
Farming Intensity Affects Soil Seedbank Composition and Spontaneous Vegetation of Arable Weeds

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Experimental Setup
2.2. Statistical Analysis
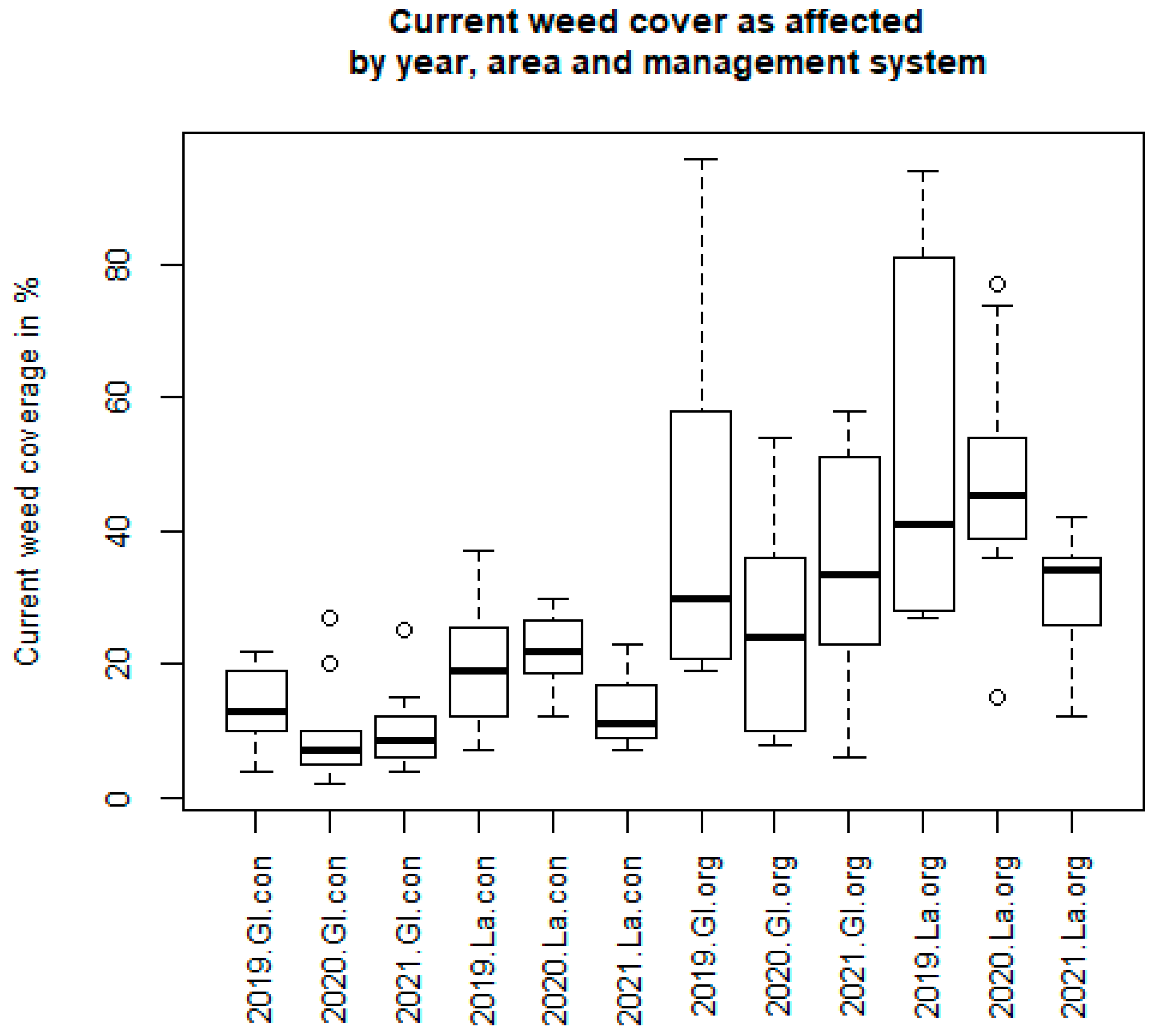
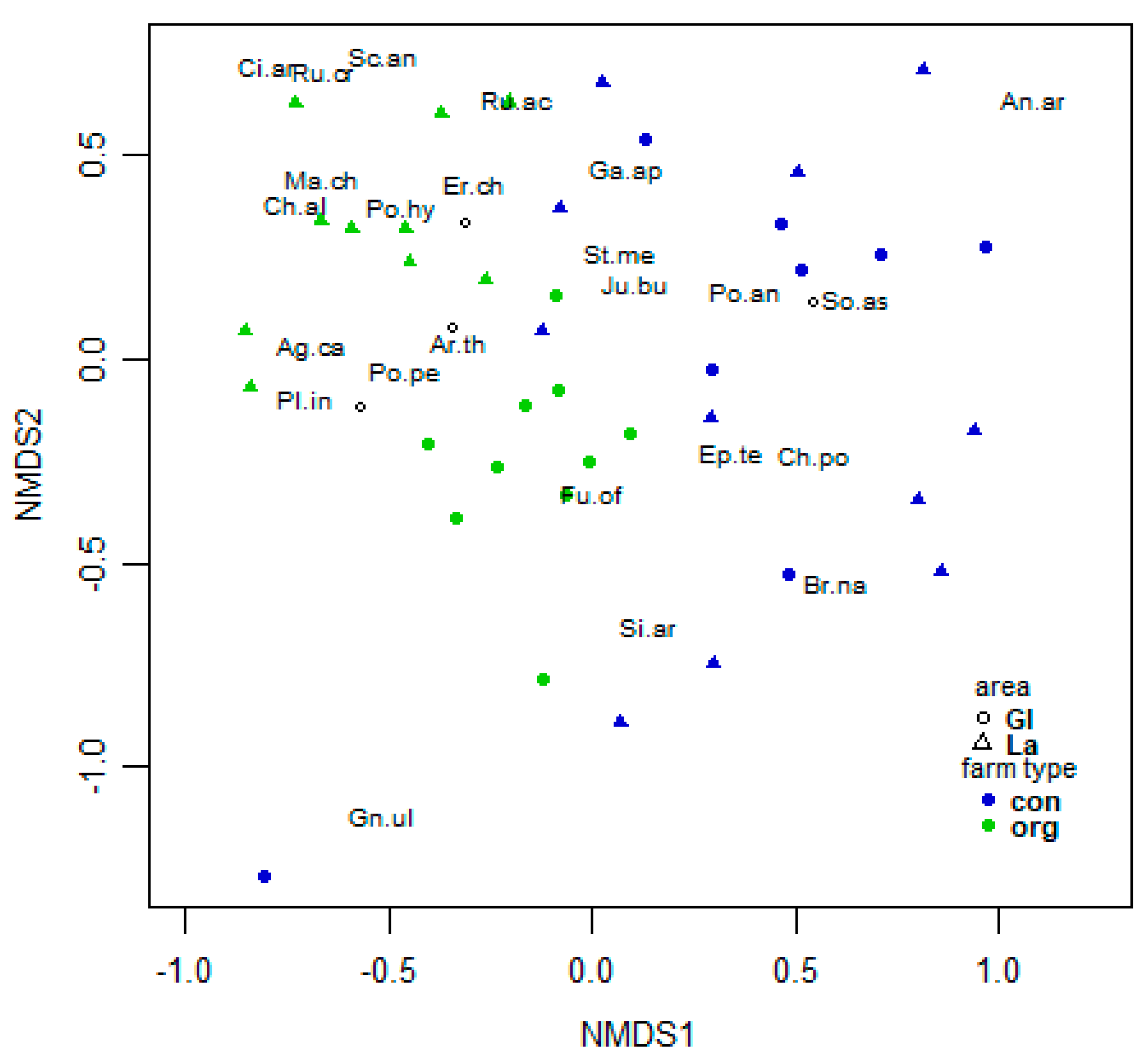
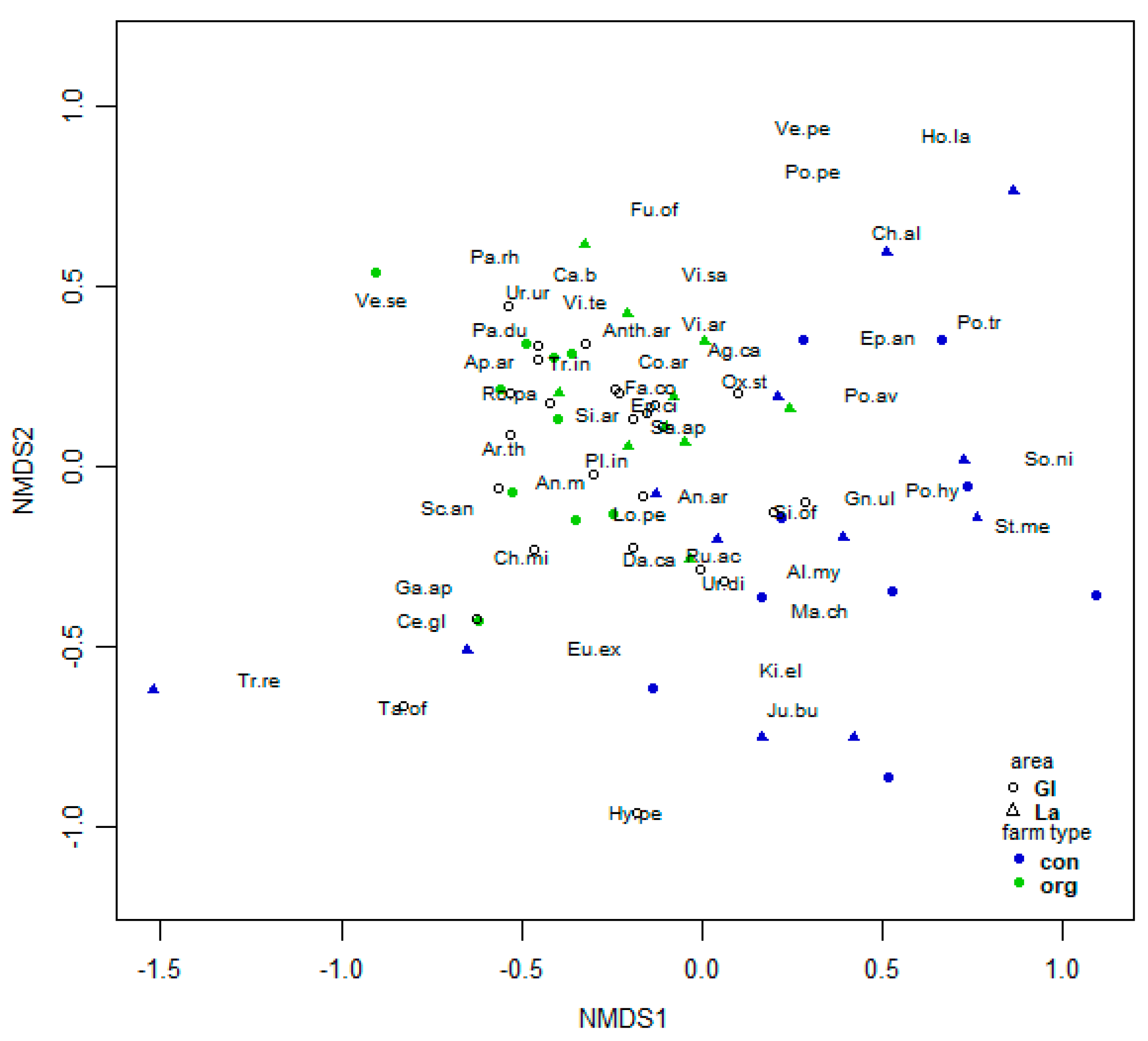
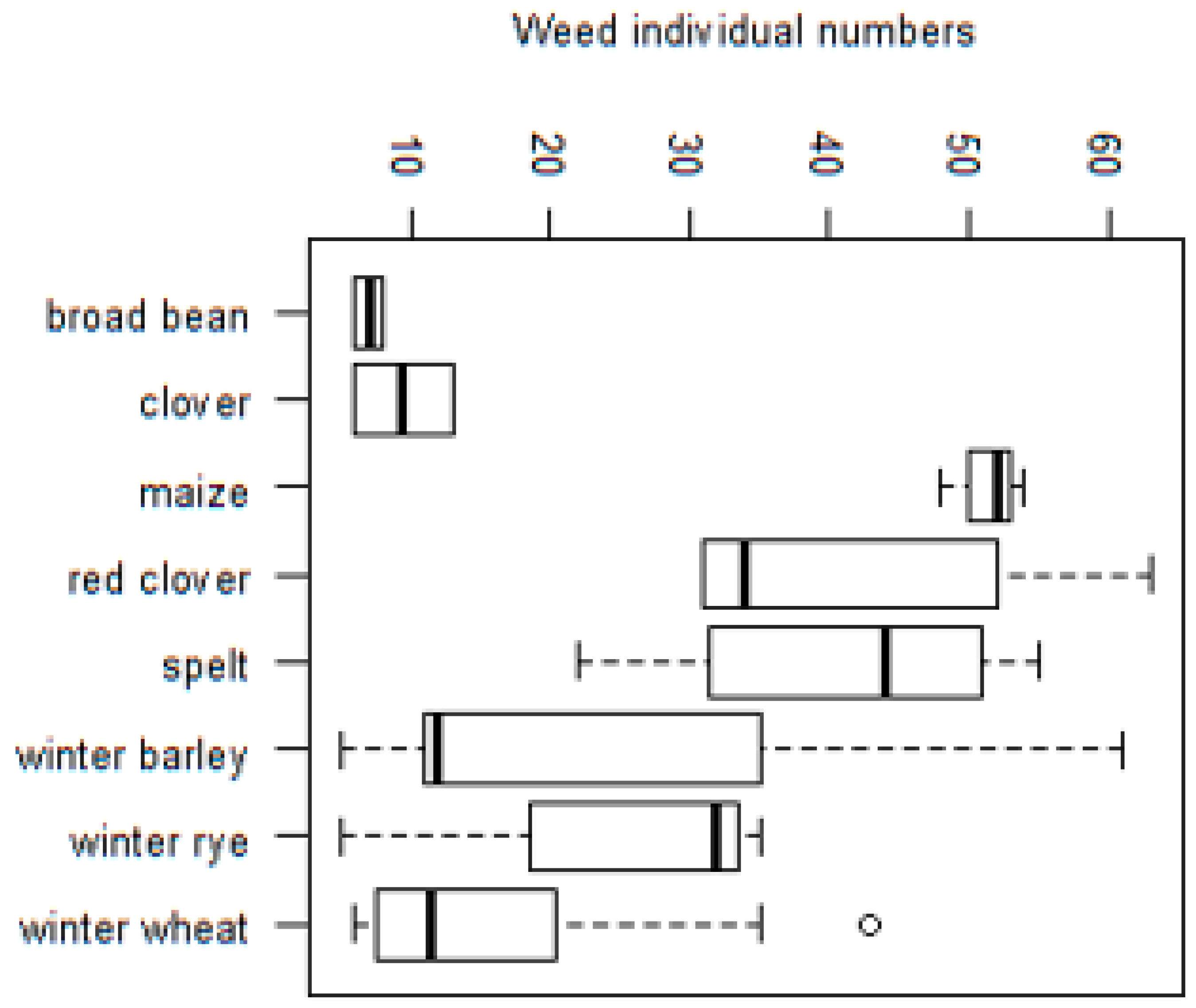
3. Results
3.1. Spontaneous Arable Weed Flora
3.2. Soil Seedbank of the Arable Weeds
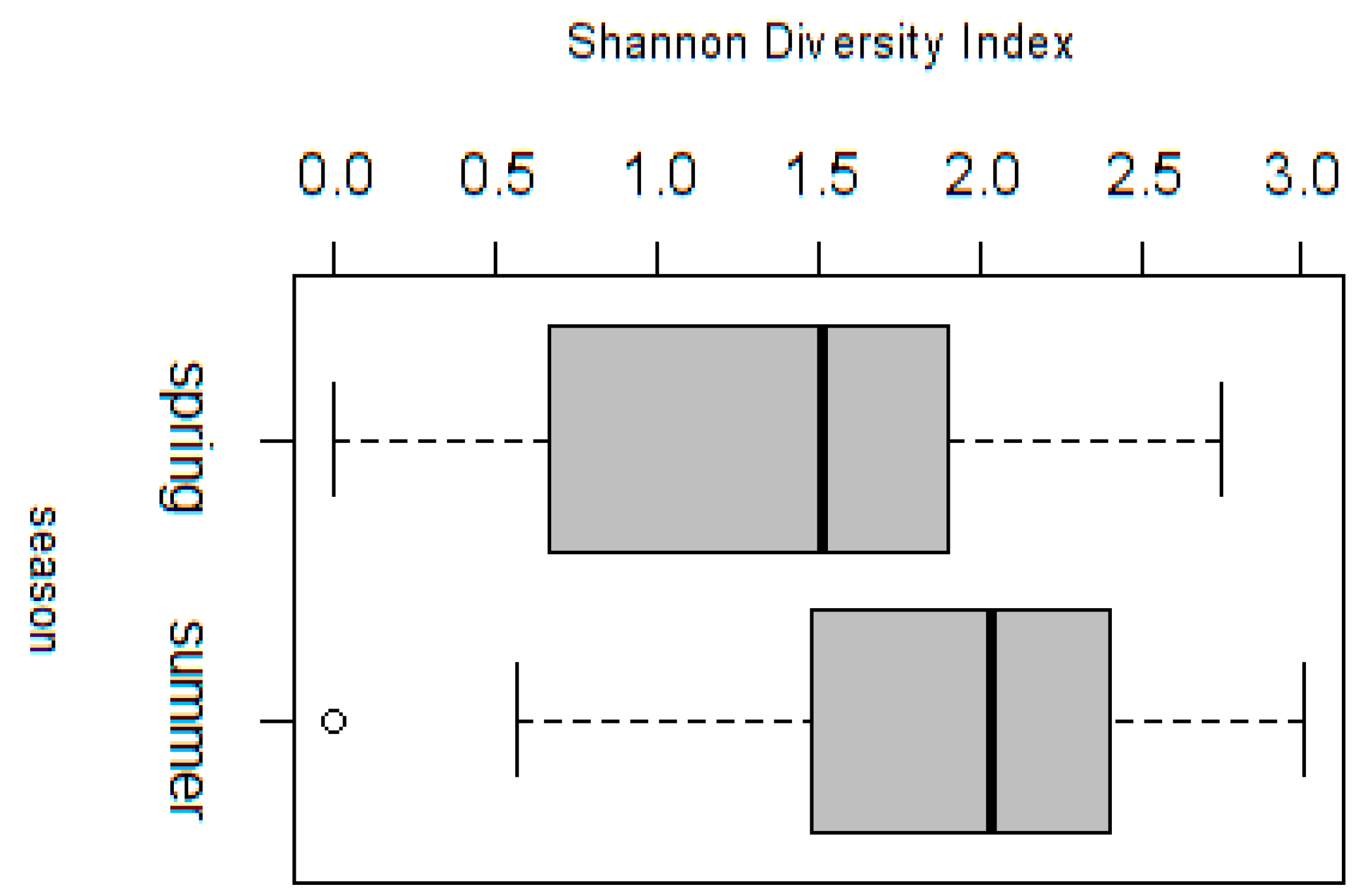
3.2.1. Analysis of Spring Vegetation
3.2.2. Indicator Species Analysis
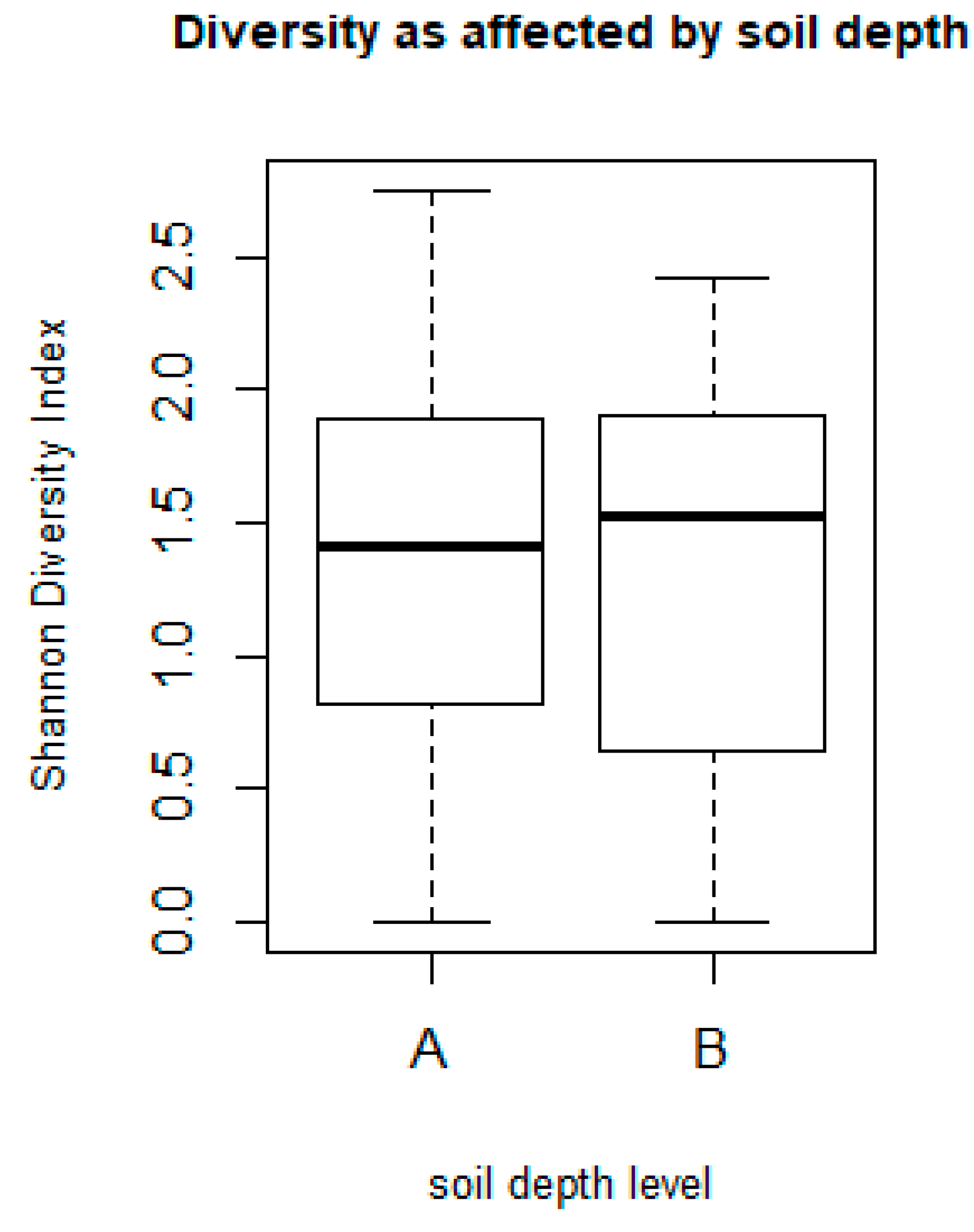
3.2.3. Impact of Crop Type on Species Richness
- A—the number of species in one of the two communities compared.
- B—the number of species in the second community compared.
- C—the number of common species in the compared communities.
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hyvönen, T.; Huusela-Veistola, E. Arable weeds as indicators of agricultural intensity–A case study from Finland. Biol. Conserv. 2008, 141, 2857–2864. [Google Scholar] [CrossRef]
- Giam, X.; Bradshaw, C.J.; Tan, H.T.; Sodhi, N.S. Future habitat loss and the conservation of plant biodiversity. Biol. Conserv. 2010, 143, 1594–1602. [Google Scholar] [CrossRef]
- Lemoine, C.; Sérusiaux, E.; Mahy, G.; Piqueray, J. Agro-environmental scheme for segetal plant conservation in Wallonia (Belgium): An assessment in conventional and organic fields. Biotechnol. Agron. Soc. Environ. 2018, 22, 35–44. [Google Scholar] [CrossRef]
- Chamorro, L.; Masalles, R.M.; Sans, F. Arable weed decline in Northeast Spain: Does organic farming recover functional biodiversity? Agric. Ecosyst. Environ. 2016, 223, 1–9. [Google Scholar] [CrossRef]
