
Recruitment Patterns and Potential Climate Change Impacts on Three Florida Hylids with Different Life Histories

Abstract
:1. Introduction
2. Materials and Methods
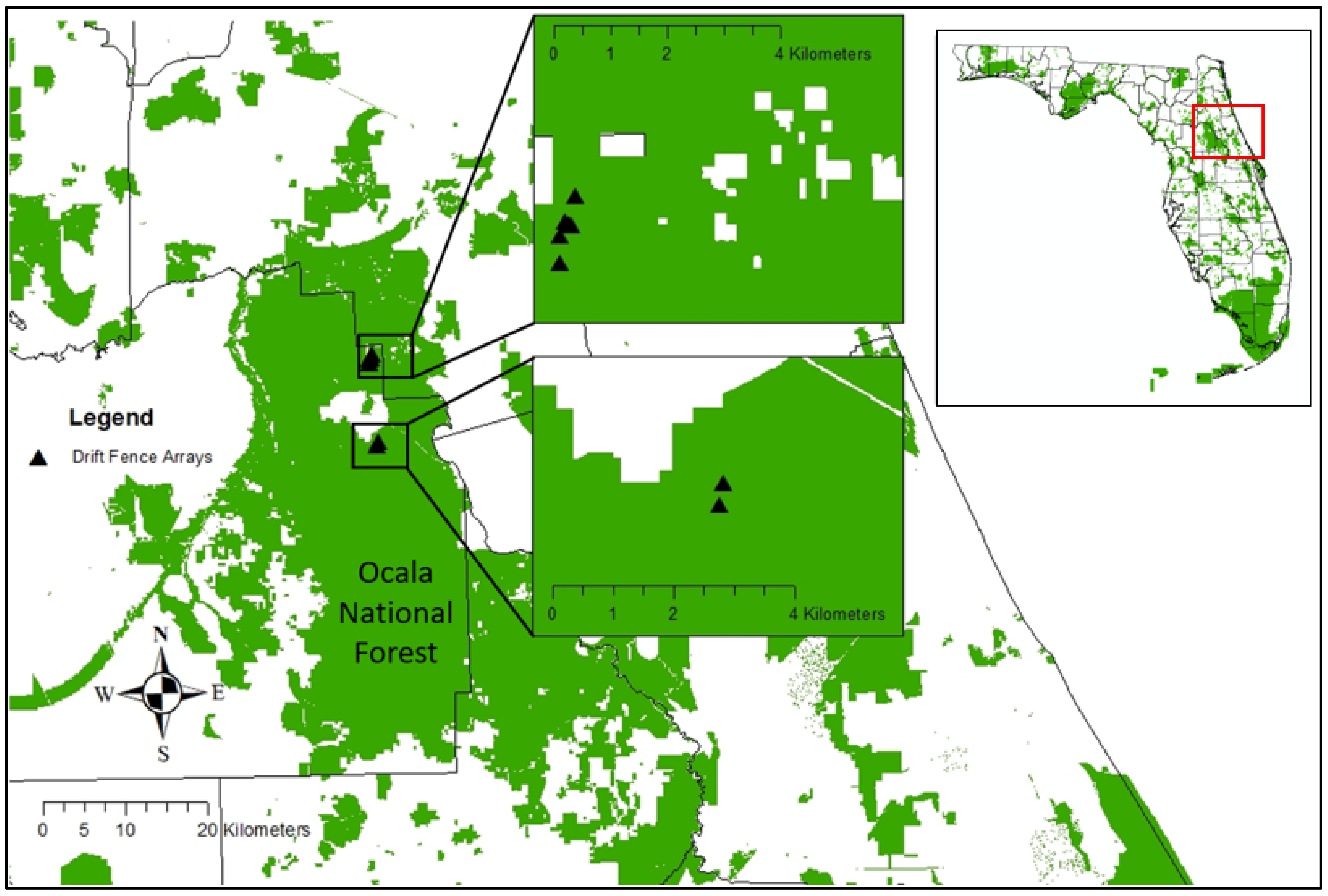
2.1. Study Area
2.2. Field Methods
2.3. Determining Breeding-to-Metamorphosis Periods (BMPs)
2.4. Data Processing
2.5. Data Analysis
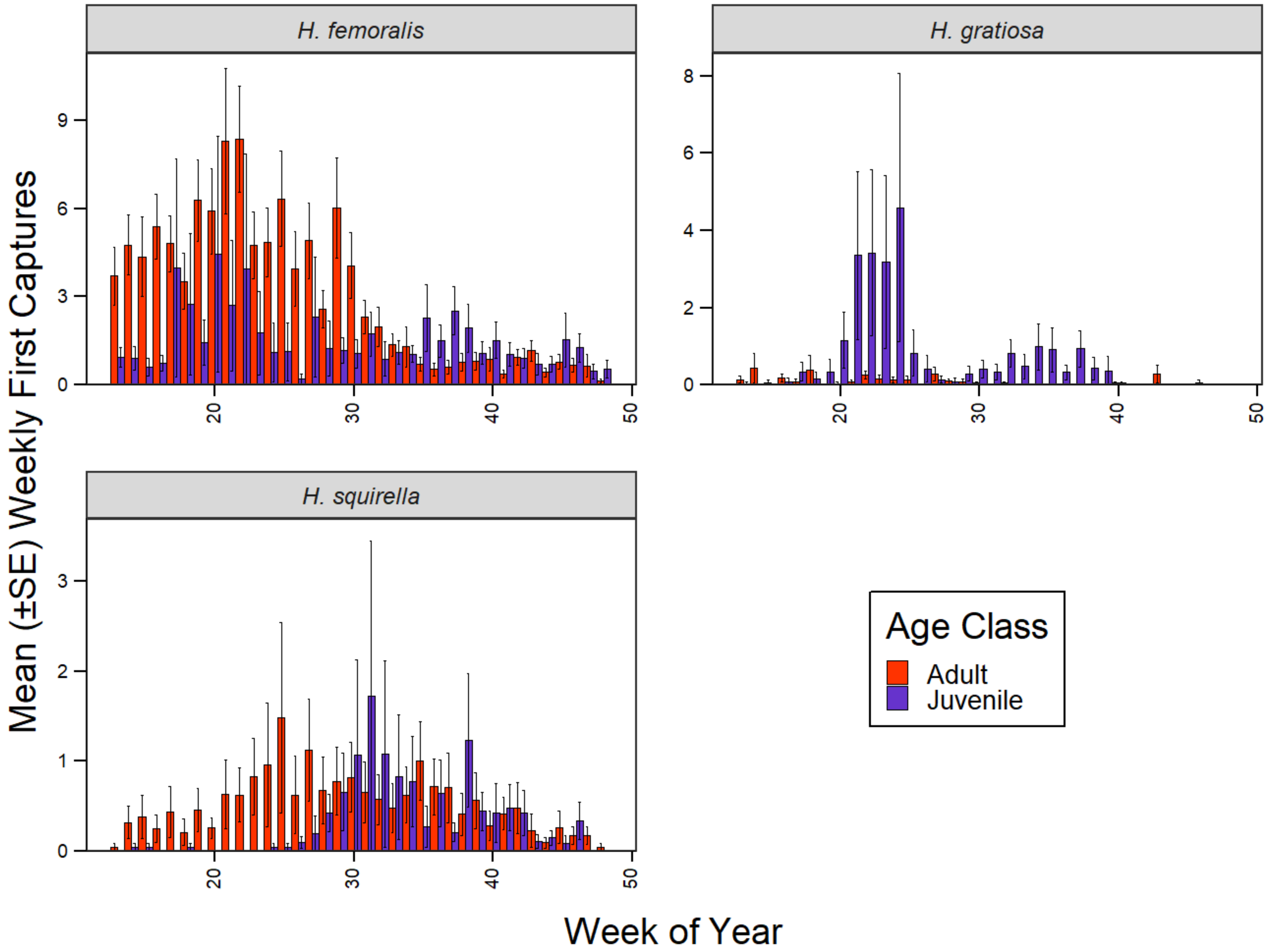
2.5.1. Phase I: Evaluating Patterns of Annual Recruitment
2.5.2. Phase II: Evaluating the Relationship between Bioclim and Recruitment-Related Variables
2.5.3. Phase III: Projecting Future Recruitment under Different Climate Change Scenarios
2.5.4. Decadal Capture Trends
3. Results
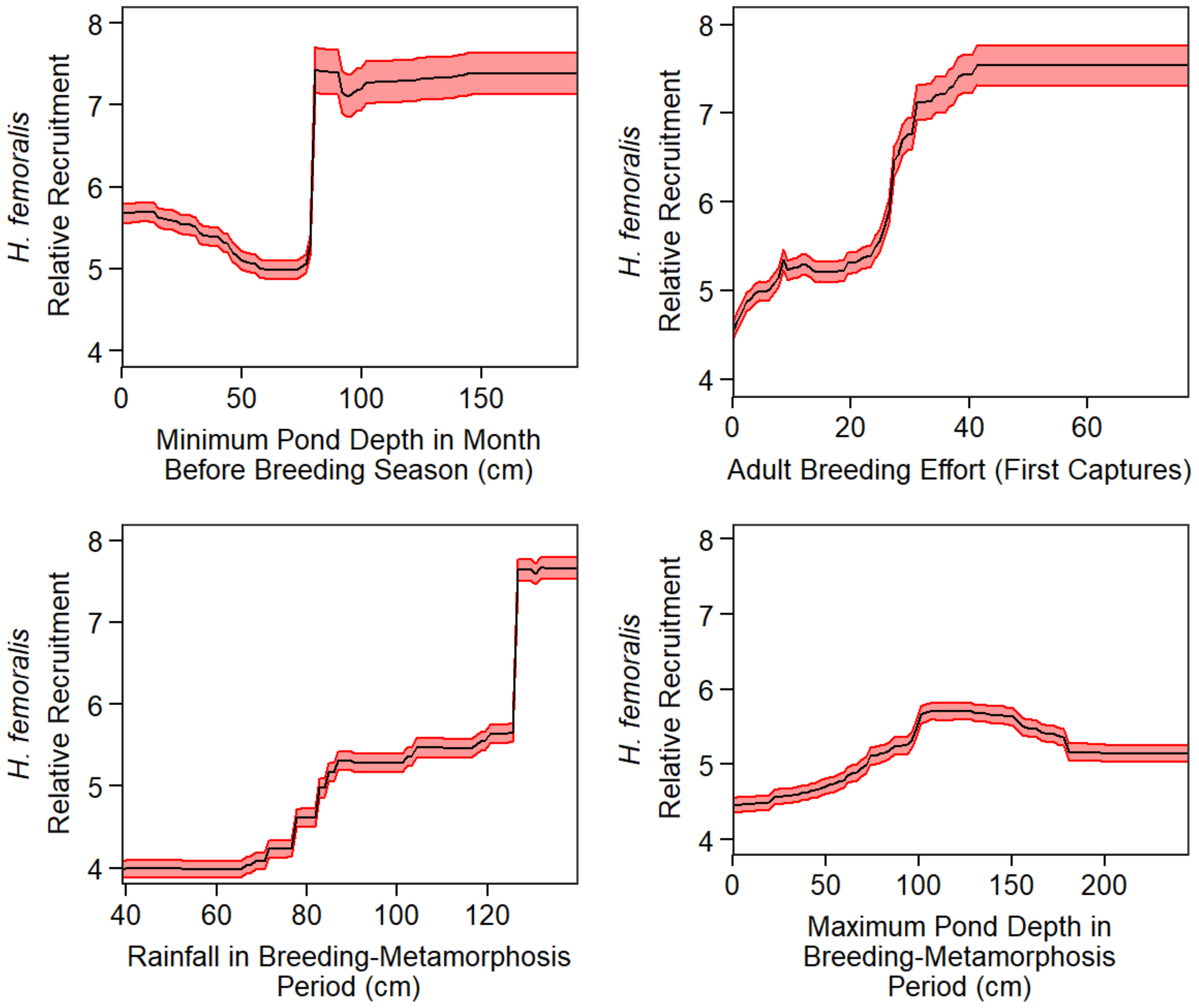
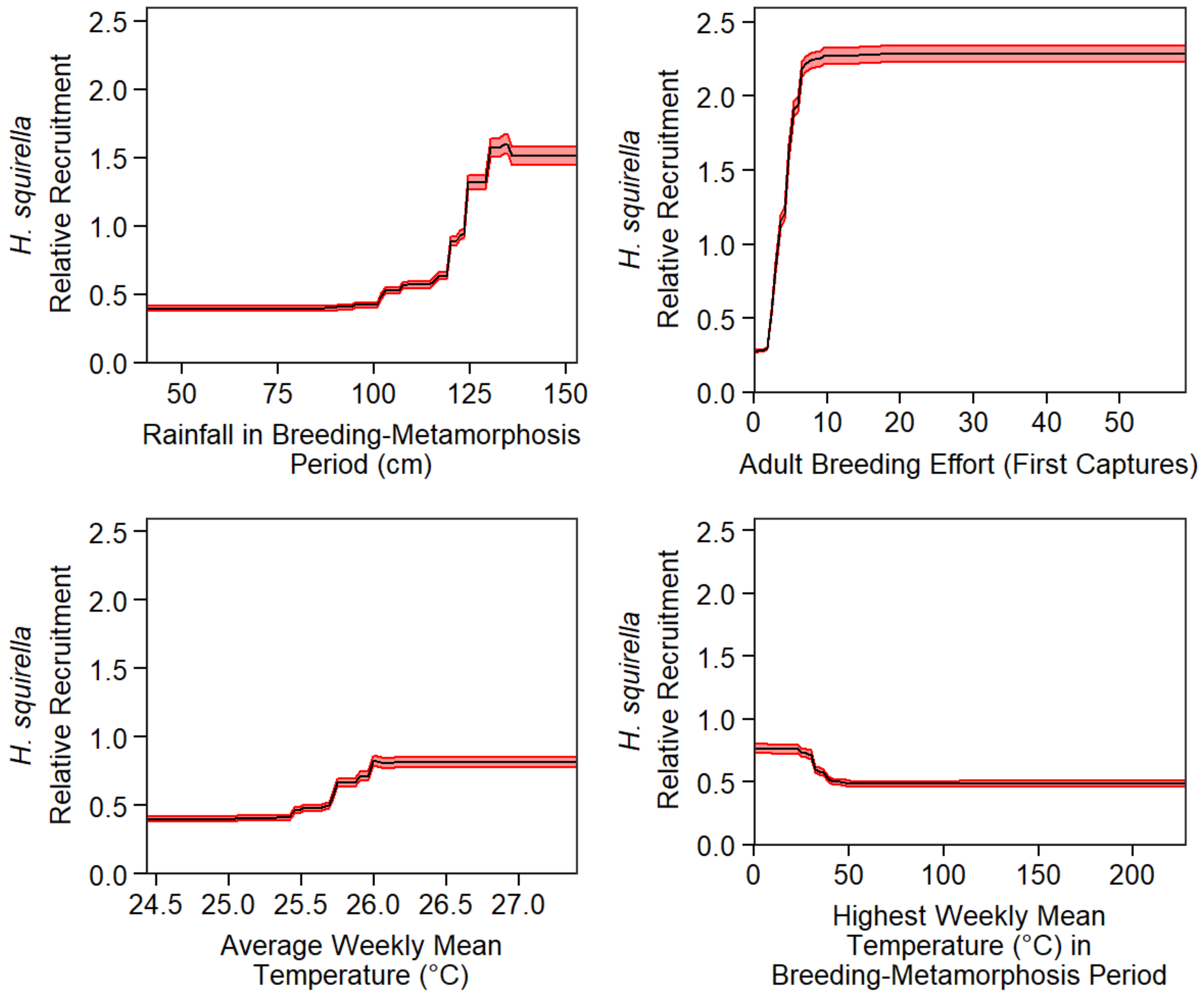
3.1. Phase I: Influence of Pond-Level Conditions on Annual Recruitment
3.2. Phase II: Predicting Pond-Level Variables Using Bioclimatic Data
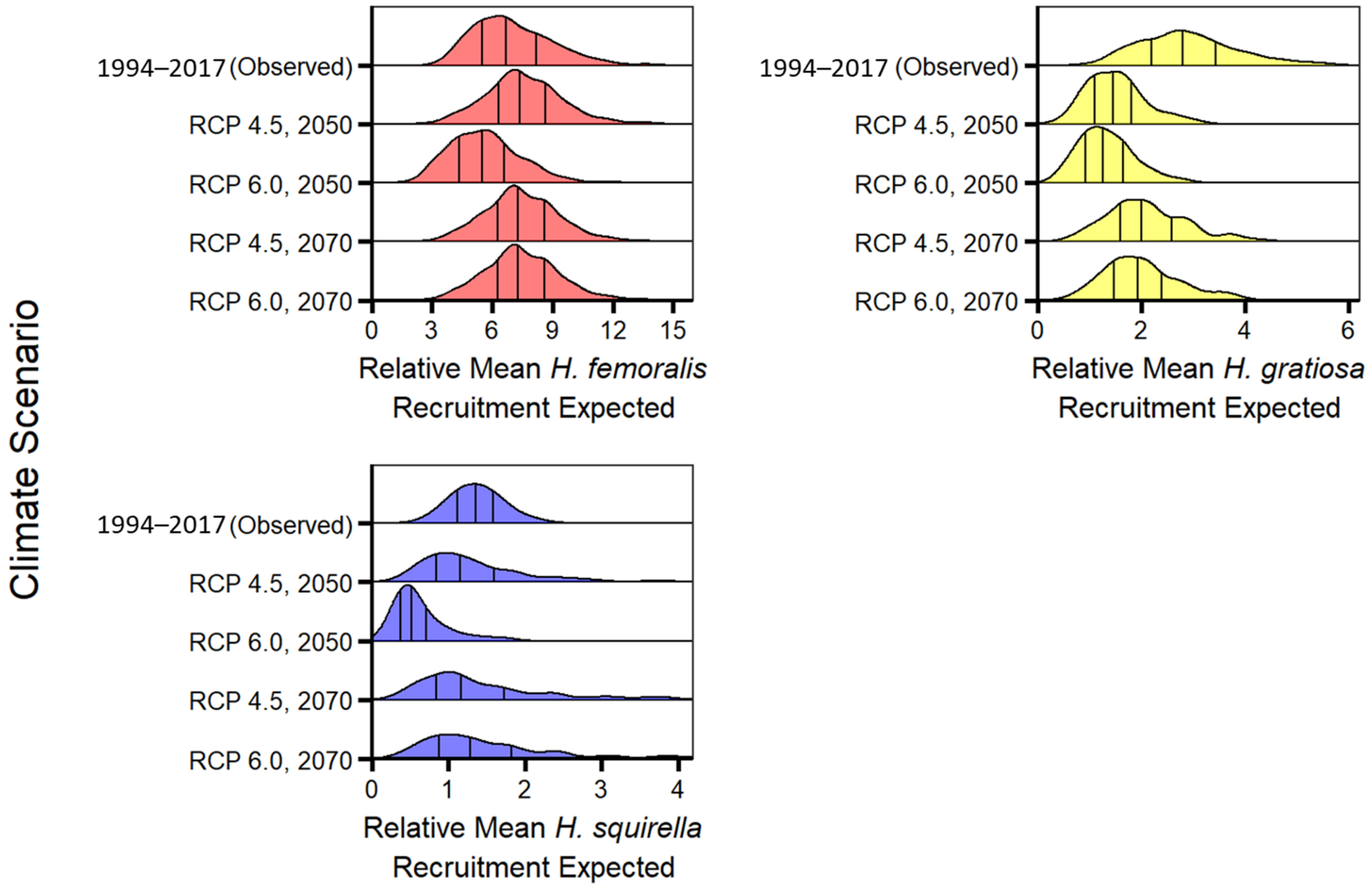
3.3. Phase III: Predicting Changes in Recruitment under Different Climate Change Scenarios
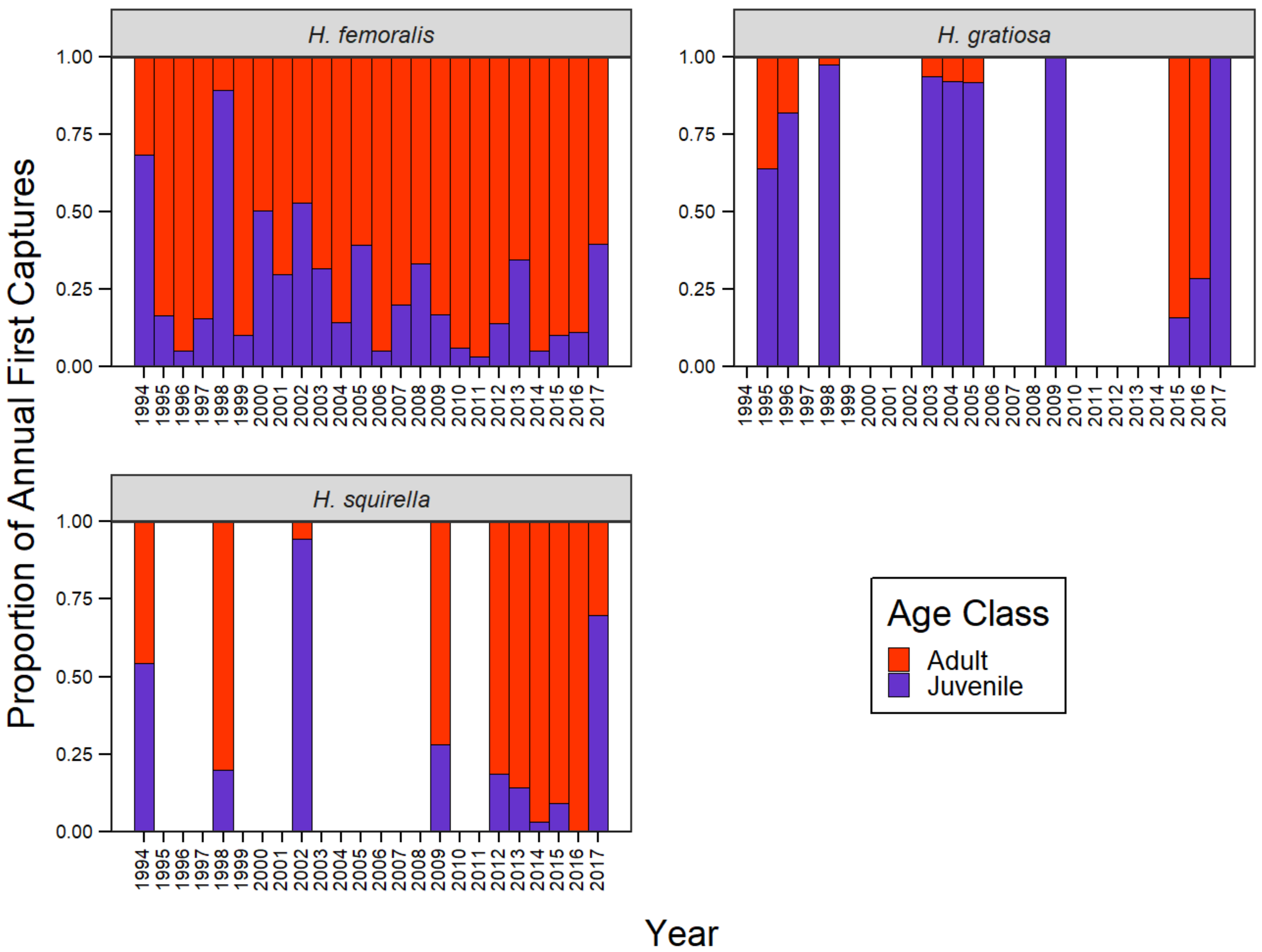
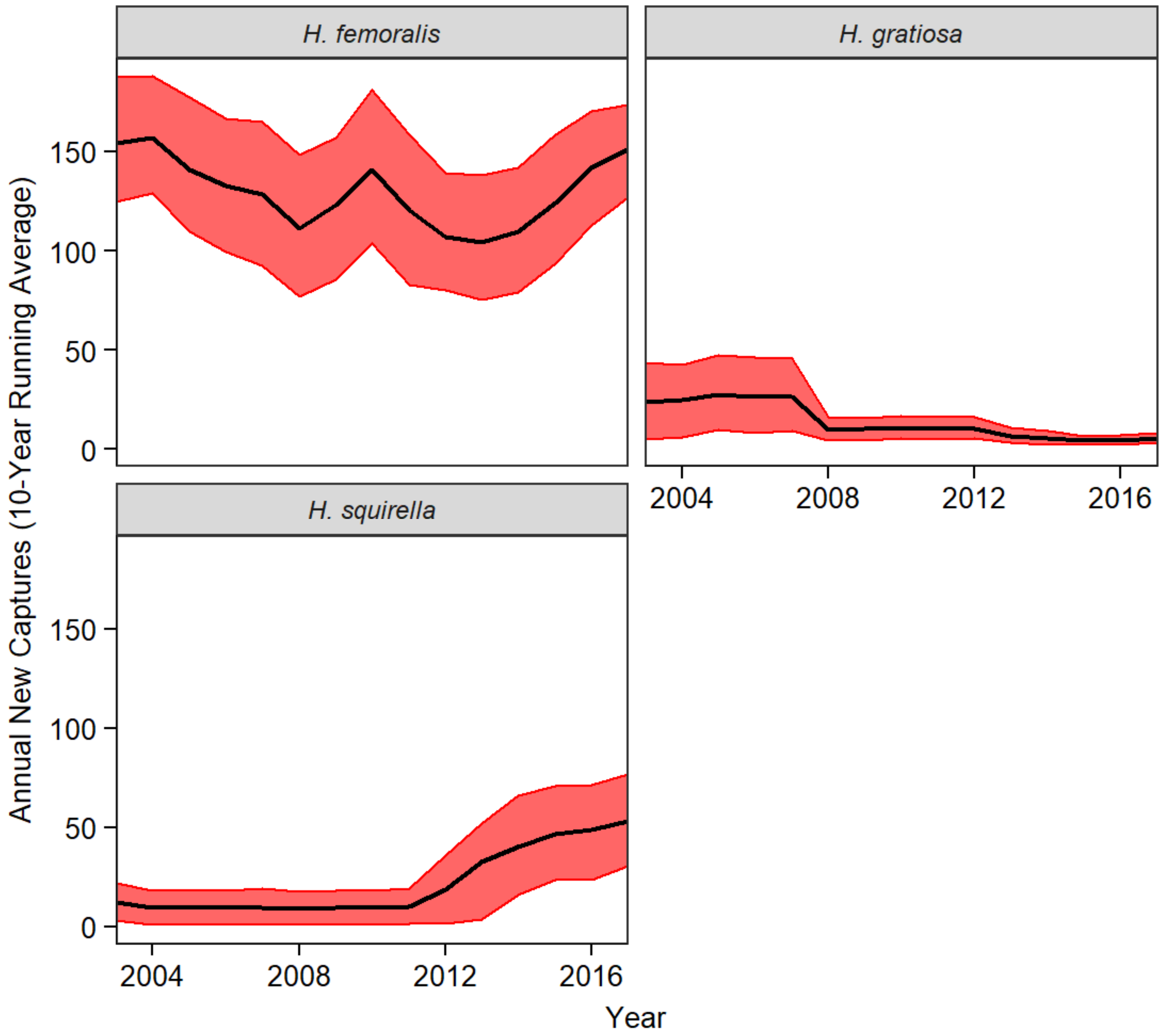
3.4. Decadal Capture Trends
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fetene, A. Aspects of climate change and its associated impacts on wetland ecosystem functions—A review. J. Am. Sci. 2012, 8, 582–596. [Google Scholar]
