] [CrossRef]
- Fürbringer, M. Untersuchungen zur Morphologie und Systematik der Vögel: Zugleich ein Beitrag zur Anatomie der Stütz-und Bewegungsorgane; T. van Holkema: Amsterdam, The Netherlands, 1888; Volume 15. [Google Scholar]
- Allen, G.M. Birds and Their Attributes; Marshall Jones Company: Boston, MA, USA, 1925. [Google Scholar]
- Stresemann, E. Aves in: Kükenthal-Krumbach, Handbuch der Zoologie. Zool. VII 1927, 7, 2. [Google Scholar]
- Verheyen, R. Outline of procedure in basic avian systematics. Gerfaut 1960, 1960, 50. [Google Scholar]
- Cracraft, J. Continental drift, paleoclimatology, and the evolution and biogeography of birds. J. Zool. 1973, 169, 455–543. [Google Scholar] [CrossRef]
- Cracraft, J. Avian evolution, Gondwana biogeography and the Cretaceous-Tertiary mass extinction event. Proc. R. Soc. B 2001, 268, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Roff, D. The evolution of flightlessness: Is history important? Evol. Ecol. 1994, 8, 639–657. [Google Scholar] [CrossRef]
- Merrem, B. Tentamen systematis naturalis avium. Abh. Der Königlichen Akad. Der Wiss. Berl. 1813, 237–259. [Google Scholar]
- Lee, K.; Feinstein, J.; Cracraft, J. Chapter 7—The Phylogeny of Ratite Birds: Resolving Conflicts between Molecular and Morphological Data Sets. In Avian Molecular Evolution and Systematics; Mindell, D.P., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 173–209. [Google Scholar]
- Livezey, B.C.; Zusi, R.L. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. Zool. J. Linn. Soc. 2007, 149, 1–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourdon, E.; De Riqles, A.; Cubo, J. A new Transantarctic relationship: Morphological evidence for a Rheidae–Dromaiidae–Casuariidae clade (Aves, Palaeognathae, Ratitae). Zool. J. Linn. Soc. 2009, 156, 641–663. [Google Scholar] [CrossRef] [Green Version]
- Elzanowski, A. Cretaceous birds and avian phylogeny. Cour. Forsch. Senckenberg 1995, 181, 37–53. [Google Scholar]
- Johnston, P. New morphological evidence supports congruent phylogenies and Gondwana vicariance for palaeognathous birds. Zool. J. Linn. Soc. 2011, 163, 959–982. [Google Scholar] [CrossRef] [Green Version]
- Owen, R. The Anatomy of Vertebrates, Volume 2: Birds and Mammals; Longmans, Green and Company: London, UK, 1866. [Google Scholar]
- Chojnowski, J.L.; Kimball, R.T.; Braun, E.L. Introns outperform exons in analyses of basal avian phylogeny using clathrin heavy chain genes. Gene 2008, 410, 89–96. [Google Scholar] [CrossRef]
- Hackett, S.J.; Kimball, R.T.; Reddy, S.; Bowie, R.C.K.; Braun, E.L.; Braun, M.J.; Chojnowski, J.L.; Cox, W.A.; Han, K.; Harshman, J.; et al. A Phylogenomic Study of Birds Reveals Their Evolutionary History. Science 2008, 320, 1763–1768. [Google Scholar] [CrossRef]
- Harshman, J.; Braun, E.L.; Braun, M.J.; Huddleston, C.J.; Bowie, R.C.K.; Chojnowski, J.L.; Hackett, S.J.; Han, K.; Kimball, R.T.; Marks, B.D.; et al. Phylogenomic evidence for multiple losses of flight in ratite birds. Proc. Natl. Acad. Sci. USA 2008, 105, 13462–13467. [Google Scholar] [CrossRef] [Green Version]
- Haddrath, O.; Baker, A. Multiple nuclear genes and retroposons support vicariance and dispersal of the palaeognaths, and an Early Cretaceous origin of modern birds. Proc. R. Soc. B: Biol. Sci. 2012, 279, 4617–4625. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.V.; Braun, E.L.; Kimball, R.T. Ratite Nonmonophyly: Independent Evidence from 40 Novel Loci. Syst. Biol. 2012, 62, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.J.; Haddrath, O.; McPherson, J.D.; Cloutier, A. Genomic Support for a Moa–Tinamou Clade and Adaptive Morphological Convergence in Flightless Ratites. Mol. Biol. Evol. 2014, 31, 1686–1696. [Google Scholar] [CrossRef] [Green Version]
- Grealy, A.; Phillips, M.; Gifford, M.; Gilbert, M.T.P.R.; Jean-Marie Lambert, D.; Bunce, M.; Haile, J. Eggshell palaeogenomics: Palaeognath evolutionary history revealed through ancient nuclear and mitochondrial DNA from Madagascan elephant bird (Aepyornis sp.) eggshell. Mol. Phylogenetics Evol. 2017, 109, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Almeida, F.C.; Porzecanski, A.L.; Cracraft, J.L.; Bertelli, S. The evolution of tinamous (Palaeognathae: Tinamidae) in light of molecular and combined analyses. Zool. J. Linn. Soc. 2021. [Google Scholar] [CrossRef]
- Feng, S.; Stiller, J.; Deng, Y.; Armstrong, J.; Fang, Q.; Reeve, A.H.; Xie, D.; Chen, G.; Guo, C.; Faircloth, B.C.; et al. Dense sampling of bird diversity increases power of comparative genomics. Nature 2020, 587, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.J.; Gibb, G.C.; Crimp, E.A.; Penny, D. Tinamous and Moa Flock Together: Mitochondrial Genome Sequence Analysis Reveals Independent Losses of Flight among Ratites. Syst. Biol. 2009, 59, 90–107. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, K.J.; Llamas, B.; Soubrier, J.; Rawlence, N.J.; Worthy, T.H.; Wood, J.; Lee, M.S.Y.; Cooper, A. Ancient DNA reveals elephant birds and kiwi are sister taxa and clarifies ratite bird evolution. Science 2014, 344, 898–900. [Google Scholar] [CrossRef] [Green Version]
- Urantówka, A.D.; Kroczak, A.; Mackiewicz, P. New view on the organization and evolution of Palaeognathae mitogenomes poses the question on the ancestral gene rearrangement in Aves. BMC Genom. 2020, 21, 874. [Google Scholar] [CrossRef]
- Jarvis, E.D.; Mirarab, S.; Aberer, A.J.; Li, B.; Houde, P.; Li, C.; Ho, S.Y.W.; Faircloth, B.C.; Nabholz, B.; Howard, J.T.; et al. Whole-genome analyses resolve early branches in the tree of life of modern birds. Science 2014, 346, 1320–1331. [Google Scholar] [CrossRef] [Green Version]
- Prum, R.O.; Berv, J.S.; Dornburg, A.; Field, D.J.; Townsend, J.P.; Lemmon, E.M.; Lemmon, A.R. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature 2015, 526, 569–573. [Google Scholar] [CrossRef]
- Yonezawa, T.; Segawa, T.; Mori, H.; Campos, P.F.; Hongoh, Y.; Endo, H.; Akiyoshi, A.; Kohno, N.; Nishida, S.; Wu, J. Phylogenomics and morphology of extinct paleognaths reveal the origin and evolution of the ratites. Curr. Biol. 2017, 27, 68–77. [Google Scholar] [CrossRef]
- Sackton, T.B.; Grayson, P.; Cloutier, A.; Hu, Z.; Liu, J.S.; Wheeler, N.E.; Gardner, P.P.; Clarke, J.A.; Baker, A.J.; Clamp, M.; et al. Convergent regulatory evolution and loss of flight in paleognathous birds. Science 2019, 364, 74–78. [Google Scholar] [CrossRef] [Green Version]
- Norberg, U.L. Evolution of flight in animals. Flow Phenom. Nat. 2007, 1, 36–48. [Google Scholar]
- Livezey, B.C. Evolution of flightlessness in rails (Gruiformes, Rallidae): Phylogenetic, Ecomorphological, and Ontogenetic Perspectives. Ornithol. Monogr. 2003, 53, iii-654. [Google Scholar] [CrossRef]
- Gaspar, J.; Gibb, G.C.; Trewick, S.A. Convergent morphological responses to loss of flight in rails (Aves: Rallidae). Ecol. Evol. 2020, 10, 6186–6207. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, H.; Frankl-Vilches, C.; Bakker, A.; Mayr, G.; Nikolaus, G.; Boerno, S.T.; Klages, S.; Timmermann, B.; Gahr, M. An Unbiased Molecular Approach Using 3′-UTRs Resolves the Avian Family-Level Tree of Life. Mol. Biol. Evol. 2020, 38, 108–127. [Google Scholar] [CrossRef] [PubMed]
- Claramunt, S.; Cracraft, J. A new time tree reveals Earth history’s imprint on the evolution of modern birds. Sci. Adv. 2015, 1, e1501005. [Google Scholar] [CrossRef] [Green Version]
- Cloutier, A.; Sackton, T.B.; Grayson, P.; Clamp, M.; Baker, A.J.; Edwards, S.V. Whole-Genome Analyses Resolve the Phylogeny of Flightless Birds (Palaeognathae) in the Presence of an Empirical Anomaly Zone. Syst. Biol. 2019, 68, 937–955. [Google Scholar] [CrossRef]
- Kimball, R.; Oliveros, C.; Wang, N.; White, N.D.; Barker, F.K.; Field, D.J.; Ksepka, D.; Chesser, R.; Moyle, R.; Braun, M.; et al. A Phylogenomic Supertree of Birds. Diversity 2019, 11, 109. [Google Scholar] [CrossRef] [Green Version]
- Reddy, S.; Kimball, R.T.; Pandey, A.; Hosner, P.A.; Braun, M.J.; Hackett, S.J.; Han, K.; Harshman, J.; Huddleston, C.J.; Kingston, S.; et al. Why Do Phylogenomic Data Sets Yield Conflicting Trees? Data Type Influences the Avian Tree of Life more than Taxon Sampling. Syst. Biol. 2017, 66, 857–879. [Google Scholar] [CrossRef]
- Faux, C.; Field, D.J. Distinct developmental pathways underlie independent losses of flight in ratites. Biol. Lett. 2017, 13. [Google Scholar] [CrossRef]
- Farlie, P.G.; Davidson, N.M.; Baker, N.L.; Raabus, M.; Roeszler, K.N.; Hirst, C.; Major, A.; Mariette, M.M.; Lambert, D.M.; Oshlack, A.; et al. Co-option of the cardiac transcription factor Nkx2.5 during development of the emu wing. Nat. Commun. 2017, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Forshaw, J.M. Encyclopedia of Animals: Birds; Merehurst Limited: London, UK, 1991. [Google Scholar]
- Houde, P. Paleognathous Birds from the Early Tertiary of the Northern Hemisphere; Paynter, J.R.A., Ed.; Nuttall Ornithological Club: Cambridge, MA, USA, 1988; Volume 22. [Google Scholar]
- Parris, D.; Hope, S. New interpretations of birds from the Navesink and Hornerstown formations, New Jersey, USA (Aves: Neornithes). In Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, Beijing, China, 1–4 June 2002; pp. 113–124. [Google Scholar]
- Nesbitt, S.J.; Clarke, J.A. The anatomy and taxonomy of the exquisitely preserved Green River formation (early Eocene) Lithornithids (Aves) and the relationships of Lithornithidae. Bull. Am. Mus. Nat. Hist. 2016, 1–91. [Google Scholar] [CrossRef]
- Torres, C.R.; Norell, M.A.; Clarke, J.A. Estimating Flight Style of Early Eocene Stem Palaeognath Bird Calciavis grandei (Lithornithidae). Anat. Rec. 2020, 303, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Cassowaries and Emu (Casuariidae), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Ostriches (Struthionidae), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Rheas (Rheidae), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Clarke, J.A.; Tambussi, C.P.; Noriega, J.I.; Erickson, G.M.; Ketcham, R.A. Definitive fossil evidence for the extant avian radiation in the Cretaceous. Nature 2005, 433, 305–308. [Google Scholar] [CrossRef]
- Ksepka, D.T.; Stidham, T.A.; Williamson, T.E. Early Paleocene landbird supports rapid phylogenetic and morphological diversification of crown birds after the K-Pg mass extinction. Proc. Natl. Acad. Sci. USA 2017, 114, 8047–8052. [Google Scholar] [CrossRef] [Green Version]
- Field, D.J.; Bercovici, A.; Berv, J.S.; Dunn, R.; Fastovsky, D.E.; Lyson, T.R.; Vajda, V.; Gauthier, J.A. Early Evolution of Modern Birds Structured by Global Forest Collapse at the End-Cretaceous Mass Extinction. Curr. Biol. 2018, 28, 1825–1831.e2. [Google Scholar] [CrossRef] [Green Version]
- Field, D.J.; Benito, J.; Chen, A.; Jagt, J.; Ksepka, D. Late Cretaceous neornithine from Europe illuminates the origins of crown birds. Nature 2020, 579, 397–401. [Google Scholar] [CrossRef]
- Longrich, N.; Tokaryk, T.; Field, D.J. Mass extinction of birds at the Cretaceous Paleogene (K-Pg) boundary. Proc. Natl. Acad. Sci. 2011, 108, 15253–15257. [Google Scholar] [CrossRef] [Green Version]
- Saupe, E.E.; Farnsworth, A.; Lunt, D.J.; Sagoo, N.; Pham, K.V.; Field, D.J. Climatic shifts drove major contractions in avian latitudinal distributions throughout the Cenozoic. Proc. Natl. Acad. Sci. USA 2019, 116, 12895–12900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouch, N.M.A.; Clarke, J.A. Body size evolution in palaeognath birds is consistent with Neogene cooling-linked gigantism. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 532, 109224. [Google Scholar] [CrossRef]
- Mayr, G. Paleogene Fossil Birds; Springer: Berlin/Heidelberg, Germany, 2009; p. 262. [Google Scholar]
- Mayr, G. Avian Evolution – The Fossil Record of Birds and Its Paleobiological Significance; Wiley Blackwell: Chichester, UK, 2017; p. 289. [Google Scholar]
- Mikhailov, K.; Zelenkov, N. The late Cenozoic history of the ostriches (Aves: Struthionidae), as revealed by fossil eggshell and bone remains. Earth-Sci. Rev. 2020, 208, 103270. [Google Scholar] [CrossRef]
- Houde, P.; Olson, S. Paleognathous Carinate Birds from the Early Tertiary of North America. Science 1981, 214, 1236–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Toit, C.J.; Chinsamy, A.; Cunningham, S. Cretaceous origins of the vibrotactile bill-tip organ in birds. Proc. R. Soc. B 2020, 287, 20202322. [Google Scholar] [CrossRef]
- Clarke, J. Morphology, Phylogenetic Taxonomy, and Systematics of Ichthyornis and Apatornis (Avialae: Ornithurae). Bull. Am. Mus. Nat. Hist. 2004, 286, 1–179. [Google Scholar] [CrossRef]
- Benito, J.; Chen, A.; Wilson, L.E.; Bhullar, B.A.S.; Burnham, D.; Field, D.J. 40 new specimens of Ichthyornis provide unprecedented insight into the postcranial morphology of crownward stem group birds. bioRxiv 2022. [Google Scholar] [CrossRef]
- Scotese, C.R. PALEOMAP PaleoAtlas for GPlates and the PaleoData Plotter Program. Available online: http://www.earthbyte.org/paleomap-paleoatlas-for-gplates/ (accessed on 3 April 2021).
- Müller, R.D.; Cannon, J.; Qin, X.; Watson, R.J.; Gurnis, M.; Williams, S.; Pfaffelmoser, T.; Seton, M.; Russell, S.H.J.; Zahirovic, S. GPlates: Building a Virtual Earth Through Deep Time. Geochem. Geophys. Geosystems 2018, 19, 2243–2261. [Google Scholar] [CrossRef]
- Olson, S.L.; Parris, D.C. The Cretaceous birds of New Jersey. Smithson. Contrib. Paleobiol. 1987, 63, 1–22. [Google Scholar] [CrossRef]
- Staron, R.; Grandstaff, D.; Grandstaff, B.; Gallagher, W. Mosasaur taphonomy and geochemistry implications for a KT bone bed in the New Jersey coastal plain. J. Vertebr. Paleontol. 1999, 19, 78A. [Google Scholar]
- Lofgren, D.L.; Lillegraven, J.; Clemens, W.; Gingerich, P.; Williamson, T.; Woodburne, M.O. Paleocene Biochronology: The Puercan Through Clarkforkian Land Mammal Ages. In Late Cretaceous and Cenozoic Mammals of North America; Woodburne, M., Ed.; Columbia University Press: New York, NY, USA, 2004. [Google Scholar]
- Stidham, T.A.; Lofgren, D.; Farke, A.A.; Paik, M.; Choi, R. A lithornithid (Aves: Palaeognathae) from the Paleocene (Tiffanian) of southern California. PaleoBios 2014, 31. [Google Scholar] [CrossRef]
- Lofgren, D.L.; Honey, J.G.; McKenna, M.C.; Zondervan, R.L.; Smith, E.E.; Wang, X.; Barnes, L. Paleocene primates from the Goler Formation of the Mojave Desert in California. In Geology and Vertebrate Paleontology of Western and Southern North America, Contributions in Honor of David P. Whistler: National History Museum of Los Angeles County Science Series; National History Museum of Los Angeles County: Los Angeles, CA, USA, 2008; Volume 41, pp. 11–28. [Google Scholar]
- Albright, L.I.; Lofgren, D.; McKenna, M. Magnetostratigraphy, mammalian biostratigraphy, and refined age assessment of the Goler Formation (Paleocene), California. Mus. North Ariz. Bull. 2009, 65, 259–278. [Google Scholar]
- Lofgren, D.; Mckenna, M.; Honey, J.; Nydam, R.; Wheaton, C.; Yokote, B.; Henn, L.; Hanlon, W.; Manning, S.; Mcgee, C. New Records of Eutherian Mammals from the Goler Formation (Tiffanian, Paleocene) of California and Their Biostratigraphic and Paleobiogeographic Implications. Am. Mus. Novit. 2014, 2014, 1–57. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.E.; Carroll, A.R.; Singer, B.S. Synoptic reconstruction of a major ancient lake system; Eocene Green River Formation, western United States. Geol. Soc. Am. Bull. 2008, 120, 54–84. [Google Scholar] [CrossRef]