- Andreasen, C.; Jensen, H.A.; Jensen, S.M. Decreasing diversity in the soil seed bank after 50 years in Danish arable fields. Agric. Ecosyst. Environ. 2018, 259, 61–71. [Google Scholar] [CrossRef]
- Hyvönen, T. Can conversion to organic farming restore the species composition of arable weed communities? Biol. Conserv. 2007, 137, 382–390. [Google Scholar] [CrossRef]
- Albrecht, H. Development of arable weed seedbanks during the 6 years after the change from conventional to organic farming. Weed Res. 2005, 45, 339–350. [Google Scholar] [CrossRef]
- Carmona, C.P.; Guerrero, I.; Peco, B.; Morales, M.B.; Oñate, J.J.; Pärt, T.; Tscharntke, T.; Liira, J.; Aavik, T.; Emmerson, M.; et al. Agriculture intensification reduces plant taxonomic and functional diversity across European arable systems. Funct. Ecol. 2020, 34, 1448–1460. [Google Scholar] [CrossRef]
- Hawes, C.; Squire, G.R.; Hallett, P.D.; Watson, C.A.; Young, M. Arable plant communities as indicators of farming practice. Agric. Ecosyst. Environ. 2010, 138, 17–26. [Google Scholar] [CrossRef]
- Fonderflick, J.; Besnard, A.; Chardès, M.-C.; Lanuzel, L.; Thill, C.; Pointereau, P. Impacts of agricultural intensification on arable plants in extensive mixed crop-livestock systems. Agric. Ecosyst. Environ. 2020, 290, 106778. [Google Scholar] [CrossRef]
- Rotchés-Ribalta, R.; Blanco-Moreno, J.M.; Armengot, L.; Chamorro, L.; Sans, F.X. Both farming practices and landscape characteristics determine the diversity of characteristic and rare arable weeds in organically managed fields. Appl. Veg. Sci. 2015, 18, 423–431. [Google Scholar] [CrossRef]
- Gaba, S.; Perronne, R.; Fried, G.; Gardarin, A.; Bretagnolle, F.; Biju-Duval, L.; Colbach, N.; Cordeau, S.; Fernández-Aparicio, M.; Gauvrit, C.; et al. Response and effect traits of arable weeds in agro-ecosystems: A review of current knowledge. Weed Res. 2017, 57, 123–147. [Google Scholar] [CrossRef]