- Greenberg, C.H.; Goodrick, S.; Austin, J.; Parresol, B.R. Hydroregime Prediction Models for Ephemeral Groundwater-Driven Sinkhole Wetlands: A Planning Tool for Climate Change and Amphibian Conservation. Wetlands 2015, 35, 899–911. [Google Scholar] [CrossRef]
- Chandler, H.C.; McLaughlin, D.L.; Gorman, T.A.; McGuire, K.J.; Feaga, J.B.; Haas, C.A. Drying Rates of Ephemeral Wetlands: Implications for Breeding Amphibians. Wetlands 2017, 37, 545–557. [Google Scholar] [CrossRef]
- Walls, S.C.; Barichivich, W.J.; Brown, M.E. Drought, Deluge and Declines: The Impact of Precipitation Extremes on Amphibians in a Changing Climate. Biology 2013, 2, 399–418. [Google Scholar] [CrossRef] [Green Version]
- Todd, B.D.; Scott, D.; Pechmann, J.H.K.; Gibbons, J.W. Climate change correlates with rapid delays and advancements in reproductive timing in an amphibian community. Proc. R. Soc. B Boil. Sci. 2010, 278, 2191–2197. [Google Scholar] [CrossRef] [Green Version]
- McMenamin, S.K.; Hadly, E.A.; Wright, C.K. Climatic change and wetland desiccation cause amphibian decline in Yellowstone National Park. Proc. Natl. Acad. Sci. USA 2008, 105, 16988–16993. [Google Scholar] [CrossRef] [Green Version]