- Murphey, P.C.; Evanoff, E. Paleontology and stratigraphy of the middle Eocene Bridger Formation, southern Green River basin, Wyoming. In Proceedings of the 9th Conference on Fossil Resources, Kemmerer, WY, USA, 26–28 April 2011; pp. 83–109. [Google Scholar]
- Smith, T.; Smith, R. Terrestrial mammals as biostratigraphic indicators in upper Paleocene-lower Eocene marine deposits of the southern North Sea Basin. Geol. Soc. Am. Spec. Pap. 2003, 369, 513–520. [Google Scholar] [CrossRef]
- De Bast, E.; Steurbaut, E.; Smith, T. New mammals from the marine Selandian of Maret, Belgium, and their implications for the age of the Paleocene continental deposits of Walbeck, Germany. Geol. Belg. 2013, 16, 4. [Google Scholar]
- Mayr, G.; Smith, T. New Paleocene bird fossils from the North Sea Basin in Belgium and France. Geol. Belg. 2019, 22, 35–46. [Google Scholar] [CrossRef]
- Aguilar, J.; Augustı, J.; Alexeeva, N.; Antoine, P.; Antunes, M.; Archer, M. Syntheses and correlation tables. In Proceedings of the Actes du Congres BiochroM, Montpellier, France, 14–17 April 1997; pp. 769–805. [Google Scholar]
- Mayr, G. The Birds from the Paleocene Fissure Filling of Walbeck (Germany). J. Vertebr. Paleontol. 2007, 27, 394–408. [Google Scholar] [CrossRef]
- Steurbaut, E. High-resolution holostratigraphy of Middle Paleocene to Early Eocene strata in Belgium and adjacent areas. Palaeontogr. Abt. A: Palaozool. Stratigr. 1997, 247, 91–156. [Google Scholar]
- Moreau, F.; Mathis, S. Les élasmobranches du Thanétien (Paléocène) du Nord de la France, des carrières de Templeuve et de Leforest. Cossmanniana 2000, 7, 1–18. [Google Scholar]
- Heilmann-Clausen, C.; Schmitz, B. The late Paleocene thermal maximum δ13C excursion in Denmark? Gff 2000, 122, 70. [Google Scholar] [CrossRef]
- Bourdon, E.; Lindow, B.E.K. A redescription of Lithornis vulturinus (Aves, Palaeognathae) from the Early Eocene Fur Formation of Denmark. Zootaxa 2015, 4032, 493–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, L.; Pringle, M.; Fitton, G.; Larsen, L.; Pedersen, A.; Parrish, R. Recalibration of the Palaeocene-Eocene boundary (PE) using high precision U-Pb and Ar-Ar isotopic dating. In Proceedings of the EGS-AGU-EUG Joint Assembly, Nice, France, 6–11 April 2003; p. 9681. [Google Scholar]
- Leonard, L.; Dyke, G.; van Tuinen, M. A New Specimen of the Fossil Palaeognath Lithornis from the Lower Eocene of Denmark. Am. Mus. Novit. 2005, 3491, 1–11. [Google Scholar] [CrossRef]
- King, C. The stratigraphy of the London Clay and associated deposits. Tert. Res. Spec. Pap. 1981, 6, 1–158. [Google Scholar]
- Ellison, R.A.; Woods, M.A.; Allen, D.J.; Forster, A.; Pharaoh, T.C.; King, C. Palaeogene–Eocene. In Geology of London; British Geological Survey: London, UK, 2004; pp. 44–54. [Google Scholar]
- Friedman, M.; Beckett, H.T.; Close, R.A.; Johanson, Z. The English Chalk and London Clay: Two remarkable British bony fish Lagerstätten. Geol. Soc. Lond. Spec. Publ. 2016, 430, 165–200. [Google Scholar] [CrossRef]
- Franzen, J.; Haubold, H. The middle Eocene of European mammalian stratigraphy. Definition of the Geiseltalian. Mod. Geol. 1986, 10, 159–170. [Google Scholar]
- Schaal, S.; Ziegler, W. Messel: Ein Schaufenster in die Geschichte der Erde und des Lebens; Waldemar Kramer: Frankfurt, Germany, 1988. [Google Scholar]
- Lenz, O.K.; Wilde, V.; Mertz, D.F.; Riegel, W. New palynology-based astronomical and revised 40Ar/39Ar ages for the Eocene maar lake of Messel (Germany). Int. J. Earth Sci. 2015, 104, 873–889. [Google Scholar] [CrossRef]
- Mayr, G. First substantial Middle Eocene record of the Lithornithidae (Aves): A postcranial skeleton from Messel (Germany). Ann. De Paleontol. 2008, 94, 29–37. [Google Scholar] [CrossRef]
- Mayr, G. Towards the complete bird—The skull of the middle Eocene Messel lithornithid (Aves, Lithornithidae). Bull. De L’Institute R. Des. Sci. Nat. De Belg. 2009, 79, 167–173. [Google Scholar]
- Blondel, J.; Mourer-Chauviré, C. Evolution and history of the western Palaearctic avifauna. Trends Ecol. Evol. 1998, 13, 488–492. [Google Scholar] [CrossRef]
- Mayr, G. Birds–The most species-rich vertebrate group in Messel. Senckenberg Ges. Für Nat. Frankf. Am Main 2018, 169–214. [Google Scholar]
- Harrison, C. A new cathartid vulture from the lower Eocene of Wyoming. Tert. Res. 1979, 5, 7–10. [Google Scholar]
- Grande, L. Paleontology of the Green River Formation, with a Review of the Fish Fauna, 2nd ed.; University of Wyoming: Laramie, WY, USA, 1984. [Google Scholar]
- Wetmore, A. Fossil birds from the Green River deposits of eastern Utah. Ann. Carnegie Mus. 1926, 16, 391–497. [Google Scholar]
- Brodkorb, P. An eocene puffbird from Wyoming. Rocky Mt. Geol. 1970, 9, 13–15. [Google Scholar]
- Olson, S. A Lower Eocene Frigatebird from the Green River Formation of Wyoming (Pelecaniformes: Fregatidae). Smithson. Contrib. Paleobiol. 1977, 35. [Google Scholar] [CrossRef]
- Olson, S. An early Eocene oilbird from the Green River Formation of Wyoming (Caprimulgiformes: Steatornithidae). In Proceedings of the Table Ronde internationale du CNRS, Lyon, France, 18–21 September 1985; pp. 57–69. [Google Scholar]
- Olson, S. A new family of primitive landbirds from the early Eocene Green River Formation of Wyoming. Pap. Avian Paleontol. Honor. Pierce Brodkorb 1992, 36, 127–136. [Google Scholar]
- Mayr, G.; Daniels, M. A new short-legged landbird from the Early Eocene of Wyoming and contemporaneous European sites. Acta Palaeontol. Pol. 2001, 46, 393–402. [Google Scholar]
- Olson, S.; Matsuoka, H. New specimens of the early Eocene frigatebird Limnofregata (Pelecaniformes: Fregatidae), with the description of a new species. Zootaxa 2005, 1046, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ksepka, D.T.; Clarke, J.A. Affinities of Palaeospiza bella and the Phylogeny and Biogeography of Mousebirds (Coliiformes). Auk 2009, 126, 245–259. [Google Scholar] [CrossRef]
- Ksepka, D.T.; Clarke, J.A. Primobucco mcgrewi (Aves: Coracii) from the Eocene Green River Formation: New anatomical data from the earliest constrained record of stem rollers. J. Vertebr. Paleontol. 2010, 30, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Weidig, I. New birds from the Lower Eocene Green River Formation, North America. In Proceedings of the VII International Meeting of the Society of Avian Paleontology and Evolution, ed. W.E. Boles and T.H. Worthy. Rec. Aust. Mus. 2010, 62, 29–44. [Google Scholar]
- Ksepka, D.T.; Clarke, J.A.; Grande, L. Stem Parrots (Aves, Halcyornithidae) from the Green River Formation and a Combined Phylogeny of Pan-Psittaciformes. J. Paleontol. 2011, 85, 835–852. [Google Scholar] [CrossRef]
- Smith, N.D.; Grande, L.; Clarke, J.A. A new species of Threskiornithidae-like bird (Aves, Ciconiiformes) from the Green River Formation (Eocene) of Wyoming. J. Vertebr. Paleontol. 2013, 33, 363–381. [Google Scholar] [CrossRef]
- Owen, R. Description of the remains of a mammal, a bird, and a serpent from the London Clay. Proc. Geol. Soc. Lond. 1840, 3, 162–166. [Google Scholar]
- Owen, R. Description of the Fossil Remains of a Mammal (Hyracotherium leporinum) and of a Bird (Lithornis vulturinus) from the London Clay. Trans. Geol. Soc. 1841, 2, 203–208. [Google Scholar] [CrossRef]
- Harrison, C.J.O.; Walker, C.A. Birds of the British Lower Eocene. Tert. Res. Spec. Pap. 1977, 3, 1–52. [Google Scholar]
- Harrison, C.J.O. Rail-like cursorial birds of the British Lower Eocene, with descriptions of two new species. Lond. Nat. 1984, 63, 14–23. [Google Scholar]
- Bledsoe, A.H. A Phylogenetic Analysis of Postcranial Skeletal Characters of the Ratite Birds. Ann. Carnegie Mus. 1988, 57, 73–90. [Google Scholar]
- Mayr, G.; Clarke, J. The deep divergences of neornithine birds: A phylogenetic analysis of morphological characters. Cladistics 2003, 19, 527–553. [Google Scholar] [CrossRef]
- Clarke, J.; Zhou, Z.; Zhang, F. Insight into the evolution of avian flight from a new clade of Early Cretaceous ornithurines from China and the morphology of Yixianornis grabaui. J. Anat. 2006, 208, 287–308. [Google Scholar] [CrossRef]
- Worthy, T.H.; Mitri, M.; Handley, W.D.; Lee, M.S.Y.; Anderson, A.; Sand, C. Osteology supports a stem-galliform affinity for the giant extinct flightless bird Sylviornis neocaledoniae (Sylviornithidae, Galloanseres). PLoS ONE 2016, 11, e0150871. [Google Scholar] [CrossRef] [Green Version]
- Worthy, T.H.; Degrange, F.J.; Handley, W.D.; Lee, M.S.Y. The evolution of giant flightless birds and novel phylogenetic relationships for extinct fowl (Aves, Galloanseres). R. Soc. Open Sci. 2017, 4, 170975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Hoyo, J.; Collar, N.; Garcia, E.F.J. Somali Ostrich (Struthio molybdophanes), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Janz, L.; Feathers, J.K.; Burr, G.S. Dating surface assemblages using pottery and eggshell: Assessing radiocarbon and luminescence techniques in Northeast Asia. J. Archaeol. Sci. 2015, 57, 119–129. [Google Scholar] [CrossRef]
- Khatsenovich, A.; Rybin, E.; Gunchinsuren, B.; Bolorbat, T.; Odsuren, D.; Angaragdulguun, G.; Margad-Erdene, G. Human and Struthio asiaticus: One page of Paleolithic art in the eastern part of Central Asia. Irkutsk State Univ. Bull. Ser. Geoarcheol. Ethnol. Anthropol. 2017, 21, 80–106. [Google Scholar]
- Wang, S.; Hu, Y.; Wang, L. New ratite eggshell material from the Miocene of Inner Mongolia, China. Chin. Birds 2011, 2, 18–26. [Google Scholar] [CrossRef]
- Mayr, G. The middle Eocene European “ratite” Palaeotis (Aves, Palaeognathae) restudied once more. Paläontologische Z. 2015, 89, 503–514. [Google Scholar] [CrossRef]
- Houde, P.; Haubold, H. Palaeotis weigelti restudied; a small middle Eocene ostrich (Aves: Struthioniformes). Palaeovertebrata 1987, 17, 27–42. [Google Scholar]
- Lemoine, V. Recherches sur les Oiseaux Fossiles des Terrains Tertiaires Inférieurs des Environs de Reims, Deuxième Partie; Matot-Braine: Reims, France, 1881. [Google Scholar]
- Lambrecht, K. Palaeotis weigelti n. g. n. sp., eine fossile Trappe aus der mitteleozänen Braunkohle des Geiseltales. Jahrb. Des Halleschen Verb. Für Die Erforsch. Mitteldtsch. Bodenschätze 1928, 7, 1–11. [Google Scholar]
- Peters, D.S. Ein vollständiges Exemplar von Palaeotis weigelti (Aves, Palaeognathae). Cour. Forsch. Senckenberg 1988, 107, 223–233. [Google Scholar]
- Dyke, G.J. The fossil record and molecular clocks: Basal radiations within Neornithes. In Telling the Evolutionary Time—Molecular Clocks and the Fossil Record; Smith, P., Donoghue, P., Eds.; Taylor and Francis: London, UK, 2003; pp. 263–278. [Google Scholar]
- Mayr, G. Hindlimb morphology of Palaeotis suggests palaeognathous affinities of the Geranoididae and other crane-like birds from the Eocene of the Northern Hemisphere. Acta Palaeontol. Pol. 2019, 64, 669–678. [Google Scholar] [CrossRef]
- Mayr, G. On the taxonomy and osteology of the Early Eocene North American Geranoididae (Aves, Gruoidea). Swiss J. Palaeontol. 2016, 135, 315–325. [Google Scholar] [CrossRef]
- Bourdon, E.; Mourer-Chauviré, C.; Laurent, Y. Early Eocene Birds from La Borie, Southern France. Acta Palaeontol. Pol. 2016, 61, 175–190. [Google Scholar] [CrossRef] [Green Version]
- Cracraft, J. Systematics and evolution of the Gruiformes (Class, Aves). 1. The Eocene family Geranoididae and the early history of the Gruiformes. Am. Mus. Novit. 1969, 2388, 1–41. [Google Scholar]
- Kurochkin, E. A survey of the Paleogene birds of Asia. Smithson. Contrib. Paleobiol. 1976, 27, 75–86. [Google Scholar]
- Burchak-Abramovich, N.I. Urmiornis (Urmiornis maraghanus Mecq.) strausopodobnaya ptitsa gipparionovoi fauny Zakavkaz’a i yuzhnoi Ukrainy [Urmiornis (Urmiornis maraghanus Mecq.), an ostrich-like bird from the Hipparion fauna of Transcaucasia and southern Ukraine]. Izv. Akedemii Nauk Azerbaidzhanskoi SSR 1951, 6, 83–94. [Google Scholar]
- Feduccia, A. The Age of Birds; Harvard University Press: Cambridge, MA, USA, 1980. [Google Scholar]
- Olson, S.L. The fossil record of birds. Avian Biol. 1985, 79–238. [Google Scholar]
- Clarke, J.A.; Norell, M.A.; Dashzėvėg, D. New Avian Remains from the Eocene of Mongolia and the Phylogenetic Position of the Eogruidae (Aves, Gruoidea). Am. Mus. Novit. 2005, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Elzanowski, A.; Paul, G.S.; Stidham, T.A. An avian quadrate from the Late Cretaceous Lance Formation of Wyoming. J. Vertebr. Paleontol. 2001, 20, 712–719. [Google Scholar] [CrossRef]
- Vandenberghe, N.; Hilgen, F.J.; Speijer, R.P.; Ogg, J.G.; Gradstein, F.M.; Hammer, O.; Hollis, C.J.; Hooker, J.J. Chapter 28—The Paleogene Period. In The Geologic Time Scale; Gradstein, F.M., Ogg, J.G., Schmitz, M., Ogg, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 855–921. [Google Scholar]
- Lambrecht, K. Drei neue Vogelformen aus dem Lutétian des Geiseltales. Nova Acta Leopold. 1935, 3, 361–367. [Google Scholar]
- Buffetaut, E.; Angst, D. Stratigraphic distribution of large flightless birds in the Palaeogene of Europe and its palaeobiological and palaeogeographical implications. Earth-Sci. Rev. 2014, 138, 394–408. [Google Scholar] [CrossRef]
- Martin, L.D. The status of the Late Paleocene birds Gastornis and Remiornis. Nat. Hist. Mus. Los Angeles Cty. Sci. Ser. 1992, 36, 97–108. [Google Scholar]
- Smith, T.; Quesnel, F.; De Plöeg, G.; De Franceschi, D.; Métais, G.; De Bast, E.; Solé, F.; Folie, A.; Boura, A.; Claude, J.; et al. First Clarkforkian equivalent Land Mammal Age in the latest Paleocene basal Sparnacian facies of Europe: Fauna, flora, paleoenvironment and (bio)stratigraphy. PLoS ONE 2014, 9, e86229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buffetaut, E.; de Ploëg, G. Giant Birds from the Uppermost Paleocene of Rivecourt (Oise, Northern France). Bol. Do Cent. Port. De Geo-História E Pré-História 2020, 2, 1. [Google Scholar]
- Laurent, Y.; Adnet, S.; Bourdon, E.; Corbalan, D.; Danilo, L.; Duffaud, S.; Fleury, G.; Garcia, G.; Godinot, M.; Le Roux, G. La Borie (Saint-Papoul, Aude): Un gisement exceptionnel dans l’Éocène basal du Sud de la France. Bull. De La Société D’histoire Nat. De Toulouse 2010, 146, 89–103. [Google Scholar]
- Danilo, L.; Remy, J.A.; Vianey-Liaud, M.; Marandat, B.; Sudre, J.; Lihoreau, F. A new Eocene locality in southern France sheds light on the basal radiation of Palaeotheriidae (Mammalia, Perissodactyla, Equoidea). J. Vertebr. Paleontol. 2013, 33, 195–215. [Google Scholar] [CrossRef]
- Zelenkov, N.; Kurochkin, E.N. Class. Aves, Iskopaemye Pozvonochnye Rossii i Sopredel’nyh Stran. Iskopaemye Reptilii i Ptitsy. Chast’3; Kurochkin, E., Lopatin, A., Zelenkov, N., Eds.; GEOS: Moscow, Russia, 2015. [Google Scholar]
- Zelenkov, N.; Boev, Z.; Lazaridis, G. A large ergilornithine (Aves, Gruiformes) from the Late Miocene of the Balkan Peninsula. Paläontologische Z. 2016, 90, 145–151. [Google Scholar] [CrossRef]
- Lydekker, R. Catalogue of the Fossil Birds in the British Museum (Natural History); Order of the Trustees: London, UK, 1891. [Google Scholar]