- Cavan, G.; Cussans, J.; Moss, S.R. Modelling different cultivation and herbicide strategies for their effect on herbicide resistance in Alopecurus myosuroides. Weed Res. 2000, 40, 561–568. [Google Scholar] [CrossRef]
- Menchari, Y.; Camilleri, C.; Michel, S.; Brunel, D.; Dessaint, F.; Le Corre, V.; Délye, C. Weed response to herbicides: Regional-scale distribution of herbicide resistance alleles in the grass weed Alopecurus myosuroides. New Phytol. 2006, 171, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Abdallah, I.; Han, H.; Owen, M.; Powles, S. Distinct non-target site mechanisms endow resistance to glyphosate, ACCase and ALS-inhibiting herbicides in multiple herbicide-resistant Lolium rigidum. Planta 2009, 230, 713–723. [Google Scholar] [CrossRef]
- Storkey, J.; Neve, P. What good is weed diversity? Weed Res. 2018, 58, 239–243. [Google Scholar] [CrossRef]
- Seifert, C.; Leuschner, C.; Culmsee, H. Arable plant diversity on conventional cropland—The role of crop species, management and environment. Agric. Ecosyst. Environ. 2015, 213, 151–163. [Google Scholar] [CrossRef]
- Albrecht, H.; Cambecèdes, J.; Lang, M.; Wagner, M. Management options for the conservation of rare arable plants in Europe. Bot. Lett. 2016, 163, 389–415. [Google Scholar] [CrossRef] [Green Version]
- Wood, T.J.; Holland, J.M.; Goulson, D. Providing foraging resources for solitary bees on farmland: Current schemes for pollinators benefit a limited suite of species. J. Appl. Ecol. 2017, 54, 323–333. [Google Scholar] [CrossRef]