- Lowe, K.; Castley, J.G.; Hero, J.-M. Resilience to climate change: Complex relationships among wetland hydroperiod, larval amphibians and aquatic predators in temporary wetlands. Mar. Freshw. Res. 2015, 66, 886–899. [Google Scholar] [CrossRef] [Green Version]
- Pinceel, T.; Vanschoenwinkel, B.; Brendonck, L.; Buschke, F. Modelling the sensitivity of life history traits to climate change in a temporary pool crustacean. Sci. Rep. 2016, 6, 29451. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, C.H.; Zarnoch, S.J.; Austin, J.D. Weather, hydroregime, and breeding effort influence juvenile recruitment of anurans: Implications for climate change. Ecosphere 2017, 8, e01789. [Google Scholar] [CrossRef] [Green Version]
- Beck, C. Effect of changes in resource level on age and size at metamorphosis in Hyla squirella. Oecologia 1997, 112, 187–192. [Google Scholar] [CrossRef]
- Murphy, C.G. Chorus tenure of male barking treefrogs, Hyla gratiosa. Anim. Behav. 1994, 48, 763–777. [Google Scholar] [CrossRef] [Green Version]
- Burkett, V.; Kusler, J. Climate Change: Potential Impacts and Interactions in Wetlands of the United States. J. Am. Water Resour. Assoc. 2000, 36, 313–320. [Google Scholar] [CrossRef]
- Erwin, K.L. Wetlands and global climate change: The role of wetland restoration in a changing world. Wetl. Ecol. Manag. 2008, 17, 71–84. [Google Scholar] [CrossRef]