- Tsoukala, E.; Bartsiokas, A. New Mesopithecus pentelicus specimens from Kryopigi, Macedonia, Greece. J. Hum. Evol. 2008, 54, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Lazaridis, G.; Tsoukala, E. Hipparion phlegrae, sp. nov. (Mammalia, Perissodactyla): A new species from the Turolian locality of Kryopigi (Kassandra, Chalkidiki, Greece). J. Vertebr. Paleontol. 2014, 34, 164–178. [Google Scholar] [CrossRef]
- Boev, Z.; Lazaridis, G.; Tsoukala, E. Otis hellenica sp. nov., a new Turolian bustard (Aves: Otididae) from Kryopigi (Chalkidiki, Greece). Geol. Balc. 2013, 42, 59–64. [Google Scholar] [CrossRef]
- Li, Q. Eocene fossil rodent assemblages from the Erlian Basin (Inner Mongolia, China): Biochronological implications. Palaeoworld 2016, 25, 95–103. [Google Scholar] [CrossRef]
- Wetmore, A. Fossil Birds from Mongolia and China. Am. Mus. Novit. 1934, 711, 1–16. [Google Scholar]
- Bendukidze, O. Novyj prestavitel’ semeist-va Geranoididae (Aves, Gruiformes) iz eotsenovykh otlozhenij Zaisan. Soobtzhenija Akad. Nauk Gruz. SSSR 1971, 63, 749–751. [Google Scholar]
- Russell, D.; Zhai, R. The Paleogene of Asia: Mammals and stratigraphy. Mémoires Du Muséum Natl. D’histoire Nat. Série C 1987, 52, 1–488. [Google Scholar]
- Kurochkin, E.N. New representatives and evolution of two archaic gruiform families in Eurasia. Tr. Sovmest. Sov. Mong. Paleontol. Ekspeditsija 1981, 15, 59–85. [Google Scholar]
- Dashzėvėg, D. Some carnivorous mammals from the Paleogene of the eastern Gobi Desert, Mongolia, and the application of Oligocene carnivores to stratigraphic correlation. Am. Mus. Novit. 1996, 3179, 1–14. [Google Scholar]
- Musser, G.; Li, Z.; Clarke, J.A. A new species of Eogruidae (Aves: Gruiformes) from the Miocene of the Linxia Basin, Gansu, China: Evolutionary and climatic implications. Auk 2019, 137. [Google Scholar] [CrossRef]
- Kozlova, E.V. Novye iskopaemye ptitsy iz yugovostochnoi Gobi. Tr. Probl. I Temat. Soveshanii ZIN 1960, 9, 323–329. [Google Scholar]
- Karhu, A. A new species of Urmiornis (Gruiformes: Ergilornithidae) from the early Miocene of western Kazakhstan. Paleontol. J. 1997, 31, 102–107. [Google Scholar]
- Wang, X.; Qiu, Z.; Opdyke, N.D. Litho-, bio-, and magnetostratigraphy and paleoenvironment of Tunggur Formation (Middle Miocene) in central Inner Mongolia, China. Am. Mus. Novit. 2003, 2003, 1–31. [Google Scholar] [CrossRef]
- Brodkorb, P. Catalogue of fossil birds. Part 3 (Ralliformes, Ichthyornithiformes, Charadriiformes). Bull. Fla. State Mus. Biol. Sci. II 1967, 3, 1–220. [Google Scholar]
- Cracraft, J. Systematics and evolution of the Gruiformes (class Aves). 3. Phylogeny of the suborder Grues. Bull. Am. Mus. Nat. Hist. 1973, 151, 1–128. [Google Scholar]
- Barry, J.C.; Morgan, M.E.; Flynn, L.J.; Pilbeam, D.; Behrensmeyer, A.K.; Raza, S.M.; Khan, I.A.; Badgley, C.; Hicks, J.; Kelley, J. Faunal and environmental change in the late Miocene Siwaliks of northern Pakistan. Paleobiology 2002, 28, 1–71. [Google Scholar] [CrossRef]
- Harrison, C.; Walker, C.A. Fossil birds from the Upper Miocene of northern Pakistan. Tert. Res. 1982, 4, 53–69. [Google Scholar]
- Mecquenem, R. Contribution à l’étude du gisement des vertébrés de Maragha et de ses environs. Ann. D’histoire Nat. 1908, 1, 27–29. [Google Scholar]
- Fang, X.; Wang, J.; Zhang, W.; Zan, J.; Song, C.; Yan, M.; Appel, E.; Zhang, T.; Wu, F.; Yang, Y.; et al. Tectonosedimentary evolution model of an intracontinental flexural (foreland) basin for paleoclimatic research. Glob. Planet. Chang. 2016, 145, 78–97. [Google Scholar] [CrossRef]
- Kurochkin, E.N. Birds of the central Asia in Pliocene. Trans. Jt. Sov. Mong. Paleontol. Exped. 1985, 26, 1–120. [Google Scholar]
- Mecquenem, R. Oiseaux. Contribution à l’étude des fossiles de Maragha. Ann. De Paléontologie 1925, 14, 54–56. [Google Scholar]
- Portis, A. Contribuzioni alla ornitolitologia italiana. Mem. Della R. Accad. Delle Sci. Di Torino 1884, 36, 361–384. [Google Scholar]
- Schaub, S. Ein Ratitenbecken aus dem Bohnerz von Egerkingen. Eclogae Geol. Helv. 1940, 33, 274–284. [Google Scholar]
- Gaillard, C. Un oiseau géant dans les dépôts éocènes du Mont-d’Or lyonnais. Compte Rendus De L’académie Des Sci. 1936, 202, 965–967. [Google Scholar]
- Angst, D.; Buffetaut, E.; Lécuyer, C.; Amiot, R. “Terror Birds” (Phorusrhacidae) from the Eocene of Europe imply trans-Tethys dispersal. PLoS ONE 2013, 8, e80357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, C.W. On the Pelvis and Hind-limb of Mullerornis betsilei M. Edw. & Grand.; with a Note On the Occurrence of a Ratite Bird in the Upper Eocene Beds of the Fayurn, Egypt. Proc. Zool. Soc. Lond. 1904, 74, 163–171. [Google Scholar]
- Rasmussen, D.T.; Simons, E.L.; Hertel, F.; Judd, A. Hindlimb of a giant terrestrial bird from the upper Eocene, Fayum, Egypt. Palaeontology 2001, 44, 325–337. [Google Scholar] [CrossRef]
- Pickford, M.; Senut, B. Miocene Palaeobiology of the Orange River Valley, Namibia. Mem. Geol. Surv. Namib. 2003, 19, 1–22. [Google Scholar]
- Mourer-Chauviré, C.; Senut, B.; Pickford, M.; Mein, P. The oldest representative of the genus Struthio (Aves: Struthionidae), Struthio coppensi n. sp., from the Lower Miocene of Namibia. Comptes Rendus De L’académie Des Sci. Sér. 2 Fasc. A Sci. De La Terre Des Planètes 1996, 322, 325–332. [Google Scholar]
- Mourer-Chauviré, C. Birds (Aves) from the Early Miocene of the Northern Sperrgebiet, Namibia. Mem. Geol. Surv. Namib. 2009, 20, 147–167. [Google Scholar]
- Pickford, M. Preliminary Miocene mammalian biostratigraphy for western Kenya. J. Hum. Evol. 1981, 10, 73–97. [Google Scholar] [CrossRef]
- Leonard, L.M.; Dyke, G.J.; Walker, C.A. New specimens of a fossil ostrich from the Miocene of Kenya. J. Afr. Earth Sci. 2006, 45, 391–394. [Google Scholar] [CrossRef]
- Werdelin, L. Chronology of Neogene mammal localities. In Cenozoic Mammals of Africa; Werdelin, L., Sanders, W.J., Eds.; University of California Press: Berkeley, CA, USA, 2010; pp. 27–43. [Google Scholar]
- Rich, P.V. A fossil avifauna from the Upper Miocene Beglia Formation of Tunisia. Notes De Serv. Géologique Du Tunis 1972, 35, 29–66. [Google Scholar]
- Roberts, D.L.; Matthews, T.; Herries, A.I.; Boulter, C.; Scott, L.; Dondo, C.; Mtembi, P.; Browning, C.; Smith, R.M.; Haarhoff, P. Regional and global context of the Late Cenozoic Langebaanweg (LBW) palaeontological site: West Coast of South Africa. Earth-Sci. Rev. 2011, 106, 191–214. [Google Scholar] [CrossRef]
- Rich, P. Preliminary report on the fossil avian remains from late Tertiary sediments at Langebaanweg (Cape Province), South Africa. South. Afr. J. Sci. 1980, 76, 166–170. [Google Scholar]
- Manegold, A.; Louchart, A.; Carrier, J.; Elzanowski, A. The early Pliocene avifauna of Langebaanweg (South Africa): A review and update. In Proceedings of the 8th International Meeting of the Society of Avian Paleontology and Evolution, Vienna, Austria, 11–16 June 2012; pp. 135–152. [Google Scholar]
- Geraads, D. Carnivores du Pliocène terminal de Ahl al Oughlam (Casablanca, Maroc). Geobios 1997, 30, 127–164. [Google Scholar] [CrossRef]
- Mourer-Chauviré, C.; Geraads, D. The Struthionidae and Pelagornithidae (Aves: Struthioniformes, Odontopterygiformes) from the late Pliocene of Ahl al Oughlam, Morocco [Les Struthionidae et les Pelagornithidae (Aves, Struthioniformes et Odontopterygiformes) du Pliocène final d’Ahl al Oughlam, Maroc]. In Proceedings of the 6ème Symposium international de la Society for Avian Paleontology and Evolution (SAPE), Quillan, France, 28 September–3 October 2004; pp. 169–194. [Google Scholar]
- Hay, R.L. Geology of the Olduvai Gorge; University of California Press: Berkeley, CA, USA, 1976. [Google Scholar]
- Lowe, P.R. XLI.—On Some Struthious Remains:—1. Description of some Pelvic Remains of a large Fossil Ostrich, Struthio oldawayi, sp. n., from the Lower Pleistocene of Oldaway (Tanganyika Territory); 2. Egg-shell Fragments referable to Psammornis and other Struthiones collected by Mr. St. John Philby in Southern Arabia. Ibis 1933, 75, 652–658. [Google Scholar] [CrossRef]
- Leakey, L.S.B. Olduvai Gorge. 1951–1961. Vol 1. A Preliminary Report on the Geology and Fauna; Cambridge University Press: Cambridge, UK, 1967. [Google Scholar]
- Arambourg, C. Vertébrés villafranchiens d’Afrique du Nord. (Artiodactyles, Carnivores, Primates, Reptiles, Oiseaux); Fondation Singer-Polignac: Paris, France, 1979; p. 141. [Google Scholar]
- Becker-Platen, J.D.; Benda, L.; Steffens, P. Litho-und Biostratigraphische Deutung radiometrischer Altersbestimmungen aus dem Jungtertiär der Türkei; Schweizerbart’sche Verlagsbuchhandlung: Sttutgart, Germany, 1977. [Google Scholar]
- Sauer, E.G. A Miocene Ostrich from Anatolia. Ibis 1979, 121, 494–501. [Google Scholar] [CrossRef]
- Lambrecht, K. Handbuch der Palaeornithologie; Gebrüder Borntraeger: Berlin, Germany, 1933. [Google Scholar]
- Louchart, A.; Bibi, F.; Stewart, J.R. The birds of the late Miocene Baynunah Formation, Abu Dhabi Emirate. In Sands of Time: Late Miocene Fossils from the Baynunah Formation, U.A.E.; Bibi, F., Kraatz, B., Beech, M., Hill, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Tugarinov, A.Y. Some data on Pliocene ornithofauna of Siberia. Tr. Paleozoologichaskogo Inst. 1935, 4, 79–85. [Google Scholar]
- Deng, T.; Qiu, Z.; Wang, B.; Wang, X.; Hou, S. Late Cenozoic biostratigraphy of the Linxia basin, northwestern China. In Fossil Mammals of Asia; Columbia University Press: New York, NY, USA, 2013; pp. 243–273. [Google Scholar]
- Hou, L.; Zhou, Z.; Zhang, F.; Wang, Z. A Miocene ostrich fossil from Gansu Province, northwest China. Chin. Sci. Bull. 2005, 50, 1808–1810. [Google Scholar] [CrossRef]
- Kaakinen, A.; Passey, B.H.; Zhang, Z.; Liu, L.; Pesonen, L.J.; Fortelius, M. Stratigraphy and paleoecology of the classical dragon bone localities of Baode County, Shanxi Province. In Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology; Wang, X., Flynn, L.J., Fortelius, M., Eds.; Columbia University Press: New York, NY, USA, 2013; pp. 203–217. [Google Scholar]
- Lowe, P.R. Struthious Remains from Northern China and Mongolia: With Descriptions of Struthio wimani, Struthio anderssoni and Struthio mongolicus, Spp. Nov.; Geological Survey of China: Beijing, China, 1931. [Google Scholar]
- Sahni, A.; Kumar, G.; Bajpai, S.; Srinivasan, S. Ultrastructure and taxonomy of ostrich eggshells from Upper Palaeolithic sites of India. J. Palaeontol. Soc. India 1989, 34, 91–98. [Google Scholar]
- Sahni, A.; Kumar, G.; Srinivasan, S.; Bajpai, S. Review of Late Pleistocene ostriches (Struthio sp.) of India. Man Environ. 1990, 15, 41–47. [Google Scholar]
- Stern, L.A.; Johnson, G.D.; Chamberlain, C.P. Carbon isotope signature of environmental change found in fossil ratite eggshells from a South Asian Neogene sequence. Geology 1994, 22, 419–422. [Google Scholar] [CrossRef]
- Patnaik, R.; Sahni, A.; Cameron, D.; Pillans, B.; Chatrath, P.; Simons, E.; Williams, M.; Bibi, F. Ostrich-like eggshells from a 10.1 million-yr-old Miocene ape locality, Haritalyangar, Himachal Pradesh, India. Curr. Sci. 2009, 96, 1485–1495. [Google Scholar]
- Davies, W. III.—On some Fossil Bird-Remains from the Siwalik Hills in the British Museum. Geol. Mag. 1880, 7, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Lydekker, R. Indian Tertiary and post-Tertiary Vertebrata. Siwalik and Narbada bunodont Suina. Mem. Geol. Surv. India Palaeontol. Indica Ser. 10 1884, 3, 35–104. [Google Scholar]
- Ginsburg, I.; Heintz, F.; Sen, S. Le gisement pliocène à Mammiferes de Çalta (Ankara, Turquie). Comptes Rendus De L’académie Des Sci. Paris 1974, 278, 2739–2742. [Google Scholar]
- Sen, S. La faune de rangeurs pliocènes de Çalta (Ankara, Turqïe). Bull. Du Muséum Mational D’histoire Nat. Ser. 3 Sci. De La Terre 1977, 61, 89–171. [Google Scholar]
- Janoo, A.; Sen, S. Pliocene vertebrate locality of Çalta, Ankara, Turkey. 2. Aves: Struthionidae. Geodiversitas 1998, 20, 339–351. [Google Scholar]
- Beliaeva, E.I. Catalogue of Tertiary Fossil Sites of Land Mammals in the USSR; American Geological Institute: Alexandria, VA, USA, 1962. [Google Scholar]
- Sotnikova, M.; Dodonov, A.; Pen’kov, A. Upper Cenozoic bio-magnetic stratigraphy of Central Asian mammalian localities. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 133, 243–258. [Google Scholar] [CrossRef]
- Cai, B.; Zheng, S.; Liddicoat, J.C.; Li, Q. Review of the litho-, bio-, and chronostratigraphy in the Nihewan Basin, Hebei, China. In Fossil Mammals of Asia; Wang, X., Flynn, L.J., Fortelius, M., Eds.; Columbia University Press: New York, NY, USA, 2013; pp. 218–242. [Google Scholar]
- Buffetaut, E.; Angst, D. A Giant Ostrich from the Lower Pleistocene Nihewan Formation of North China, with a Review of the Fossil Ostriches of China. Diversity 2021, 13, 47. [Google Scholar] [CrossRef]
- Hou, L. Avian fossils of Pleistocene from Zhoukoudian. Mem. Inst. Vertebr. Palaeontol. Palaeoanthropology Acad. Sin. 1993, 19, 165–297. [Google Scholar]
- Vangengeim, E.; Tesakov, A.S. Late Miocene Mammal Localities of Eastern Europe and Western Asia. In Fossil Mammals of Asia; Columbia University Press: New York, NY, USA, 2013; pp. 521–537. [Google Scholar]
- Kurochkin, E.N.; Lungu, A.N. A new ostrich from the Middle Sarmatian of Moldavia. Paleontol. Zhurnal 1970, 1, 118–123. [Google Scholar]
- Solounias, N.; Rivals, F.; Semprebon, G.M. Dietary interpretation and paleoecology of herbivores from Pikermi and Samos (late Miocene of Greece). Paleobiology 2010, 36, 113–136. [Google Scholar] [CrossRef]
- Bachmayer, F.; Zapfe, H. Reste von Struthio aus Pikermi. Proc. Acad. Athens 1962, 37, 247–253. [Google Scholar]
- Michailidis, D.; Roussiakis, S.; Theodorou, G. Palaeoavian remains from the late Miocene localities of Pikermi, Chomateri and Kerassiá: Palaeoecological implications. In Proceedings of the 9th Congress of the Carpathian-Balkan Geological Association, Thessaloniki, Greece, 23–26 September 2010; pp. 23–26. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S.; Konidaris, G.E. Palaeontology of the upper Miocene vertebrate localiti of Nikiti (Chalkidiki Peninsula, Macedonia, Greece) Foreword. Geobios 2016, 49, 29–36. [Google Scholar] [CrossRef]
- Spassov, N. The Turolian Megafauna of West Bulgaria and the character of the Late Miocene “Pikermian biome”. Boll. Soc. Paleontol. Ital. 2002, 41, 69–82. [Google Scholar]
- Boev, Z.; Spassov, N. First record of ostriches (Aves, Struthioniformes, Struthionidae) from the late Miocene of Bulgaria with taxonomic and zoogeographic discussion. Geodiversitas 2009, 31, 493–507. [Google Scholar] [CrossRef]
- Aleksejev, A. Animaux Fossiles du Village Novo-Elisavetovka; Zapiski Novorosiyskogo Institute: Odessa, Ukraine, 1915; pp. 273–453. [Google Scholar]
- Burchak-Abramovich, N. Fossil ostriches of Caucasus and Southern Ukraine. Tr. Estestvenoritoricheskogo Muzeya Im. G. Zardabi 1953, 7, 1–206. [Google Scholar]
- Forsyth Major, C. Sur un gisement d’ossements fossiles dans l’île de Samos, contemporains de l’âge de Pikermi. Comptes Rendus De L’académie Des Sci. Paris 1888, 107, 1178–1181. [Google Scholar]
- Tzankov, T.; Spassov, N.; Stoyanov, K. Neogene-Quaternary Paleogeography and Geodynamics of the Region of Middle Struma River Valley (South.-Western Bulgaria); Publishing House of the South-Western University “N. Rilski”: Blagoevgrad, Bulgaria, 2005; p. 199. [Google Scholar]