- McGinlay, J.; Parsons, D.J.; Morris, J.; Hubatova, M.; Graves, A.; Bradbury, R.B.; Bullock, J.M. Do charismatic species groups generate more cultural ecosystem service benefits? Ecosyst. Serv. 2017, 27, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.M.; Aebischer, N.J.; Ewald, J.; Moreby, S.; Potter, C.; Holland, J.M. The Potential of Arable Weeds to Reverse Invertebrate Declines and Associated Ecosystem Services in Cereal Crops. Front. Sustain. Food Syst. 2020, 3, 118. [Google Scholar] [CrossRef]
- Kolářová, M.; Tyšer, L.; Soukup, J. Impact of site conditions and farming practices on the occurrence of rare and endangered weeds on arable land in the Czech Republic. Weed Res. 2013, 53, 489–498. [Google Scholar] [CrossRef]
- Rotchés-Ribalta, R.; Armengot, L.; Mäder, P.; Mayer, J.; Sans, F.X. Long-Term Management Affects the Community Composition of Arable Soil Seedbanks. Weed Sci. 2017, 65, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Waldhardt, R.; Fuhr-Boßdorf, K.; Otte, A.; Schmidt, J.; Simmering, D. Floristisch-phytocoenotische Diversitäten einer peripheren Kulturlandschaft in Abhängigkeit von Landnutzung. Raum und Zeit. Agrar. 2000, 31, 121–147. [Google Scholar]
- Price, J.N.; Wright, B.R.; Gross, C.L.; Whalley, W.R.D.B. Comparison of seedling emergence and seed extraction techniques for estimating the composition of soil seed banks: Comparison of soil seed bank methods. Methods Ecol. Evol. 2010, 1, 151–157. [Google Scholar] [CrossRef]
- Rahman, A.; James, T.K.; Mellsop, J.; Grbavac, N. Effect of cultivation methods on weed seed distribution and seedling emergence. New Zealand Plant Prot. 2000, 53, 28–33. [Google Scholar] [CrossRef]
- Ahammad, K.; Rahman, M.; Ali, M. Effect of hydropriming method on maize (Zea mays) seedling emergence. Bangladesh J. Agric. Res. 2014, 39, 143–150. [Google Scholar] [CrossRef]
- Kolářová, M.; Tyšer, L.; Soukup, J. Weed vegetation of arable land in the Czech Republic: Environmental a management factors determining weed species composition. Biologia 2014, 69, 443–448. [Google Scholar] [CrossRef]