- Scott, D.E.; Metts, B.S. Shifts in the Amphibian Community over 30 Years at an Isolated Wetland: Has Climate Change Altered Wetland Hydrology? Georgia Institute of Technology: Atlanta, GA, USA, 2011. [Google Scholar]
- Semlitsch, R.D.; Bodie, J.R. Are small, isolated wetlands expendable? Conserv. Biol. 1998, 12, 1129–1133. [Google Scholar] [CrossRef] [Green Version]
- Semlitsch, R.D. Differentiating Migration and Dispersal Processes for Pond-Breeding Amphibians. J. Wildl. Manag. 2008, 72, 260–267. [Google Scholar] [CrossRef]
- Delis, P.R. Hyla gratiosa and H. femoralis (Anura: Hylidae) in west central Florida: A comparative study of rarity and commonness. In Biology; University of South Florida: Tampa, FL, USA, 2002. [Google Scholar]
- Homan, R.N.; Windmiller, B.S.; Reed, J.M. Critical thresholds associated with habitat loss for two vernal pool-breeding amphibians. Ecol. Appl. 2004, 14, 1547–1553. [Google Scholar] [CrossRef]
- Hether, T.D.; Hoffman, E.A. Machine learning identifies specific habitats associated with genetic connectivity in Hyla squirella. J. Evol. Biol. 2012, 25, 1039–1052. [Google Scholar] [CrossRef]
- Knowles, L., Jr.; O’Reilly, A.M.; Adamski, J.C. Hydrogeology and simulated effects of the ground-water withdrawals from the Floridan aquifer system in lake county and in the Ocala National Forest and vicinity, north-central Florida. Water-Resour. Investig. Rep. 2002, 2, 4207. [Google Scholar]