- Spassov, N.; Tzankov, T.; Geraads, D. Late Neogene stratigraphy, biochronology, faunal diversity and environments of South-West Bulgaria (Struma River Valley). Geodiversitas 2006, 28, 477–498. [Google Scholar]
- Theodorou, G.; Athanassiou, A.; Roussiakis, S.; Iliopoulos, G. Preliminary remarks on the late Miocene herbivores of Kerassia (Northern Euboea, Greece). Deinsea 2003, 10, 519–530. [Google Scholar]
- Kampouridis, P.; Michailidis, D.; Kargopoulos, N.; Roussiakis, S.; Theodorou, G. First description of an ostrich from the late Miocene of Kerassia (Euboea, Greece): Remarks on its cervical anatomy. Hist. Biol. 2020, 33, 1–8. [Google Scholar] [CrossRef]
- Burchak-Abramovich, N. New data on the Tertiary ostriches of Southern Ukraine. Priroda 1939, 5, 94–97. [Google Scholar]
- Burchak-Abramovich, N.I.; Vekua, A.K. The Fossil Ostrich from Akchagil Layers of Georgia. Acta Zool. Crac. 1971, 16, 1–26. [Google Scholar]
- Tesakov, A. Biostratigraphy of Middle Pliocene–Eopleistocene of Eastern Europe (based on small mammals). Trans. Geol. Inst. 2004, 1–247. [Google Scholar]
- Tesakov, A.S.; Dodonov, A.E.; Titov, V.V.; Trubikhin, V.M. Plio-Pleistocene geological record and small mammal faunas, eastern shore of the Azov Sea, Southern European Russia. Quat. Int. 2007, 160, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Athanassiou, A. A Villafranchian Hipparion-Bearing Mammal Fauna from Sésklo (E. Thessaly, Greece): Implications for the Question of Hippario-Equus Sympatry in Europe. Quaternary 2018, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Ferring, R.; Oms, O.; Agustí, J.; Berna, F.; Nioradze, M.; Shelia, T.; Tappen, M.; Vekua, A.; Zhvania, D.; Lordkipanidze, D. Earliest human occupations at Dmanisi (Georgian Caucasus) dated to 1.85–1.78 Ma. Proc. Natl. Acad. Sci. USA 2011, 108, 10432–10436. [Google Scholar] [CrossRef] [Green Version]
- Burchak-Abramovich, N.; Vekua, A. The fossil ostrich Struthio dmanisensis sp. n. from the Lower Pleistocene of eastern Georgia. Acta Zool. Crac. 1990, 33, 121–132. [Google Scholar]
- Lopatin, A.V.; Vislobokova, I.A.; Lavrov, A.V.; Startsev, D.B.; Gimranov, D.O.; Zelenkov, N.V.; Maschenko, E.N.; Sotnikova, M.V.; Tarasenko, K.K.; Titov, V.V. The Taurida Cave, a New Locality of Early Pleistocene Vertebrates in Crimea. Dokl. Biol. Sci. 2019, 485, 40–43. [Google Scholar] [CrossRef]
- Zelenkov, N.V.; Lavrov, A.V.; Startsev, D.B.; Vislobokova, I.A.; Lopatin, A.V. A giant early Pleistocene bird from eastern Europe: Unexpected component of terrestrial faunas at the time of early Homo arrival. J. Vertebr. Paleontol. 2019, 39, e1605521. [Google Scholar] [CrossRef]
- Mayhew, D.F. Revision of the fossil vole assemblage (Mammalia, Rodentia, Arvicolidae) from Pleistocene deposits at Kisláng, Hungary. Palaeontology 2012, 55, 11–29. [Google Scholar] [CrossRef]
- Kretzoi, M. Bericht über die Calabrische (Villafranchische) Fauna von Kisláng, Kom. Feyér. Jahresber. Der Ung. Geol. Anst. 1954, 1953, 212–264. [Google Scholar]
- Rich, P.V. Significance of the Tertiary avifaunas from Africa (with emphasis on a mid to late Miocene avifauna from southern Tunisia). Ann. Geol. Surv. Egypt 1974, 4, 167–210. [Google Scholar]
- Wang, S. Rediscussion in the taxonomic assignment of “Struthio linxiaensis” Hou, et al., 2005. Acta Paleotologica Sin. 2008, 47, 362–368. [Google Scholar]
- Milne-Edwards, A. Recherches Anatomiques et Paléontologiques pour Servir à l’histoire des Oiseaux Fossiles de la France, Tome Second; Victor Masson: Paris, France, 1869–1871; Volume 1, p. 632. [Google Scholar]
- Mlíkovsky, J.; Chenzychenova, F.; Filippov, A. Quaternary birds of the Baikal region, East Siberia. Acta Soc. Zool. Bohem. 1997, 61, 151–156. [Google Scholar]
- Stimpson, C.M.; Lister, A.; Parton, A.; Clark-Balzan, L.; Breeze, P.S.; Drake, N.A.; Groucutt, H.S.; Jennings, R.; Scerri, E.M.L.; White, T.S.; et al. Middle Pleistocene vertebrate fossils from the Nefud Desert, Saudi Arabia: Implications for biogeography and palaeoecology. Quat. Sci. Rev. 2016, 143, 13–36. [Google Scholar] [CrossRef] [Green Version]
- Field, D.J.; Lynner, C.; Brown, C.; Darroch, S.A. Skeletal correlates for body mass estimation in modern and fossil flying birds. PLoS ONE 2013, 8, e82000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buffetaut, E.; Angst, D. How large was the giant ostrich of China. EVOLUÇÃO Rev. De Geistória E Pré-História 2017, 2, 6–8. [Google Scholar]
- Folch, A.; del Hoyo, J.; Christie, D.A.; Collar, N.; Jutglar, F.; Garcia, E.F.J. Lesser Rhea (Rhea pennata), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Kirwan, G.M.; Korthals, A.; Hodes, C.E. Greater Rhea (Rhea americana), version 2.0. Birds World 2021. [Google Scholar] [CrossRef]
- Delsuc, F.; Superina, M.; Ferraris, G.; Tilak, M.K.; Douzery, E. Molecular evidence for hybridisation between the two living species of South American ratites: Potential conservation implications. Conserv. Genet. 2007, 8, 503–507. [Google Scholar] [CrossRef] [Green Version]
- Miranda-Ribeiro, A. Notas ornithologicas, tinamidae. Rev. Do Mus. Paul. 1938, 23, 667–788. [Google Scholar]
- Bertelli, S.; Chiappe, L.M.; Mayr, G. Phylogenetic interrelationships of living and extinct Tinamidae, volant palaeognathous birds from the New World. Zool. J. Linn. Soc. 2014, 172, 145–184. [Google Scholar] [CrossRef]
- Rayner, J.M. Form and function in avian flight. In Current Ornithology; Springer: New York, NY, USA, 1988; pp. 1–66. [Google Scholar]
- Widrig, K.E.; Watanabe, J.; Bhullar, B.S.; Field, D.J. Three-dimensional atlas of pectoral musculoskeletal anatomy in the extant tinamou Nothoprocta pentlandii (Palaeognathae: Tinamidae). In Proceedings of the The Society of Vertebrate Paleontology 80th Annual Meeting, Virtual, 12–16 October 2020. [Google Scholar]
- Alvarenga, H.M. Uma ave ratitae do paleoceno brasileiro: Bacia calcaŕia de itaboraí, estado do Rio de Janeiro, Brasil. Bol. Do Mus. Nacional. Nova Ser. Geol. 1983, 41, 1–8. [Google Scholar]
- Woodburne, M.O.; Goin, F.J.; Raigemborn, M.S.; Heizler, M.; Gelfo, J.N.; Oliveira, E.V. Revised timing of the South American early Paleogene land mammal ages. J. South. Am. Earth Sci. 2014, 54, 109–119. [Google Scholar] [CrossRef]
- Agnolín, F.L. Unexpected diversity of ratites (Aves, Palaeognathae) in the early Cenozoic of South America: Palaeobiogeographical implications. Alcheringa: An. Australas. J. Palaeontol. 2017, 41, 101–111. [Google Scholar] [CrossRef]
- Alvarenga, H. Diogenornis fragilis Alvarenga, 1985, restudied: A South American ratite closely related to Casuariidae. In Proceedings of the 25th International Ornithological Congress, Campos do Jordão, Brazil, 22–28 August 2010. [Google Scholar]
- Tambussi, C. The fossil Rheiformes from Argentina. Cour. Forsch. Senckenberg 1995, 181, 121–129. [Google Scholar]
- Pascual, R.; Ortiz-Jaureguizar, E. The Gondwanan and South American episodes: Two major and unrelated moments in the history of the South American mammals. J. Mamm. Evol. 2007, 14, 75–137. [Google Scholar] [CrossRef]
- Raigemborn, M.S.; Krause, J.M.; Bellosi, E.; Matheos, S.D. Redefinición estratigráfica del grupo Río Chico (Paleógeno Inferior), en el norte de la cuenca del golfo San Jorge, Chubut. Rev. De La Asoc. Geológica Argent. 2010, 67, 239–256. [Google Scholar]
- Krause, J.; Bellosi, E. Paleosols from the Koluel Kaike Formation (Lower-Middle Eocene) in the South-central Chubut, Argentina. A preliminary analysis. In Proceedings of the Actas del IV Congreso Latinoamericano de Sedimentología, La Plata, Argentina, 11–24 November 2006; pp. 125–136. [Google Scholar]
- Paredes, J.M.; Colombo, F.; Foix, N.; Allard, J.O.; Nillni, A.; Allo, M. Basaltic Explosive Volcanism in a tuff-dominated intraplate setting, Sarmiento formation (Middle Eocene-lower Miocene), Patagonia, Argentina. Lat. Am. J. Sedimentol. Basin Anal. 2008, 15, 77–92. [Google Scholar]
- Kramarz, A.; Garrido, A.; Ribeiro, A.; Ortiz, R. Nuevos registros de vertebrados fósiles de la Formación Chichinales, Mioceno Temprano de la provincia de Río Negro, Argentina. Ameghiniana 2004, 41, 53R. [Google Scholar]
- Agnolín, F.; Chafrat, P. New fossil bird remains from the Chichinales Formation (Early Miocene) of northern Patagonia, Argentina. Ann. De Paleontol. 2015, 101, 87–94. [Google Scholar] [CrossRef]
- Marshall, L.G.; Patterson, B. Geology and Geochronology of the Mammal-Bearing Tertiary of the Valle de Santa María and Río Corral Quemado, Catamarca Province, Argentina; Field Museum of Natural History: Chicago, IL, USA, 1981. [Google Scholar]
- Fleagle, J.G.; Perkins, M.E.; Heizler, M.T.; Nash, B.; Bown, T.M.; Tauber, A.A.; Dozo, M.T.; Tejedor, M.F.; Vizcaíno, S.F.; Kay, R.F. Absolute and relative ages of fossil localities in the Santa Cruz and Pinturas Formations. In Early Miocene Paleobiology in Patagonia: High-Latitude Paleocommunities of the Santa Cruz Formation; Vizcaíno, S.F., Kay, R.F., Bargo, M.S., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 41–58. [Google Scholar]
- Blisniuk, P.M.; Stern, L.A.; Chamberlain, C.P.; Idleman, B.; Zeitler, P.K. Climatic and ecologic changes during Miocene surface uplift in the Southern Patagonian Andes. Earth Planet. Sci. Lett. 2005, 230, 125–142. [Google Scholar] [CrossRef]
- Perkins, M.E.; Fleagle, J.G.; Heizler, M.T.; Nash, B.; Bown, T.; Tauber, A.A.; Dozo, M.T. Tephrochronology of the Miocene Santa Cruz and Pinturas Formations, Argentina. In Early Miocene Paleobiology in Patagonia: High-Latitude Paleocommunities of the Santa Cruz Formation; Vizcaíno, S.F., Kay, R.F., Bargo, M.S., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 23–40. [Google Scholar]
- Cuitiño, J.I.; Fernicola, J.C.; Kohn, M.J.; Trayler, R.; Naipauer, M.; Bargo, M.S.; Kay, R.F.; Vizcaíno, S.F. U-Pb geochronology of the Santa Cruz Formation (early Miocene) at the Río Bote and Río Santa Cruz (southernmost Patagonia, Argentina): Implications for the correlation of fossil vertebrate localities. J. South. Am. Earth Sci. 2016, 70, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Ameghino, F. Enumeración de las aves fósiles de la República Argentina. Rev. Argent. De Hist. Nat. 1891, 1, 441–453. [Google Scholar]
- Buffetaut, E. Tertiary ground birds from Patagonia (Argentina) in the Tournouër collection of the Muséum National d’Histoire Naturelle, Paris. Bull. De La Soc. Geol. De Fr. 2014, 185, 207–214. [Google Scholar] [CrossRef]
- Diederle, J.M.; Noriega, J. New Records Of Birds In The Santa Cruz Formation (Early-Middle Miocene) At The Río Santa Cruz Valley, Patagonia, Argentina. Publicación Electrónica De La Asoc. Paleontológica Argent. 2020, 61, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Forasiepi, A.M.; Martinelli, A.G.; de la Fuente, M.; Dieguez, S.; Bond, M. Paleontology and stratigraphy of the Aisol Formation (Neogene), San Rafael, Mendoza. Cenozoic Geol. Cent. Andes Argentina. SCS Publ. Salta 2011, 135–154. [Google Scholar]
- Agnolín, F.L.; Noriega, J.I. Una Nueva Especie de Ñandú (Aves: Rheidae) del Mioceno Tardío de la Mesopotamia Argentina. Ameghiniana 2012, 49, 236–246. [Google Scholar] [CrossRef]
- Ganduglia, P. Observaciones geológicas en la región de Ingeniero Jacobacci provincia de Río Negro (con énfasis en el Cretácico y Terciario). Bachelor’s Dissertation, Universidad Nacional de Buenos Aires, Buenos Aires, Argentina, 1977. [Google Scholar]
- Cione, A.L.; Azpelicueta, M.d.l.M.; Bond, M.; Carlini, A.A.; Casciotta, J.R.; Cozzuol, M.A.; de la Fuente, M.; Gasparini, Z.; Goin, F.J.; Noriega, J. Miocene vertebrates from Entre Ríos province, eastern Argentina. El Neógeno De Argentina. Ser. Correlación Geológica 2000, 14, 191–237. [Google Scholar]
- Cerdeño, E.; Montalvo, C. Los Mesotheriinae (Mesotheriidae, Notoungulata) del Mioceno superior de La Pampa, Argentina. Rev. Española De Paleontol. 2001, 16, 63–75. [Google Scholar] [CrossRef]
- Verzi, D.; Deschamps, C.; Montalvo, C. Bioestratigrafía y biocronología del Mioceno tardío de Argentina central. Ameghiniana 2004, 41, 21R. [Google Scholar]
- Cenizo, M.M.; Tambussi, C.; Montalvo, P.C.I. Late Miocene continental birds from the Cerro Azul Formation in the Pampean region (central-southern Argentina). Alcheringa: An. Australas. J. Palaeontol. 2012, 36, 47–68. [Google Scholar] [CrossRef]
- Bossi, G.E.; Muruaga, C.M. Estratigrafía e inversión tectónica del ’rift’ neógeno en el Campo del Arenal, Catamarca, NO Argentina. Andean Geol. 2009, 36, 311–340. [Google Scholar]
- Reguero, M.A.; Candela, A.M. Late Cenozoic mammals from the northwest of Argentina. Cenozoic Geol. Cent. Andes Argent. 2011, 458, 411–426. [Google Scholar]
- Noriega, J.I.; Jordan, E.A.; Vezzosi, R.I.; Areta, J.I. A new species of Opisthodactylus Ameghino, 1891 (Aves, Rheidae), from the late Miocene of northwestern Argentina, with implications for the paleobiogeography and phylogeny of rheas. J. Vertebr. Paleontol. 2017, 37, e1278005. [Google Scholar] [CrossRef]
- Deschamps, C.M.; Vucetich, M.G.; Verzi, D.H.; Olivares, A.I. Biostratigraphy and correlation of the Monte Hermoso Formation (early Pliocene, Argentina): The evidence from caviomorph rodents. J. South. Am. Earth Sci. 2012, 35, 1–9. [Google Scholar] [CrossRef]
- Tomassini, R.L.; Montalvo, C.I.; Deschamps, C.M.; Manera, T. Biostratigraphy and biochronology of the Monte Hermoso Formation (early Pliocene) at its type locality, Buenos Aires Province, Argentina. J. South. Am. Earth Sci. 2013, 48, 31–42. [Google Scholar] [CrossRef]
- Rovereto, G. Los estratos araucanos y sus fósiles. An. Del Mus. Nac. De Hist. Nat. De Buenos Aires 1914, 25, 1–247. [Google Scholar]
- Palazzesi, L.; Barreda, V. Fossil pollen records reveal a late rise of open-habitat ecosystems in Patagonia. Nat. Commun. 2012, 1294, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Tambussi, C.P.; Degrange, F.J. South. American and Antarctic Continental Cenozoic Birds: Paleobiogeographic Affinities and Disparities; Springer: Dordrecht, The Netherlands, 2013; p. 113. [Google Scholar]
- Picasso, M.B.J.; Mosto, M.C. New insights about Hinasuri nehuensis (Aves, Rheidae, Palaeognathae) from the early Pliocene of Argentina. Alcheringa: An. Australas. J. Palaeontol. 2016, 40, 244–250. [Google Scholar] [CrossRef]
- Ameghino, C.; Rusconi, C. Nueva subespecie de avestruz fósil del Plioceno de Buenos Aires. An. De La Soc. Científica Argent. 1932, 114, 38–42. [Google Scholar]
- Moreno, F.P.; Mercerat, A. Catálogo de los pájaros fósiles de la República Argentina conservados en el Museo de La Plata. An. Del Mus. De La Plata Paleontol. 1891, 1, 7–71. [Google Scholar]
- Picasso, M.B.J.; Mosto, C. The new taxonomic status of Rhea anchorenensis (Ameghino and Rusconi, 1932) (Aves, Palaeognathae) from the Pleistocene of Argentina. Ann. De Paleontol. 2016, 102, 237–241. [Google Scholar] [CrossRef]
- Picasso, M.B.J. Diversity of extinct Rheidae (Aves, Palaeognathae): Historical controversies and the new taxonomic status of Rhea pampeana Moreno and Mercerat 1891 from the Pleistocene of Argentina. Hist. Biol. 2016, 28, 1101–1107. [Google Scholar] [CrossRef]
- Chiappe, L. Fossil birds from the Miocene Pinturas Formation of southern Argentina. J. Vertebr. Paleontol. 1991, 11, 21A–22A. [Google Scholar]
- Bertelli, S.; Chiappe, L.M. Earliest Tinamous (Aves: Palaeognathae) from the Miocene of Argentina and their Phylogenetic Position; Natural History Museum of Los Angeles County: Los Angeles, CA, USA, 2005. [Google Scholar]