- Hyvönen, T.; Ketoja, E.; Salonen, J.; Jalli, H.; Tiainen, J. Weed species diversity and community composition in organic and conventional cropping of spring cereals. Agric. Ecosyst. Environ. 2003, 97, 131–149. [Google Scholar] [CrossRef]
- Feledyn-Szewczyk, B.; Smagacz, J.; Kwiatkowski, C.A.; Harasim, E.; Woźniak, A. Weed Flora and Soil Seed Bank Composition as Affected by Tillage System in Three-Year Crop Rotation. Agriculture 2020, 10, 186. [Google Scholar] [CrossRef]
- Berbeć, A.K.; Staniak, M.; Feledyn-Szewczyk, B.; Kocira, A.; Stalenga, J. Organic but Also Low-Input Conventional Farming Systems Support High Biodiversity of Weed Species in Winter Cereals. Agriculture 2020, 10, 413. [Google Scholar] [CrossRef]
- van Elsen, T. Species diversity as a task for organic agriculture in Europe. Agric. Ecosyst. Environ. 2000, 77, 101–109. [Google Scholar] [CrossRef]
- Chamorro, L.; Armengot, L.; José-María, L.; Sans, F.X. Organic farming enhances the recovery of ancient crops and segetal weeds in Catalonia (NE of Spain). Build. Org. Bridges 2014, 3, 979–982. [Google Scholar]
- Mahé, I.; Cordeau, S.; Bohan, D.A.; Derrouch, D.; Dessaint, F.; Millot, D.; Chauvel, B. Soil seedbank: Old methods for new challenges in agroecology? Ann. Appl. Biol. 2021, 178, 23–38. [Google Scholar] [CrossRef]
- Schwartz-Lazaro, L.M.; Copes, J.T. A Review of the Soil Seedbank from a Weed Scientists Perspective. Agronomy 2019, 9, 369. [Google Scholar] [CrossRef] [Green Version]
- Lososová, Z.; Chytrý, M.; Cimalová, S.; Kropáč, Z.; Otýpková, Z.; Pyšek, P.; Tichý, L. Weed vegetation of arable land in Central Europe: Gradients of diversity and species composition. J. Veg. Sci. 2004, 15, 415–422. [Google Scholar] [CrossRef]