- Lee, S.; Yeo, I.-Y.; Lang, M.; Sadeghi, A.; McCarty, G.; Moglen, G.; Evenson, G. Assessing the cumulative impacts of geographically isolated wetlands on watershed hydrology using the SWAT model coupled with improved wetland modules. J. Environ. Manag. 2018, 223, 37–48. [Google Scholar] [CrossRef]
- Winter, T.C. The vulnerability of wetlands to climate change: A hydrologic landscape perspective. JAWRA J. Am. Water Resour. Assoc. 2000, 36, 305–311. [Google Scholar] [CrossRef]
- McCoy, E.D.; Delis, P.R.; Mushinsky, H.R. The importance of determining species sensitivity to environmental change: A tree frog example. Ecosphere 2021, 12, e03526. [Google Scholar] [CrossRef]
- Krysko, K.L.; Enge, K.M.; Moler, P.E. Amphibians and Reptiles of Florida; University of Florida Press: Gainesville, FL, USA, 2019; p. 728. [Google Scholar]
- Busby, J.R. BIOCLIM—A bioclimate analysis and prediction system. Plant Prot. Q. 1991, 61, 8–9. [Google Scholar]
- Winsberg, M.D. Florida Weather; University Press of Florida: Gainesville, FL, USA, 2003. [Google Scholar]