- Chandler, R. A New Species of Tinamou (Aves: Tinamiformes, Tinamidae) From the Early-Middle Miocene of Argentina. PalArch’s J. Vertebr. Palaeontol. 2012, 9, 1–8. [Google Scholar]
- Woodburne, M.O.; Goin, F.J.; Bond, M.; Carlini, A.A.; Gelfo, J.N.; López, G.M.; Iglesias, A.; Zimicz, A.N. Paleogene Land Mammal Faunas of South America; a Response to Global Climatic Changes and Indigenous Floral Diversity. J. Mamm. Evol. 2014, 21, 1–73. [Google Scholar] [CrossRef]
- Brodkorb, P. Notes on fossil Tinamous. Auk 1961, 78, 257. [Google Scholar] [CrossRef]
- Tonni, E.P. Los Tinamidos Fosiles Argentinos I. El Genero Tinamisornis Rovereto, 1914. Ameghiniana 1977, 14, 224–232. [Google Scholar]
- Tambussi, C.; Tonni, E. Un Tinamidae (Aves: Tinamiformes) del Mioceno tardío de La Pampa (República Argentina) y comentarios sobre los tinámidos fósiles argentinos. Rev. De La Asoc. Paleontológica Argent. 1985, 14, 4. [Google Scholar]
- Tambussi, C.P. Catalogo Critico De Los Tinamidae (Aves: Tinamiformes) Fosiles De La Republica Argentina. Ameghiniana 1987, 24, 241–244. [Google Scholar]
- Tambussi, C.P. Las Aves del Plioceno Tardío-Pleistoceno Temprano de la Provincia de Buenos Aires; Universidad Nacional de La Plata: Buenos Aires, Argentina, 1989. [Google Scholar]
- Marshall, L.G.; Berta, A.; Hofstetter, R.; Pascual, R.; Reig, O.; Bombin, M.; Mones, A. Mammals and Stratigraphy: Geochronology of the Continental Mammal-Bearing Quaternary of South America; Laboratoire de paléontologie des vertébrés de l’Ecole pratique des hautes études: Montpellier, France, 1983. [Google Scholar]
- Tambussi, C.; Noriega, J. Summary of the avian fossil record from southern South America. Muenchner Geowiss. Abh. 1996, 30, 245–264. [Google Scholar]
- Gill, F.; Donsker, D.; Rasmussen, P. IOC World Bird List. 2021. Available online: https://www.worldbirdnames.org/new/ (accessed on 30 December 2021).
- Parker, S.A. The extinct Kangaroo Island Emu, a hitherto-unrecognised species. Bull. Br. Ornithol. Club 1984, 104, 19–22. [Google Scholar]
- Spencer, B. The King Island Emu. Vic. Nat. 1906, 23, 140. [Google Scholar]
- Le Souëf, W.H.D. Dromaeus diemenensis. Bull. Br. Ornithol. Club 1907, 21, 13. [Google Scholar]
- Heupink, T.H.; Huynen, L.; Lambert, D.M. Ancient DNA Suggests Dwarf and ‘Giant’ Emu Are Conspecific. PLoS ONE 2011, 6, e18728. [Google Scholar] [CrossRef] [Green Version]
- Thomson, V.A.; Mitchell, K.J.; Eberhard, R.; Dortch, J.; Austin, J.J.; Cooper, A. Genetic diversity and drivers of dwarfism in extinct island emu populations. Biol. Lett. 2018, 14, 20170617. [Google Scholar] [CrossRef] [Green Version]
- Worthy, T.H.; Nguyen, J.M.T. An annotated checklist of the fossil birds of Australia. Trans. R. Soc. South. Aust. 2020, 144, 66–108. [Google Scholar] [CrossRef]
- Folch, A.; Christie, D.A.; Garcia, E.F.J. Emu (Dromaius novaehollandiae), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Boles, W. A new emu (Dromaiinae) from the Late Oligocene Etadunna Formation. Emu Austral. Ornithol. 2001, 101, 317–321. [Google Scholar] [CrossRef]
- Woodburne, M.O.; Macfadden, B.J.; Case, J.A.; Springer, M.S.; Pledge, N.S.; Power, J.D.; Woodburne, J.M.; Springer, K.B. Land mammal biostratigraphy and magnetostratigraphy of the Etadunna Formation (late Oligocene) of South Australia. J. Vertebr. Paleontol. 1994, 13, 483–515. [Google Scholar] [CrossRef]
- Megirian, D.; Prideaux, G.J.; Murray, P.F.; Smit, N. An Australian land mammal age biochronological scheme. Paleobiology 2010, 36, 658–671. [Google Scholar] [CrossRef]
- Patterson, C.; Rich, P. The fossil history of the emus, Dromaius (Aves: Dromaiinae). Rec. South. Aust. Mus. 1987, 21, 85–117. [Google Scholar]
- Boles, W. Revision of Dromaius gidju Patterson and Rich, 1987 from Riversleigh, northwestern Queensland, Australia, with a reassessment of its generic position. Pap. Avian Paleontol. Honor. Pierce Brodkorb 1992, 36, 195–208. [Google Scholar]
- Archer, M.; Hand, S.; Godthelp, H.; Creaser, P. Correlation of the Cainozoic sediments of the Riversleigh World Heritage fossil property, Queensland, Australia. Mémoires Et Trav. De L’institut De Montp. 1997, 131–152. [Google Scholar]
- Travouillon, K.J.; Archer, M.; Hand, S.J.; Godthelp, H. Multivariate analyses of Cenozoic mammalian faunas from Riversleigh, northwestern Queensland. Alcheringa: An. Australas. J. Palaeontol. 2006, 30, 323–349. [Google Scholar] [CrossRef]
- Boles, W. Hindlimb proportions and locomotion of Emuarius gidju (Patterson & Rich, 1987) (Aves: Casuariidae). Mem. Qld. Mus. 1997, 41, 235–240. [Google Scholar]
- Worthy, T.H.; Hand, S.J.; Archer, M. Phylogenetic relationships of the Australian Oligo-Miocene ratite Emuarius gidju Casuariidae. Integr. Zool 2014, 9, 148–166. [Google Scholar] [CrossRef]
- Woodburne, M.; Tedford, R.; Archer, M.; Turnbull, W.; Plane, M. Biochronology of the continental mammal record of Australia and New Guinea. Spec. Publ. South. Aust. Dep. Mines Energy 1985, 5, 347–363. [Google Scholar]
- Rich, P.V. The Dromornithidae, an extinct family of large ground birds endemic to Australia. Bur. Mineral. Resour. Geol. Geophys. (Aust.) Bull. 1979, 184, 1–196. [Google Scholar]
- Rich, P.; Van Tets, J. Fossil birds of Australia and New Guinea: Their biogeographic, phylogenetic and biostratigraphic input. In The Fossil Vertebrate Record of Australasia; Rich, P.V., Thompson, E.M., Eds.; Monash University Offset Printing Unit: Melbourne, Australia, 1982; pp. 235–385. [Google Scholar]
- Woodburne, M.O. The Alcoota Fauna, Central Australia: An Integrated Palaeontological and Geological Study; Australian Government Public Service: Canberra, Australia, 1967; Volume 87. [Google Scholar]
- Stirton, R.A.; Tedford, R.H.; Woodburne, M.O. Australian Tertiary deposits containing terrestrial mammals. Univ. Calif. Publ. Geol. Sci. 1968, 77, 1–30. [Google Scholar]
- Woods, J.T. Fossiliferous fluviatile and cave deposits. J. Geol. Soc. Aust. 1960, 7, 393–403. [Google Scholar]
- Stirton, R.; Tedford, R.; Miller, A. Cenozoic stratigraphy and vertebrate paleontology of the Tirari Desert, South Australia. Rec. South. Aust. Mus. 1961, 14, 19–61. [Google Scholar]
- Miller, A.H. Fossil ratite birds of the late Tertiary of South Australia. Rec. South. Aust. Mus. 1963, 14, 413–420. [Google Scholar]
- Hoch, E.; Holm, P. New K/Ar age determinations of the Awe fauna gangue, Papua New Guinea: Consequences for Papuaustralian late Cenozoic biostratigraphy. Mod. Geol. 1986, 10, 181–195. [Google Scholar]
- Plane, M. Stratigraphy and vertebrate fauna of the Otibanda formation, New Guinea; Bureau of Mineral Resources, Geology and Geophysics: Canberra, Australia, 1967; p. 64. [Google Scholar]
- Miller, A.H. The history and significance of the fossil Casuarius lydekkeri. Rec. Aust. Mus. 1962, 25, 235–238. [Google Scholar] [CrossRef] [Green Version]
- Rothschild, L.W.R.B. On the Former and Present Distribution of the So Called Ratitae Or Ostrich-like Birds with Certain Deductions and a Description of a New Form by CW Andrew. In Proceedings of the Fifth International Ornithological Congress, Berlin, Germany, 4 June 1910; pp. 144–169. [Google Scholar]
- Williams, P.; McDougall, I.; Powell, J. Aspects of the quaternary geology of the Tari-Koroba area, Papua. J. Geol. Soc. Aust. 1972, 18, 333–347. [Google Scholar] [CrossRef]
- Rich, P.V.; Plane, M.; Schroeder, N. A pygmy cassowary (Casuarius lydekkeri) from late Pleistocene bog deposits at Pureni, Papua New Guinea. BMR J. Aust. Geol. Geophys. 1988, 10, 377–389. [Google Scholar]
- Vickers-Rich, P.; Rich, T. Wildlife of Gondwana; Indiana University Press: Bloomington, IN, USA, 1993. [Google Scholar]
- Martin, H.A. Cenozoic climatic change and the development of the arid vegetation in Australia. J. Arid Environ. 2006, 66, 533–563. [Google Scholar] [CrossRef]
- Yates, A.M.; Worthy, T.H. A diminutive species of emu (Casuariidae: Dromaiinae) from the late Miocene of the Northern Territory, Australia. J. Vertebr. Paleontol. 2019, 39, e1665057. [Google Scholar] [CrossRef]
- Campbell, K.E.; Marcus, L. The relationship of hindlimb bone dimensions to body weight in birds. Nat. Hist. Mus. Los Angeles Cty. Sci. Ser. 1992, 36, 395–412. [Google Scholar]
- Naish, D.; Perron, R. Structure and function of the cassowary’s casque and its implications for cassowary history, biology and evolution. Hist. Biol. 2014, 28, 507–518. [Google Scholar] [CrossRef]
- Calder, W.A. The Kiwi. Sci. Am. 1978, 239, 132–143. [Google Scholar] [CrossRef]
- Kinsky, F.C. The consistent presence of paired ovaries in the Kiwi (Apteryx) with some discussion of this condition in other birds. J. Für Ornithol. 1971, 112, 334–357. [Google Scholar] [CrossRef]
- Folch, A.; Jutglar, F.; Garcia, E.F.J. Little Spotted Kiwi (Apteryx owenii), version 1.0. Birds World 2020. [Google Scholar] [CrossRef]
- Weir, J.T.; Haddrath, O.; Robertson, H.A.; Colbourne, R.M.; Baker, A.J. Explosive ice age diversification of kiwi. Proc. Natl. Acad. Sci. USA 2016, 113, E5580–E5587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huynen, L.; Suzuki, T.; Ogura, T.; Watanabe, Y.; Millar, C.D.; Hofreiter, M.; Smith, C.; Mirmoeini, S.; Lambert, D.M. Reconstruction and in vivo analysis of the extinct tbx5 gene from ancient wingless moa (Aves: Dinornithiformes). BMC Evol. Biol. 2014, 14, 75. [Google Scholar] [CrossRef] [Green Version]
- Bunce, M.; Worthy, T.; Ford, T.; Hoppitt, W.; Willerslev, E.; Drummond, A.; Cooper, A. Extreme reversed sexual size dimorphism in the extinct New Zealand moa Dinornis. Nature 2003, 425, 172–175. [Google Scholar] [CrossRef]
- Perry, G.L.W.; Wheeler, A.B.; Wood, J.R.; Wilmshurst, J.M. A high-precision chronology for the rapid extinction of New Zealand moa (Aves, Dinornithiformes). Quat. Sci. Rev. 2014, 105, 126–135. [Google Scholar] [CrossRef]
- Greenwood, R.; Atkinson, I. Evolution of divaricating plants in New Zealand in relation to moa browsing. Proc. New Zealand Ecol. Soc. 1977, 24, 21–33. [Google Scholar]
- Bond, W.; Silander, J. Springs and wire plants: Anachronistic defences against Madagascar’s extinct elephant birds. Proc. R. Soc. B 2007, 274, 1985–1992. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.R.; Richardson, S.J.; McGlone, M.S.; Wilmshurst, J.M. The diets of moa (Aves: Dinornithiformes). New Zealand J. Ecol. 2020, 44, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Allentoft, M.; Rawlence, N.J. Moa’s Ark or volant ghosts of Gondwana? Insights from nineteen years of ancient DNA research on the extinct moa (Aves: Dinornithiformes) of New Zealand. Ann. Anat. 2012, 194, 36–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tennyson, A.J.D. The origin and history of New Zealand’s terrestrial vertebrates. New Zealand J. Ecol. 2010, 34, 6–27. [Google Scholar]
- Suggate, R.P.; Stevens, G.R.; Te Punga, M.T. The Geology of New Zealand; EC Keating, Government Printer: Wellington, New Zealand, 1978; Volume 2, p. 819. [Google Scholar]
- Landis, C.; Campbell, H.; Begg, J.; Mildenhall, D.; Paterson, A.; Trewick, S. The Waipounamu Erosion Surface: Questioning the antiquity of the New Zealand land surface and terrestrial fauna and flora. Geol. Mag. 2008, 145, 173–197. [Google Scholar] [CrossRef] [Green Version]
- Waters, J.M.; Craw, D. Goodbye Gondwana? New Zealand biogeography, geology, and the problem of circularity. Syst. Biol. 2006, 55, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Trewick, S.A.; Paterson, A.M.; Campbell, H.J. Hello New Zealand. J. Biogeogr. 2007, 34, 1–6. [Google Scholar] [CrossRef]
- Cooper, A.; Cooper, R. The Oligocene bottleneck and New Zealand biota: Genetic record of a past environmental crisis. Proc. R. Soc. B 1995, 261, 293–302. [Google Scholar] [CrossRef]
- Carr, L.; McLenachan, P.; Waddell, P.; Gemmell, N.; Penny, D. Analyses of the mitochondrial genome of Leiopelma hochstetteri argues against the full drowning of New Zealand. J. Biogeogr. 2015, 42, 1066–1076. [Google Scholar] [CrossRef]
- Edgecombe, G.D.; Giribet, G. A New Zealand species of the trans-Tasman centipede order Craterostigmomorpha (Arthropoda: Chilopoda) corroborated by molecular evidence. Invertebr. Syst. 2008, 22, 1–15. [Google Scholar] [CrossRef]
- Boyer, S.L.; Giribet, G. Welcome back New Zealand: Regional biogeography and Gondwanan origin of three endemic genera of mite harvestmen (Arachnida, Opiliones, Cyphophthalmi). J. Biogeogr. 2009, 36, 1084–1099. [Google Scholar] [CrossRef]
- Buckley, T.R.; Lord, N.; Ramón-Laca, A.; Allwood, J.; Leschen, R. Multiple lineages of hyper-diverse Zopheridae beetles survived the New Zealand Oligocene Drowning. J. Biogeogr. 2020, 47, 927–940. [Google Scholar] [CrossRef]
- Wallis, G.P.; Jorge, F. Going under down under? Lineage ages argue for extensive survival of the Oligocene marine transgression on Zealandia. Mol. Ecol. 2018, 27, 4368–4396. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.P.; Wheeler, W.C. Revenant clades in historical biogeography: The geology of New Zealand predisposes endemic clades to root age shifts. J. Biogeogr. 2013, 40, 1609–1618. [Google Scholar] [CrossRef]
- Strogen, D.P.; Bland, K.J.; Nicol, A.; King, P.R. Paleogeography of the Taranaki Basin region during the latest Eocene–Early Miocene and implications for the ‘total drowning’ of Zealandia. N. Z. J. Geol. Geophys. 2014, 57, 110–127. [Google Scholar] [CrossRef]
- Mildenhall, D.C.; Pocknall, D.T. Miocene-Pleistocene spores and pollen from Central Otago, South Island, New Zealand. New Zealand Geol. Surv. Paleontol. Bull. 1989, 1–128. [Google Scholar]
- Pole, M.; Douglas, B. A quantitative palynostratigraphy of the Miocene Manuherikia Group, New Zealand. J. R. Soc. New Zealand 1998, 28, 405–420. [Google Scholar] [CrossRef]
- Worthy, T.H.; Worthy, J.P.; Tennyson, A.; Salisbury, S.; Hand, S.; Scofield, R. Miocene fossils show that kiwi (Apteryx, Apterygidae) are probably not phyletic dwarves. In Paleornithological Research 2013—Proceedings of the 8th International Meeting of the Society of Avian Paleontology and Evolution; Natural History Museum: Vienna, Austria, 2013; pp. 63–80. [Google Scholar]
- Calder, W.A. Size, Function, and Life History; Harvard University Press: Cambridge, MA, USA, 1984; p. 448. [Google Scholar]
- Gould, S.J. Of kiwi eggs and the Liberty Bell. Nat. Hist. 1986, 95, 20–29. [Google Scholar]
- Ducatez, S.; Field, D.J. Disentangling the avian altricial-precocial spectrum: Quantitative assessment of developmental mode, phylogenetic signal, and dimensionality. Evolution 2021, 75, 2717–2735. [Google Scholar] [CrossRef]
- Worthy, T.H.; Holdaway, R.N. The Lost World of the Moa: Prehistoric Life of New Zealand; Indiana University Press: Bloomington, IN, USA, 2002; p. 718. [Google Scholar]
- Tennyson, A.J.D.; Tomotani, B.M. A new fossil species of kiwi (Aves: Apterygidae) from the mid-Pleistocene of New Zealand. Hist. Biol. 2021, 1–9. [Google Scholar] [CrossRef]
- Worthy, T. A mid-Pleistocene rail from New Zealand. Alcheringa 1997, 21, 71–78. [Google Scholar] [CrossRef]
- Worthy, T.H.; Tennyson, A.J.D.; Jones, C.; McNamara, J.A.; Douglas, B.J. Miocene waterfowl and other birds from central Otago, New Zealand. J. Syst. Palaeontol. 2007, 5, 1–39. [Google Scholar] [CrossRef] [Green Version]