- Mennan, H.; Ngouajio, M. Seasonal cycles in germination and seedling emergence of summer and winter populations of catchweed bedstraw (Galium aparine) and wild mustard (Brassica kaber). Weed Sci. 2006, 54, 114–120. [Google Scholar] [CrossRef]
- Swanton, C.J.; Shrestha, A.; Knezevic, S.Z.; Roy, R.C.; Ball-Coelho, B.R. Influence of tillage type on vertical weed seedbank distribution in a sandy soil. Can. J. Plant Sci. 2000, 80, 455–457. [Google Scholar] [CrossRef]
- Schmitzberger, I.; Wrbka, T.; Steurer, B.; Aschenbrenner, G.; Peterseil, J.; Zechmeister, H.G. How farming styles influence biodiversity maintenance in Austrian agricultural landscapes. Agric. Ecosyst. Environ. 2005, 108, 274–290. [Google Scholar] [CrossRef]
- Clements, D.R.; Benoit, D.L.; Murphy, S.D.; Swanton, C.J. Tillage Effects on Weed Seed Return and Seedbank Composition. Weed Sci. 1996, 44, 314–322. [Google Scholar] [CrossRef]
- Buhler, D.D.; Kohler, K.A.; Thompson, R.L. Weed Seed Bank Dynamics During a Five-Year Crop Rotation. Weed Technol. 2001, 15, 170–176. [Google Scholar] [CrossRef]
- Armengot, L.; José-María, L.; Blanco-Moreno, J.M.; Bassa, M.; Chamorro, L.; Sans, F.X. A novel index of land use intensity for organic and conventional farming of Mediterranean cereal fields. Agron. Sustain. Dev. 2011, 31, 699–707. [Google Scholar] [CrossRef] [Green Version]
- Waldhardt, R.; Otte, A. Abschätzung der zur Erhaltung einer lokaltypischen Ackerflora benötigten Fläche am Beispiel der Gemarkung Erda (Lahn-Dill-Bergland, Hessen). Peckiana 2001, 1, 101–108. [Google Scholar]
- Chytrý, M.; Schaminée, J.H.J.; Schwabe, A. Vegetation survey: A new focus for Applied Vegetation Science: Vegetation survey. Appl. Veg. Sci. 2011, 14, 435–439. [Google Scholar] [CrossRef]