- Button, S.T.; Sovie, A.R.; Greenberg, C.H.; Austin, J. Evaluating the Ecology of Tantilla relicta in Florida Pine–Wiregrass Sandhills Using Multi-Season Occupancy Models. S. Am. J. Herpetol. 2019, 53, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, C.; Johnson, S.; Owen, R.; Storfer, A. Amphibian breeding phenology and reproductive outcome: An examination using terrestrial and aquatic sampling. Can. J. Zoöl. 2017, 95, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Stekhoven, D.J.; Bühlmann, P. MissForest—Non-parametric missing value imputation for mixed-type data. Bioinformatics 2012, 28, 112–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Einem, G.E.; Ober, L.D. The seasonal behavior of certain Floridian Salientia. Herpetologica 1956, 12, 205–212. [Google Scholar]
- Murphy, C.G. Determinants of chorus tenure in barking treefrogs (Hyla gratiosa). Behav. Ecol. Sociobiol. 1994, 34, 285–294. [Google Scholar] [CrossRef]
- Dray, S.; Legendre, P.; Neto, P.P. Spatial modelling: A comprehensive framework for principal coordinate analysis of neighbour matrices (PCNM). Ecol. Model. 2006, 196, 483–493. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef]
- Kohavi, R. A study of cross-validation and bootstrap for accuracy estimation and model selection. In Proceedings of the International Joint Conference on Artificial Intelligence, Montreal, QC, Canada, 20–25 August 1995. [Google Scholar]