- Tennyson, A.; Worthy, T.; Jones, C.M.; Scofield, R.; Hand, S. Moa’s Ark: Miocene fossils reveal the great antiquity of moa (Aves: Dinornithiformes) in Zealandia. In Proceedings of the VII International Meeting of the Society of Avian Paleontology and Evolution, ed. W.E. Boles and T.H. Worth. Rec. Aust. Mus. 2010, 62, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Tennyson, A.J.D.; Martinson, P. Extinct birds of New Zealand; Te Papa Press: Wellington, New Zealand, 2006. [Google Scholar]
- Worthy, T.H.; Edwards, A.R.; Millener, P.R. The fossil record of moas (Aves: Dinornithiformes) older than the Otira (last) Glaciation. J. R. Soc. New Zealand 1991, 21, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Forbes, H. On avian remains found under a lava-flow near Timaru. Trans. Proc. New Zealand Inst. 1891, 23, 366–372. [Google Scholar]
- Mathews, W.H.; Curtis, G. Date of the Pliocene-Pleistocene boundary in New Zealand. Nature 1966, 212, 979–980. [Google Scholar] [CrossRef]
- Beu, A.; Edwards, A. New Zealand Pleistocene and late Pliocene glacio-eustatic cycles. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1984, 46, 119–142. [Google Scholar] [CrossRef]
- Oliver, W.R.B. The Moas of New Zealand and Australia; Dominion Museum: Wellington, New Zealand, 1949. [Google Scholar]
- Marshall, P. Occurence of Fossil Moa-bones in the Lower Wanganui Strata. Trans. Proc. New Zealand Inst. 1919, 51, 250–253. [Google Scholar]
- Coates, G. The Rise and Fall of the Southern Alps; Canterbury University Press: Christchurch, New Zealand, 2002; p. 80. [Google Scholar]
- Dewar, R.E. Extinctions in Madagascar: The loss of the subfossil fauna. In Quaternary Extinctions: A Prehistoric Revolution; Martin, P.S., Klein, R.G., Eds.; The University of Arizona Press: Tuscon, CA, USA, 1984; pp. 574–593. [Google Scholar]
- Samonds, K.E.; Godfrey, L.R.; Ali, J.R.; Goodman, S.M.; Vences, M.; Sutherland, M.R.; Irwin, M.T.; Krause, D.W. Spatial and temporal arrival patterns of Madagascar’s vertebrate fauna explained by distance, ocean currents, and ancestor type. Proc. Natl. Acad. Sci. USA 2012, 109, 5352–5357. [Google Scholar] [CrossRef] [Green Version]
- Samonds, K.E. Late Pleistocene bat fossils from Anjohibe Cave, northwestern Madagascar. Acta Chiropterologica 2007, 9, 39–65. [Google Scholar] [CrossRef]
- Angst, D.; Buffetaut, É. Palaeobiology of Giant Flightless Birds; Elsevier: London, UK, 2017; p. 281. [Google Scholar]
- Clarke, S.; Miller, G.; Fogel, M.; Chivas, A.; Murray-Wallace, C. The amino acid and stable isotope biogeochemistry of elephant bird (Aepyornis) eggshells from southern Madagascar. Quat. Sci. Rev. 2006, 25, 2343–2356. [Google Scholar] [CrossRef]
- Torres, C.R.; Clarke, J.A. Nocturnal giants: Evolution of the sensory ecology in elephant birds and other palaeognaths inferred from digital brain reconstructions. Proc. R. Soc. B 2018, 285, 20181540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewar, R.E.; Radimilahy, C.; Wright, H.T.; Jacobs, Z.; Kelly, G.O.; Berna, F. Stone tools and foraging in northern Madagascar challenge Holocene extinction models. Proc. Natl. Acad. Sci. USA 2013, 110, 12583–12588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansford, J.; Wright, P.C.; Rasoamiaramanana, A.; Pérez, V.R.; Godfrey, L.R.; Errickson, D.; Thompson, T.; Turvey, S.T. Early Holocene human presence in Madagascar evidenced by exploitation of avian megafauna. Sci. Adv. 2018, 4, eaat6925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, A.; Clark, G.; Haberle, S.; Higham, T.; Nowak-Kemp, M.; Prendergast, A.; Radimilahy, C.; Rakotozafy, L.M.; Ramilisonina; Schwenninger, J.L.; et al. New evidence of megafaunal bone damage indicates late colonization of Madagascar. PLoS ONE 2018, 13, e0204368. [Google Scholar] [CrossRef] [PubMed]
- Douglass, K.; Hixon, S.; Wright, H.T.; Godfrey, L.R.; Crowley, B.E.; Manjakahery, B.; Rasolondrainy, T.; Crossland, Z.; Radimilahy, C. A critical review of radiocarbon dates clarifies the human settlement of Madagascar. Quat. Sci. Rev. 2019, 221, 105878. [Google Scholar] [CrossRef]
- Hixon, S.W.; Douglass, K.G.; Crowley, B.E.; Rakotozafy, L.M.A.; Clark, G.; Anderson, A.; Haberle, S.; Ranaivoarisoa, J.F.; Buckley, M.; Fidiarisoa, S. Late Holocene spread of pastoralism coincides with endemic megafaunal extinction on Madagascar. Proc. R. Soc. B 2021, 288, 20211204. [Google Scholar] [CrossRef]
- Goodman, S.M.; Patterson, B.D. Natural Change and Human Impact in Madagascar; Smithsonian Institution Press: Washington, DC, USA, 1997. [Google Scholar]
- Crowley, B.E. A refined chronology of prehistoric Madagascar and the demise of the megafauna. Quat. Sci. Rev. 2010, 29, 2591–2603. [Google Scholar] [CrossRef]
- Buffetaut, E. Elephant Birds Under the Sun King? Etienne de Flacourt and the Vouron patra. Bol. Do Cent. Port. De Geo-História E Pré-História 2018, 1, 1. [Google Scholar]
- Barrett, P. Antarctic palaeoenvironment through Cenozoic times-a review. Terra Antarct. 1996, 3, 103–119. [Google Scholar]
- Pross, J.; Contreras, L.; Bijl, P.K.; Greenwood, D.R.; Bohaty, S.M.; Schouten, S.; Bendle, J.A.; Röhl, U.; Tauxe, L.; Raine, J.I.; et al. Persistent near-tropical warmth on the Antarctic continent during the early Eocene epoch. Nature 2012, 488, 73–77. [Google Scholar] [CrossRef]
- Contreras, L.; Pross, J.; Bijl, P.K.; Koutsodendris, A.; Raine, J.; van de Schootbrugge, I.; Bas Brinkhuis, H. Early to Middle Eocene vegetation dynamics at the Wilkes Land Margin (Antarctica). Rev. Palaeobot. Palynol. 2013, 197, 119–142. [Google Scholar] [CrossRef]
- Poole, I.; Hunt, R.J.; Cantrill, D. A Fossil Wood Flora from King George Island: Ecological Implications for an Antarctic Eocene Vegetation. Ann. Bot. 2001, 88, 33–54. [Google Scholar] [CrossRef] [Green Version]
- Tambussi, C.; Noriega, J.; Gbzdzicki, A.; Tatur, A.; Reguero, M.; Vizcaíno, S. Ratite bird from the Paleogene La Meseta formation, Seymour Island, Antarctica. Pol. Polar Res. 1994, 15, 15–20. [Google Scholar]
- Field, D.J. Preliminary paleoecological insights from the Pliocene avifauna of Kanapoi, Kenya: Implications for the ecology of Australopithecus anamensis. J. Hum. Evol. 2017, 140, 1–10. [Google Scholar] [CrossRef]
- Case, J.A.; Woodburne, M.O.; Chaney, D.S. A gigantic phororhacoid(?) bird from Antarctica. J. Paleontol. 1987, 61, 1280–1284. [Google Scholar] [CrossRef]
- Alvarenga, H.M.; Höfling, E. Systematic revision of the Phorusrhacidae (Aves: Ralliformes). Papéis Avulsos De Zool. 2003, 43, 55–91. [Google Scholar] [CrossRef] [Green Version]
- Chávez, M. Fossil birds of Chile and Antarctic Peninsula. Arq. Do Mus. Nac. Rio De Jan. 2007, 65, 551–572. [Google Scholar]
- Tambussi, C.; Acosta Hospitaleche, C. Antarctic birds (Neornithes) during the Cretaceous-Eocene times. Rev. De La Asoc. Geológica Argent. 2007, 62, 604–617. [Google Scholar]
- Alvarenga, H.M.F.; Chiappe, L.M.; Bertelli, S. Phorusrhacids: The Terror Birds. In Living Dinosaurs: The Evolutionary History of Modern Birds; Dyke, G., Kaiser, G., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 187–208. [Google Scholar]
- Cenizo, M. Review of the putative Phorusrhacidae from the Cretaceous and Paleogene of Antarctica: New records of ratites and pelagornithid birds. Pol. Polar Res. 2012, 33, 239–258. [Google Scholar] [CrossRef]
- Acosta Hospitaleche, C.; Jadwiszczak, P.; Clarke, J.; Cenizo, M. The fossil record of birds from the James Ross Basin, West Antarctica. Adv. Polar Sci. 2019, 30, 251–273. [Google Scholar] [CrossRef]
- Pujana, R.R.; Santillana, S.N.; Marenssi, S.A. Conifer fossil woods from the La Meseta Formation (Eocene of Western Antarctica): Evidence of Podocarpaceae-dominated forests. Rev. Palaeobot. Palynol. 2014, 200, 122–137. [Google Scholar] [CrossRef]
- Amenábar, C.R.; Montes, M.; Nozal, F.; Santillana, S. Dinoflagellate cysts of the La Meseta Formation (middle to late Eocene), Antarctic Peninsula: Implications for biostratigraphy, palaeoceanography and palaeoenvironment. Geol. Mag. 2020, 157, 351–366. [Google Scholar] [CrossRef]
- Tambussi, C.; Noriega, J.; Santillana, S.; Marenssi, S. Falconid bird from the Middle Eocene La Meseta Formation, Seymour Island. West Antarctica. J. Vertebr. Paleontol. 1995, 15, 55A. [Google Scholar]
- Cenizo, M.; Noriega, J.I.; Reguero, M.A. A stem falconid bird from the Lower Eocene of Antarctica and the early southern radiation of the falcons. J. Ornithol. 2016, 157, 885–894. [Google Scholar] [CrossRef]
- Goin, F.J.; Reguero, M.A.; Pascual, R.; von Koenigswald, W.; Woodburne, M.O.; Case, J.A.; Marenssi, S.A.; Vieytes, C.; Vizcaíno, S.F. First gondwanatherian mammal from Antarctica. Geol. Soc. Lond. Spec. Publ. 2006, 258, 135–144. [Google Scholar] [CrossRef]
- Gelfo, J.; Goin, F.; Bauzá, N.; Reguero, M. The fossil record of Antarctic land mammals: Commented review and hypotheses for future research. Adv. Polar Sci. 2019, 30, 274–292. [Google Scholar] [CrossRef]
- Woodburne, M.O.; Zinsmeister, W.J. The First Land Mammal from Antarctica and Its Biogeographic Implications. J. Paleontol. 1984, 58, 913–948. [Google Scholar]
- Case, J.A.; Woodburne, M.O.; Chaney, D.S. A new genus of polydolopid marsupial from Antarctica. Geol. Soc. Am. Mem. 1988, 169, 505–522. [Google Scholar]
- Goin, F.J.; Carlini, A.A. An early Tertiary microbiotheriid marsupial from Antarctica. J. Vertebr. Paleontol. 1995, 15, 205–207. [Google Scholar] [CrossRef]
- Goin, F.J.; Case, J.A.; Woodburne, M.O.; Vizcaíno, S.F.; Reguero, M.A. New Discoveries of “Opposum-Like” Marsupials from Antarctica (Seymour Island, Medial Eocene). J. Mamm. Evol. 1999, 6, 335–365. [Google Scholar] [CrossRef]
- Bond, M.; Reguero, M.; Vizcaíno, S.; Marenssi, S. A new ‘South American ungulate’ (Mammalia: Litopterna) from the Eocene of the Antarctic Peninsula. Geol. Soc. Lond. Spec. Publ. 2006, 258, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Bond, M.; Pascual, R.; Reguero, M.; Santillana, S.; Marenssi, S. Los primeros ungulados extinguidos sudamericanos de la Antártida. Ameghiniana 1990, 16, 240. [Google Scholar]
- Bond, M.; Kramarz, A.; Macphee, R.D.E.; Reguero, M. A New Astrapothere (Mammalia, Meridiungulata) from La Meseta Formation, Seymour (Marambio) Island, and a Reassessment of Previous Records of Antarctic Astrapotheres. Am. Mus. Novit. 2011, 2011, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Hooker, J.J. An additional record of a placental mammal (Order Astrapotheria) from the Eocene of West Antarctica. Antarct. Sci. 1992, 4, 107–108. [Google Scholar] [CrossRef]
- Gelfo, J.N.; López, G.M.; Santillana, S.N. Eocene ungulate mammals from West Antarctica: Implications from their fossil record and a new species. Antarct. Sci. 2017, 29, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Vizcaíno, S.F.; Reguero, M.A.; Goin, F.J.; Tambussi, C.P.; Noriega, J.I. Community structure of Eocene terrestrial vertebrates from Antarctic Peninsula. Publicación Electrónica De La Asoc. Paleontológica Argent. 1998, 5. [Google Scholar]
- Scher, H.D.; Martin, E.E. Timing and climatic consequences of the opening of Drake Passage. Science 2006, 312, 428–430. [Google Scholar] [CrossRef] [Green Version]
- Bijl, P.K.; Bendle, J.A.P.; Bohaty, S.M.; Pross, J.; Schouten, S.; Tauxe, L.; Stickley, C.E.; McKay, R.M.; Röhl, U.; Olney, M.; et al. Eocene cooling linked to early flow across the Tasmanian Gateway. Proc. Natl. Acad. Sci. USA 2013, 110, 9645–9650. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zhang, J.; Xu, X.; Witt, C.; Deng, Y.; Chenc, G.; Meng, G.; Feng, S.; Xu, L.; Szekely, T. Phylogeny and Sex Chromosome Evolution of Palaeognathae. J. Genet. Genom. 2021. [Google Scholar] [CrossRef]
- Kimball, R.T.; Wang, N.; Heimer-McGinn, V.; Ferguson, C.; Braun, E.L. Identifying localized biases in large datasets: A case study using the avian tree of life. Mol. Phylogenetics Evol. 2013, 69, 1021–1032. [Google Scholar] [CrossRef]
- Field, D.J.; Berv, J.; Hsiang, A.; Lanfear, R.; Landis, M.; Dornburg, A. Timing the Extant Avian Radiation: The Rise of Modern Birds, and the Importance of Modeling Molecular Rate Variation. Bull. Am. Mus. Nat. History 2020, 440. [Google Scholar] [CrossRef]
- Berv, J.S.; Field, D.J. Genomic Signature of an Avian Lilliput Effect across the K-Pg Extinction. Syst. Biol. 2017, 67, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaulieu, J.M.; O’Meara, B.C.; Crane, P.; Donoghue, M.J. Heterogeneous Rates of Molecular Evolution and Diversification Could Explain the Triassic Age Estimate for Angiosperms. Syst. Biol. 2015, 64, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Mayr, G.; De Pietri, V.L.; Scofield, R.P.; Worthy, T.H. On the taxonomic composition and phylogenetic affinities of the recently proposed clade Vegaviidae Agnolín et al., 2017—Neornithine birds from the Upper Cretaceous of the Southern Hemisphere. Cretac. Res. 2018, 86, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Field, D.J.; Hsiang, A.Y. A North American stem turaco, and the complex biogeographic history of modern birds. BMC Evol. Biol. 2018, 18. [Google Scholar] [CrossRef] [Green Version]
- Mayr, G. Old World Fossil Record of Modern-Type Hummingbirds. Science 2004, 304, 861–864. [Google Scholar] [CrossRef] [Green Version]
- Mayr, G.; Alvarenga, H.; Mourer-Chauviré, C. Out of Africa: Fossils shed light on the origin of the hoatzin, an iconic Neotropic bird. Naturwissenschaften 2011, 98, 961–966. [Google Scholar] [CrossRef]
- Olson, S.L. Evolution of the rails of the South Atlantic islands (Aves: Rallidae). Smithson. Contrib. Zool. 1973, 152, 1–153. [Google Scholar] [CrossRef]
- Wright, N.A.; Steadman, D.W.; Witt, C.C. Predictable evolution toward flightlessness in volant island birds. Proc. Natl. Acad. Sci. USA 2016, 113, 4765–4770. [Google Scholar] [CrossRef] [Green Version]
- McNab, B.K. Energy Conservation and the Evolution of Flightlessness in Birds. Am. Nat. 1994, 144, 628–642. [Google Scholar] [CrossRef]
- Paxinos, E.E.; James, H.F.; Olson, S.L.; Sorenson, M.D.; Jackson, J.; Fleischer, R.C. mtDNA from fossils reveals a radiation of Hawaiian geese recently derived from the Canada goose (Branta canadensis). Proc. Natl. Acad. Sci. USA 2002, 99, 1399–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, K.; Archer, M.; Hand, S.; Godthelp, H. The Rise of Australian Marsupials: A Synopsis of Biostratigraphic, Phylogenetic, Palaeoecologic and Palaeobiogeographic Understanding. In Earth and Life: Global Biodiversity, Extinction Intervals and Biogeographic Perturbations Through Time; Talent, J., Mulder, E.F.J.d., Derbyshire, E., Eds.; International Year of Planet Earth; Springer: Berlin/Heidelberg, Germany, 2012; pp. 983–1078. [Google Scholar]
- Lyson, T.; Miller, I.; Bercovici, A.; Weissenburger, K.; Fuentes, A.J.; Clyde, W.; Hagadorn, J.W.; Butrim, M.J.; Johnson, K.R.; Fleming, R.; et al. Exceptional continental record of biotic recovery after the Cretaceous Paleogene mass extinction. Science 2019, 366, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Van Valkenburgh, B. Major patterns in the history of carnivorous mammals. Annu. Rev. Earth Planet. Sci. 1999, 27, 463–493. [Google Scholar] [CrossRef] [Green Version]