- Kuht, J.; Eremeev, V.; Talgre, L.; Madsen, H.; Toom, M.; Mäeorg, E. Soil weed seed bank and factors influencing the number of weeds at the end of conversion period to organic production. Agron. Res. 2016, 14, 1372–1379. [Google Scholar]
- Oksanen, J. Multivariate Analysis of Ecological Communities in R: Vegan tutorial. R Package Version 2011, 1, 1–43. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimate | Std.Error | z | Pr(>|z|) | Significance | |
|---|---|---|---|---|---|
| (Intercept) | 7.161 | 0.002555 | 2802.65 | 2e × 10−16 | *** |
| area.La | −1.252 | 0.004270 | −293.20 | 2e × 10−16 | *** |
| man.org | 0.944 | 0.002259 | 417.83 | 2e × 10−16 | *** |
| sd.2 | 1.031 | 0.002249 | 458.35 | 2e × 10−16 | *** |
| area.La × man.org | 1.094 | 0.003770 | 290.25 | 2e × 10−16 | *** |
| area.La × sd.2 | 0.213 | 0.003399 | 62.56 | 2e × 10−16 | *** |
| Estimate | Std.Error | t-Value | Pr(>|t|) | Significance | |
|---|---|---|---|---|---|
| Intercept | 10 | 3.383.923 | 3.046 | 0.00285 | * |
| year | −5.097 | 1.675 | −3.043 | 0.00288 | * |
| area | 7.008 | 3.808 | 1.840 | 0.06819 | n.s. |
| management | 24.024 | 3.974 | 6.045 | 1.73e-08 | * |
| Estimate | Std.Error | z | Pr(>|z|) | Significance | |
|---|---|---|---|---|---|
| (Intercept) | 7.161 | 0.002555 | 2802.65 | 2e × 10−16 | *** |
| area.La | −1.252 | 0.004270 | −293.20 | 2e × 10−16 | *** |
| man.org | 0.944 | 0.002259 | 417.83 | 2e × 10−16 | *** |
| sd.2 | 1.031 | 0.002249 | 458.35 | 2e × 10−16 | *** |
| area.La × man.org | 1.094 | 0.003770 | 290.25 | 2e × 10−16 | *** |
| area.La × sd.2 | 0.213 | 0.003399 | 62.56 | 2e × 10−16 | *** |
| 14 Years | 18 Years | 27 Years | |
|---|---|---|---|
| 14 years | - | 0.035 * | - |
| 27 years | 0.025 * | 0.403 | - |
| 38 years | 0.403 | 0.402 | 0.207 |
| Group nr. | A | B | Stat | p.Value | ||
|---|---|---|---|---|---|---|