- National Weather Service. Climate Data Online; N.O.A.A. Administration: Washington, DC, USA, 2020.
- Zhao, P.; Yu, B. On model selection consistency of Lasso. J. Mach. Learn. Res. 2006, 7, 2541–2563. [Google Scholar]
- Wong, T.-T. Performance evaluation of classification algorithms by k-fold and leave-one-out cross validation. Pattern Recognit. 2015, 48, 2839–2846. [Google Scholar] [CrossRef]
- Taylor, K.E.; Stouffer, R.J.; Meehl, G.A. An overview of CMIP5 and the experiment design. Bull. Am. Meteorol. Soc. 2012, 93, 485–498. [Google Scholar] [CrossRef] [Green Version]
- Meinshausen, M.; Smith, S.J.; Calvin, K.; Daniel, J.S.; Kainuma, M.L.T.; Lamarque, J.F.; Matsumoto, K.; Montzka, S.A.; Raper, S.C.B.; Riahi, K.; et al. The RCP greenhouse gas concentrations and their extensions from 1765 to 2300. Clim. Chang. 2011, 109, 213. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.H. Life-Histories of the Frogs of Okefinokee Swamp, Georgia: North American Salientia (Anura); Cornell University Press: Ithaca, NY, USA, 1932; Volume 2. [Google Scholar]
- Travis, J. Anuran Size at Metamorphosis: Experimental Test of a Model Based on Intraspecific Competition. Ecology 1984, 65, 1155–1160. [Google Scholar] [CrossRef]
- USGS. Water Data for the Nation; U.S. Geological Survey: Reston, VA, USA, 2022.