- Figueirido, B.; Palmqvist, P.; Pérez-Claros, J.A.; Janis, C.M. Sixty-six million years along the road of mammalian ecomorphological specialization. Proc. Natl. Acad. Sci. USA 2019, 116, 12698–12703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxwell, E.E.; Larsson, H.C.E. Osteology and myology of the wing of the Emu (Dromaius novaehollandiae), and its bearing on the evolution of vestigial structures. J. Morphol. 2007, 268, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Hudson, G.; Schreiweis, D.; Wang, S.C.; Lancaster, D. A numerical study of the wing and leg muscles of tinamous (Tinamidae). Northwest. Sci. 1972, 46, 207–255. [Google Scholar]
- Suzuki, D.; Chiba, K.; VanBuren, C.S.; Ohashi, T. The Appendicular Anatomy of the Elegant Crested Tinamou (Eudromia elegans); Kitakyushu Museum of Natural History and Human History: Kitakyushu, Japan, 2014. [Google Scholar]
- Witmer, L.M. The extant phylogenetic bracket and the importance of reconstructing soft tissues in fossils. Funct. Morphol. Vertebr. Paleontol. 1995, 1, 19–33. [Google Scholar]
- Hosner, P.A.; Tobias, J.A.; Braun, E.L.; Kimball, R.T. How do seemingly non-vagile clades accomplish trans-marine dispersal? Trait and dispersal evolution in the landfowl (Aves: Galliformes). Proc. R. Soc. B 2017, 284. [Google Scholar] [CrossRef]
- Chiappe, L.M.; Walker, C.A. Skeletal morphology and systematics of the Cretaceous Euenantiornithes (Ornithothoraces: Enantiornithes). In Mesozoic Birds: Above the Heads of Dinosaurs; Chiappe, L.M., Witmer, L.M., Eds.; University of California Press: Berkeley, CA, USA, 2002; pp. 240–267. [Google Scholar]
- O’Connor, J.K.; Chiappe, L.M.; Bell, A. Pre-modern birds: Avian divergences in the Mesozoic. In Living Dinosaurs: The Evolutionary History of Modern Birds; Dyke, G., Kaiser, G., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2011; pp. 39–114. [Google Scholar]
- Chiappe, L.M.; Qingjin, M. Birds of Stone: Chinese Avian Fossils from the Age of Dinosaurs; JHU Press: Baltimore, MD, USA, 2016. [Google Scholar]
- Tschudy, R.H.; Pillmore, C.L.; Orth, C.J.; Gilmore, J.S.; Knight, J.D. Disruption of the Terrestrial Plant Ecosystem at the Cretaceous-Tertiary Boundary, Western Interior. Science 1984, 225, 1030–1032. [Google Scholar] [CrossRef]
- Vajda, V.; Raine, J.I.; Hollis, C.J. Indication of Global Deforestation at the Cretaceous-Tertiary Boundary by New Zealand Fern Spike. Science 2001, 294, 1700–1702. [Google Scholar] [CrossRef]
- Vajda, V.; Bercovici, A. The global vegetation pattern across the Cretaceous–Paleogene mass extinction interval: A template for other extinction events. Glob. Planet. Chang. 2014, 122, 29–49. [Google Scholar] [CrossRef] [Green Version]
- Klein, C.G.; Pisani, D.; Field, D.J.; Lakin, R.; Wills, M.A.; Longrich, N.R. Evolution and dispersal of snakes across the Cretaceous-Paleogene mass extinction. Nat. Commun. 2021, 12, 5335. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.J.; Berv, J.S.; Chester, S.G.B.; Sargis, E.J.; Field, D.J. Ecological selectivity and the evolution of mammalian substrate preference across the K-Pg boundary. Ecol. Evol. 2021, 11, 14540–14554. [Google Scholar] [CrossRef] [PubMed]
- Anders, E.; Wolbach, W.S.; Gilmour, I. Major wildfires at the Cretaceous-Tertiary boundary. In Global Biomass Burning; Levine, J.S., Ed.; The MIT Press: Cambridge, MA, USA, 1991; pp. 485–492. [Google Scholar]
- Robertson, D.S.; Lewis, W.M.; Sheehan, P.M.; Toon, O.B. K-Pg extinction: Reevaluation of the heat-fire hypothesis. J. Geophys. Res. Biogeosci. 2013, 118, 329–336. [Google Scholar] [CrossRef]
- Clyde, W.C.; Ramezani, J.; Johnson, K.R.; Bowring, S.A.; Jones, M.M. Direct high-precision U–Pb geochronology of the end-Cretaceous extinction and calibration of Paleocene astronomical timescales. Earth Planet. Sci. Lett. 2016, 452, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.R.; Ellis, B. A Tropical Rainforest in Colorado 1.4 Million Years After the Cretaceous-Tertiary Boundary. Science 2002, 296, 2379–2383. [Google Scholar] [CrossRef]
- Bell, A.; Chiappe, L.M. A species-level phylogeny of the Cretaceous Hesperornithiformes (Aves: Ornithuromorpha): Implications for body size evolution amongst the earliest diving birds. J. Syst. Palaeontol. 2014, 14, 239–251. [Google Scholar] [CrossRef]
- Dumont, M.; Tafforeau, P.; Bertin, T.; Bhullar, B.; Field, D.; Schulp, A.; Strilisky, B.; Thivichon-Prince, B.; Viriot, L.; Louchart, A. Synchrotron imaging of dentition provides insights into the biology of Hesperornis and Ichthyornis, the last toothed birds. BMC Evol. Biol. 2016, 16, 178. [Google Scholar] [CrossRef] [Green Version]
- Field, D.J.; Hanson, M.; Burnham, D.; Wilson, L.E.; Super, K.; Ehret, D.; Ebersole, J.A.; Bhullar, B.A.S. Complete Ichthyornis skull illuminates mosaic assembly of the avian head. Nature 2018, 557, 96–100. [Google Scholar] [CrossRef]
- Alegret, L.; Thomas, E.; Lohmann, K.C. End-Cretaceous marine mass extinction not caused by productivity collapse. Proc. Natl. Acad. Sci. USA 2012, 109, 728–732. [Google Scholar] [CrossRef] [Green Version]
- Gottschaldt, K. Structure and function of avian somatosensory receptors. In Form and Function in Birds; King, A.S., McLelland, J., Eds.; Academic Press: London, UK, 1985; Volume 3, pp. 375–461. [Google Scholar]
- Cunningham, S.; Castro, I.; Alley, M. A new prey detection mechanism for kiwi (Apteryx spp.) suggests convergent evolution between paleognathous and neognathous birds. J. Anat. 2007, 211, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Crole, M.; Soley, J. Bony Pits in the Ostrich (Struthio camelus) and Emu (Dromaius novaehollandiae) Bill Tip. Anat. Rec. 2017, 300, 1705–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelenitsky, D.K.; Therrien, F.; Ridgely, R.C.; McGee, A.R.; Witmer, L.M. Evolution of olfaction in non-avian theropod dinosaurs and birds. Proc. R. Soc. B 2011, 278, 3625–3634. [Google Scholar] [CrossRef] [PubMed] [Green Version]













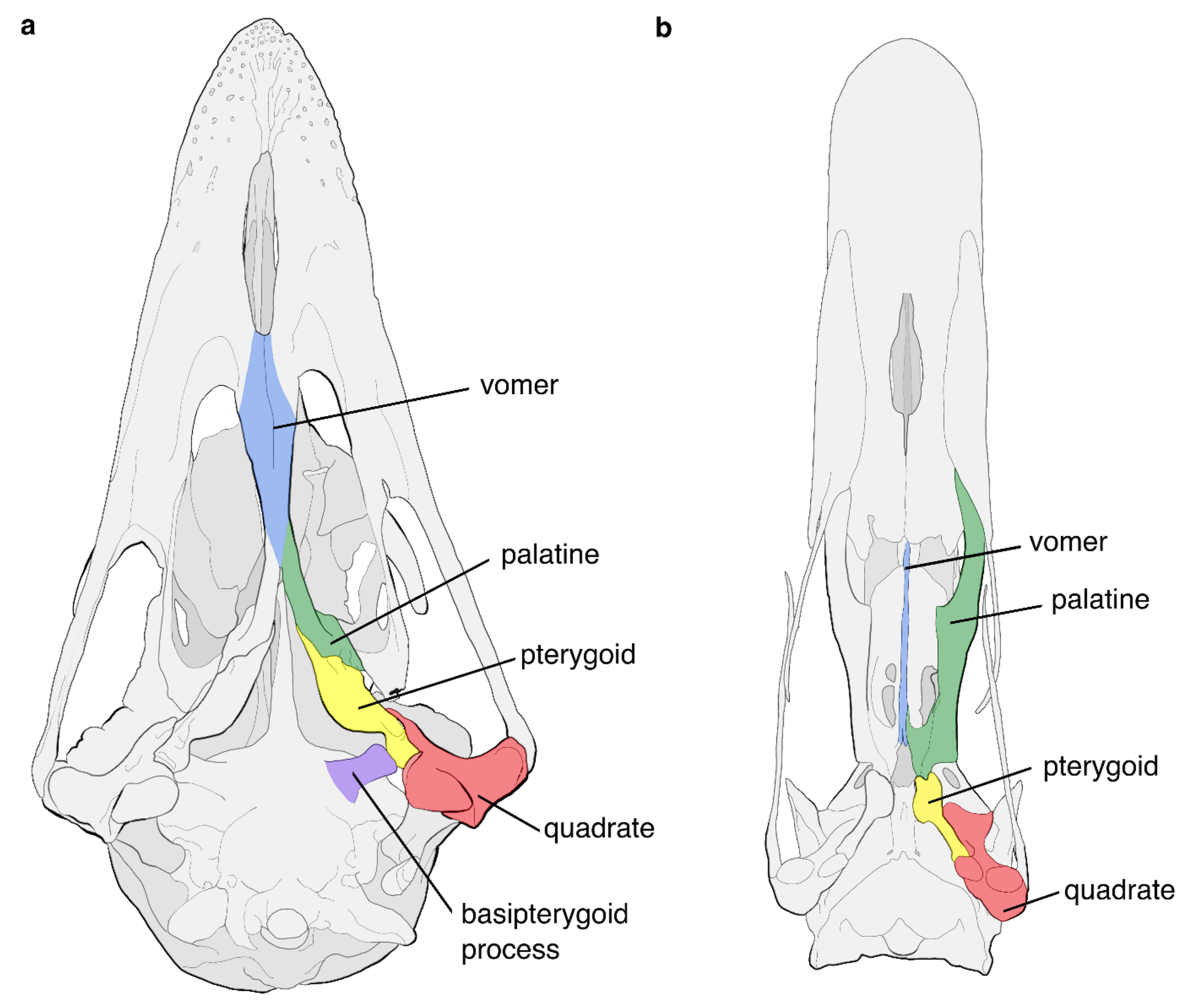{kind=link}
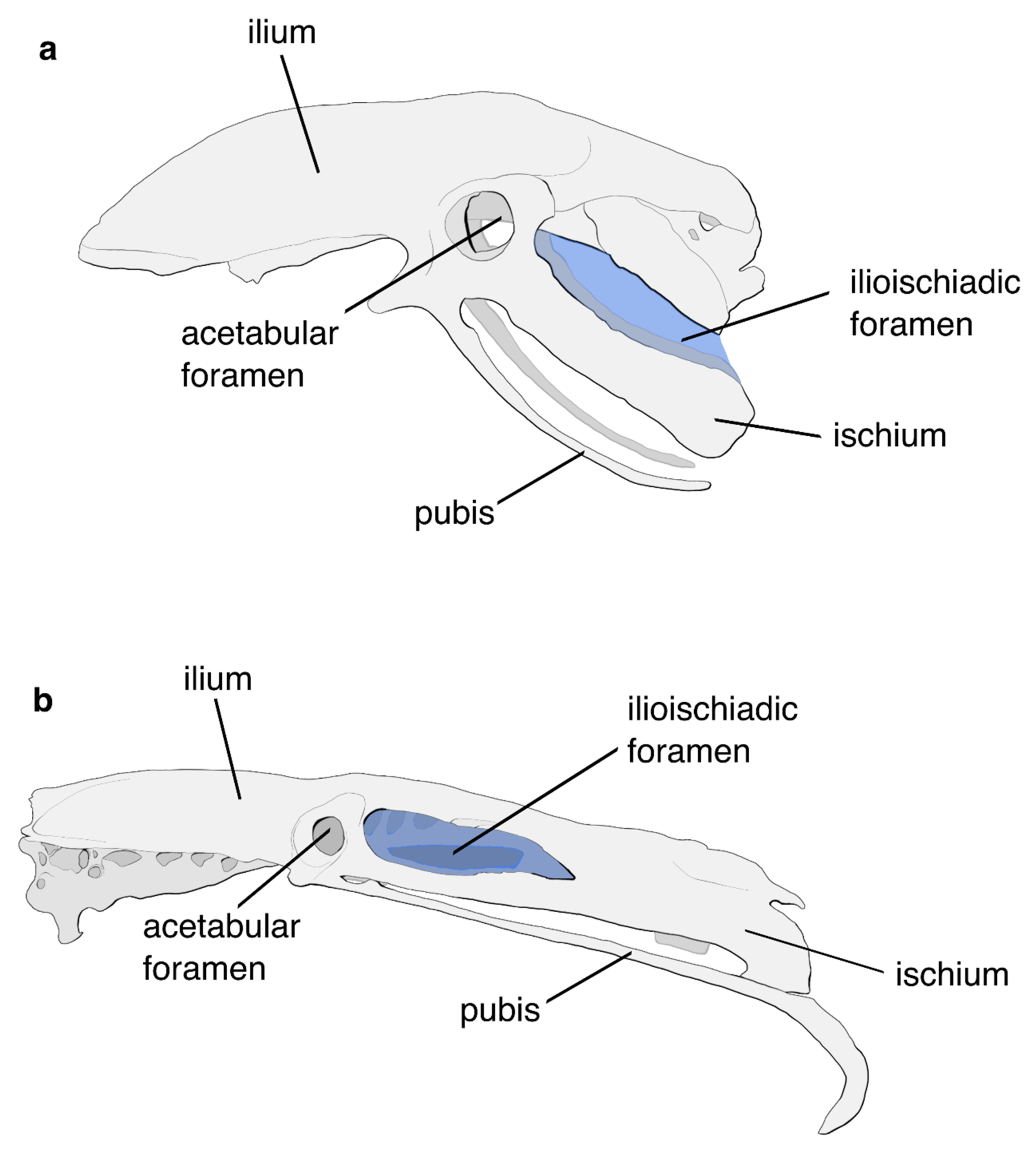{kind=link}
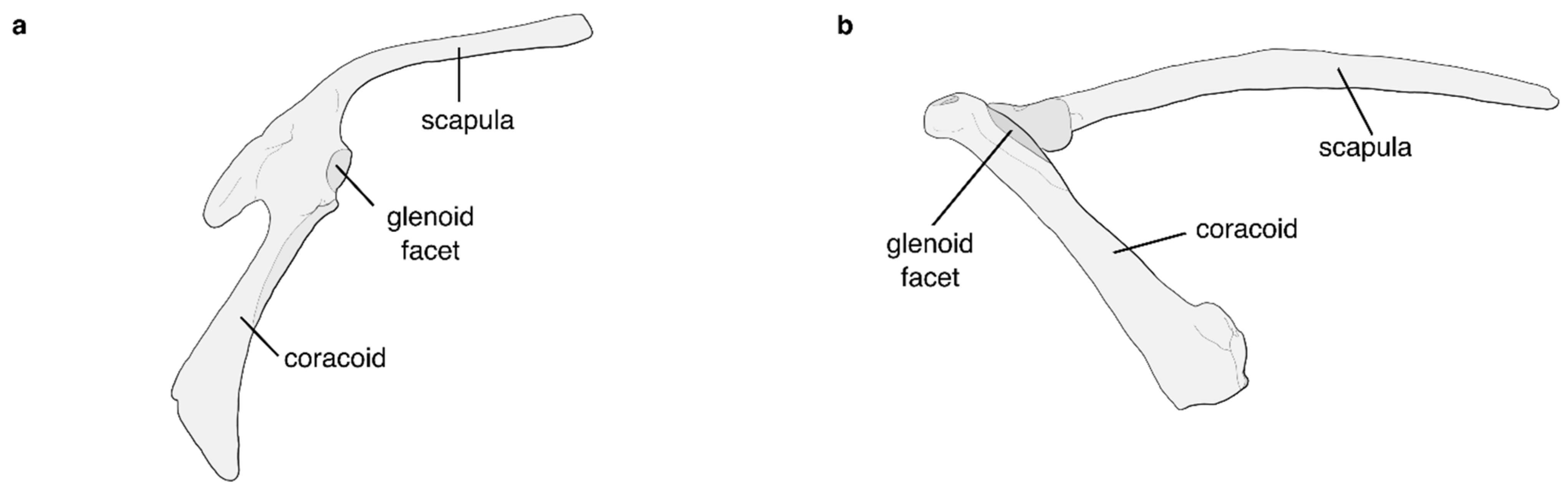{kind=link}
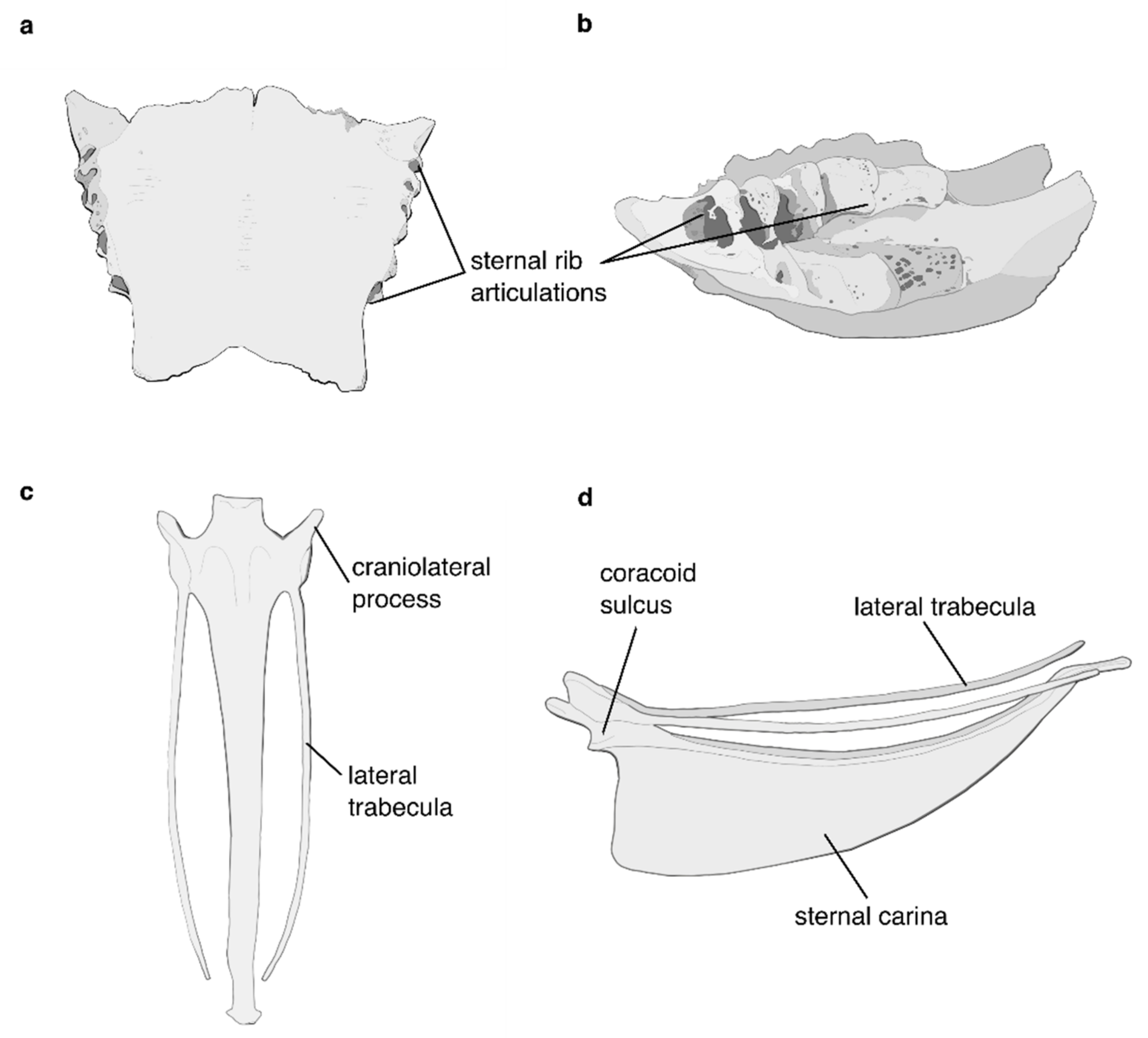{kind=link}
{kind=link}
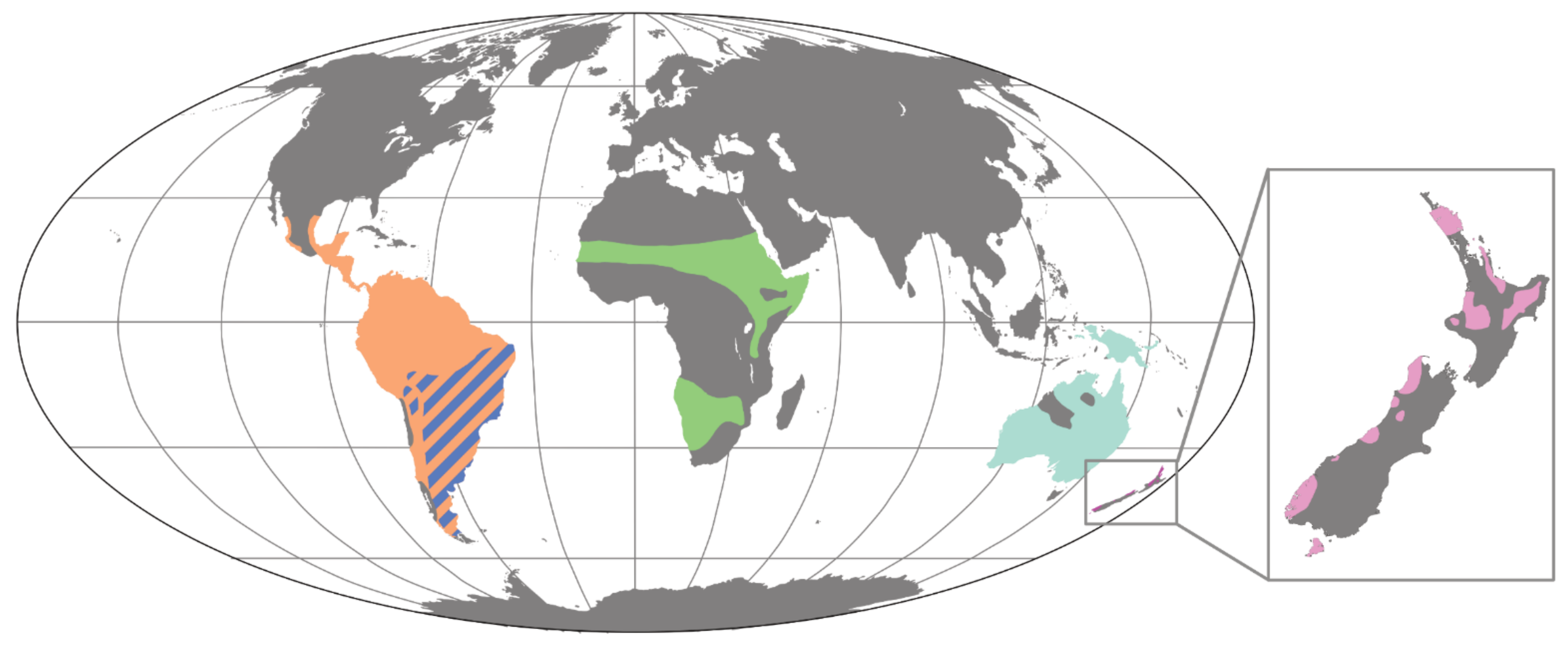{kind=link}
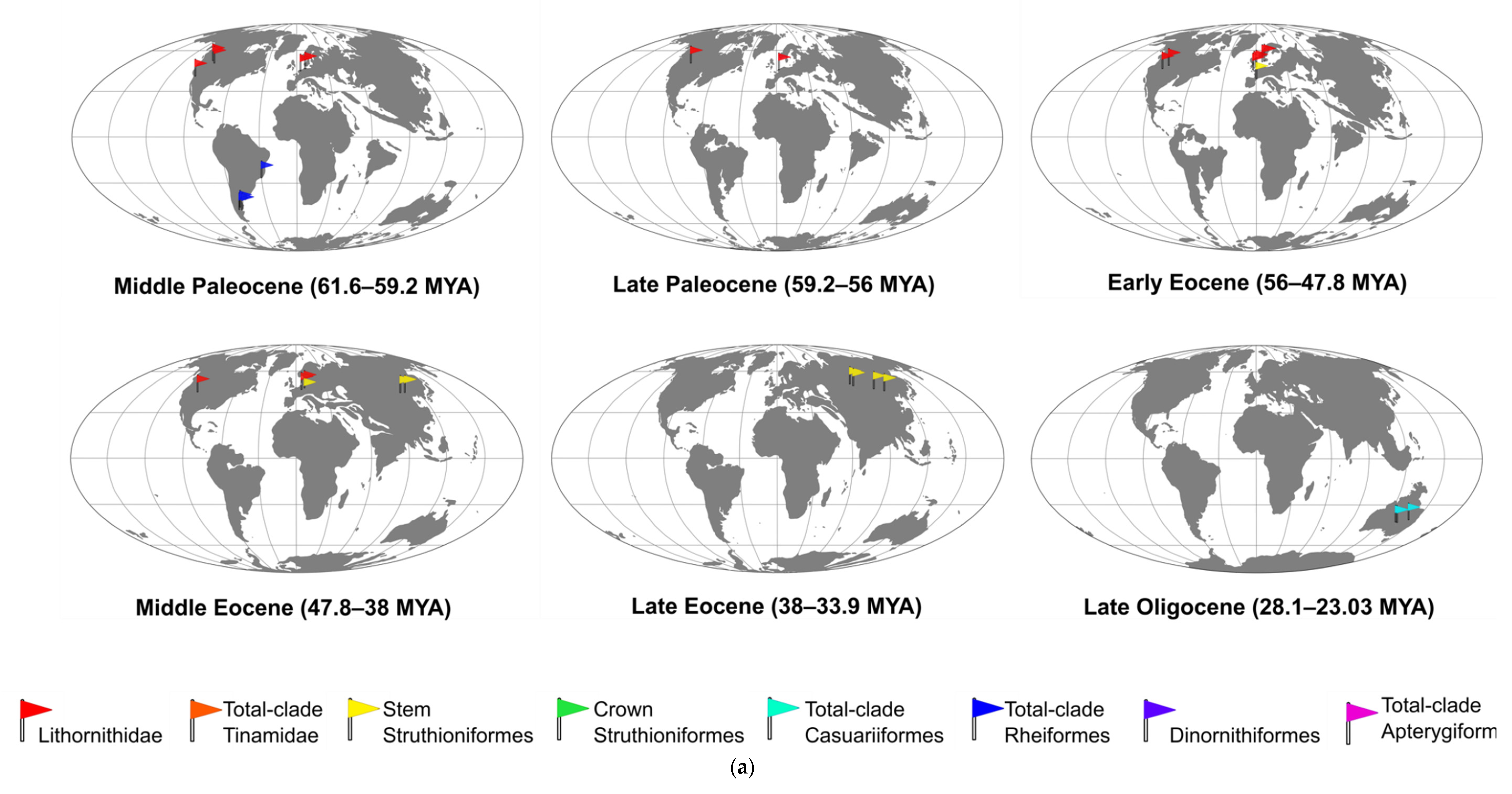{kind=link}
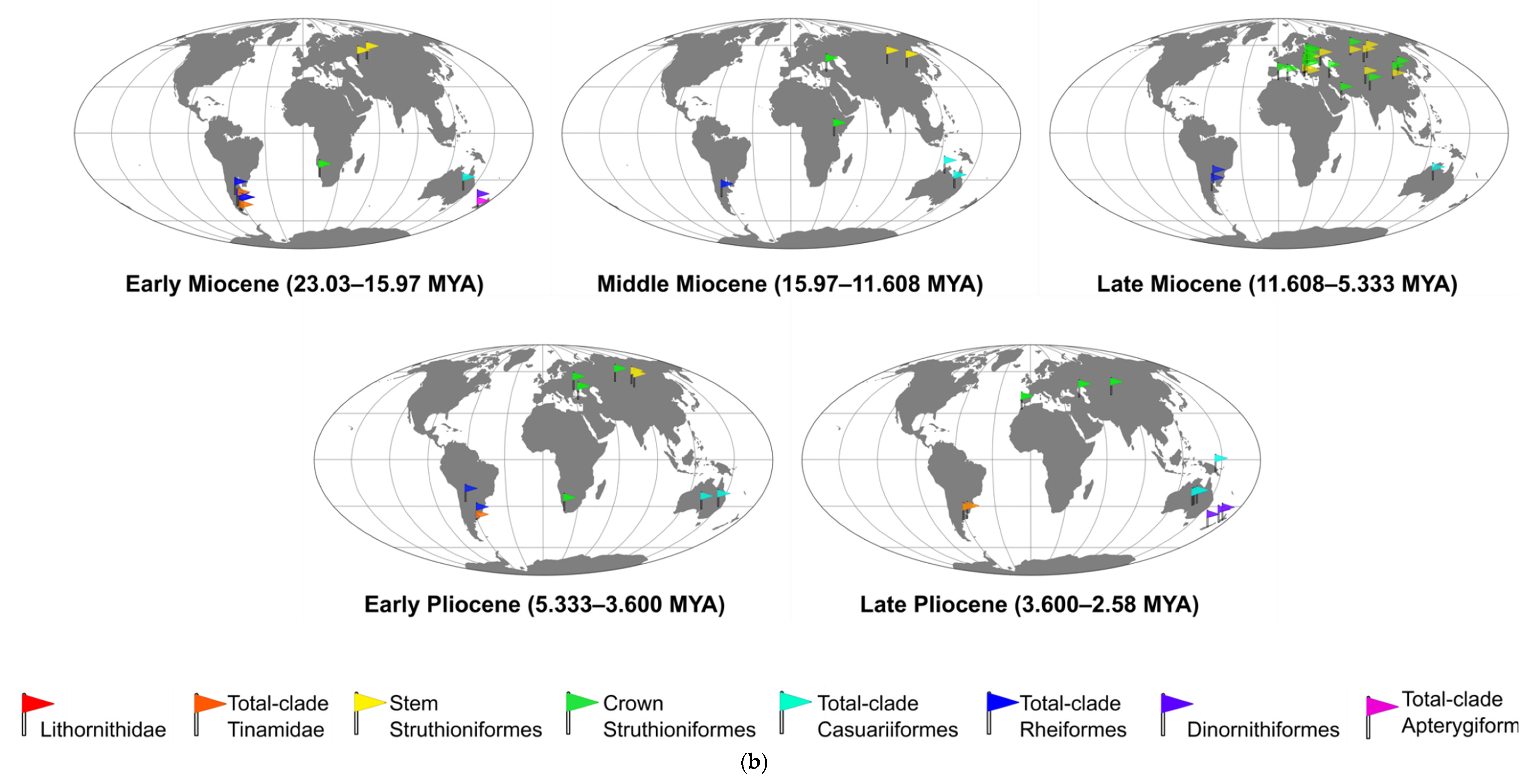{kind=link}
{kind=link}
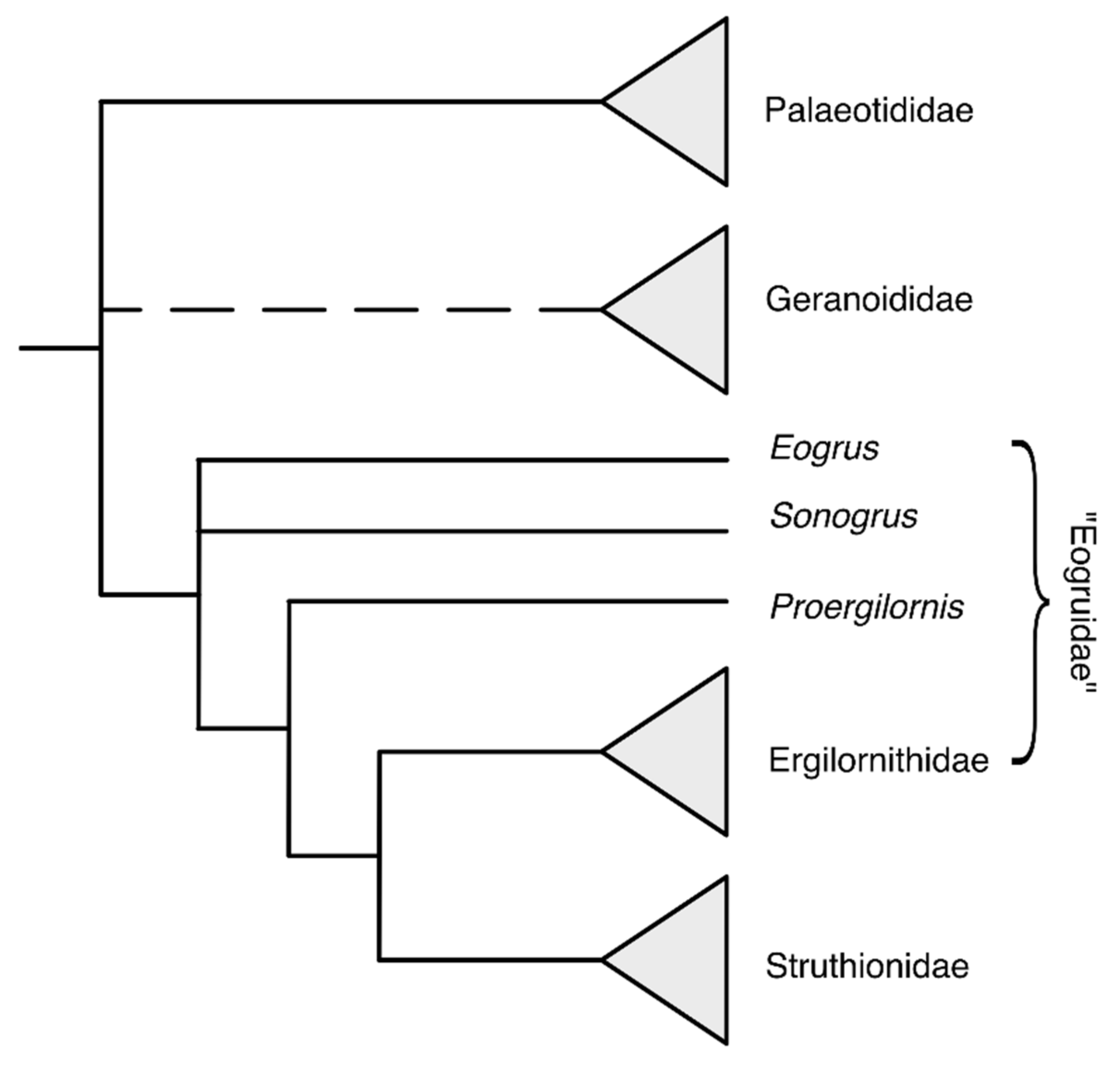{kind=link}
{kind=link}
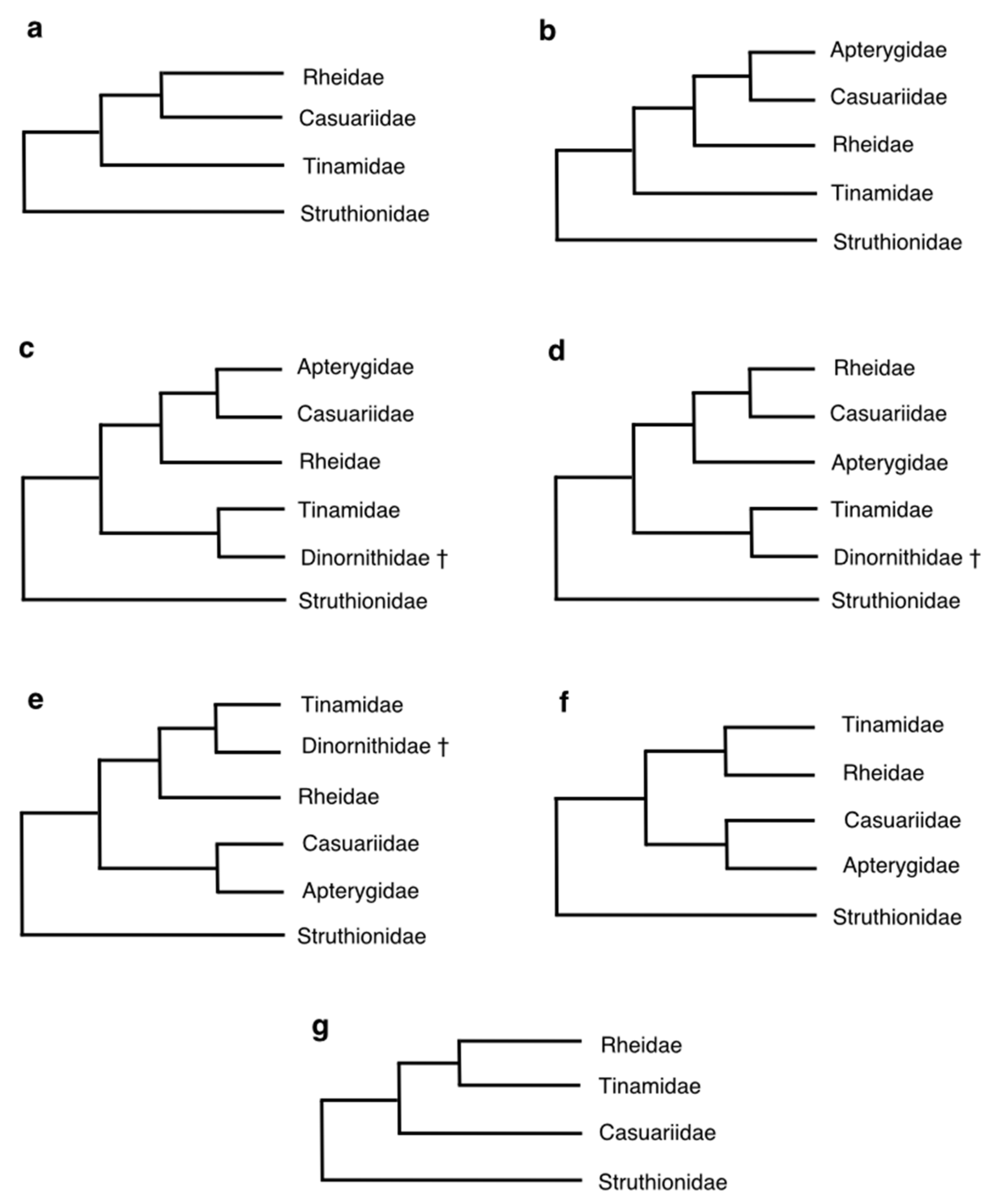{kind=link}
| Continent | Geological Unit | Location | Epoch | Stage | Age Reference | Taxa | Institutions | Reference |
|---|---|---|---|---|---|---|---|---|
| North America | Hornerstown Formation | New Jersey, USA | Late Cretaceous–early Paleocene | Maastrichtian-Danian | Olson and Parris [85]; Staron, et al. [86] | ?Palaeognathae | NJSM | Parris and Hope [63] |
| Fort Union Formation | Park County, Montana, USA | middle Paleocene | Selandian | Lofgren, et al. [87]; Stidham, et al. [88] | Lithornis celetius | USNM, PU | Houde [62] | |
| Polecat Bench Formation | Wyoming, USA | middle Paleocene | Selandian | Lofgren, et al. [87]; Stidham, et al. [88] | Lithornis celetius | PU, UM | Houde [62] | |
| Goler Formation | Kern County, California, USA | middle Paleocene | Selandian | Lofgren, et al. [89]; Albright, et al. [90]; Lofgren, et al. [91] | Lithornis sp. | RAM | Stidham, et al. [88] | |
| Willwood Formation, Sand Coulee beds | Park County, Wyoming, USA | late Paleocene | Thanetian | Lofgren, et al. [87] | Lithornis promiscuus, Lithornis plebius | USNM, UM, AMNH | Houde [62] | |
| Willwood Formation | Basin, Wyoming, USA | early Eocene | Ypresian | Lofgren, et al. [87] | Lithornis nasi (provisional), Paracathartes howardae | UM, ROM, USNM | Houde [62] | |
| Green River Formation, Fossil Butte member | Lincoln County, Wyoming, USA | early Eocene | Ypresian | Smith, et al. [92] | Calciavis grandei, Pseudocrypturus cercanaxius | AMNH, USNM | Houde [62]; Nesbitt and Clarke [64] | |
| Bridger Formation | Bridger Basin, Wyoming, USA | middle Eocene | Ypresian-Lutetian | Murphey and Evanoff [93] | incertae sedis | YPM | Houde [62] | |
| Europe | Heers Formation, Orp Sand member | Maret, Belgium | middle Paleocene | Selandian | Smith and Smith [94], De Bast, et al. [95] | cf. Lithornithidae | IRSNB | Mayr and Smith [96] |
| Fissure filling of Walbeck | Helmstedt, Germany | middle Paleocene | Selandian | Aguilar, et al. [97] | Fissuravis weigelti | GMH | Mayr [98] | |
| Tuffeau de Saint-Omer | Templeuve, France | late Paleocene | Thanetian | Steurbaut [99]; Moreau and Mathis [100]; Smith and Smith [94] | Lithornithidae gen. et sp. indet. | IRSNB | Mayr and Smith [96] | |
| Ølst Formation | Limfjord region, Denmark | early Eocene | Ypresian | Heilmann-Clausen and Schmitz [101] | Lithornis nasi, Lithornis vulturinus | MGUH | Bourdon and Lindow [102] | |
| Fur Formation | Denmark | early Eocene | Ypresian | Chambers, et al. [103] | Lithornis vulturinus | DK, MGUH | Leonard, et al. [104]; Bourdon and Lindow [102] | |
| London Clay Formation | Kent, Essex, Sussex, England | early Eocene | Ypresian | King [105]; Ellison, et al. [106]; Friedman, et al. [107] | Lithornis vulturinus, Lithornis nasi, ?Lithornis hookeri, Pseudocrypturus cercanaxius (provisional) | NHMUK, WN, PU | Houde [62] | |
| Messel Formation | Messel, Germany | middle Eocene | Ypresian-Lutetian | Franzen and Haubold [108]; Schaal and Ziegler [109]; Lenz, et al. [110] | Lithornis sp. | SGPIMH, IRSNB | Mayr [111]; Mayr [112] |
| Continent | Geological Unit | Location | Epoch | Stage | Age Reference | Taxa | Institutions | Reference |
|---|---|---|---|---|---|---|---|---|
| Europe | Châlons-sur-Vesles Formation | Cernay and Berru, Marne, France | late Paleocene | Thanetian | Buffetaut and Angst [160] | Remiornis heberti | MNHN | Lemoine [144]; Martin [161]; Mayr [76] |
| Sables de Bracheux Formation | Rivecourt, France | late Paleocene | Thanetian | Smith, et al. [162] | Remiornis heberti | MV | Buffetaut and de Ploëg [163] | |
| Argiles rutilantes d’Issel et de Saint-Papoul | Saint-Papoul, France | early Eocene | Ypresian | Laurent, et al. [164]; Danilo, et al. [165] | Galligeranoides boriensis | MHNT | Bourdon, et al. [150]; Mayr [148] | |
| Messel Formation | Messel, Germany | middle Eocene | Ypresian-Lutetian | Franzen and Haubold [108]; Schaal and Ziegler [109]; Lenz, et al. [110] | Palaeotis weigelti | HLMD | Peters [146]; Houde and Haubold [143]; Mayr [142] | |
| Geiseltal brown coal | Geisel Valley lignite pits, Germany | middle Eocene | Lutetian | Franzen and Haubold [108] | Palaeotis weigelti | GMH | Lambrecht [145]; Houde and Haubold [143]; Mayr [142]; Mayr [148] | |
| unlisted | Kolkotova Balka, Tiraspol, Moldova | late Miocene | Tortonian-Messinian | Zelenkov and Kurochkin [166] | Urmiornis ukrainus | PIN | Zelenkov and Kurochkin [166] | |
| unlisted | Hrebeniki, Odessa Oblast, Ukraine | late Miocene | Tortonian-Messinian | Zelenkov and Kurochkin [166] | Urmiornis ukrainus | NNPM | Zelenkov and Kurochkin [166] | |
| unlisted | Morozovka, Odessa Oblast, Ukraine | late Miocene | Tortonian-Messinian | Zelenkov and Kurochkin [166] | Urmiornis ukrainus | NNPM | Zelenkov and Kurochkin [166] | |
| unlisted | Armavir, Krasnodar Krai, Russia | late Miocene | Tortonian-Messinian | Zelenkov and Kurochkin [166] | Urmiornis ukrainus | Armavir Regional Museum | Zelenkov and Kurochkin [166] | |
| unlisted | Samos, Greece | late Miocene | Tortonian | Zelenkov, et al. [167] | Ampipelargus majori | NHMUK | Lydekker [168]; Zelenkov, et al. [167] | |
| Triglia Formation | Kryopigi, Chalkidiki, Greece | late Miocene | Tortonian-Messinian | Tsoukala and Bartsiokas [169]; Lazaridis and Tsoukala [170] | ?Ampipelargus sp. | AUG | Boev, et al. [171]; Zelenkov, et al. [167] | |
| Asia | Irdin Manha Formation | Shara Murun region, Inner Mongolia, China | middle Eocene | Lutetian | Li [172] | Eogrus aeola | AMNH, PIN | Wetmore [173]; Kurochkin [152]; Zelenkov and Kurochkin [166] |
| Khaichin Formation | Omnogvi Province, Mongolia | middle Eocene | Lutetian | Zelenkov and Kurochkin [166] | Eogrus aeola | PIN | Zelenkov and Kurochkin [166] | |
| Obayla Formation | Kalmakpai River, East Kazakstan | late Eocene | Priabonian | Clarke, et al. [156] | Eogrus turanicus | PIN | Bendukidze [174]; Zelenkov and Kurochkin [166] | |
| unlisted | Tsagan Khutel, Bayanhongor Province, Mongolia | late Eocene | Priabonian | Russell and Zhai [175] | Eogrus crudus | PIN | Kurochkin [176]; Zelenkov and Kurochkin [166] | |
| unlisted | Alag Tsav, Dornogovi Province, Mongolia | late Eocene | Priabonian | Dashzėvėg [177]; Clarke, et al. [156] | Eogruidae incertae sedis | IGM | Clarke, et al. [156] | |
| Kustovskaya Formation | East Kazakstan | late Eocene | Priabonian | Musser, et al. [178] | Eogrus sp. | PIN | Kozlova [179]; Kurochkin [176]; Musser, et al. [178] | |
| Ergilin Dzo Formation | Dornogovi Province, Mongolia | latest Eocene-earliest Oligocene | Priabonian-Rupelian | Dashzėvėg [177] | Eogrus sp., Ergilornis rapidus, Ergilornis minor, Ergilornis sp., Ergilornithidae incertae sedis, Sonogrus gregalis | PIN | Wetmore [173]; Kozlova [179]; Kurochkin [152]; Kurochkin [176]; Zelenkov and Kurochkin [166]; Mayr and Zelenkov [6] | |
| unlisted | Mynsualmas, Kazakstan | early Miocene | Aquitanian-Burdigalian | Karhu [180]; Zelenkov and Kurochkin [166] | Urmiornis brodkorbi | PIN | Karhu [180]; Zelenkov and Kurochkin [166] | |
| Upper Aral Formation | Altynshokysu, Kazakstan | early Miocene | Aquitanian-Burdigalian | Karhu [180]; Zelenkov and Kurochkin [166] | Urmiornis brodkorbi | PIN | Karhu [180]; Zelenkov and Kurochkin [166] | |
| Tunggur Formation | Shara Murun region, Inner Mongolia, China | middle Miocene | Serravallian | Wang, et al. [181] | Eogrus wetmorei | AMNH | [173]; Brodkorb [182]; Cracraft [183] | |
| unlisted | Sharga, Govi-Altai Province, Mongolia | middle Miocene | Serravallian | Musser, et al. [178] | Ergilornis sp. | Zelenkov, et al. [167]; Musser, et al. [178] | ||
| Nagri and Chinji Formations | Gilgit-Baltistan, Pakistan | late middle- early late Miocene | Serravallian-Tortonian | Barry, et al. [184] | ? Urmiornis cracrafti | Harrison and Walker [185]; Musser, et al. [178] | ||
| unlisted | Maragheh, Iran | late Miocene | Tortonian-Messinian | Musser, et al. [178] | Urmiornis maraghanus | MNHN | Mecquenem [186] | |
| Lower Pavlodar Formation | Pavlodar, Kazakstan | late Miocene | Tortonian-Messinian | Zelenkov and Kurochkin [166] | Urmiornis sp. | PIN | Kurochkin [176]; Zelenkov and Kurochkin [166] | |
| Karabulak Formation | Kalmakpai, Zaisan, East Kazakstan | late Miocene | Tortonian-Messinian | Zelenkov and Kurochkin [166] | Urmiornis orientalis | PIN | Kurochkin [176]; Zelenkov and Kurochkin [166] | |
| Liushu Formation | Zhuangeji town, Gansu, China | late Miocene | Messinian | Fang, et al. [187] | Sinoergilornis guanheensis | IVPP | Musser, et al. [178] | |
| Khirgis-Nur Formation | Khirgis-Nur, Sunur Province, Mongolia | late Miocene | Messinian | Zelenkov and Kurochkin [166] | Urmiornis sp. | PIN | Kurochkin [176]; Zelenkov and Kurochkin [166] | |
| Khirgis-Nur Formation | Chono-Khariakh, Kobdos Province, Mongolia | early Pliocene | Zanclean | Zelenkov and Kurochkin [166] | Urmiornis dzabghanensis | PIN | Kurochkin [188]; Zelenkov and Kurochkin [166] | |
| Khirgis-Nur Formation | Dzagso-Khairkhan-Obo, Ubsunur Province, Mongolia | early Pliocene | Zanclean | Zelenkov and Kurochkin [166] | Urmiornis dzabghanensis | PIN | Kurochkin [188]; Zelenkov and Kurochkin [166] |
| Continent | Geological Unit | Location | Epoch | Stage | Age Reference | Taxa | Institutions | Reference |