| 1 | Pap.rho | 0.9732 | 1.0 | 0.987 | 0.001 | *** |
| Cen.cya. | 0.9784 | 0.8333 | 0.903 | 0.001 | *** | |
| Ver.arv. | 0.6816 | 1.0 | 0.826 | 0.001 | *** | |
| Fal.con. | 0.8128 | 0.5000 | 0.638 | 0.001 | *** | |
| Vio.arv. | 0.4722 | 0.6667 | 0.561 | 0.004 | ** | |
| Dac.glo. | 0.6598 | 0.3333 | 0.469 | 0.013 | * | |
| Gal.apa. | 0.9010 | 0.1667 | 0.388 | 0.011 | * | |
| Sis.off. | 0.8987 | 0.1667 | 0.387 | 0.039 | * | |
| Pap.dub. | 0.8812 | 0.1667 | 0.383 | 0.025 | * | |
| 2 | Sag.pr. | 0.87429 | 0.24490 | 0.463 | 0.032 | * |
| Rum.obt. | 100.000 | 0.06122 | 0.247 | 0.041 | * | |
| 3 | Tri.pra. | 0.91054 | 0.43478 | 0.629 | 0.001 | *** |
| Alop.myo. | 0.75716 | 0.26087 | 0.444 | 0.033 | * | |
| Laps.comm. | 100.000 | 0.08696 | 0.295 | 0.025 | * | |
| 4 | Matr.cham. | 0.8640 | 0.9636 | 0.912 | 0.001 | *** |
| Cha.mi. | 0.9445 | 0.1455 | 0.371 | 0.029 | * |
| Conventional | Gl.Organic | |
|---|---|---|
| organic | 57.3 | - |
| La.organic | - | 65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Köllmann, P.; Waldhardt, R. Farming Intensity Affects Soil Seedbank Composition and Spontaneous Vegetation of Arable Weeds. Diversity 2022, 14, 111. https://doi.org/10.3390/d14020111
Köllmann P, Waldhardt R. Farming Intensity Affects Soil Seedbank Composition and Spontaneous Vegetation of Arable Weeds. Diversity. 2022; 14(2):111. https://doi.org/10.3390/d14020111
Chicago/Turabian StyleKöllmann, Philipp, and Rainer Waldhardt. 2022. "Farming Intensity Affects Soil Seedbank Composition and Spontaneous Vegetation of Arable Weeds" Diversity 14, no. 2: 111. https://doi.org/10.3390/d14020111
APA StyleKöllmann, P., & Waldhardt, R. (2022). Farming Intensity Affects Soil Seedbank Composition and Spontaneous Vegetation of Arable Weeds. Diversity, 14(2), 111. https://doi.org/10.3390/d14020111