- Melillo, J.M.; Richmond, T.T.; Yohe, G. Climate change impacts in the United States. In Natioanl Climate Assessments; Federal National Climate Assessment and Development Advisory Committee: Washington, DC, USA, 2014. [Google Scholar]
- Nagel, L.D.; McNulty, S.A.; Schlesinger, M.D.; Gibbs, J.P. Breeding Effort and Hydroperiod Indicate Habitat Quality of Small, Isolated Wetlands for Amphibians under Climate Extremes. Wetlands 2021, 41, 1–11. [Google Scholar] [CrossRef]
- Greenberg, C.; Zarnoch, S.; Austin, J. Long term amphibian monitoring at wetlands lacks power to detect population trends. Biol. Conserv. 2018, 228, 120–131. [Google Scholar] [CrossRef]
- Rowe, C.L.; Dunson, W.A. Relationships among abiotic parameters and breeding effort by three amphibians in temporary wetlands of central Pennsylvania. Wetlands 1993, 13, 237–246. [Google Scholar] [CrossRef]
- Wells, K.D. The social behaviour of anuran amphibians. Anim. Behav. 1977, 25, 666–693. [Google Scholar] [CrossRef]
- Boswell, J.S.; Olyphant, G.A. Modeling the hydrologic response of groundwater dominated wetlands to transient boundary conditions: Implications for wetland restoration. J. Hydrol. 2007, 332, 467–476. [Google Scholar] [CrossRef]
- Griebler, C.; Avramov, M. Groundwater ecosystem services: A review. Freshw. Sci. 2015, 34, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Hamer, A.J.; Parris, K.M. Predation Modifies Larval Amphibian Communities in Urban Wetlands. Wetlands 2013, 33, 641–652. [Google Scholar] [CrossRef]
- Jara, F.G. The impact of phenology on the interaction between a predaceous aquatic insect and larval amphibians in seasonal ponds. Hydrobiologia 2019, 835, 49–61. [Google Scholar] [CrossRef]
- Brand, A.B.; Snodgrass, J.W. Value of Artificial Habitats for Amphibian Reproduction in Altered Landscapes. Conserv. Biol. 2010, 24, 295–301. [Google Scholar] [CrossRef]
- Le Viol, I.; Chiron, F.; Julliard, R.; Kerbiriou, C. More amphibians than expected in highway stormwater ponds. Ecol. Eng. 2012, 47, 146–154. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metamorphs | Juveniles | Adults | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | BMP | Breeding Period | Metam. Period * | Larval Duration (Days) | Size (mm) | n | Size (mm) | n | Size (mm) | n | n |
| H. femoralis | 1 May–26 October | 1 May–31 August | 22 June–26 October | 50–75 | <13 | 131 | <25 | 958 | ≥25 | 2271 | 3229 |
| H. gratiosa | 15 April–9 October | 15 April–28 August | 28 May–9 October | 41–160 | <25 | 44 | <44 | 285 | ≥44 | 60 | 345 |
| H. squirella | 1 April–16 October | 1 April–31 August | 10 May–16 October | 40–50 | <15 | 100 | <23 | 190 | ≥23 | 466 | 656 |
| Total | 275 | 1433 | 2797 | 4230 | |||||||
| Field-Derived Variable * | Correlation between Past Annual Values (1994–2017) and Predictions from Bioclim Variables in LASSO Model ** | ||
|---|---|---|---|
| H. femoralis | H. gratiosa | H. squirella | |
| Average weekly mean temperature | — | — | — |
| Total precipitation | 0.95 | 0.96 | 0.95 |
| Median pond depth | 0.45 | — | — |
| Minimum pond depth | — | — | — |
| Maximum pond depth | 0.62 | 0.68 | 0.64 |
| Minimum pond depth in month prior to breeding season | — | 0.56 | 0.57 |
| Maximum consecutive days with water in pond | 0.72 | 0.69 | 0.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Button, S.T.; Greenberg, C.H.; Austin, J.D. Recruitment Patterns and Potential Climate Change Impacts on Three Florida Hylids with Different Life Histories. Diversity 2022, 14, 129. https://doi.org/10.3390/d14020129
Button ST, Greenberg CH, Austin JD. Recruitment Patterns and Potential Climate Change Impacts on Three Florida Hylids with Different Life Histories. Diversity. 2022; 14(2):129. https://doi.org/10.3390/d14020129
Chicago/Turabian StyleButton, Sky T., Cathryn H. Greenberg, and James D. Austin. 2022. "Recruitment Patterns and Potential Climate Change Impacts on Three Florida Hylids with Different Life Histories" Diversity 14, no. 2: 129. https://doi.org/10.3390/d14020129
APA StyleButton, S. T., Greenberg, C. H., & Austin, J. D. (2022). Recruitment Patterns and Potential Climate Change Impacts on Three Florida Hylids with Different Life Histories. Diversity, 14(2), 129. https://doi.org/10.3390/d14020129