|---|---|---|---|---|---|---|---|---|
| Africa | Elisabethfeld silts | Northern Sperrgebiet, Namibia | early Miocene | Aquitanian | Pickford and Senut [196] | Struthio coppensi | Mourer-Chauviré, et al. [197]; Mourer-Chauviré [198] | |
| unlisted | Kadianga West, Kenya | middle Miocene | Langhian | Pickford [199] | Struthio sp. | KNM | Leonard, et al. [200] | |
| unlisted | Central Nyanza, Kenya | middle Miocene | Serravallian | Pickford [199] | Struthio sp. | KNM | Leonard, et al. [200] | |
| unlisted | Ngorora, Kenya | middle Miocene | Serravallian | Pickford [199] | Struthio sp. | KNM | Leonard, et al. [200] | |
| Beglia Formation | Bled el Douarah, Tunisia | late Miocene | Tortonian | Werdelin [201] | Struthio sp. | Rich [202] | ||
| Varswater Formation | Langebaanweg, South Africa | early Pliocene | Zanclean | Roberts, et al. [203] | Struthio cf. asiaticus | Rich [204]; Manegold, et al. [205], but see Mikhailov and Zelenkov [78] | ||
| unlisted | Ahl al Oughlam, Casablanca, Morocco | late Pliocene | Piacenzian | Geraads [206] | Struthio asiaticus | Mourer-Chauviré and Geraads [207], but see Mikhailov and Zelenkov [78] | ||
| Olduvai series | Olduvai Gorge Bed I, Tanzania | early Pliestocene | Gelasian | Hay [208] | Struthio oldawayi | Lowe [209]; Leakey [210] | ||
| unlisted | Aïn Boucherit, Algeria | early Pleistocene | Gelasian | Werdelin [201] | Struthio barbarus | Arambourg [211]; Mikhailov and Zelenkov [78] | ||
| Asia | Turgut strata | Çandir, Turkey | middle Miocene | Langhian | Becker-Platen, et al. [212] | Struthio cf. brachydactylus | BGR | Sauer [213] |
| unlisted | Maragha, Iran | late Miocene | Tortonian | Palaeostruthio karatheodoris | Mecquenem [189]; Lambrecht [214]; Mikhailov and Zelenkov [78] | |||
| Baynunah Formation | United Arab Emirates | late Miocene | Tortonian | Palaeostruthio karatheodoris | Louchart, et al. [215] | |||
| unlisted | Pavlodar, Kazakhstan | late Miocene | Messinian (?) | Palaeostruthio karatheodoris | Tugarinov [216]; Kurochkin [188]; Mikhailov and Zelenkov [78] | |||
| Liushu Formation | Gansu province, China | late Miocene | Tortonian-Messinian | Deng, et al. [217] | Struthio (Orientornis) linxiaensis | Hou, et al. [218] | ||
| unlisted | Baode county, China | late Miocene | Messinian | Kaakinen, et al. [219] | Struthio wimani | Lowe [220]; Mikhailov and Zelenkov [78] | ||
| Dhok Pathan Formation?, Siwalik series | Siwalik Hills, India | late Miocene-early Pliocene | Messinian-Zanclean | Sahni, et al. [221]; Sahni, et al. [222]; Stern, et al. [223]; Patnaik, et al. [224] | Struthio asiaticus | Davies [225]; Lydekker [226]; Mikhailov and Zelenkov [78] | ||
| unlisted | Çalta, Ankara, Turkey | early Pliocene | Zanclean | Ginsburg, et al. [227]; Sen [228]; Janoo and Sen [229] | Struthio sp. | Janoo and Sen [229] | ||
| unlisted | Pavlodar, Kazakhstan | early Pliocene | Zanclean | Struthio chersonensis | Beliaeva [230] | |||
| upper Issykulian Formation | Akterek, Kyrgyzstan | late Pliocene | Piacenzian | Sotnikova, et al. [231] | Pachystruthio transcaucasius | Sotnikova, et al. [231] | ||
| Nihewan Formation | Nihewan Basin, China | early Pleistocene | Gelasian | Cai, et al. [232] | Pachystruthio indet. | MNHN | Buffetaut and Angst [233] | |
| unlisted | Zhoukoudian, China | middle-late Pleistocene | Calabrian- Chibanian | “Struthio anderssoni” | Hou [234] | |||
| Europe | unlisted | Varnitsa, Moldova | late Miocene | Tortonian | Vangengeim and Tesakov [235] | Struthio orlovi | Kurochkin and Lungu [236] | |
| unlisted | Pikermi, Greece | late Miocene | Tortonian | Solounias, et al. [237] | Palaeostruthio cf. karatheodoris | Bachmayer and Zapfe [238]; Michailidis, et al. [239] | ||
| Nikiti Formation | Nikiti, Greece | late Miocene | Tortonian | Palaeostruthio cf. karatheodoris | Koufos, et al. [240] | |||
| unlisted | Hadzhidimovo, Bulgaria | late Miocene | Tortonian | Spassov [241] | Palaeostruthio karatheodoris | NMNHS | Boev and Spassov [242] | |
| unlisted | Novoelizavetovka, Ukraine | late Miocene | Tortonian- Messinian | Vangengeim and Tesakov [235] | Struthio novorossicus | ONU | Aleksejev [243]; Mikhailov and Zelenkov [78] | |
| unlisted | Kuyal’nik, Ukraine | late Miocene | Tortonian-Messinian | Struthio sp. | Burchak-Abramovich [244]; Mikhailov and Zelenkov [78] | |||
| unlisted | Samos, Greece | late Miocene | Tortonian-Messinian | Palaeostruthio karatheodoris | MGL | Forsyth Major [245]; Mikhailov and Zelenkov [78] | ||
| Strumyani Genetic Lithocomplex | Kamimantsi, Bulgaria | late Miocene | Tortonian- Messinian | Tzankov, et al. [246]; Spassov, et al. [247] | Palaeostruthio cf. karatheodoris | NMNHS | Boev and Spassov [242] | |
| unlisted | Kerassia, Greece | late Miocene | Tortonian-Messinian | Theodorou, et al. [248] | Palaeostruthio karatheodoris | Kampouridis, et al. [249] | ||
| unlisted | Grebeniki, Ukraine | late Miocene | Tortonian | Vangengeim and Tesakov [235] | Palaeostruthio karatheodoris, Struthio brachydactylus | Burchak-Abramovich [250]; Mikhailov and Zelenkov [78] | ||
| Odessa Catacombs | Odessa, Ukraine | early Pliocene | Zanclean | Struthio sp. “Odessa Ostrich” | ONU | Burchak-Abramovich [244]; Mikhailov and Zelenkov [78] | ||
| unlisted | Kvabebi, Georgia | late Pliocene | Piacenzian | Pachystruthio transcaucasius | Burchak-Abramovich and Vekua [251]; Mikhailov and Zelenkov [78] | |||
| Khapry Formation | Liventzovka, Rostovskaya Oblast, Russia | early Pleistocene | Gelasian | Tesakov [252]; Tesakov, et al. [253] | Struthio sp. “Odessa Ostrich” | Kurochkin and Lungu [236] | ||
| Sésklo basin sedimentary fill | Sésklo, Thessaly, Greece | early Pleistocene | Gelasian | Struthio cf. chersonensis | Athanassiou [254] | |||
| unlisted | Dmanisi, Georgia | early Pleistocene | Gelasian | Ferring, et al. [255] | Pachystruthio dmanisensis | Burchak-Abramovich and Vekua [256]; Mikhailov and Zelenkov [78] | ||
| Taurida Cave | Taurida, Crimea | early Pleistocene | Gelasian | Lopatin, et al. [257] | Pachystruthio dmanisensis | Lopatin, et al. [257]; Zelenkov, et al. [258] | ||
| unlisted | Kisláng, Hungary | early-middle Pleistocene | Gelasian-Calabrian | Mayhew [259] | Pachystruthio pannonicus | GMB | Kretzoi [260]; Mikhailov and Zelenkov [78] |
| Continent | Geological Unit | Location | Epoch | Stage | Age Reference | Taxa | Institutions | Reference |
|---|---|---|---|---|---|---|---|---|
| South America | Itaboraí Formation | São José, Brazil | late Paleocene | Selandian | Pascual and Ortiz-Jaureguizar [280] | Diogenornis fragilis | Alvarenga [275] | |
| Rio Chico Formation | Chubut province, Argentina | late Paleocene | Thanetian | Raigemborn, et al. [281] | Diogenornis sp., Rheidae indet. | MACN | Tambussi [279]; Agnolín [277] | |
| Koluel Kaike Formation | El Gauchito, Chubut province, Argentina | late Paleocene | Thanetian | Krause and Bellosi [282] | gen. et sp. indet. | MLP | Agnolín [277] | |
| Sarmiento Formation | Chubut province, Argentina | middle Eocene to early Miocene | unknown | Paredes, et al. [283] | gen. et sp. Indet. | MACN | Agnolín [277] | |
| Chichinales Formation | Río Negro province, Argentina | early Miocene | Burdigalian | Kramarz, et al. [284] | Opisthodactylus horacioperezi | MPCN | Agnolín and Chafrat [285] | |
| Santa Cruz Formation | Santa Cruz province, Argentina | early Miocene | Burdigalian-Langhian | Marshall and Patterson [286]; Fleagle, et al. [287]; Blisniuk, et al. [288]; Perkins, et al. [289]; Cuitiño, et al. [290] | Opisthodactylus patagonicus | NHMUK, MPM, YPM, MNHN | Ameghino [291]; Buffetaut [292]; Diederle and Noriega [293] | |
| Aisol Formation | Mendoza province, Argentina | early Miocene | Burdigalian-Langhian | Forasiepi, et al. [294] | Pterocnemia cf. mesopotamica | FMNH | Agnolín and Noriega [295] | |
| Level 13 of Ganduglia (1977) | Río Negro province, Argentina | middle Miocene | Langhian | Ganduglia [296] | gen et sp. indet. | MLP | Agnolín [277] | |
| Ituzaingó Formation | Entre Ríos province, Argentina | late Miocene | Messinian | Cione, et al. [297] | Pterocnemia mesopotamica, Pterocnemia sp., Rheidae indet. | MACN, MASP, CICYTTP | Agnolín and Noriega [295] | |
| Cerro Azul Formation | La Pampa province, Argentina | late Miocene | Messinian | Cerdeño and Montalvo [298]; Verzi, et al. [299] | Pterocnemia sp. | GHUNLP | Cenizo, et al. [300] | |
| Andalhuala Formation | Tucumán province, Argentina | late Miocene-early Pliocene | Messinian-Zanclean | Marshall and Patterson [286]; Bossi and Muruaga [301]; Reguero and Candela [302] | Opisthodactylus kirchneri | MUFYCA | Noriega, et al. [303] | |
| Monte Hermoso Formation | Buenos Aires province, Argentina | early Pliocene | Zanclean | Deschamps, et al. [304]; Tomassini, et al. [305] | Heterorhea dabbenei, Hinasuri nehuensis | MLP | Rovereto [306]; Tambussi [279] |
| Continent | Geological Unit | Location | Epoch | Stage | Age Reference | Taxa | Institutions | Reference |
|---|---|---|---|---|---|---|---|---|
| South America | Pinturas Formation | Santa Cruz province, Argentina | early Miocene | Burdigalian | Fleagle, et al. [287] | Tinamidae gen. et sp. indet | MACN | Bertelli and Chiappe [315] |
| Santa Cruz Formation | Santa Cruz province, Argentina | early Miocene | Burdigalian | Marshall and Patterson [286]; Fleagle, et al. [287]; Blisniuk, et al. [288]; Perkins, et al. [289]; Cuitiño, et al. [290] | Crypturellus reai, Tinamidae gen. et sp. indet | MPM, MACN, AMNH | Bertelli and Chiappe [315]; Chandler [316] | |
| Cerro Azul Formation | La Pampa province, Argentina | late Miocene | Messinian | Cerdeño and Montalvo [298]; Verzi, et al. [299] | Eudromia sp., Nothura sp. | MLP, GHUNLP | Cenizo, et al. [300] | |
| Monte Hermoso Formation | Buenos Aires province, Argentina | early Pliocene | Zanclean | Deschamps, et al. [304]; Tomassini, et al. [305] | Eudromia olsoni, Eudromia cf. elegans, Roveretornis intermedius, Tinamisornis parvulus | MACN | Brodkorb [318]; Tambussi and Tonni [320]; Tomassini, et al. [305] | |
| Chapadmalal Formation | Buenos Aires province, Argentina | late Pliocene | Zanclean-Piacenzian | Marshall, et al. [323];Deschamps, et al. [304] | Eudromia elegans, Eudromia sp., Nothura parvula, Nothura darwinii | MLP | Tambussi and Noriega [324]; Tambussi and Degrange [308] |
| Continent | Geological Unit | Location | Epoch | Stage | Age Reference | Taxa | Institutions | Reference |
|---|---|---|---|---|---|---|---|---|
| Australia | Etadunna Formation | Lake Palankarinna, South Australia, Australia | late Oligocene | Chattian | Woodburne, et al. [334]; Megirian, et al. [335] | Emuarius guljaruba | SAM | Boles [333] |
| Wipajiri Formation | Etadunna Station, South Australia, Australia | latest Oligocene-early Miocene | Chattian-Aquitanian | Woodburne, et al. [334]; Megirian, et al. [335] | Emuarius gidju | SAM, AM | Patterson and Rich [336]; Boles [337] | |
| Riversleigh faunal zones A-C | Riversleigh, Queensland, Australia | latest Oligocene-middle Miocene | Chattian-Langhian | Archer, et al. [338]; Travouillon, et al. [339]; Megirian, et al. [335] | Emuarius gidju | AM, QM | Boles [337]; Boles [340]; Worthy, et al. [341] | |
| Camfield beds | Bullock Creek, Northern Territory, Australia | middle Miocene | unknown | Woodburne, et al. [342] | Dromaius sp. | Rich [343]; Rich and Van Tets [344] | ||
| Waite Formation | Alcoota, Northern Territory, Australia | late Miocene | unknown | Rich [343] | Dromaius sp. | QM, UCMP | Woodburne [345]; Stirton, et al. [346]; Rich [343]; Rich and Van Tets [344] | |
| Chinchilla Sands | Chinchilla, Queensland, Australia | early Pliocene | Zanclean | Rich and Van Tets [344] | Dromaius novaehollandiae | QM | Woods [347]; Stirton, et al. [346]; Rich and Van Tets [344] | |
| Tirari Formation | Lake Palankarinna, South Australia, Australia | late Pliocene-early Pleistocene | Piacenzian-Gelasian | Stirton, et al. [348]; Rich and Van Tets [344] | Dromaius ocypus | UCMP | Miller [349]; Rich [343]; Rich and Van Tets [344] | |
| New Guinea | Otibanda Formation | Morobe, Papua New Guinea | late Pliocene | Piacenzian | Hoch and Holm [350] | Casuarius sp. | UCMP | Plane [351]; Rich and Van Tets [344] |
| Cave deposits | unknown | Pleistocene? | Unknown | Lydekker [168]; Miller [352] | Casuarius lydekkeri | AM | Lydekker [168]; Rothschild [353]; Miller [352]; Worthy, et al. [341] | |
| Pleistocene swamp deposits | Pureni, Papua New Guinea | late Pleistocene | Chibanian | Williams, et al. [354] | Casuarius lydekkeri | CPC | Rich, et al. [355] |
| Continent | Geological Unit | Location | Epoch | Stage | Age Reference | Taxa | Institutions | Reference |
|---|---|---|---|---|---|---|---|---|
| New Zealand | Bannockburn Formation | Otago, South Island, New Zealand | late early Miocene | Burdigalian | Mildenhall and Pocknall [385]; Pole and Douglas [386] | Proapteryx micromeros | NMNZ | Worthy, et al. [387] |
| Kaimatira Pumice | Marton, North Island, New Zealand | middle Pleistocene | Calabrian | Worthy [393] | Apteryx littoralis | NMNZ | Tennyson and Tomotani [392] |
| Continent | Geological Unit | Location | Epoch | Stage | Age Reference | Taxa | Institutions | Reference |
|---|---|---|---|---|---|---|---|---|
| New Zealand | Bannockburn Formation | Otago, South Island, New Zealand | late early Miocene | Burdigalian | Mildenhall and Pocknall [385]; Pole and Douglas [386] | Dinornithidae indet. | NMNZ | Tennyson, et al. [395] |
| unlisted | Timaru, South Island, New Zealand | early Pleistocene | Gelasian | Mathews and Curtis [399] | Anomalopteryx didiformis | Forbes [398]; Worthy, et al. [397] | ||
| unlisted | Hawke’s Bay, North Island, New Zealand | early Pleistocene? | Gelasian? | Beu and Edwards [400] | Eurapteryx curtus | AIM | Worthy, et al. [397] | |
| unlisted | Wairapa, North Island, New Zealand | early Pleistocene? | Gelasian? | Oliver [401]; Beu and Edwards [400] | “Eurapteryx geranoides” | NMNZ | Worthy, et al. [397] | |
| Tewkesbury Formation | Wanganui, North Island, New Zealand | early Pleistocene | Calabrian | Beu and Edwards [400] | Dinornis novaezealanidae, Emeidae indet. | NMNZ | Marshall [402]; Worthy, et al. [397] |
| Continent | Geological Unit | Location | Epoch | Stage | Age Reference | Taxa | Institutions | Reference |
|---|---|---|---|---|---|---|---|---|
| Antarctica | La Meseta Formation | Seymour Island | late Eocene | Lutetian- Priabonian | Amenábar, et al. [432] | “ratititae” | MLP, UCR | Tambussi, et al. [422]; Cenizo [429]; Acosta Hospitaleche, et al. [430] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Widrig, K.; Field, D.J. The Evolution and Fossil Record of Palaeognathous Birds (Neornithes: Palaeognathae). Diversity 2022, 14, 105. https://doi.org/10.3390/d14020105
Widrig K, Field DJ. The Evolution and Fossil Record of Palaeognathous Birds (Neornithes: Palaeognathae). Diversity. 2022; 14(2):105. https://doi.org/10.3390/d14020105
Chicago/Turabian StyleWidrig, Klara, and Daniel J. Field. 2022. "The Evolution and Fossil Record of Palaeognathous Birds (Neornithes: Palaeognathae)" Diversity 14, no. 2: 105. https://doi.org/10.3390/d14020105
APA StyleWidrig, K., & Field, D. J. (2022). The Evolution and Fossil Record of Palaeognathous Birds (Neornithes: Palaeognathae). Diversity, 14(2), 105. https://doi.org/10.3390/d14020105